
Abstract
Changes in ovarian hormones predict changes in emotional eating across the menstrual cycle. However, in prior studies, researchers have not examined whether the nature of associations varies across dysregulated-eating severity. In the current study, we determined whether the strength or nature of hormone/dysregulated-eating associations differ on the basis of the presence of clinically diagnosed binge episodes (BEs). Participants included 28 women with BEs and 417 women without BEs who provided salivary hormone samples, ratings of emotional eating, and BE frequency for 45 days. Results revealed stronger associations between dysregulated eating and ovarian hormones in women with BEs relative to women without BEs. The nature of associations also differed, as progesterone moderated the effects of lower estradiol levels on dysregulated eating in women with BEs only. Although hormone/dysregulated-eating associations are present across the spectrum of pathology, the nature of associations may vary in ways that have implications for etiological models and treatment.
Past research has suggested that changes in ovarian hormones significantly predict within-person changes in binge-eating phenotypes across the menstrual cycle in women (Edler, Lipson, & Keel, 2007; Klump, Culbert, Edler, & Keel, 2008). Researchers in these studies initially examined associations indirectly by investigating changes in food intake and binge eating across menstrual-cycle phases. Results across samples, including samples from the community and samples composed of participants with bulimia nervosa (BN), were remarkably consistent in suggesting postovulatory peaks in food intake, binge eating, and phenotypes strongly related to binge eating (e.g., emotional eating, or the tendency to overeat in response to negative emotions; Edler et al., 2007; Gladis & Walsh, 1987; Gong, Garrel, & Calloway, 1989; Klump et al., 2008; Lester, Keel, & Lipson, 2003; Price, Torem, & Dimarzio, 1987).
The consistency of cycle-phase findings prompted researchers to examine whether natural changes in ovarian hormones across the menstrual cycle predict these within-person changes in food intake and binge-eating phenotypes. Although sample sizes were small (maximum N = 9), initial pilot studies indicated that changes in ovarian hormones accounted for the cycle-phase differences observed previously (Edler et al., 2007; Klump et al., 2008). Lower levels of estradiol and higher levels of progesterone were associated with increased binge frequency and emotional-eating scores in women with BN as well as in women drawn from the community (Edler et al., 2007; Klump et al., 2008). Importantly, these hormonal influences were found to be independent of within-person changes in negative affect and body mass index (BMI; Edler et al., 2007; Klump et al., 2008; Klump et al., 2013a), suggesting that the effects of hormones on binge eating risk were direct rather than indirectly related to these potentially important third variables.
Until recently, larger-scale studies were not available to replicate these direct effects. However, our group recently published the first large-scale study of within-person changes in hormones and emotional eating across the menstrual cycle in a community-based sample (Klump, Keel, et al., 2013). Our aim in that study was to replicate and extend prior work by examining both the main and the interactive effects of hormones on binge-eating risk. The focus on interactive effects emanated from results from experimental studies in animals, in which high levels of estrogen caused decreases in food intake when progesterone levels were low, whereas when estrogen and progesterone levels both were high, food intake increased as a result of the antagonizing effects of progesterone on estrogen (Blaustein & Wade, 1976; Gray & Wade, 1981; Kemnitz, Gibber, Lindsey, & Eisele, 1989; Varma et al., 1999). Using a sample of more than 190 women drawn from the community, we replicated the findings from animal studies by showing that interactions between estrogen and progesterone were the strongest hormonal predictors of within-person changes in emotional eating (Klump, Keel, et al., 2013). Emotional-eating scores were highest when both estradiol and progesterone levels were high (Klump, Keel, et al., 2013). These interactive effects appeared to account for postovulatory- and midluteal-phase increases in emotional-eating scores, given that the midluteal phase is characterized by the highest levels of progesterone and the second highest levels of estradiol across the entire menstrual cycle. Overall, these findings were significant in suggesting that estrogen and progesterone act in concert to increase emotional-eating levels across the menstrual cycle, most likely through the antagonizing effects of progesterone on estrogen (Klump, Keel, et al., 2013).
Nonetheless, our conclusions were limited by the exclusive focus on unselected, community samples of women. Effect sizes were somewhat small in our community sample, given that many women did not engage in significant amounts of emotional eating (Klump, Keel, et al., 2013). Subgroup analyses of more severely affected participants (e.g., the top 33% of emotional-eating scorers) revealed similar patterns of hormone/emotional-eating associations, but sample sizes were still modest, and conclusions were limited by the focus on emotional eating rather than on more traditional indices of binge eating (e.g., binge-eating frequency). Thus, although our previous study contributed important insights into the nature of hormone/dysregulated-eating associations in a community sample of women, the generalizability of findings to individuals suffering from more severe types of binge eating that are common in clinical eating disorders, such as BN and binge-eating disorder (BED), was unclear.
To date, only one small study (N = 9 participants) has investigated binge frequency in a clinically diagnosed sample (i.e., women with BN), and this study did not focus on interactions between estrogen and progesterone (Edler et al., 2007). Thus, it remains unknown whether the main effects of estrogen, the main effects of progesterone, or the interactive effects of the hormones are most predictive of changes in binge eating in women with clinical levels of pathology. Information regarding similarities and differences in the hormonal milieus that are “risky” for dysregulated eating across the spectrum of severity is needed to inform dimensional models of etiology (e.g., Research Domain Criteria; Cuthbert, 2005) and the development of novel treatments that consider biological triggers for binge eating (Nillni, Toufexis, & Rohan, 2011; van Elburg & Treasure, 2013).
Therefore, our aim in the current study was to examine hormone/dysregulated-eating associations across the menstrual cycle in a clinically diagnosed sample of women. Following previous recommendations (Cuthbert, 2005), we took a dimensional approach by studying women from our community-based sample who endorsed binge episodes (BEs) during study interviews. Although some of these women met criteria for a Diagnostic and Statistical Manual of Mental Disorders (5th ed.; DSM–5; American Psychiatric Association, 2013) eating disorder (i.e., BN or BED), others did not meet full diagnostic criteria but nonetheless endorsed engaging in BEs. We compared the magnitude and nature of hormone/emotional-eating associations in our BE sample participants (i.e., those women with and without eating disorders) with associations observed in our community sample participants who did not endorse BEs but who still varied in levels of emotional eating, a prospective risk factor for the development of BEs (Stice, Presnell, & Spangler, 2002). We also examined binge frequencies in the women with BEs to investigate whether emotional eating and BE frequency exhibit similar patterns of hormone/behavior associations across the menstrual cycle among women with more severe pathology.
Method
Participants
Participants were 445 female twins (aged 16–25 years) who participated in the Twin Study of Hormones and Behavior Across the Menstrual Cycle project (Klump, Keel, et al., 2013) within the Michigan State University Twin Registry (for a description of recruitment methods, see Burt & Klump, 2013; Klump & Burt, 2006). It is notable that a subset of participants in our full sample (n = 196, 44%) were included in our previous report (Klump, Keel, et al., 2013) that showed significant interactive effects of estrogen and progesterone on emotional-eating scores (reviewed in the previous section).
The current report focused on 28 women with a lifetime history of BEs (6.3%) and 417 women without BEs (93.7%). The diagnostic criteria used to determine the presence of BEs is outlined later in the Measures section. Of the 445 participants, 15 (3.4%) of the women in the BE sample endorsed a lifetime history of BN (n = 8, 1.8%) or BED (n = 7, 1.6%) during our diagnostic interviews. Given the small number of participants reporting BN and BED, our analyses focused on the full BE sample (regardless of BN or BED status) to maximize power to detect significant effects. This data aggregation is supported by analyses that showed no subgroup differences (i.e., differences between women with BEs only vs. women with BEs within the context of BN or BED) in ovarian-hormone levels or negative-affect scores (all ps > .10, average d = 0.26). Although women with BN and BED did report higher emotional-eating scores and a higher number of BEs than did women with BEs only (ps < .05, d = 0.47–0.77), these group differences would be expected, given the more severe pathology in the BN and BED groups.
