
Abstract
Cadherin 26 (CDH26) is a recently identified member of the cadherin superfamily. Although CDH26 gene expression has been reported in association with allergic inflammatory responses, the protein expression levels and the signaling pathways mediated through its interactions with other proteins remain poorly understood. This is primarily due to the lack of monoclonal antibodies (mAbs) that can recognize the intact, cell surface-expressed form of CDH26. In this study, we developed an antihuman CDH26 mAb, Ca26Mab-6 (IgM, kappa), using the Cell-Based Immunization and Screening (CBIS) method. Ca26Mab-6 demonstrated high sensitivity and specificity for CDH26 in flow cytometry and did not bind to Chinese hamster ovary (CHO)-K1 cells, which overexpress any of the other type I or type II cadherins. Ca26Mab-6 successfully detected endogenous CDH26 protein expression in HepG2, U-87 MG, MCF7, and 293FT cells. The apparent dissociation constant of Ca26Mab-6 was determined to be 9.8 ± 4.8 × 10−9 M for CDH26-overexpressed CHO-K1 (CHO/CDH26) cells and 3.6 ± 1.0 × 10−7 M for HepG2 cells. The detection of CDH26 expression in cancer cells may offer new insights into the potential relationship between inflammatory responses and malignant transformation. Therefore, Ca26Mab-6, developed using the CBIS method, is expected to facilitate functional studies of CDH26 and contribute to the development of CDH26-targeted antibody-based therapies.
Introduction
Cadherins are defined as cell surface glycoproteins responsible for calcium-dependent cell–cell adhesion.1–5 Large-scale comparative genome sequencing projects have identified over 20,000 cadherin protein sequences, which are now classified into a cadherin major branch, subdivided into 2 families and 8 subfamilies, and a cadherin-related major branch, subdivided into 4 families and 11 subfamilies. 6 The major branch includes 21 classical human cadherins, which are further classified into type I and type II cadherins based on their similar structures. 6 These cadherins share a common architecture comprising five extracellular cadherin repeat domains with conserved calcium-binding motifs, a single-pass transmembrane domain, and a highly conserved cytoplasmic domain.6,7 The extracellular repeat domains mediate homophilic cell–cell interactions, while the cytoplasmic domain interacts with β-catenin and p120-catenin to mediate intracellular signaling. 7 In addition to type I and type II cadherins, cadherin 26 (CDH26) is a relatively newly identified member of the cadherin family and is classified as a nonclassical cadherin. 6 CDH26 has five extracellular cadherin repeat domains, but its cytoplasmic domain is unique. Phylogenetic analysis of its amino acid sequence, based on the extracellular repeat domains of cadherins, reveals that CDH26 forms a distinct branch, diverging after the desmogleins and before the desmocollins and type I cadherins. 6
Cadherins play a crucial role in cell recognition and adhesion processes, and cadherin-mediated adhesion plays a major role in development and tissue morphogenesis.8,9 Their function has been reported as maintaining tissue architecture,8,10 stabilizing intercellular adhesion, 11 participating in Wnt/β-catenin signaling, 12 and being essential for the development of the nervous 13 and cardiovascular systems. 14 In addition, cadherins are associated with cancer invasion and metastasis15,16 and are involved in inflammatory and allergic diseases. 17 In this context, the inhibition of cadherin signaling has significant implications for various diseases and has been investigated as a therapeutic target.18–21
CDH26 has been reported to be expressed in human allergic gastrointestinal tissues, such as those from patients with eosinophilic gastritis and eosinophilic esophagitis. 17 Moreover, CDH26 mRNA expression is significantly upregulated (approximately 115-fold) in the esophageal tissues of patients with active eosinophilic esophagitis compared with controls. 17 Immunohistochemical staining using anti-CDH26 polyclonal antibodies has shown that CDH26 is localized almost exclusively to the surface and glandular epithelial cells in the gastric tissues of patients with eosinophilic gastritis. 17 In addition, CDH26 has been implicated in the inflammatory responses associated with allergic rhinitis and asthma.22,23 CDH26 is expressed in airway epithelial cells and has been implicated in actin cytoskeleton maintenance and bone edge polarity. 24 Although these findings have been derived primarily from knockdown experiments22,24 and RNA sequencing analyses, 23 the protein expression levels of CDH26 and signal transduction elicited by CDH26 interactions with proteins remain poorly understood. Therefore, the development of monoclonal antibodies (mAbs) that can recognize the intact form of CDH26 on the cell surface is urgently needed.
In this study, we developed anti-CDH26 mAbs by the Cell-Based Immunization and Screening method. 25 The obtained mAb, Ca26Mab-6 (IgM, kappa), showed a clear response of CDH26 expression in flow cytometry. This mAb is anticipated to open new avenues from basic studies of CDH26 function to the development of therapeutic applications.
