
Abstract
Study Design
Retrospective Study.
Objective
Spinal cord injury (SCI) is a severe central nervous system trauma. To explore the regulatory mechanism of long non-coding RNA (lncRNA) NORAD in SCI.
Methods
Eighty healthy controls and 100 SCI patients were enrolled. Meanwhile, SCI animal models and LPS-induced cell models were established. Basso–Beattie–Bresnahan (BBB) scoring was performed to monitor hind-limb motor recovery. The NORAD biological function was evaluated through RT-qPCR, CCK-8 experiments, flow cytometry, enzyme-linked immunosorbent assay (ELISA), and the measurement of oxidative stress and metabolism-related indicators. The targeting relationships were verified through RIP experiments and dual-luciferase experiments.
Results
NORAD exhibited high expression in SCI patients and possessed good diagnostic utility for this disease. Its high expression may also be associated with the severity of injury. In SCI rats, inhibiting NORAD significantly improved the recovery of motor function and alleviated oxidative stress, cellular damage, and neuroinflammation, indicating a pro-injury role of NORAD during SCI pathogenesis. In addition, NORAD was highly expressed in the LPS-induced cell model, and promoted cellular damage, apoptosis, inflammatory responses, and oxidative stress by targeting and inhibiting miR-22-3p. While miR-22-3p directly targeted and inhibited PTEN, which was also highly expressed in the LPS-induced cell model. Functional rescue experiments further confirmed that the miR-22-3p inhibitor could reverse the protective effect of interfering with NORAD on SCI-related pathological damage, while silencing PTEN could offset the antagonistic effect of the miR-22-3p inhibitor, suggesting that the NORAD/miR-22-3p/PTEN axis constituted a positive feedback pro-injury loop.
Conclusions
NORAD sequestered miR-22-3p to relieve PTEN repression and exacerbate SCI.
Introduction
Spinal cord injury (SCI) refers to a series of varying degrees of motor, sensory, and autonomic nervous system dysfunction caused by damage to the spinal cord. 1 The pathological evolution of SCI is characterized by a biphasic pattern of “primary mechanical destruction-secondary cascade injury”.2,3 SCI is associated with high morbidity, disability, and mortality rates, imposing a heavy economic burden on families and society. 4 If SCI is not treated effectively and timely after spinal cord injury, a series of pathological changes will occur at the injured site, such as inflammatory response, oxidative stress, and apoptosis, etc., which will lead to further deterioration of the microenvironment at the injured site, damage to neurological function, and slow down the rate of nerve regeneration.5,6 Currently, no drugs have been proven to be effective in treating SCI. 6
Long non-coding RNAs (lncRNAs) orchestrate a spectrum of biological processes—including chromatin remodeling, transcriptional interference, and “molecular sponging”—thereby governing cell differentiation, development, inflammation, and oxidative stress.7,8 Notably, lncRNAs are involved in the progression of SCI.9-11 For instance, NEAT1 mitigates neuronal apoptosis and fosters post-SCI regeneration, 12 whereas TSIX is markedly up-regulated in patient serum and lesioned mouse spinal cord. 13 In addition, NORAD, as a highly conserved lncRNA, has received widespread attention in the field of cancer: NORAD is involved in the progression of breast cancer, 14 esophageal squamous cell carcinoma, 15 and other cancers. 16 In addition, NORAD is also involved in a variety of inflammation-related diseases. For example, NORAD, as a miR-552-3p sponge, is involved in the role of inflammation and oxidative stress in ulcerative colitis (UC). 17 Elevated NORAD is related to the worsening of coronary heart disease stenosis, inflammatory state, and blood lipids. 18 Nevertheless, the biological role of NORAD in SCI remains completely unexplored.
It is worth noting that lncRNAs commonly act as molecular sponges that sequester MicroRNAs (miRNAs) and thereby de-repress downstream mRNAs.19,20 MicroRNAs can promote the degradation of target mRNAs or inhibit their translation process.21,22 The ENCORI online database predicts that miR-22-3p is a target miRNA of NORAD. Previous studies have shown that increased miR-22-3p expression inhibits the progression of spinal cord ischemia-reperfusion (I/R) injury by suppressing IRF5 in macrophages, 23 enhances the intrinsic regenerative capacity of primary sensory neurons, 24 and is involved in neuronal apoptosis following SCI. 10 ENCORI and TargetScan bioinformatics database jointly show that phosphatase and tensin homolog deleted on chromosome ten (PTEN) is a potential target gene of miR-22-3p. PTEN plays a key role in regulating the PI3K/Akt pathway and is involved in cell growth, proliferation, and survival. 25 Notably, in astrocytes surrounding synaptophysin puncta (APAS), the PTEN expression is significantly increased after SCI. 26 Knocking out PTEN using adeno-associated viruses (AAVrg) can restore motor function in chronic SCI. 27 Moreover, PTEN upregulation in the spinal cord tissues of SCI mice inactivates the PI3K/AKT pathway and exacerbates SCI. 28 In view of the above evidence, we speculate that in the SCI microenvironment, NORAD may competitively adsorb miR-22-3p through a “sponge” mechanism, relieve its inhibition of PTEN, and participate in regulating the progression of SCI.