All participants were required to meet the following inclusion criteria: (a) menstruation every 22 to 32 days for the past 6 months, (b) no hormonal contraceptive use within the past 3 months, (c) no psychotropic or steroid medications within the past 4 weeks, (d) no pregnancy or lactation within the past 6 months, and (e) no history of genetic or medical conditions known to influence hormone functioning or appetite/weight. Despite these inclusion criteria, participants did not differ meaningfully on continuous measures of binge eating or on other disordered-eating symptoms (e.g., weight preoccupation, body dissatisfaction) from participants from previous Michigan State University Twin Registry studies not requiring these inclusion criteria (average d = 0.12). Women without BEs were demographically representative of the recruitment region (see Table 1; Klump, Keel, et al., 2013; Racine et al., 2013b). Although women with BEs were primarily Caucasian, there was a similar percentage of individuals of Hispanic ethnicity in the BE sample relative to the no-BE sample. The predominance of Caucasian descent and a more equitable distribution of Hispanic ethnicity in the women with BEs would be expected, given that previous research has suggested increased rates of binge eating in Caucasians and individuals of Hispanic ethnicity (e.g., Alegria et al., 2007).
Descriptive Statistics
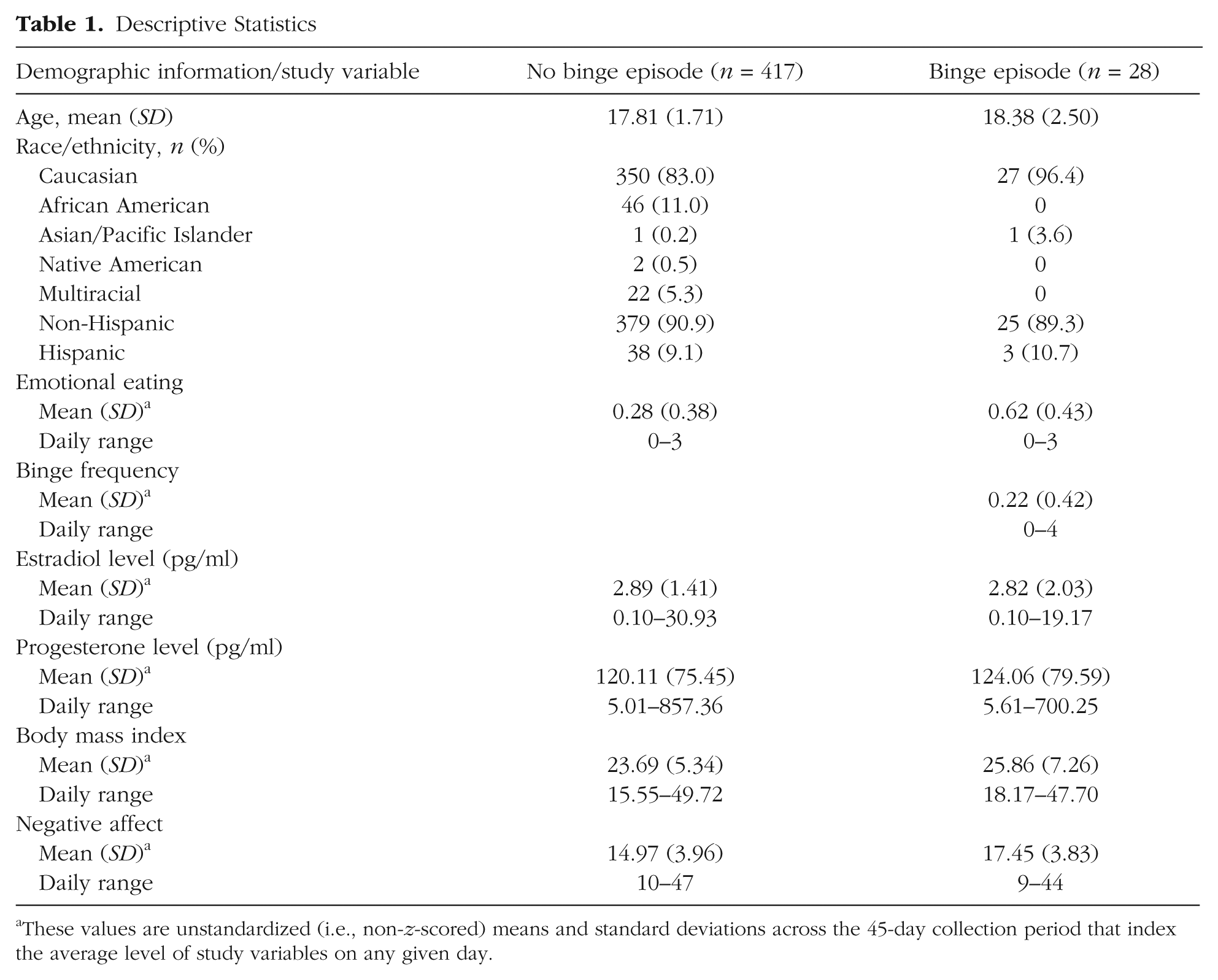
These values are unstandardized (i.e., non-z-scored) means and standard deviations across the 45-day collection period that index the average level of study variables on any given day.
Procedure
All study measures and procedures were approved by the Institutional Review Board of Michigan State University. Participants provided daily behavioral and hormone data for 45 consecutive days. Salivary samples were used to assay ovarian hormones and were collected within the first 30 min of waking using previously established methods (Edler et al., 2007; Klump et al., 2008). Questionnaires were completed each evening (after 5:00 p.m.) using an online data system or preprinted Scantrons. The timing of data collection ensured that a given day’s hormone collection preceded that day’s behavioral ratings (Edler et al., 2007; Klump et al., 2008).
In addition to daily data collection, all participants completed three in-person visits occurring at the start of the study, halfway through the study (approximately day 23), and at the end of data collection (approximately day 45). During these in-person assessments, eligibility was reassessed, height and weight were measured, and completed materials were collected from participants. The diagnostic interview for BEs was administered during the last study visit to ensure that we captured all symptoms that were present during the 45-day study period. Between visits, staff contacted participants once per week to answer questions and confirm continued protocol adherence. These procedures were effective at minimizing participant dropout (< 3%) and missing data (< 6%) as well as at identifying individuals who were no longer eligible to participate as a result of missed periods, medication use, or pregnancy (< 3%).
Measures
BEs
A modified version of the Eating Disorder Module from the Structured Clinical Interview for DSM–IV Axis I Disorders (First, Spitzer, Gibbon, & Williams, 1996) was used to assess the lifetime presence of BEs. Modifications were made to assess both DSM–IV (American Psychiatric Association, 1994) and DSM–5 criteria and to provide additional symptom probes for use with a community sample (e.g., multiple questions were added to assess loss of control over BEs).