Materials and Methods
Cell lines
Chinese hamster ovary (CHO)-K1, mouse myeloma P3X63Ag8U.1 (P3U1), human liver carcinoma HepG2, and human glioblastoma U-87 MG cells were obtained from the American Type Culture Collection (Manassas, VA, USA). The human breast cancer cell line MCF7 was obtained from the Cell Resource Center for Biomedical Research, Institute of Development, Aging and Cancer, Tohoku University (Miyagi, Japan). The human embryonic kidney cell line 293FT was purchased from Thermo Fisher Scientific Inc. (Waltham, MA, USA). All cell lines were maintained as previously described. 25
Plasmid construction and establishment of stable transfectants
Stable CHO-K1 transfectants that overexpressed human type I cadherins, including CHO/CDH1, CHO/PA16-CDH2 (CHO/CDH2), CHO/CDH3, CHO/PA16-CDH4 (CHO/CDH4), and CHO/PA16-CDH15 (CHO/CDH15) cells, were previously established. 25 CHO-K1 transfectants that overexpressed human type II cadherins and CDH26 were also established. Genes encoding human CDH6 (NM_004932.4), CDH7 (NM_004361.5), CDH17 (NM_004063.4), CDH19 (NM_021153), and CDH24 (NM_022478) were purchased from OriGene Technologies, Inc. (Rockville, MD, USA). Genes encoding CDH8 (NM_001796.5), CDH12 (NM_004061), CDH13 (NM_001257), CDH16 (NM_004062), CDH18 (NM_004934), and CDH26 (NM_177980) were obtained from the RIKEN BioResource Research Center (Ibaraki, Japan). Genes encoding CDH5 (NM_001795.5), CDH9 (NM_016279.4), CDH10 (NM_006727.5), CDH11 (NM_001797), CDH20 (NM_031891), and CDH22 (NM_021248) were synthesized by Eurofins Genomics KK (Tokyo, Japan).
The cDNAs of CDH5, CDH7, CDH8, CDH9, CDH10, CDH11, CDH12, CDH13, CDH15, CDH16, CDH18, CDH19, CDH20, CDH22, CDH24, and CDH26, with the signal sequence and propeptide removed, were subcloned into the pCAG-Ble vector (FUJIFILM Wako Pure Chemical Corporation, Osaka, Japan) with an N-terminal PA16 tag (GLEGGVAMPGAEDDVV). 26 In addition, the cDNA of CDH26 with an N-terminal MAP16 tag (PGTGDGMVPPGIEDKI) 27 was constructed. The cDNAs of CDH6 and CDH17 were supplied in pCMV6 vectors containing a C-terminal mycDDK tag (OriGene Technologies, Inc.). These plasmids were transfected into CHO-K1 or LN229 cells, and stable transfectants were sorted using an anti-CDH6 mAb (clone 427909, Thermo Fisher Scientific Inc.), an anti-CDH11 mAb (clone 16G5, BioLegend, San Diego, CA, USA), an anti-CDH17 mAb (clone 2618, Novus Biologicals, Centennial, CO, USA), an anti-PA16 tag mAb (clone NZ-1), 26 and an anti-MAP16 tag mAb (PMab-1) 27 using an SH800 cell sorter (Sony Corporation, Tokyo, Japan). The CDH-overexpressed stable transfectants were established, including CHO/PA16-CDH5 (CHO/CDH5), CHO/CDH6-mycDDK (CHO/CDH6), CHO/PA16-CDH7 (CHO/CDH7), CHO/PA16-CDH8 (CHO/CDH8), CHO/PA16-CDH9 (CHO/CDH9), CHO/PA16-CDH10 (CHO/CDH10), CHO/PA16-CDH11 (CHO/CDH11), CHO/PA16-CDH12 (CHO/CDH12), CHO/PA16-CDH13 (CHO/CDH13), CHO/PA16-CDH15 (CHO/CDH15), CHO/PA16-CDH16 (CHO/CDH16), CHO/CDH17-mycDDK (CHO/CDH17), CHO/PA16-CDH18 (CHO/CDH18), CHO/PA16-CDH19 (CHO/CDH19), CHO/PA16-CDH20 (CHO/CDH20), CHO/PA16-CDH22 (CHO/CDH22), CHO/PA16-CDH24 (CHO/CDH24), CHO/PA16-CDH26 (CHO/CDH26), and LN229/MAP16-CDH26 (LN229/CDH26) cells. To confirm the expression of CDHs in these transfectants, 1 μg/mL of commercially available mAbs or 0.1 μg/mL of NZ-33, 28 a derivative of NZ-1, was used.