To verify the above conjecture, we further analyzed the expression pattern and potential molecular mechanisms of NORAD in SCI through clinical data of SCI patients, SCI animal models, and LPS-induced in vitro cell experiments, which may provide new ideas for the progression of SCI.
Materials and Methods
Participants
This was a retrospective study. To clarify the expression pattern and clinical significance of long non-coding RNA NORAD in spinal cord injury (SCI), we consecutively included 100 patients with acute spinal cord injury who visited our hospital from August 2021 to August 2024 and met the following criteria. Patient selection strictly adhered to pre-defined objective criteria to ensure the representativeness of the study population and to avoid selection bias. Inclusion criteria: (1) > 18 years old, (2) patients with SCI confirmed by CT and other imaging examinations, (3) Time from injury to admission ≤12 h. Exclusion criteria: (1) History of spinal or craniocerebral trauma or surgery within the past 6 months, (2) Subjects with severe cardiovascular diseases, such as myocardial infarction, arrhythmia, (3) Pregnant women, (4) Patients with malignant tumors, liver and kidney insufficiency, cerebrovascular diseases, and infectious diseases.
According to the ASIA Impairment Scale in the International Standards for Neurological Classification of SCI, all patients were divided into three groups: neurologically normal group (ASIA-E, n = 29), incomplete SCI group (ASIA-B, C, and D, n = 48), and complete SCI group (ASIA-A, n = 23). The normal neurological function group corresponds to Grade E on the ASIA scale, referring to patients with normal sensory and motor functions after SCI.
During the same period, 80 healthy controls were recruited from the physical examination center. The age and gender distribution of the control group matched that of the SCI patient group to control the potential influence of these baseline characteristics on the results. They had no history of spinal cord injury, no spinal disease, normal motor function of the limbs, no limb deformity, paralysis, or sensory impairment, and had not received drug treatment that may affect the nervous system in the past 3 months, and had no serious systemic diseases.
Venous blood samples were collected from all participants at admission or during physical examination. Serum was separated by centrifugation and stored at −80°C.
This study was approved by the Ethics Committee of author’s institution (No. 202106), and all participants signed informed consent forms.
SCI Rat Model
Forty specific-pathogen-free (SPF) male Sprague–Dawley rats, 7-8 weeks old and weighing 250 ± 8.79 g, were housed under controlled conditions (22-25°C, 50-60 % humidity, 12 h light/dark cycle) with ad libitum access to food and water. The animals were divided into four groups according to the random number table method (with n = 10 in each group) according to the experimental protocol.
The Allen’s weight-drop method was used to establish SCI models.29,30 After disinfecting the dorsal skin of rats, ketamine (60 mg/kg) and xylazine (5 mg/kg) were injected for anesthesia. Subsequently, the skin and subcutaneous tissue of the rats were dissected at the T10 vertebral body. After dissecting the muscles, the spinous processes and spinal lamina were exposed. After removing the T10 spinal plate with a laminar curette, a heavy hammer (10 g) was used to impact the exposed spinal cord from a height of 5 cm, resulting in an impact force of 50 g, which severely damaged the spinal cord. Tail spasticity and hindlimb paralysis were characteristic features of the SCI model. The incision was then sutured. To prevent infection, rats were injected intramuscularly with penicillin for 3 consecutive days after surgery. All animals had free access to food and water. Rats in the sham-operated group underwent only a laminectomy without any SCI. Five days after the surgery, the lentiviral vectors (produced by GenePharma) expressing si-NC and si-NORAD plasmids were injected into the rat. The choice to administer lentiviral vectors on day 5 post-injury was based on the following considerations: by this time, the peak period of acute hemorrhage and edema has subsided, and the blood-spinal cord barrier showed a degree of recovery, which facilitates vector delivery. Simultaneously, the molecular and cellular events of secondary injury (such as sustained inflammation and oxidative stress) remain active during this phase, making it a critical window for molecular intervention.
At 6, 24, 48, and 72 hours after SCI modeling, the motor function of the rat hind limbs was independently assessed by two researchers blinded to the group allocation using the Basso, Beattie, and Bresnahan (BBB) scoring scale, and the final score was the average of the two evaluators’ scores. The full score was 21 points, with 0 points indicating complete paralysis and 21 points indicating normal movement. Postoperatively, manual bladder emptying was assisted twice daily until the autonomous urination function recovered.
After anesthesia with 4% pentobarbital, the rats were sacrificed, and the spinal cord tissues were collected.
Cell Culture and Transfection
Neuronal PC12 cells (Beijing Biowit NTCC, China) were maintained in DMEM medium supplemented with 10 % fetal bovine serum (FBS) and 1 % penicillin–streptomycin at 37°C, 95% humidity, and 5 % CO2.