All participants with BEs were required to endorse both the consumption of a large amount of food during a short period and the experience of loss of control over eating. Threshold BE participants (n = 15, 54%) met full criteria for both symptoms; that is, they consumed 1,000 calories or more during a BE and reported both behavioral and psychological indicators of loss of control. Subthreshold BE participants (n = 13, 46%) were defined as those who reported both BEs and a loss of control, but the level of severity was diminished in terms of either the amount of food eaten (i.e., 600–999 calories) or the degree of loss of control reported (i.e., some, but not all, behavioral indicators of loss of control were accompanied by endorsement of significant sadness, guilt, or disgust about the potential effects of the BE on body weight or shape). Because of the younger age of participants, and the community-based nature of the sample, we did not enforce DSM–5 frequency requirements for the presence/absence of BEs. However, the range of BE frequencies (i.e., 6 per year to 11 per week) indicated that the full spectrum of severity was present in our participants. Individuals who endorsed current BEs (i.e., BEs in the past month; n = 22, 79%) and individuals who endorsed past BEs (i.e., no BEs in the past month; n = 6, 21%) were included in the study to maximize sample size. The inclusion of participants who reported past BEs is supported by prior research that has suggested the existence of continued difficulties with dysregulated eating (e.g., emotional eating) in individuals with histories of BE, BN, and BED (Fairburn, Cooper, Doll, Norman, & O’Connor, 2000; Keel, Mitchell, Miller, Davis, & Crow, 1999). However, to ensure that the inclusion of individuals with histories of BE did not unduly influence our results, we conducted analyses with and without these 6 participants. The pattern of results was identical across analyses (data not shown) and, thus, only findings with the full BE sample (i.e., individuals who endorsed current BEs and individuals who endorsed past BEs) are reported herein.
The Structured Clinical Interview for DSM–IV Axis I Disorders was administered by trained interviewers with a master’s level education in clinical psychology or a related mental-health field. Training involved viewing instruction videos, studying/reviewing the symptom coding manual, listening to audiotapes of previous interviews, conducting practice interviews, and receiving feedback on interviewing skills via review of audiotapes and live observation. At the end of the training period (i.e., 2–3 months), all interviewers were required to pass a “checkout” interview with a senior interviewer on the project.
All interviews were audiotaped, and each co-twin was assessed by a different interviewer to ensure independence of symptom coding within twin pairs. Codes for all symptoms were reviewed during a weekly diagnostic case conference attended by the interview team and the principal investigator (K. L. Klump). Audiotapes of each interview were reviewed (as necessary), and participants were recontacted if there was insufficient information to code symptoms or make diagnoses. As with the interviews themselves, symptom reviews were conducted blind to co-twin status, given that co-twin interviews were never reviewed during the same case-conference session. Interrater reliability was assessed by comparing diagnoses made by an independent interviewer (i.e., an interviewer who was blind to study diagnosis but who listened to the audiotaped interview) with the final diagnosis. The kappa coefficient (.82) for the presence of BEs (as defined earlier) indicated substantial interrater agreement.
Emotional eating
Emotional eating was assessed daily using the Emotional Eating scale of the Dutch Eating Behavior Questionnaire (DEBQ; Van Strien, Frijters, Bergers, & Defares, 1986). The Emotional Eating scale assesses eating in response to negative emotions (example item: “Did you have a desire to eat when you were depressed?”; responses were made using scales from 1 (not at all) to 5 (very often). Internal consistencies for the DEBQ Emotional Eating scale are excellent in previous research (α = .93; Klump et al., 2008; Racine et al., 2012; Van Strien et al., 1986) and in the current study (45-day average α = .90). It is important to note that eating in response to negative emotions is thought to be a core feature of binge eating, and the DEBQ Emotional Eating scale has demonstrated validity in differentiating among individuals with BN or binge eating, overweight individuals, and college students. Furthermore, the DEBQ Emotional Eating scale is correlated with established measures of binge eating (rs = .55–.69; Racine, Culbert, Larson, & Klump, 2009; Van Strien, 2000) as well as with palatable food intake (i.e., ice cream) in a laboratory setting (Van Strien, 2000). Similar to previous research (Klump et al., 2008), the instructions for the scale were modified with permission to ask about emotional eating over the current day.
Binge frequency
Participants reported the daily frequency of BEs (0–9 or more episodes). To help ensure that participants reported accurately on binge frequency, we provided participants with a detailed definition of BEs during the first study session. Specifically, participants were told that binge eating was eating an unusually large amount of food (i.e., something that most people would think is larger than a normal meal) with a sense of loss of control over eating that often is experienced as feeling driven or compelled to eat, not being able to stop eating once started, or not being able to keep from eating large amounts of certain kinds of food in the first place. Participants were also quizzed on their understanding of BEs, in that they were provided with four case examples and asked to report on whether BEs or loss of control were present for each case according to the definitions given. Past research has suggested that providing BE definitions significantly increases the accuracy of self-reported BEs (Celio, Wilfley, Crow, Mitchell, & Walsh, 2004).
Ovarian hormones
Estradiol and progesterone were assayed from daily saliva samples. Saliva samples are preferred over other methods (e.g., blood spots) because they represent a less invasive collection method, particularly if repeated samples are needed. Previous research has shown that saliva samples are associated with higher compliance and more robust hormone-behavior associations than is blood-spot sampling (Edler et al., 2007).
Saliva samples were processed by Salimetrics, LLC (State College, PA) using enzyme immunoassay kits designed specifically for analyzing saliva. These assays show excellent intra- and interassay coefficients of variation (estradiol = 7.1% and 7.5%; progesterone = 6.2% and 7.6%), as well as assay sensitivity (measured by interpolating the mean optical density minus 2 SDs of 10–20 replicates at the 0-pg/ml level; estradiol = 0.10 pg/ml; progesterone = 5 pg/ml) and method accuracy (determined by spike recovery and linearity; estradiol = 104.2% and 99.4%; progesterone = 99.6% and 91.8%). To conserve resources, we assayed samples only every other day during menstrual bleeding and the early follicular phase when hormones are expected to be low and stable. This process ensured that we captured periods of maximum hormonal change across the menstrual cycle (e.g., the mid-to-late follicular through premenstrual phase) while, in turn, maximizing the number of participant samples assayed.
Negative affect
The Negative Affect scale from the Positive and Negative Affect Schedule (Watson, Clark, & Tellegen, 1988) was used to assess daily negative affect. This scale consists of 10 items that assess the full range of daily negative emotions (e.g., distress, nervousness, irritability, fear). Participants rated the degree to which each emotion was experienced; responses were made using scales from 1 (very slightly/not at all) to 5 (extremely). The Negative Affect scale has exhibited excellent internal consistency as well as good convergent and discriminant validity (Watson et al., 1988). Internal consistency in the current study was excellent (45-day average α = .85).
BMI
Participants’ height and weight were measured during the three in-person study visits using a wall-mounted ruler and a digital scale, respectively. BMI was calculated using the following formula: BMI = weight (in kilograms)/height (in meters)2.
Statistical analyses
Data preparation
Data preparation followed methods used in previous studies on the relationship between ovarian hormones and binge eating/emotional eating across the menstrual cycle (Edler et al., 2007; Klump et al., 2008; Klump, Keel, et al., 2013). For our repeated measures (i.e., emotional eating, binge frequency, hormones, negative affect, and BMI), 5-day rolling averages were calculated and standardized within-person. Rolling averages have been used in previous research and are preferred because of their ability to minimize random variation that is present in behavioral data as a result of environmental circumstances (Gladis & Walsh, 1987). To accommodate the fact that BMI was assessed at only three time points across the study, we calculated rolling averages using Visit 1 BMI for days in between the first and second in-person assessments, Visit 2 BMI for days in between the second and third in-person assessment, and Visit 3 BMI for the last day of the study. Rolling-average variables were then converted to within-person standardized scores based on each individual participant’s overall standard deviation across the study. This standardization meant that statistical-modeling results (see the following section) represent standardized estimates of effects that index the degree to which changes in a woman’s ovarian hormones, relative to her equilibrium, predict changes away from the woman’s equilibrium in emotional eating and binge eating.