Production of hybridomas
All animal experiments were conducted in accordance with the guidelines for animal care and use and were approved by the Animal Care and Use Committee of Tohoku University (Permit No. 2022MdA-001). Every effort was made to minimize animal suffering and distress throughout the study. Animals were housed under specific pathogen-free conditions. Two 6-week-old female BALB/cAJcl mice (obtained from CLEA Japan [Tokyo, Japan]) were intraperitoneally immunized with LN229/CDH26 (1 × 108 cells/mouse/week, five times in total). Alhydrogel adjuvant 2% (InvivoGen, San Diego, CA, USA) was used in the first immunization. Hybridomas were generated as described previously. 25
Flow cytometry
Cells were detached using 1 mM ethylenediaminetetraacetic acid (Nacalai Tesque, Inc., Kyoto, Japan) to prevent enzymatic degradation of surface proteins. The cells were washed with 0.1% bovine serum albumin in phosphate-buffered saline (blocking buffer) and incubated with mAbs at 4°C for 30 minutes. CHO/CDH26 was transfected with CDH26 siRNAs (s34067, s34068, and s34069, Thermo Fisher Scientific Inc.) by Lipofectamine RNAiMAX reagent (Thermo Fisher Scientific Inc.) for 24 hours and incubated with 1 µg/mL of Ca26Mab-6. The cells were stained with anti-mouse IgG (H + L)-Alexa Fluor 488 conjugate (1:2,000 dilution) (Cell Signaling Technology, Inc., Danvers, MA, USA). Data were acquired using the SA3800 Cell Analyzer (Sony Corporation, Tokyo, Japan) and analyzed as described previously. 25
Determination of the binding affinity by flow cytometry
CHO/CDH26 cells were treated with serial dilutions of Ca26Mab-6 (100–0.0012 μg/mL). The cells were stained with anti-mouse IgG (H + L)-Alexa Fluor 488 conjugate (1:200 dilution). The apparent dissociation constant (KD) values of Ca26Mab-6 were determined using GraphPad Prism 6 software (GraphPad Software, Inc., La Jolla, CA, USA), as described previously. 25
Results
Development of anti-CDH26 mAbs
Two female BALB/cAJcl mice were immunized with LN229/CDH26 cells (Fig. 1A). Splenocytes from the immunized mice were then fused with P3U1 myeloma cells to generate hybridomas (Fig. 1B). These hybridomas were seeded into 96-well plates, and their culture supernatants were screened using a flow cytometry-based high-throughput screening to identify supernatants that were positive for CHO/CDH26 cells but negative for parental CHO-K1 cells (Fig. 1C). Positive clones were subjected to limiting dilution to establish mAbs, resulting in the isolation of Ca26Mab-6 (mouse IgM, kappa) (Fig. 1D).
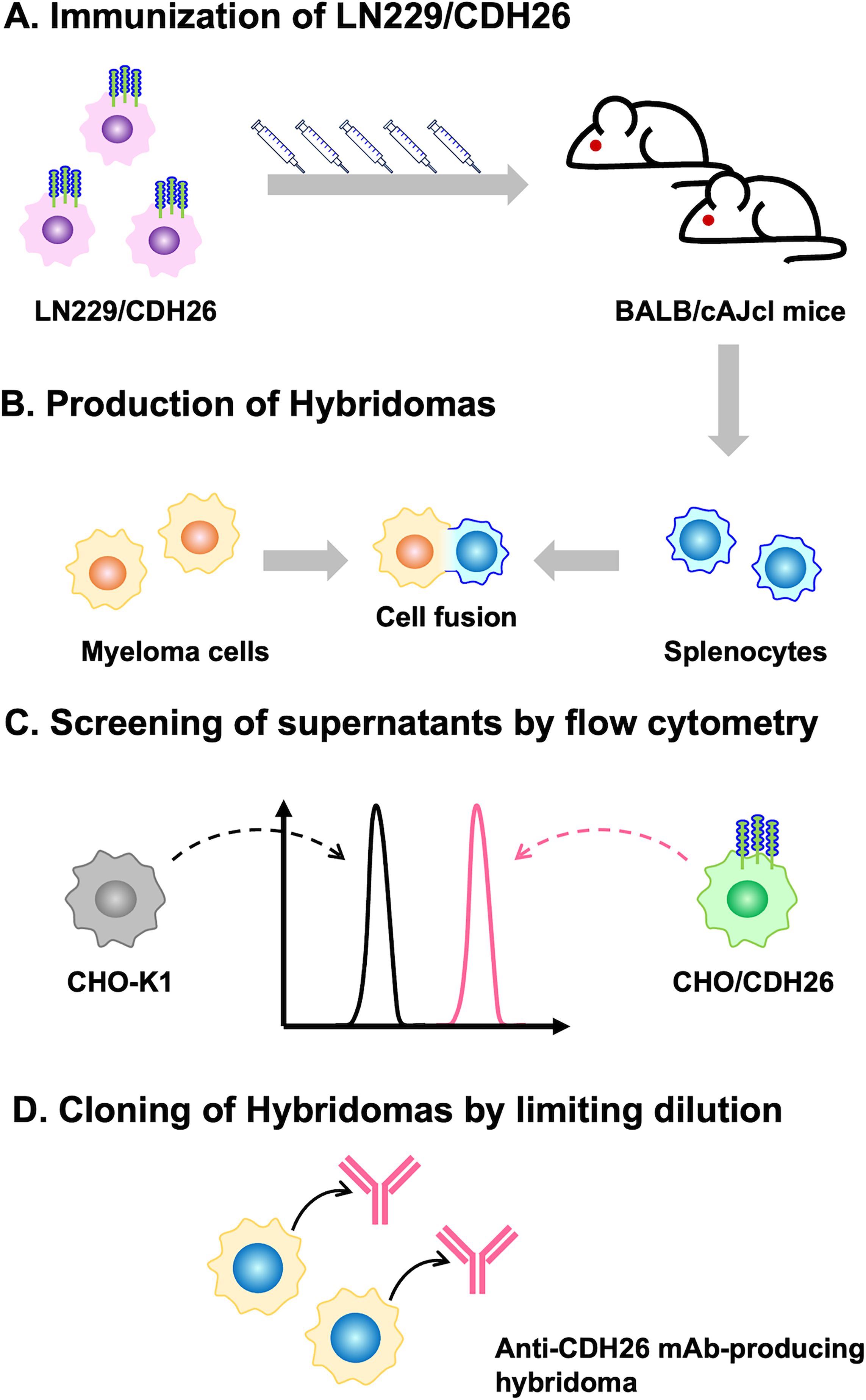
Schematic illustration of anti-CDH26 mAb production.