Si-NC and si-NORAD/PTEN, inhibitor/mimic-NC, miR-22-3p-inhibitor/mimic, and the corresponding negative control were all synthesized by Shanghai GenePharma. The co-transfection was performed using Lipofectamine 2000 (Invitrogen) according to different experimental groups.
To simulate the inflammatory microenvironment after SCI in vitro, PC12 cells were treated with lipopolysaccharide (LPS; 100 ng/ml) for 24 hours.
RT-qPCR
Total RNA was extracted from spinal cord tissues or cells using Trizol reagent. 1 μg RNA was reverse-transcribed into cDNA using a reverse transcription kit. RT-qPCR was performed on an ABI PRISM 7500 system (Applied Biosystems) with SYBR Premix Ex Taq II (Takara) using the above cDNA as template. Gene expression was calculated by the 2–ΔΔCt method and normalized to U6 (for miRNA) or GAPDH (for mRNA).
Cell Proliferation
Cell viability was analyzed using the Cell Counting Kit-8 (CCK-8; Beyotime, China). Transfected PC12 cells were seeded into 96-well plates, and at each specified time point, 10 μL of CCK-8 solution was added to each well. The absorbance of each well at 450 nm was measured.
Cell Apoptosis
Apoptosis was quantified with the Annexin V-FITC/PI kit (Invitrogen). PC12 cells were harvested, resuspended in 100 μL binding buffer, and stained with 5 μL Annexin V-FITC plus 5 μL PI for 15 min in the dark. The apoptotic fraction (%) was immediately analyzed on a FACSCalibur flow cytometer (BD Biosciences).
Enzyme-Linked Immunosorbent Assay (ELISA)
The contents of tumor necrosis factor-α (TNF-α), interleukin-1β (IL-1β), and interleukin-6 (IL-6) in spinal cord tissue or cell culture supernatant were measured using corresponding ELISA kits (Invitrogen).
Measurement of Oxidative Stress Indicators
Malondialdehyde (MDA) content, lactate content, and lactate dehydrogenase (LDH) activity in spinal cord tissues and cells were determined using the MDA, lactate, or LDH detection kit (Nanjing Jiancheng Bioengineering Institute).
RNA Immunoprecipitation Assay (RIP)
RIP was performed using the RNA Immunoprecipitation Kit. Cell pellets were lysed in 1 mL ice-cold RIP buffer. The lysate was incubated overnight with 5 µg anti-Ago2 antibody or control IgG pre-conjugated to protein A/G magnetic beads. After protein removal by Proteinase K treatment at 55°C for 30 min, co-precipitated RNA was extracted, and the enrichment of target transcripts was quantified by RT-qPCR.
Dual Luciferase Assay
Binding sequences complementary to miR-22-3p in NORAD or PTEN were predicted using bioinformatics tools (TargetScan and ENCORI). Wild-type luciferase reporter vectors (pmirGLO-NORAD-WT and pmirGLO-PTEN-WT) containing NORAD/PTEN fragments with sequences complementary to miR-22-3p, as well as mutant vectors (pmirGLO-NORAD-MUT and pmirGLO-PTEN-MUT) with site-directed mutations at the binding sites, were constructed. These vectors were co-transfected into HEK293 T cells with miR-22-3p mimic/inhibitor or corresponding negative controls using Lipofectamine 2000.
Gene Ontology (GO) and Kyoto Encyclopedia of Genes and Genomes (KEGG) Analysis
The target genes were selected through the intersection of the ENCORI, miRDB, and TargetScan databases. Perform signal pathway analysis on the common target genes of the three databases using GO and KEGG.
Statistical Analysis
Data analysis was performed using SPSS 21.0 software. Measurement data were expressed as mean ± standard deviation. Comparisons between two groups were performed using the independent samples t-test; comparisons among multiple groups were performed using one-way ANOVA, with post hoc pairwise comparisons conducted using the LSD test. Categorical data were expressed as frequencies, and comparisons between groups were performed using the chi-square test. The receiver operating characteristic curve (ROC) was used to evaluate the diagnostic value of NORAD for SCI, and Pearson correlation coefficient analysis was used to verify the relationship between expression levels. P < 0.05 was considered statistically significant.