Statistical models
Using methods identical to those outlined in Klump, Keel, et al. (2013), we employed mixed linear models (MLMs) to examine changes in emotional eating and binge eating across the menstrual-cycle phase (i.e., pre- vs. postovulation) as well as changes in ovarian hormones as predictors of changes in binge eating. MLMs are ideal for these analyses, given that we could examine predictive associations while controlling for covariates (i.e., negative affect and BMI) and the nonindependence of the repeated measures and twin data. We controlled for the nonindependence of the twin data by estimating random intercepts and allowing them to correlate. Because most co-twins were assessed across the same 45-day period, we also estimated a time-specific dyadic correlation that allowed the twins’ residual scores for emotional eating to correlate from day to day. For each of these random effects, we specified a compound-symmetry covariance structure that estimates a single intercept variance or a single residual variance across twins and time. The models also allowed for random slopes for each of the predictors (i.e., hormones, negative affect, BMI). However, because there was no evidence that these slopes correlated across twins, we fixed the cross-twin correlation to 0 for all predictors.
In our first set of MLMs, we examined changes in emotional-eating scores and, for women with BEs, binge frequency across pre- versus postovulation, given that previous research has revealed higher binge-eating rates in postovulatory phases of the menstrual cycle (Edler et al., 2007; Gladis & Walsh, 1987; Klump et al., 2008; Lester et al., 2003; Price et al., 1987). We investigated these changes by including a preovulation (i.e., from the follicular phase through the ovulatory phase) versus postovulation (i.e., from postovulation through the premenstrual phase) variable as the predictor in the MLMs. Pre- versus postovulation was coded on the basis of dates of menstrual bleeding and increases/decreases in hormones (see Edler et al., 2007, for coding methods). To examine whether there were BE/no-BE group differences in the magnitude or nature of the ovulation/dysregulated-eating association, we included a BE “sample” variable (i.e., women with BEs vs. women without BEs) as a moderator in the models. Significant interactions between the “sample” moderator and the ovulation phase indicated the presence of significant group differences in the nature or magnitude of changes in emotional eating across ovulation in women who did binge eat versus women who did not binge eat. It is important to note that we conducted these moderator analyses only for emotional-eating scores, and not for binge frequency, because women without BEs, by definition, would not be expected to report daily BEs. Consequently, for binge frequency, the models included only the women with BEs and did not include a “sample” moderator variable.
After examining ovulation-phase effects, we then tested whether changes in ovarian hormones (rather than menstrual-cycle phase) predicted within-person changes in dysregulated eating by examining estrogen, progesterone, and the Estrogen × Progesterone interaction as predictors of within-person changes in emotional-eating scores and binge frequencies. We initially fit a model that tested only the main effects of estradiol, progesterone, and the covariates (i.e., negative affect and BMI). We then fit a second model that included the Estradiol × Progesterone interaction in addition to all of the main effects and covariates. Significant hormone-interaction effects were graphed using the MLM beta weights (see Dawson & Richter, 2006) to determine the specific hormonal conditions (e.g., high vs. low levels of estradiol and progesterone) that were associated with the highest emotional-eating scores and binge frequencies. As before, we tested for BE/no-BE group differences in the nature or magnitude of hormone effects by including a BE “sample” moderator (i.e., women with BEs vs. women without BEs) in the models examining emotional-eating scores. Significant interactions between the “sample” variable and the hormones indicated the presence of significant group differences in hormonal influences on emotional eating between women who did and did not report BEs.
Results
Descriptive statistics
Table 1 includes descriptive statistics for daily, average levels of emotional eating, binge frequencies, ovarian hormones, BMI, and negative affect in the no-BE and BE groups. Age differences across the two samples were minimal, t(423) = 1.61, p = .11, d = 0.27, but, as expected, the women with BEs had significantly higher levels of emotional eating, t(443) = 4.49, p = .001, d = 0.84, higher BMIs, t(443) = 2.00, p = .047, d = 0.34, and higher levels of negative affect, t(443) = 3.21, p = .001, d = 0.67, relative to women without BEs. Importantly, levels of estradiol, t(443) = 0.27, p = .79, d = 0.04, and progesterone, t(443) = 0.31, p = .76, d = 0.05, were not significantly different between groups.
Changes across pre- versus postovulation
Within-person changes in emotional-eating scores, binge frequencies, and ovarian hormones across the menstrual-cycle phase are shown in Figures 1 and 2. Changes in estradiol and progesterone levels across the menstrual cycle were remarkably similar across groups and followed expectations for hormonal changes in all women (i.e., estradiol peaks before ovulation; progesterone peaks and a secondary estradiol peak in postovulation). Within-person changes in emotional eating and binge frequencies were similar across groups as well. There was no significant interaction between the BE “sample” variable (i.e., no BEs vs. BEs present) and the ovulation-phase variable (i.e., pre- vs. postovulation) for emotional-eating scores, β = 0.08, SE = 0.08, p = .30. In the combined participant group (i.e., women with and without BEs), the ovulation-phase variable was significant, β = −0.03, SE = 0.02, p = .04, and results showed significantly lower levels of emotional eating in preovulation (across the phase M = −0.06, SD = 0.95) compared with postovulation (across the phase M = 0.01, SD = 1.06). The same general pattern was observed for binge frequencies in the BE group only, that is, lower frequencies in preovulation (across the phase M = −0.02, SD = 0.92) compared with postovulation (across the phase M = 0.11, SD = 1.03), although smaller sample sizes in the BE group resulted in a nonsignificant beta estimate despite a higher beta value, β = −0.07, SE = 0.10, p = .49.