Investigation of the reactivity of Ca26Mab-6 using flow cytometry
The binding of purified Ca26Mab-6 to CHO/CDH26 and CHO-K1 cells was analyzed by flow cytometry. Ca26Mab-6 exhibited dose-dependent reactivity with CHO/CDH26 cells at concentrations ranging from 20 to 0.1 μg/mL but showed no binding to CHO-K1 cells at any concentration (Fig. 2A). An siRNA against CDH26 (s34069) reduces the reactivity of Ca26Mab-6 (Fig. 2B). These results indicate that Ca26Mab-6 specifically recognizes CDH26 on the cell surface. Furthermore, Ca26Mab-6 bound to HepG2 cells in a dose-dependent manner (Fig. 3A), indicating that this cell endogenously expresses CDH26. Binding to U-87 MG, MCF7, and 293FT cells was also observed as a weak but significant signal (Fig. 3B–D). These findings suggest that various types of cancer cells and immortalized cells may endogenously express CDH26.
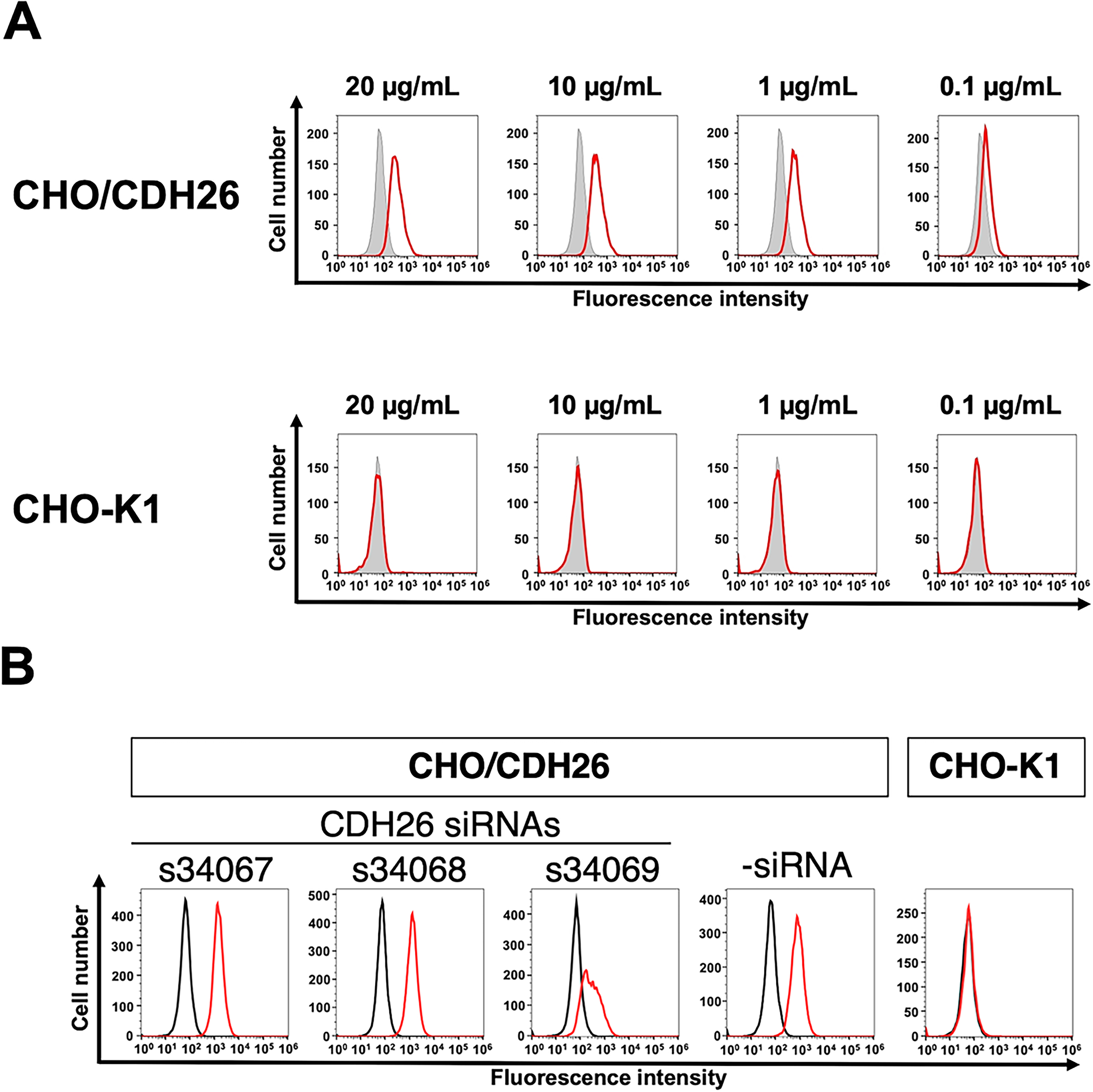
Flow cytometry analysis of an anti-CDH26 mAb against CHO/CDH26 and CHO-K1 cells.
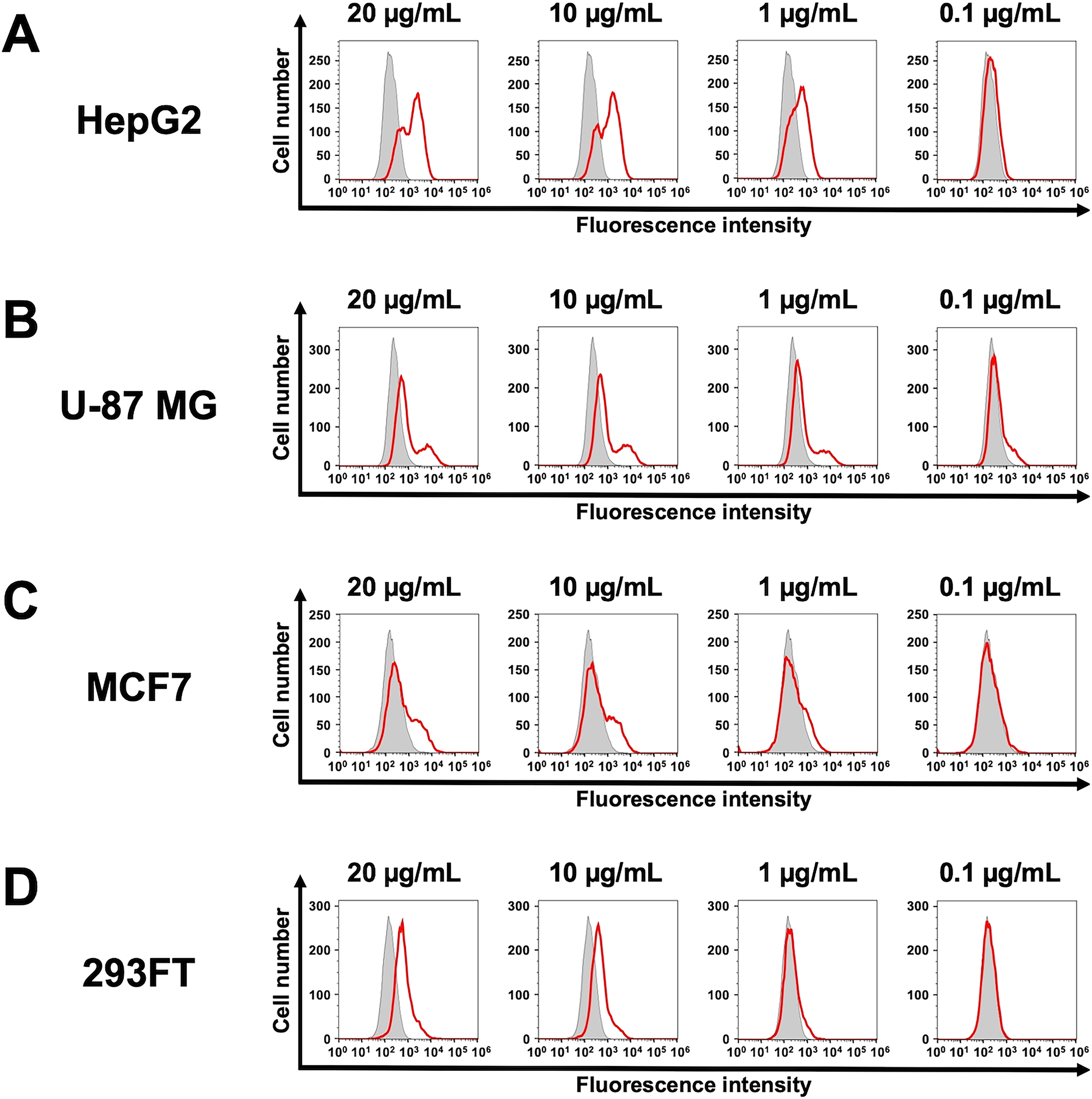
Flow cytometry analysis of an anti-CDH26 mAb against cells endogenously expressing CDH26. HepG2
Analysis of the specific reactivity of Ca26Mab-6 to CDH26 using CHO-K1 cells expressed various cadherins
We previously established CHO-K1 cells expressed type I cadherins (Supplementary Fig. S1A). 25 Furthermore, we generated stable CHO-K1 transfectants that overexpressed type II and other cadherins (Supplementary Fig. S1B). Among these cadherins-overexpressed CHO-K1, Ca26Mab-6 recognized only CHO/CDH26 cells, but not closely related type I cadherins-overexpressed CHO-K1 cells (Fig. 