Results
Expression and Diagnostic Value of NORAD in SCI
The General Clinical Characteristics of the Subjects
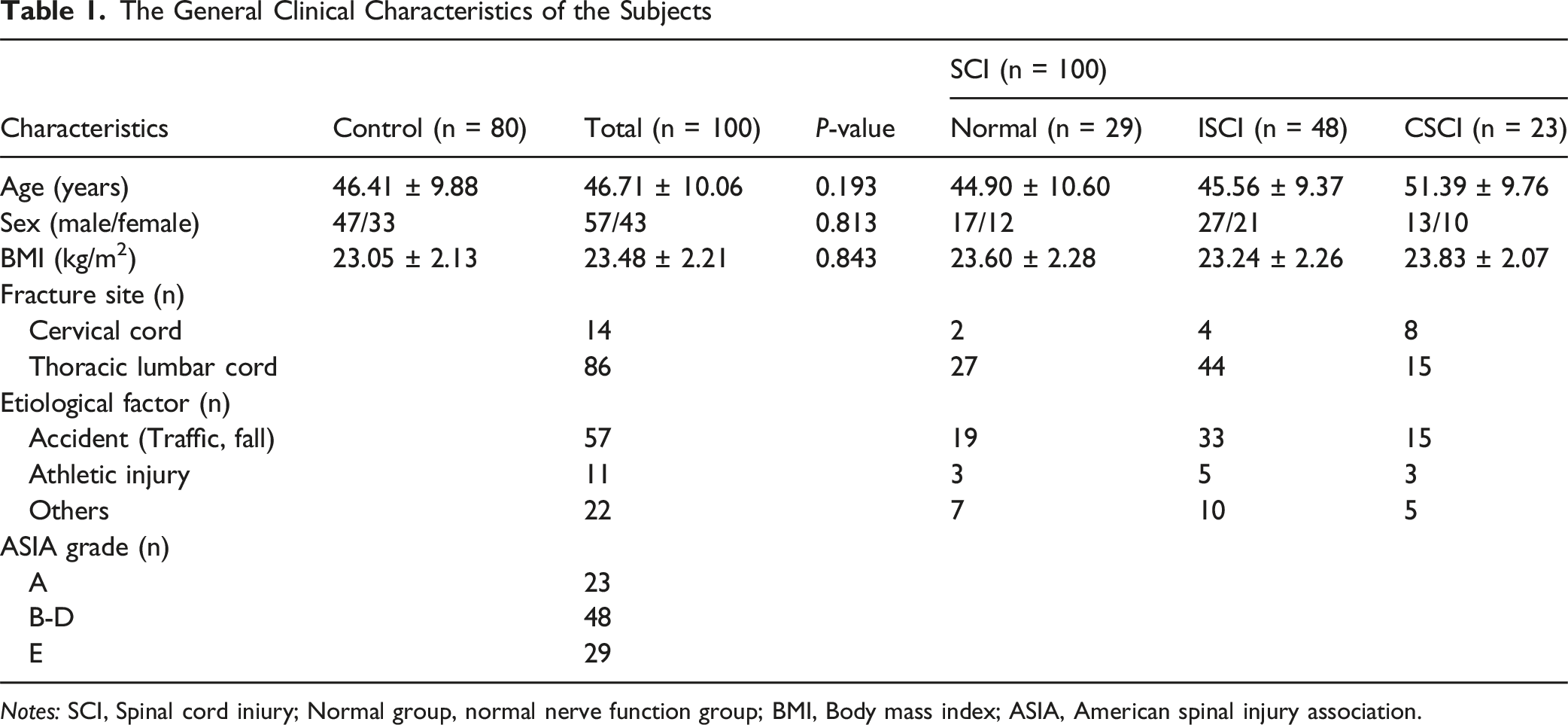
Notes: SCI, Spinal cord iniury; Normal group, normal nerve function group; BMI, Body mass index; ASIA, American spinal injury association.
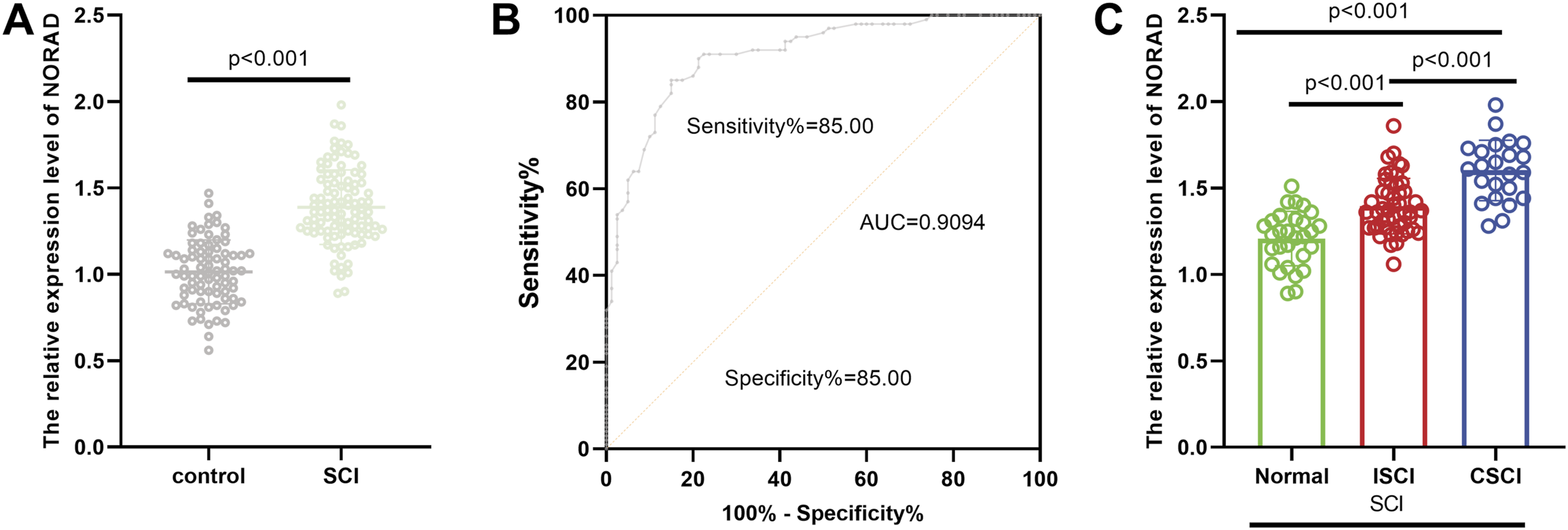
Expression and diagnostic value of NORAD in SCI. (A) NORAD expression was significantly upregulated in the SCI group compared to the control group. (B) ROC curve analysis demonstrated that NORAD has considerable diagnostic potential for SCI. (C) The expression level of NORAD was positively correlated with the severity of SCI, suggesting its clinical relevance
The Influence of NORAD on the SCI Rat Model
The expression of NORAD was low in the Sham (sham operation) group and significantly increased in the SCI group. After interfering with NORAD, its expression was significantly reduced (Figure 2A). Motor function was evaluated by the BBB score (spinal cord function score). Scores were significantly lower in the SCI group, and interfering with NORAD improved motor function after SCI (Figure 2B). The MDA level in the SCI group was significantly higher than that in the Sham group, and interfering with NORAD could alleviate oxidative stress damage after SCI (Figure 2C). LDH activity and lactate were significantly increased in the SCI group, and interfering with NORAD could reduce this increase (Figure 2D–E). The levels of various inflammatory factors (TNF-α, IL-6, IL-1β) in the SCI group were significantly higher than those in the Sham group. After interfering with NORAD, the levels of these inflammatory factors were significantly