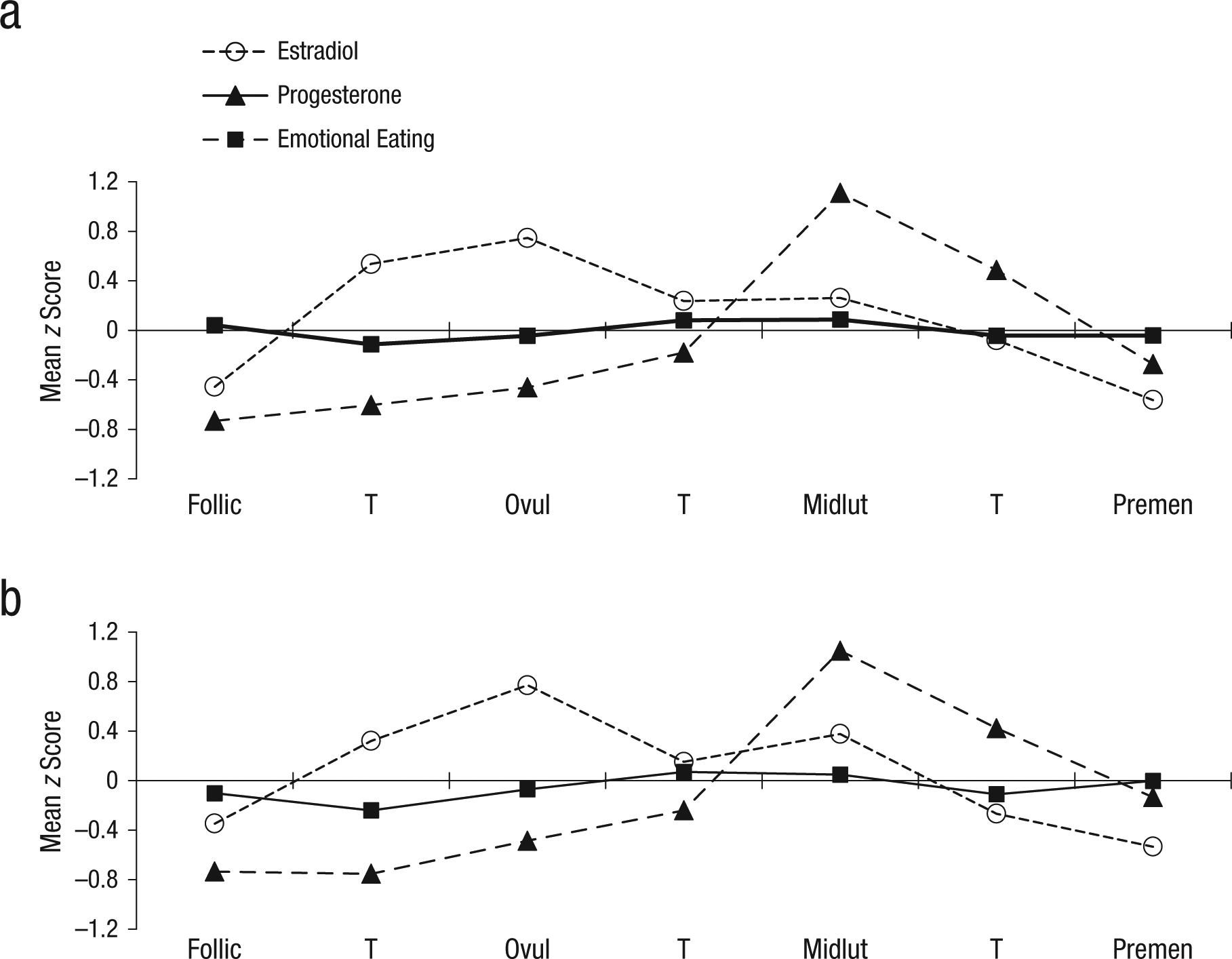
Changes in ovarian-hormone levels and emotional eating across the menstrual cycle in (a) women without binge episodes (n = 417) and (b) women with binge episodes (n = 28). Mean z score = the mean of the 5-day rolling averages calculated within-subjects and then averaged across participants; Follic = follicular phase; Ovul = ovulatory phase; Midlut = midluteal phase; Premen = premenstrual phase; T = transition days that are in between phases. The number of days included in each phase varied by participant based on their cycle length, but the days roughly corresponded to the following (1st day of menstrual bleeding = +1; previous day = −1): Follicular = +3 to +12; Ovulatory = −15 to −12; Midluteal = −9 to −5; Premenstrual = −3 to +1.
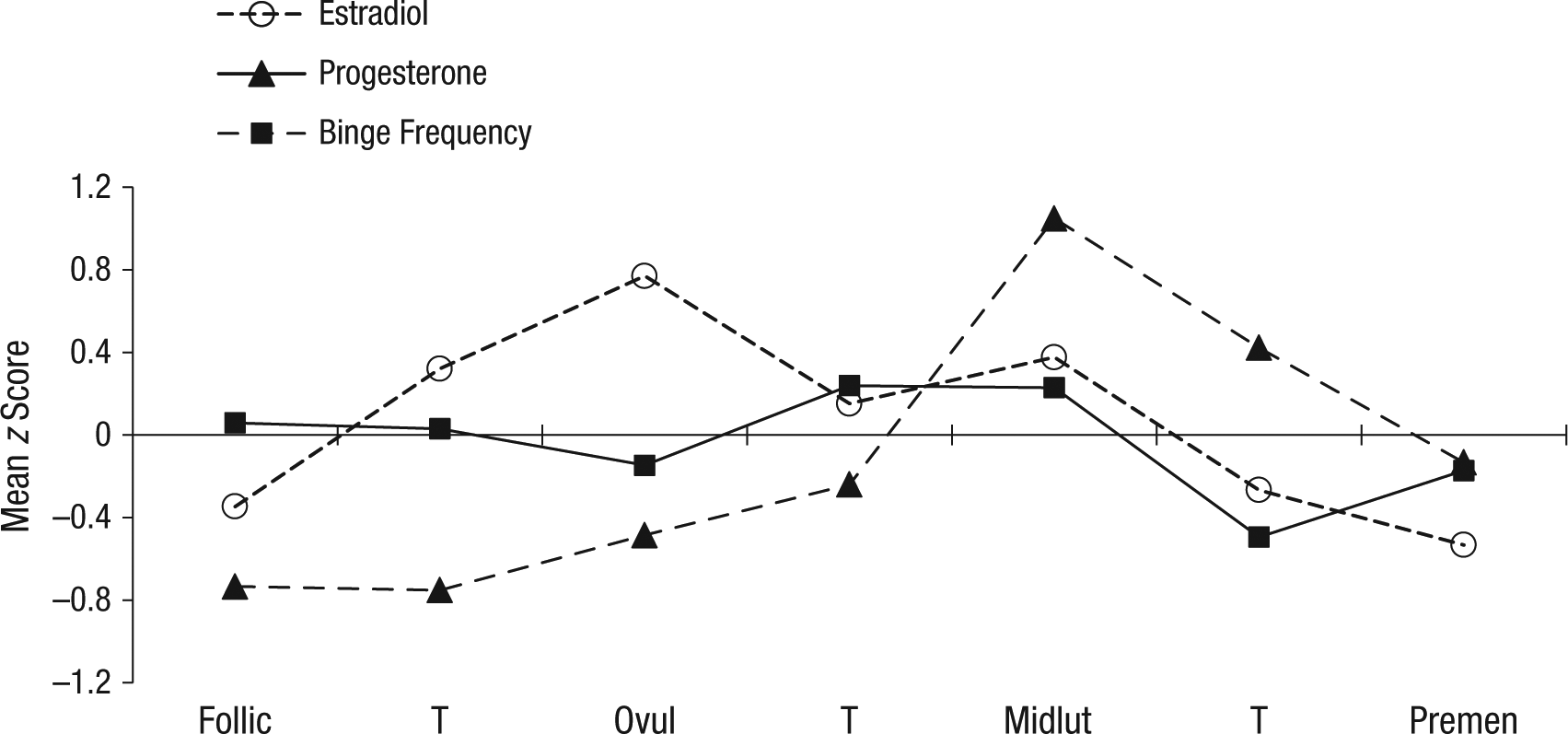
Changes in ovarian-hormone levels and binge frequencies across the menstrual cycle in women with binge episodes (n = 28). Mean z score = the mean of the 5-day rolling averages calculated within-subjects and then averaged across participants; Follic = follicular phase; Ovul = ovulatory phase; Midlut = midluteal phase; Premen = premenstrual phase; T = transition days that are in between phases. The number of days included in each phase varied by participant based on their cycle length, but the days roughly corresponded to the following (1st day of menstrual bleeding = +1; previous day = −1): Follicular = +3 to +12; Ovulatory = −15 to −12; Midluteal = −9 to −5; Premenstrual = −3 to +1.
Associations between changes in ovarian hormones and changes in dysregulated eating
Unlike analyses of cycle phase, there was a significant interaction between the “sample” variable and ovarian-hormone levels in the prediction of emotional-eating scores. Specifically, the three-way interaction (“Sample” × Estradiol × Progesterone) was statistically significant, β = 0.14, SE = 0.04, p = .004, which suggests that the influence of ovarian hormones on emotional-eating scores differed in women with BEs versus women without BEs. Given these significant interactions, in the remaining MLMs, we examined the no-BE and BE groups separately.