4A). Furthermore, Ca26Mab-6 did not show binding to type II and other cadherins-overexpressed CHO-K1 cells (Fig. 4B). Phylogram of CDH26 and closely related type I CDHs is shown in Fig. 4C. These results indicated that Ca26Mab-6 specifically recognizes CDH26.
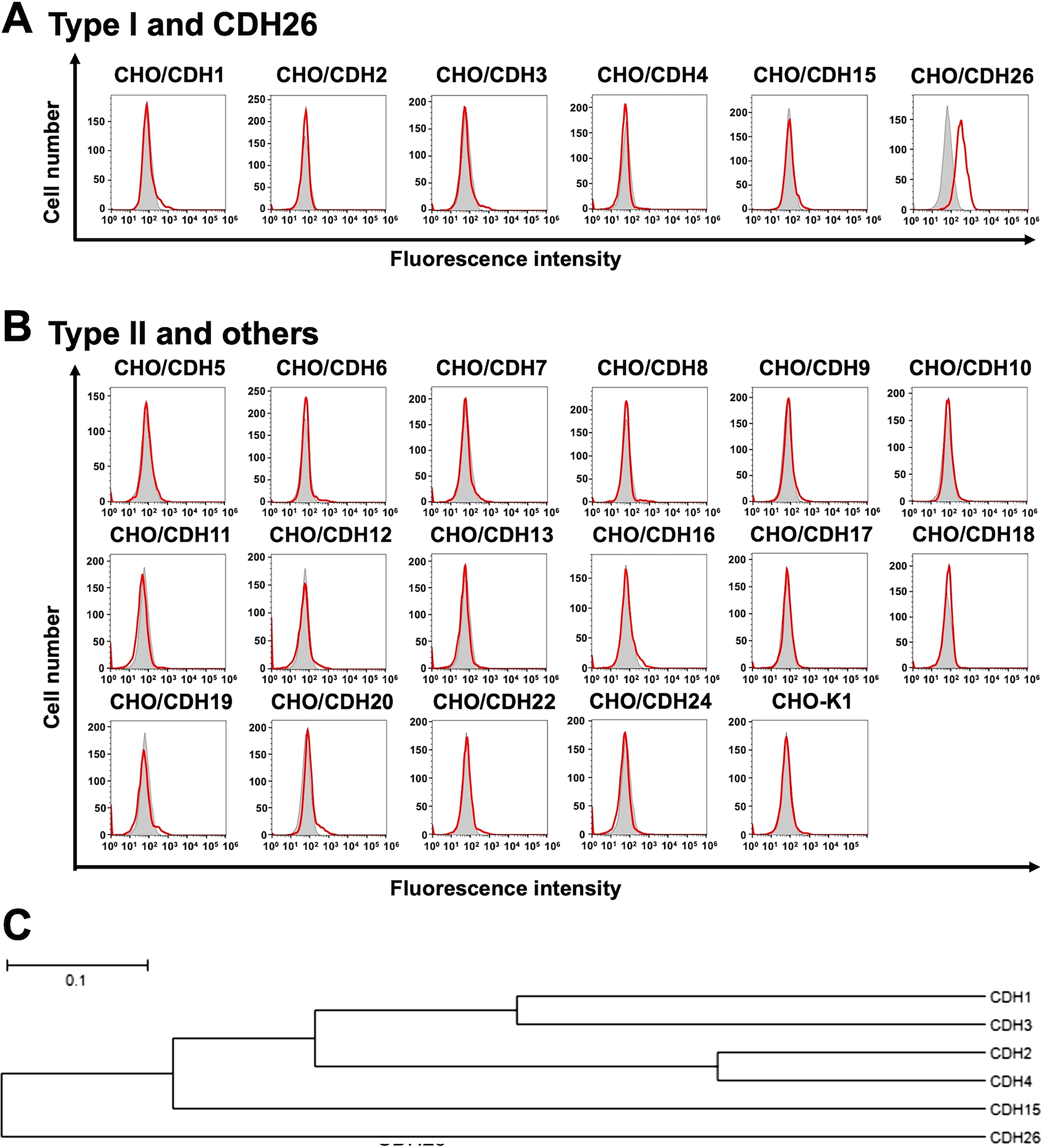
Flow cytometry analysis of cross-reactivity among cadherin-expressed cells. Type I cadherins and CDH26-overexpressed CHO-K1
Determination of apparent KD values of Ca26Mab-6 by flow cytometry
The binding affinity of Ca26Mab-6 was evaluated by flow cytometry using serial dilutions ranging from 100 to 0.0012 μg/mL. The apparent KD value of Ca26Mab-6 was 9.8 ± 4.8 × 10−9 M for CHO/CDH26 cells (Fig. 5A and Supplementary Fig. S2A) and was 3.6 ± 1.0 × 10−7 M for HepG2 cells (Fig. 5B and Supplementary Fig. S2B).