reduced, suggesting that interfering with NORAD can inhibit the inflammatory response after SCI (Figure 2F). Effects of NORAD knockdown on functional outcomes in a rat model of SCI. (A) NORAD expression was low in the sham group, markedly elevated in the SCI group, and effectively reduced following NORAD interference. (B) Silencing NORAD significantly improved motor function recovery after SCI. (C) ORAD interference attenuated oxidative stress injury post-SCI. (D–E) SCI induced a significant increase in LDH activity and lactate levels, both of which were substantially reduced by NORAD knockdown. (F) Inhibition of NORAD suppressed the inflammatory response following SCI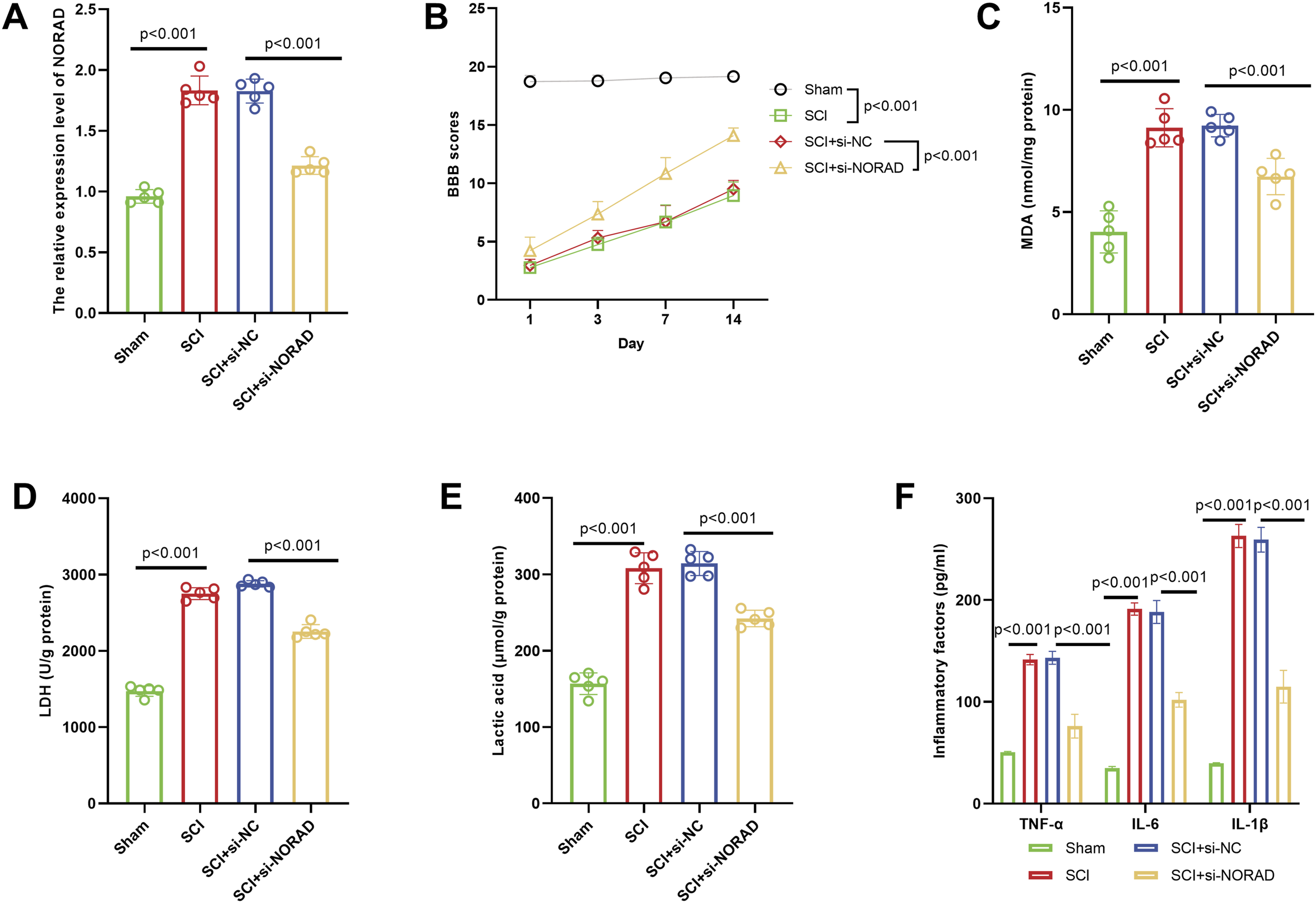
NORAD Participated in LPS-Induced Inflammatory Injury by Targeting miR-22-3p
After LPS treatment, the expression of NORAD significantly increased, and after interfering with NORAD, its expression decreased significantly (Figure 3A). In the LPS group, cell proliferation was inhibited, and the apoptosis rate was increased. NORAD silencing significantly restored cell proliferation capacity and reduced the apoptosis rate (Figure 3B–C). Levels of inflammatory factors were markedly elevated in the LPS group, while NORAD silencing notably downregulated these factors (Figure 3D). Additionally, the LPS group exhibited significantly increased levels of LDH (cellular damage), MDA (oxidative stress), and lactic acid (metabolic disorders), and these indicators were obviously decreased following NORAD silencing (Figure 3E–G). The ENCORI database revealed that there is a binding site between NORAD and miR-22-3p (Figure 3H). The RIP experiment