Results for the women without BEs generally corroborated our initial findings from this data set (Klump, Keel, et al., 2013) by showing an interaction (but no main effects) between estrogen and progesterone in the prediction of emotional-eating scores (see Table 2). The Estrogen × Progesterone interaction just missed statistical significance (p = .10), but it took the form of interactions observed in our previous analysis of the full sample (Klump, Keel, et al., 2013). Specifically, emotional-eating scores for women without BEs were elevated when both progesterone and estradiol levels were high (see Fig. 3a). Although the magnitude of the interaction was diminished compared with that reported previously (see Klump, Keel, et al., 2013), this would be expected, given that in the current sample, we excluded all participants with clinical levels of binge eating. Thus, the variability and severity of emotional eating (and, consequently, associations with ovarian hormones) would be expected to be reduced in the no-BE sample.
Predictive Associations Between Hormones and Dysregulated Eating
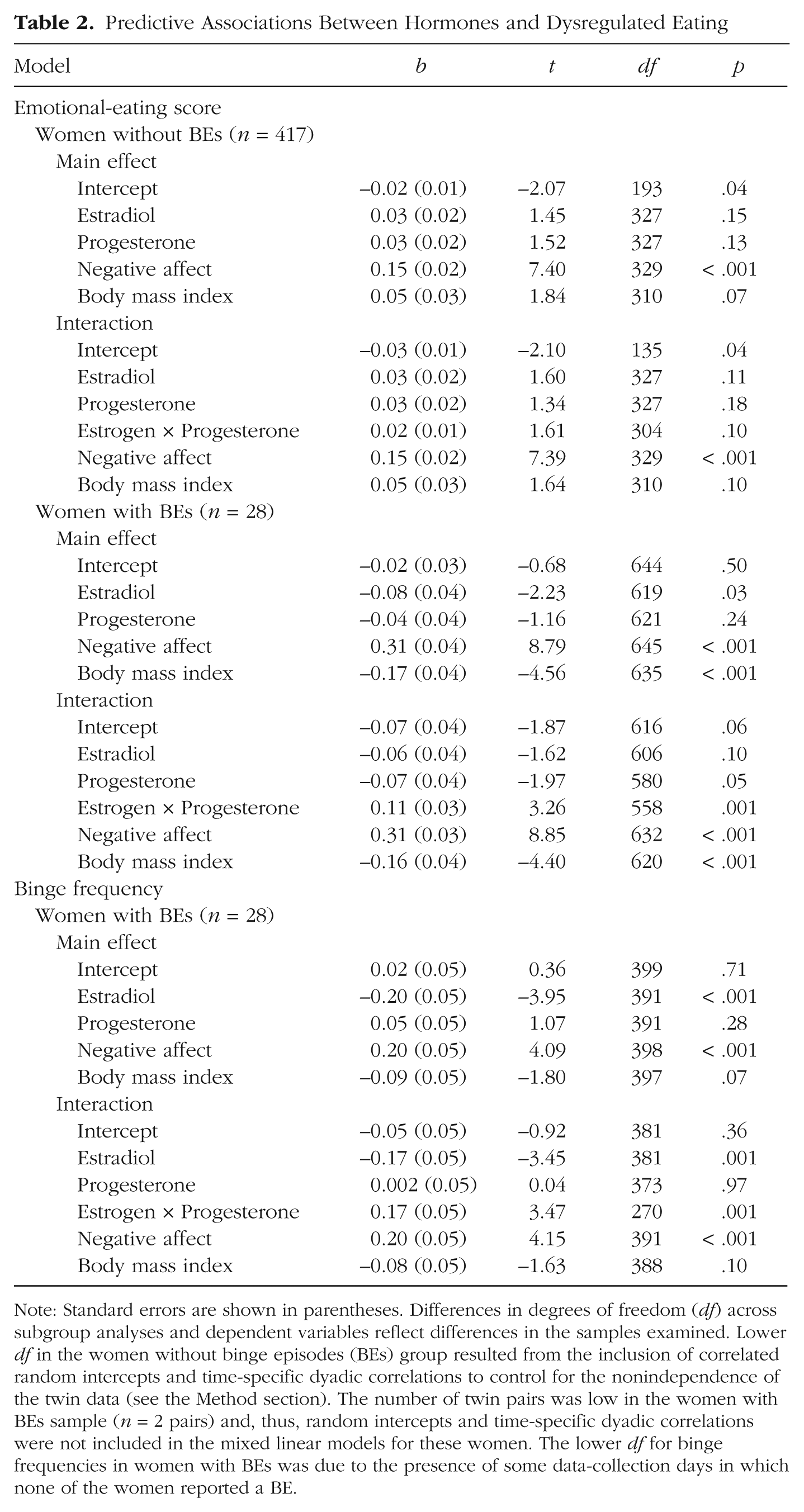
Note: Standard errors are shown in parentheses. Differences in degrees of freedom (df) across subgroup analyses and dependent variables reflect differences in the samples examined. Lower df in the women without binge episodes (BEs) group resulted from the inclusion of correlated random intercepts and time-specific dyadic correlations to control for the nonindependence of the twin data (see the Method section). The number of twin pairs was low in the women with BEs sample (n = 2 pairs) and, thus, random intercepts and time-specific dyadic correlations were not included in the mixed linear models for these women. The lower df for binge frequencies in women with BEs was due to the presence of some data-collection days in which none of the women reported a BE.
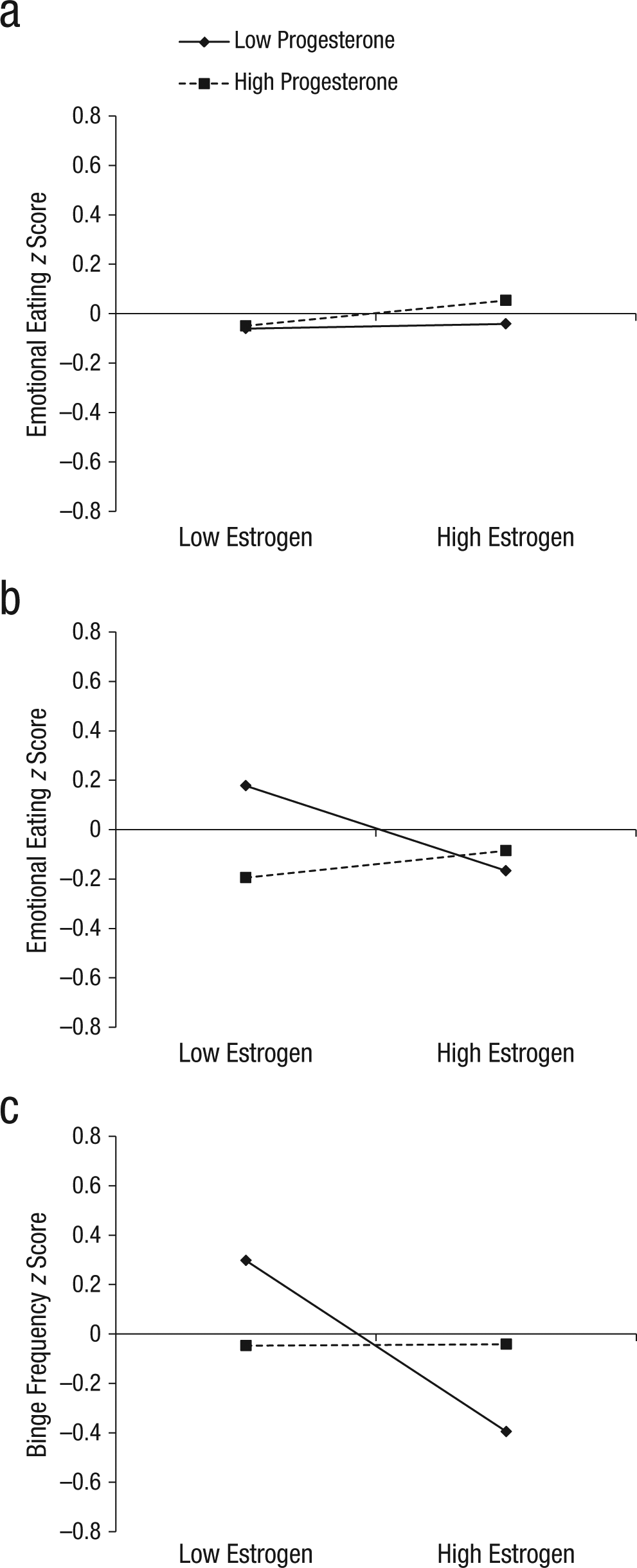
Interactions between estradiol and progesterone in the prediction of (a) emotional-eating scores in women without binge episodes (n = 417) and (b) emotional-eating scores and (c) binge frequencies in women with binge episodes (n = 28). z scores = 5-day rolling average calculated within-subjects and then averaged across participants.