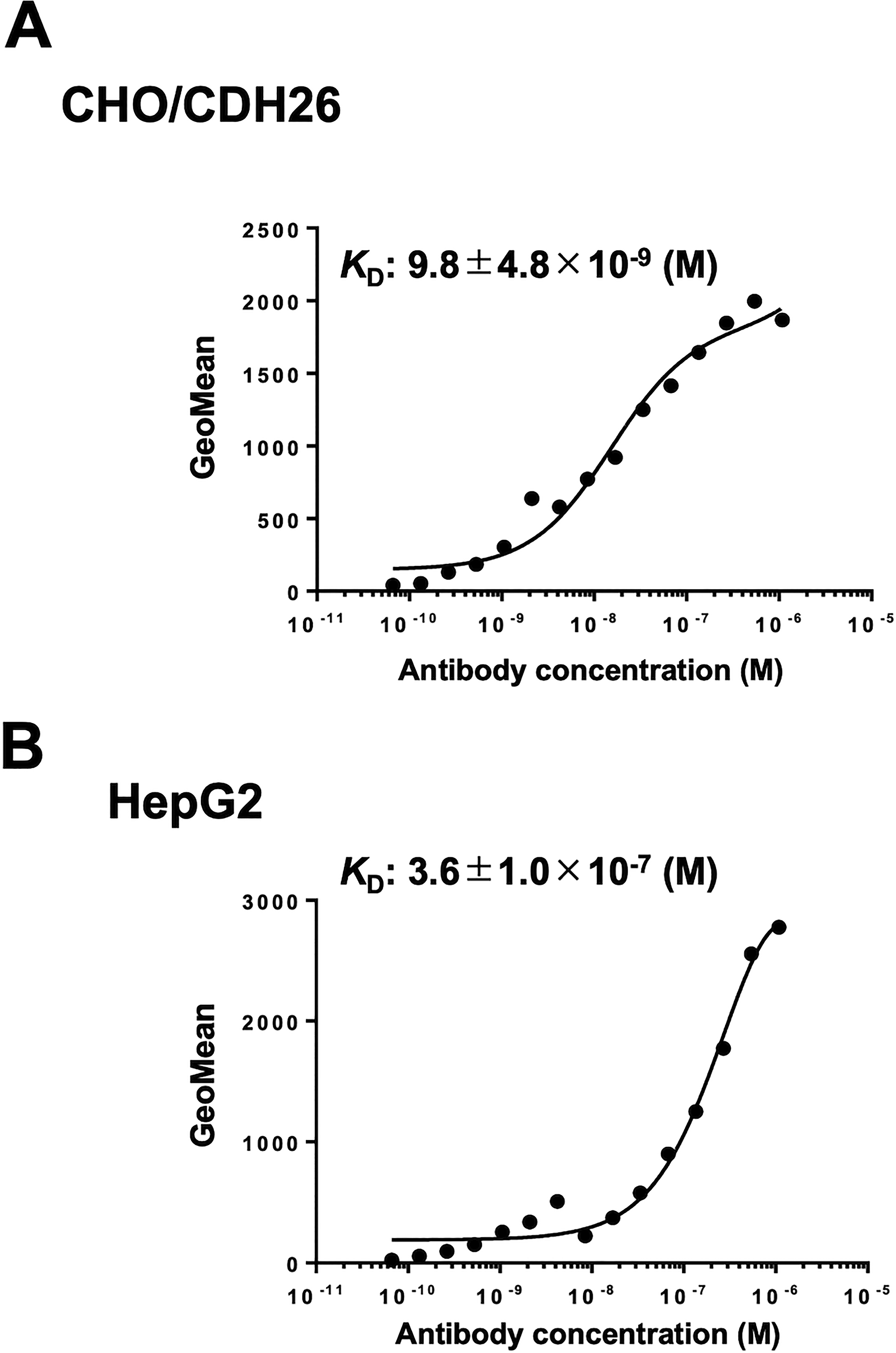
Measurement of the binding affinity of Ca26Mab-6. CHO/CDH26
Discussion
We established anti-CDH26 mAbs and demonstrated that Ca26Mab-6 is suitable for flow cytometry (Fig. 2). Ca26Mab-6 did not bind to any CHO-K1 cells that overexpressed type I or type II cadherins other than CDH26 (Fig. 4), indicating its high specificity for CDH26. We also assessed whether Ca26Mab-6 is suitable for Western blotting and immunohistochemistry. However, we were unable to detect CDH26 in these applications, indicating that Ca26Mab-6 likely recognizes a conformational epitope. Considering the absence of previous reports describing mAbs that recognize cell surface-expressed CDH26, our results provide a valuable mAb for further basic research on CDH26 expression and for the development of diagnostic and therapeutic agents.