showed that the Ago2 antibody could significantly enrich NORAD and miR-22-3p, indicating a direct binding between the two (Figure 3I). The dual luciferase reporter assay demonstrated that the miR-mimic could inhibit the luciferase activity of WT-NORAD, but did not affect MUT-NORAD, further confirming the targeted binding of NORAD and miR-22-3p (Figure 3J). The miR-22-3p expression was significantly lower in the SCI group, suggesting a potential anti-inflammatory role of miR-22-3p in SCI (Figure 3K). Correlation analysis revealed a significant negative correlation between NORAD and miR-22-3p expression (Figure 3L). Additionally, miR-22-3p expression was significantly decreased after LPS treatment, while it was notably increased following NORAD silencing, which further supported the negative regulatory effect of NORAD on miR-22-3p (Figure 3M). NORAD exacerbated LPS-induced inflammatory injury by targeting miR-22-3p. (A) LPS treatment significantly increased NORAD expression, which was reversed upon NORAD interference. (B-C) LPS inhibited cell proliferation and promoted apoptosis, these effects were reversed by NORAD silencing, as evidenced by restored proliferation and reduced apoptosis. (D) Levels of pro-inflammatory cytokines (TNF-α, IL-6, and IL-1β) were significantly elevated in the LPS group but markedly decreased after NORAD knockdown. (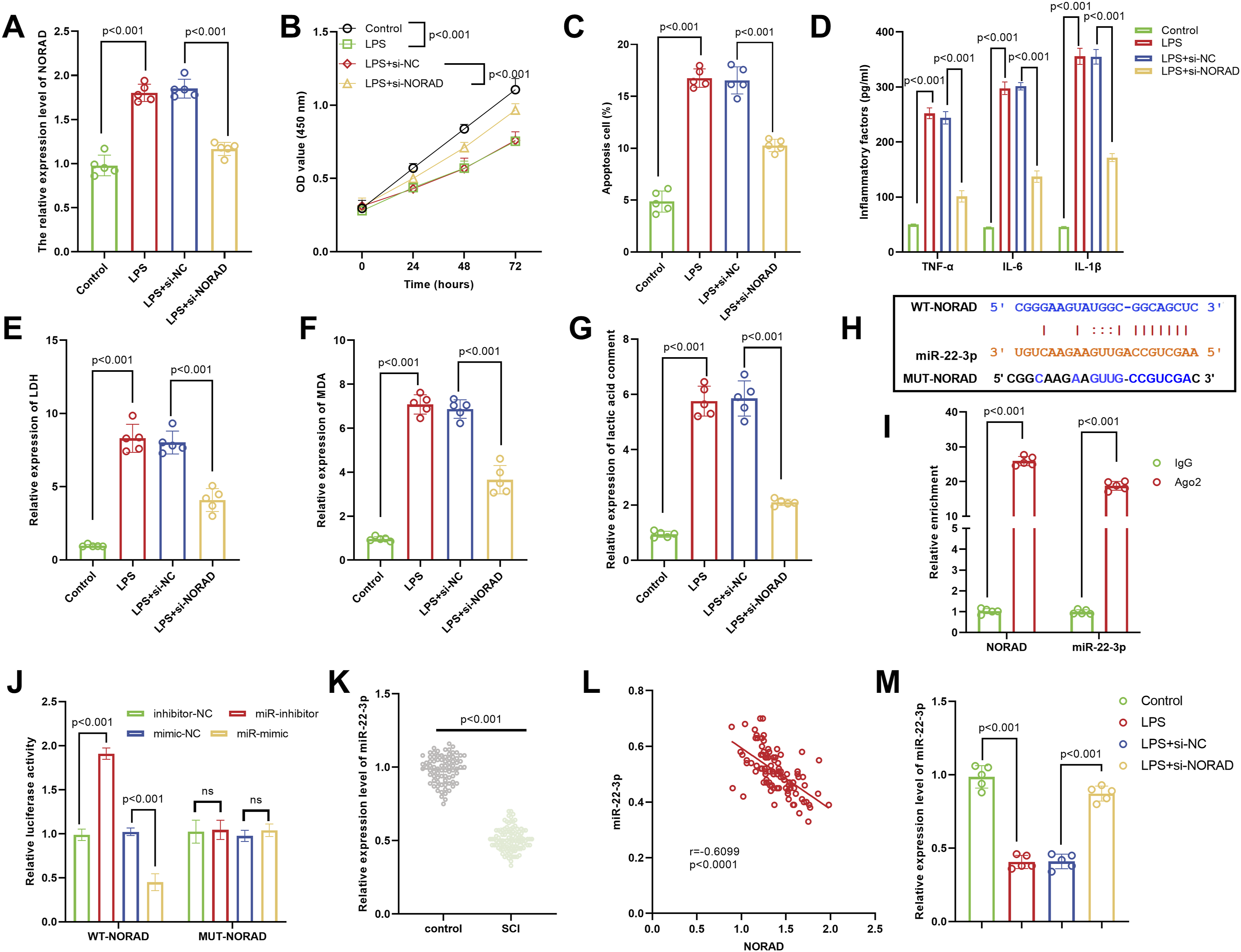
Mechanism of the NORAD/miR-22-3p/PTEN Axis in LPS-Induced Inflammatory Injury