Findings for the women with BEs (see Table 2) were somewhat different: The magnitude of the hormone effects appeared to be larger than were those in the women without BEs (see Figs. 1–3), and the nature of the effects appeared to be somewhat different. Specifically, in the main-effects models, estrogen showed a significant main effect in women with BEs, with lower levels of estradiol predicting higher emotional-eating scores and binge frequencies, even after we controlled for changes in negative affect and BMI. These estrogen effects were not observed in the women without BEs or in analyses of unselected twins from this sample (see earlier discussion and Klump, Keel, et al., 2013), but they are consistent with a previous study of women with BN in which inverse associations between estradiol levels and binge frequency were observed (Edler et al., 2007).
Nonetheless, follow-up models indicated that the interactive effects of estradiol and progesterone were also predictive of within-person changes in both types of dysregulated eating in women with BEs. As shown in Figures 3b and 3c, emotional-eating scores and binge frequencies were elevated when both progesterone and estradiol levels were low. This pattern was observed for emotional-eating scores and binge frequencies, which suggests that the findings are robust and present across a range of dysregulated-eating symptoms.
At first glance, these differential hormone effects across the no-BE/BE group might seem to contradict findings from analyses of ovulation, given that postovulation peaks were observed in both women with BEs and women without BEs (with no significant group differences in effects). However, closer inspection of Figures 1b and 2 reveals that although emotional-eating scores and binge frequencies were highest during postovulation in both groups, the postovulation peaks and valleys differed somewhat between women with BEs versus women without BEs. In women without BEs, the highest postovulatory peak was in the midluteal phase, whereas the highest peaks for the women with BEs were in the midluteal phase as well as the premenstrual phase and the transition phase from ovulation. These latter two phases tend to include lower levels of both hormones (particularly the premenstrual phase), which suggests that the hormonal milieus that are risky for women with BEs include both high levels (during the midluteal phase) and low levels (during the premenstrual phase) of hormones. This interpretation is supported by data that show that trends for higher levels of hormones to be associated with the second highest elevations in dysregulated eating in women with BEs (see Fig. 3). Interestingly, there was a tendency for high estradiol and low progesterone levels to be associated with the lowest levels of dysregulated eating (see Figs. 3b and 3c). This latter finding replicates results from the women without BEs (see Fig. 3a) and previous analyses (see Klump, Keel, et al., 2013) in which the lowest levels of emotional eating were observed at high estradiol and low progesterone levels (Klump, Keel, et al., 2013).
Discussion
Our results reveal important similarities as well as differences in hormone/dysregulated-eating associations across the spectrum of binge-eating severity. Emotional-eating scores and binge-eating frequencies peaked in the postovulatory phases of the menstrual cycle for women with BEs and women without BEs, and high levels of hormones appeared to partially account for these peaks. In addition, in the BE sample only, lower levels of hormones (in particular, estradiol) predicted within-person increases in emotional-eating scores and binge frequencies. Critically, in the BE group, patterns observed for fluctuations in emotional eating were identical to those for fluctuations in binge frequency, which suggests that the continuous measure of emotional eating taps clinically relevant dimensions of binge eating (Racine et al., 2013a; Wardle, 1987) that likely share risk factors with full-threshold BEs (Stice et al., 2002).
Nonetheless, variations in hormone/binge-eating associations between the no-BE and BE samples deserve note. Overall, findings suggested a stronger effect of lower levels of hormones in women with BEs than in women without BEs. Estrogen exerted a significant main effect in nearly all models, and interaction graphs indicated that low levels of estrogen and progesterone were most predictive of within-person changes in dysregulated eating in women with BEs. With regard to physiology, these findings are consistent with previous suggestions that progesterone has few direct effects on binge eating or food intake but, instead, acts indirectly by antagonizing estrogen. These antagonizing effects would be expected to be diminished if progesterone levels are low, particularly if estrogen levels also are low (Asarian & Geary, 2006). In these types of hormonal milieus, low levels of progesterone likely act as permissive factors in allowing for the behavioral effects of low estrogen levels (i.e., increased binge eating and food intake; see Asarian & Geary, 2006) to be manifested.
However, it remains unclear why progesterone would moderate both high and low levels of estradiol in women with BEs but not in women without BEs. Only one previous study has investigated women with BEs (Edler et al., 2007). In that study, researchers assessed 9 women with BN and found that low levels of estrogen, and high levels of progesterone, were associated with increased binge frequencies across the menstrual cycle. However, they did not examine Estrogen × Progesterone interactions and, thus, could not investigate how the two hormones act in concert to influence changes in binge eating. In our sample of women with BEs, we saw the same pattern of main effects for binge frequency (e.g., low estradiol and high progesterone levels were associated with increased binge frequency; see Table 2), even though the progesterone main effects were not significant. Thus, although our main effects are largely consistent with the prior study of women with BN, they do not help explain the somewhat different pattern of hormone interactions in women with BEs versus women without BEs.
One possible contributing factor is the aberrant eating patterns (e.g., dieting) that often characterize women with BEs. Dieting could affect changes in hormone levels or modify associations between hormone changes and changes in dysregulated eating in women with BEs versus women without BEs (even if daily hormone levels are equivalent; see Table 1). To directly examine this possibility, we conducted post hoc tests to investigate whether the effects of hormones became less significant when we accounted for dietary restraint in the women with BEs. We included rolling averages for dietary restraint scores (measured at three time points using the Eating Disorders Examination Questionnaire; Fairburn & Beglin, 1994) as covariates and found that the Estradiol × Progesterone interactions continued to be significant in all models (ps < .05).
Consequently, the bulk of the data thus far indicates that the dual risky hormonal milieus in women with BEs (higher and lower hormone levels) are robust and present despite changes in other variables across the cycle (e.g., dietary restraint). Both hormonal milieus would be expected to increase dysregulated eating via the antagonistic effects (at high hormone levels) or permissive effects (at low hormone levels) of progesterone on estrogen influences on behavior. However, future research is needed to determine why the different hormonal milieus are risky for women with BEs versus those without BEs and to identify factors (e.g., psychological, genetic, neurobiological) that might contribute to this differential pattern of risk.