Because Ca26Mab-6 is an IgM antibody, the higher antigen density on CHO/CDH26 cells likely facilitates multipoint binding of Ca26Mab-6. In contrast, the lower antigen density on HepG2 cells does not. This difference results in the distinct KD values observed for CHO/CDH26 and HepG2 (Fig. 5). Furthermore, differences in endogenous CDH26, such as heterogeneous glycosylation and/or cell-cycle-dependent changes in expression, may influence Ca26Mab-6 binding, as shown in Figure 3.
The expression of CDH26 in various cancer cells suggests that the CDH26 expression may play a role in carcinogenesis and that anti-CDH26 mAbs have potential as therapeutic agents. CDH26 expression has been reported in allergic gastrointestinal tissues 17 and in allergic inflammatory conditions such as rhinitis and asthma22,23; however, its role in cancer remains unclear. Since chronic inflammation is a major risk factor for cancer development and progression, 18 the correlation between CDH26 expression and malignant transformation of cells may be elucidated in future studies. For preclinical studies using mouse xenograft models, Ca26Mab-6 (IgM) should be class-switched to mouse IgG2a or human IgG1 to confer the antibody-dependent cellular cytotoxicity.
In HepG2 cells, the flow cytometry histogram displayed two distinct peaks (Fig. 3A). This may be attributed to the aneuploidy of HepG2 cells, which are known to exhibit high chromosomal instability, 29 potentially affecting CDH26 protein expression levels. Similarly distinct or broadened peaks were observed in U-87 MG and MCF7 cells (Fig. 3B and C), suggesting that similar ploidy variations of U-87 MG 30 and MCF7 31 may influence the expression level of CDH26.
Ca26Mab-6 also bound to 293FT (Fig. 3D). This cell is a derivative of HEK293, an immortalized epithelial cell derived from human embryonic kidney, suggesting that CDH26 is expressed not only in cancer cells but also in noncancer cells. Since Ca26Mab-6 is suitable for flow cytometry but is not applicable to Western blot or immunohistochemistry, it remains challenging to comprehensively evaluate the relationship between CDH26 expression levels and tumor malignancy using tissue sections. To advance research into the role of CDH26 in cancer, the development of additional anti-CDH26 mAbs that are compatible with multiple applications is essential.
In the context of developing antibody-based therapeutics targeting CDH26, the expression of CDH26 in normal cells raises concerns about potential off-target effects. To eliminate this risk, the development of cancer-specific mAbs (CasMabs) is a promising strategy. 32 CasMabs are designed to recognize aberrant conformational epitopes of overexpressed and misfolded proteins found specifically in cancer cells. 33 Therefore, the development of anti-CDH26 CasMabs is also anticipated to enhance the specificity and safety of CDH26-targeted therapies.
In conclusion, Ca26Mab-6 is a sensitive mAb suitable for basic research and holds promise for proof-of-concept studies in preclinical models aimed at developing CDH26-targeted antibody therapies.
Authors’ Contributions
H. Satofuka and H. Suzuki performed the experiments. M.K.K. and Y.K. designed the experiments. H. Satofuka, H. Suzuki, and Y.K. wrote the article. All authors have read and agreed to the published version of the article.
Footnotes
Author Disclosure Statement
No competing financial interests exist.
Funding Information
This research was supported in part by the Japan Agency for Medical Research and Development, under grant numbers JP25am0521010 (to Y.K.), JP25ama121008 (to Y.K.), JP25ama221153 (to Y.K.), JP25ama221339 (to Y.K.), and JP25bm1123027 (to Y.K.), and by the Japan Society for the Promotion of Science Grants-in-Aid for Scientific Research (KAKENHI) grant nos. 24K11652 (H. Satofuka) and 25K10553 (to Y.K.).
Supplemental Material
Supplemental Material
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.