After interfering with NORAD, the miR-22-3p expression was increased. Co-transfection with miR-22-3p inhibitor could significantly reduce its expression (Figure 4A). In the LPS-induced cell model, the partial reversal effect of miR-22-3p inhibitor on the improvement of LPS damage caused by interference with NORAD was manifested as follows: co-transfection with miR-22-3p inhibitor could reduce cell proliferation (Figure 4B), promote cell apoptosis (Figure 4C), promote inflammatory response (Figure 4D), and increase LDH (cell damage), MDA (oxidative stress), and lactate (metabolic disorder) levels (Figure 4E–G). Enrichment analysis indicated that NORAD-related pathways are significantly associated with the PI3K-Akt signaling pathway (Figure 4H). Bioinformatics prediction revealed putative binding sites between miR-22-3p and the 3′-UTR of PTEN (Figure 4I). RIP assays demonstrated that the Ago2 antibody markedly enriched both miR-22-3p and PTEN, confirming their direct interaction within the RNA-induced silencing complex (Figure 4J). Dual-luciferase reporter assays further showed that miR-22-3p mimics specifically suppressed the luciferase activity of wild-type PTEN-3′-UTR, whereas no effect was observed with mutant constructs, validating the direct targeting of PTEN by miR-22-3p (Figure 4K). In the SCI group, the PTEN expression was significantly higher, suggesting its pro-damaging effect (Figure 4L). A significant negative correlation between miR-22-3p and PTEN expression (Figure 4M). After interfering with NORAD, the PTEN expression decreased, and the miR-22-3p inhibitor could partly reverse this reduction effect, further confirming that NORAD regulated PTEN through miR-22-3p (Figure 4N). Mechanistic insights into the NORAD/miR-22-3p/PTEN axis in LPS-induced inflammation. (A) NORAD knockdown increased miR-22-3p expression, an