One potentially interesting possibility in this regard is differential hormone regulation of genetic expression for binge eating in women with BEs versus women without BEs. As we have proposed previously (Klump, 2013; Klump, Keel, et al., 2013), it is likely that hormones influence dysregulated eating in all women via their regulatory effects on gene expression within the central nervous system. Ovarian hormones in general, and estrogen in particular, act as gene-transcription factors to activate and deactivate genes within several neurotransmitter systems (e.g., serotonin, dopamine, opioids) that are believed to be important for binge eating and eating disorders characterized by binge eating (e.g., BN; Becker, 1999; Craft, 2008; Hildebrandt, Alfano, Tricamo, & Pfaff, 2010; Ostlund, Keller, & Hurd, 2003). Effects of ovarian hormones on dysregulated eating in all women therefore might result from the hormones’ effects on the production of these important neurotransmitters, their receptors, or their signal-transduction mechanisms. In these scenarios, the key individual differences variable is the presence or absence of susceptibility alleles that are regulated by ovarian hormones. These individual differences would be most evident in hormonal milieus (e.g., during the midluteal phase when estradiol and progesterone levels are high) that trigger gene activation or deactivation and result in increased rates of dysregulated eating.
With our current data in hand, we can move beyond theories regarding general effects that might be present in all women and theorize about specific Gene × Hormone effects that might be present in women who go on to develop full-threshold BEs. As noted earlier, estrogen regulation of neurobiological systems that are important for binge eating appears to be more robust than progesterone regulation of these systems. The differential strength of estrogen versus progesterone maps nicely onto the pattern of phenotypic results presented herein. In all models of women with BEs, estrogen showed stronger phenotypic associations with dysregulated eating than did progesterone, and interaction graphs clearly depicted more direct effects of estrogen than of progesterone. These strong and direct effects of estrogen in women with BEs was in contrast to the lack of direct estrogen effects observed in women without BEs, where the interaction between estrogen and progesterone was most important for dysregulated eating.
In aggregate, these findings suggest that perhaps women with BEs have a stronger or a different genetic loading for dysregulated eating that translates into even stronger effects of estrogen on binge-eating risk. Women with BEs may have a larger number of estrogen-responsive risk genes or different genetic variants relative to women without BEs. Given that genetic influences are present across the spectrum of binge eating (Bulik, Sullivan, & Kendler, 1998; Klump, McGue, & Iacono, 2000; Racine, Burt, Iacono, McGue, & Klump, 2011; Racine et al., 2013a), and that we see hormone effects across the full range of dysregulated eating (see earlier discussion and Klump, Keel, et al., 2013), we are not proposing that genetic effects are present in women with BEs but not in women without BEs. It is clear that genetic influences are present in both groups. But what might be different is the relative number or type of estrogen-responsive risk genes in women with BEs that translates into greater risk for BEs and a wider range of estrogen milieus that are risky.
What is interesting about this potential mechanistic picture is that in women with BEs, low estradiol levels are risky, particularly in the presence of low progesterone levels. Presumably, it is the lack of regulation of gene expression in women with BEs that translates into greater risk for BEs. This direction of association may seem counterintuitive (lower gene regulation equals greater risk); however, estrogen can either activate or deactivate gene transcription. Thus, at lower estradiol levels, there may be lower or higher transcription depending on the particular role for estrogen within that neurobiological system.
It is clear that future studies are needed to test all of these mechanistic hypotheses, but such studies are possible using both nonhuman animal and human research. Nonhuman animal studies could allow researchers to examine the differential effects of estradiol dose on binge eating and gene expression in ovariectomized female animals known to be prone versus resistant to binge eating (see Boggiano et al., 2007; Klump, Racine, Hildebrandt, & Sisk, 2013; Klump, Suisman, Culbert, Kashy, Keel, & Sisk, 2011; Klump, Suisman, Culbert, Kashy, & Sisk, 2011). A stronger or differential effect of estradiol dose on binge eating and the number/type of genes expressed in binge-prone animals would suggest that estrogen regulation may contribute to differences in binge-eating proneness. In human studies, researchers could use our longitudinal, menstrual-cycle design to examine changes in heritability and gene expression across the menstrual cycle in large samples of female twins with and without binge eating. Differential changes in heritability and gene expression by estradiol levels would suggest that differences in gene regulation may contribute to differences in the effects of ovarian hormones on dysregulated eating in women with BEs versus women without BEs. It is important to note that both the nonhuman animal and the human studies could include strong “control” hormonal conditions (high hormone levels for increased risk; high estradiol and low progesterone levels for decreased risk) that do not differ in women with BEs and women without BEs to determine the specific effect of low estradiol levels on genetic risk in women with BEs.
The trick to completing all of these studies is to identify the most important risk genes for analysis. Given the state of molecular genetic studies of eating disorders (see Trace, Baker, Peñas-Lledó, & Bulik, 2013), there unfortunately are no replicated risk genes for binge eating or eating disorders characterized by binge eating (e.g., BN). Consequently, the most fruitful approach might be to select genes that already known to be regulated by estrogen that are within key neurobiological systems that likely contribute to the development and phenomenology of binge eating. In our view, genes involved in the reward circuitry contained in the Positive Valence Domain in the Research Domain Criteria (Insel et al., 2010) are likely to be helpful in this regard. Past studies have suggested that differential activation of reward areas in the brain (e.g., the nucleus accumbens) likely contribute to binge eating in nonhuman animals (Berridge, 2009; Gradl, Klump, & Sisk, 2014) as well as in humans (Gearhardt et al., 2011; Stice, Spoor, Ng, & Zald, 2009). Key neurotransmitters in these systems (e.g., dopamine and opioids) show strong estrogen regulation (Becker, 1999; Craft, 2008), which highlights the potential for genes in these systems to exhibit differential responsivity to estrogen in women with BEs and women without BEs. Given uncertainty as to which particular dopaminergic and opioid genes may be important, a biological pathway analysis in which all genes within a particular circuit are examined might be a good first step in this line of work (Bralten et al., 2013).
Alternatively, a sequential design, in which a series of nonhuman animal studies are used to identify genetic candidates that are then explored further in studies of women both with and without BEs, might be useful. We particularly like the last combination of nonhuman animal and human studies because this approach would identify replicable Gene × Hormone effects across species that could advance the science of translational research and highlight new areas of interface between basic animal science and neurobiological research in humans.
Before ending, two key study limitations should be noted. First, although our sample of women with BEs was larger than those samples examined previously, the sample was relatively small for a longitudinal study. Our use of a 45-day collection period somewhat ameliorated these concerns, given that the use of repeated measures tends to decrease measurement error (Kazdin, 2002). Nonetheless, replication of our findings in larger samples is needed.
Second, because our sample was community based, we were limited to examining participants with subthreshold in addition to threshold BEs who might have lacked the severity of binge eating present in treatment-seeking participants. Data shown in Table 1 suggest that the daily frequency of BEs was lower than that observed in clinical samples (Edler et al., 2007; Lester et al., 2003). Our inclusion of participants who endorsed past BEs (n = 6, 21%) also likely limited variability in our outcome measures. Although our pattern of results was unchanged when we excluded these participants (see the Measures section), additional research is needed to replicate our results and extend them to individuals more severely affected by BEs.
Footnotes
Declaration of Conflicting Interests
The authors declared that they had no conflicts of interest with respect to their authorship or the publication of this article.
Funding
This work was supported by National Institute of Mental Health (NIMH) Grants R01 MH082054, T32 MH018269, and T32 MH070343. The content of this article is solely the responsibility of the authors and does not necessarily represent the official views of the NIMH.