effect reversed by co-transfection with a miR-22-3p inhibitor. (B–G) In the LPS-induced injury model, inhibition of miR-22-3p partially reversed the protective effects of NORAD silencing, as shown by suppressed cell proliferation (B), enhanced apoptosis (C), exacerbated inflammation (D), and elevated levels of LDH (E), MDA (F), and lactate (G). (H) NORAD may regulate the PI3K-Akt signaling pathway. (I) Public databases (ENCORI and TargetScan) predicted conserved binding sites between miR-22-3p and the 3'UTR of PTEN. (J) RIP assays confirmed direct interaction between miR-22-3p and PTEN via Ago2 immunoprecipitation. (K) Dual-luciferase reporter assays confirmed that miR-22-3p directly targeted the 3'UTR of PTEN. (L) PTEN expression was significantly higher in SCI tissues than in controls, indicating its detrimental role in injury progression. (M) (A strong negative correlation was observed between miR-22-3p and PTEN expression. (N) NORAD interference reduced PTEN expression, and this reduction was reversed by miR-22-3p inhibition, confirming that NORAD regulates PTEN in a miR-22-3p–dependent manner. ns indicates no statistical significance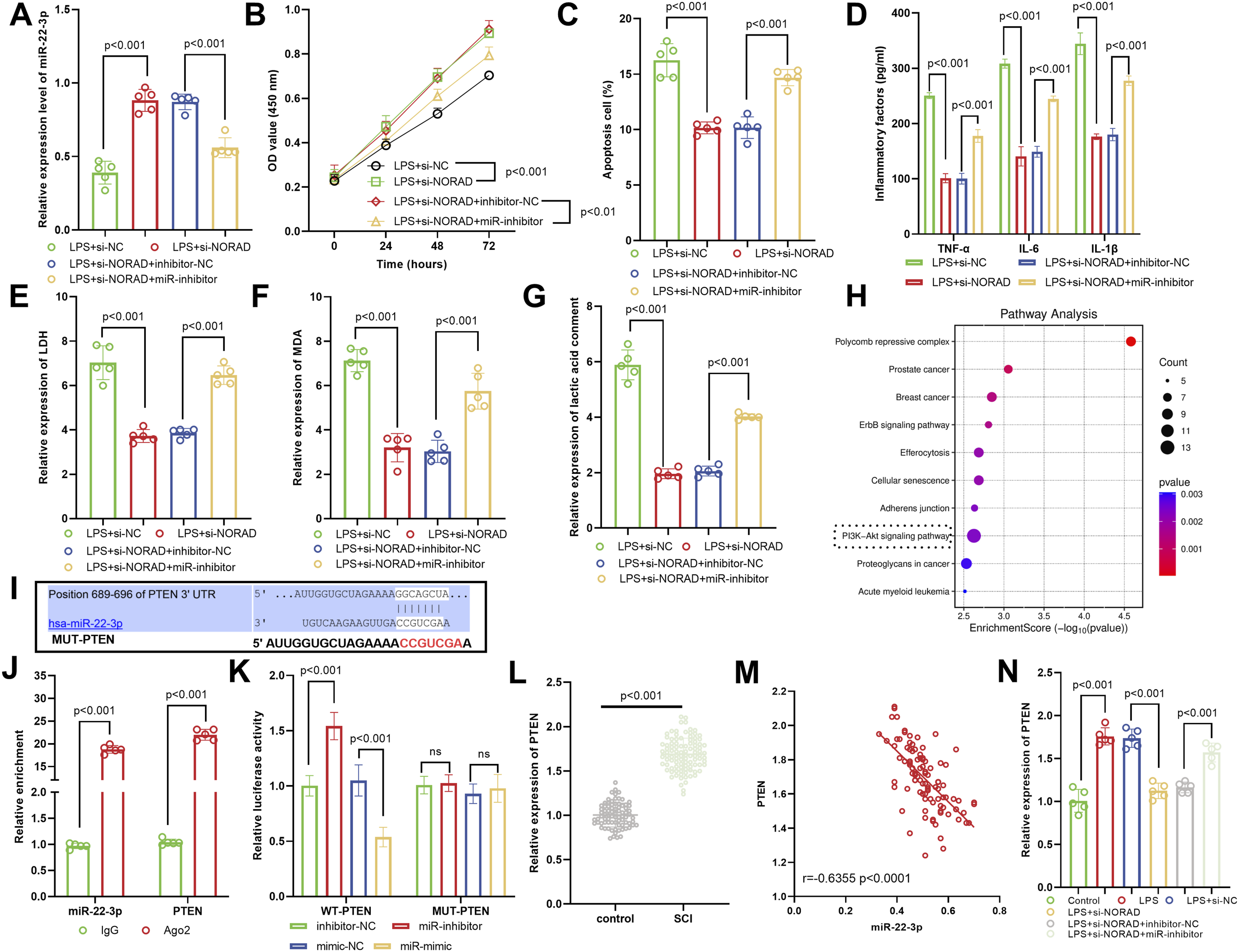
Functional Verification of the NORAD/miR-22-3p/PTEN Axis in LPS-Induced Inflammatory Injury
In the NORAD-silenced + miR-22-3p-inhibitor group, PTEN expression was markedly elevated, and co-transfection with si-PTEN efficiently abolished this increase, confirming the knockdown efficacy of si-PTEN (Figure 5A). Functionally, PTEN depletion reversed the antagonistic effects imposed by the miR-22-3p inhibitor on NORAD silencing: cell proliferation was restored (Figure 5B), apoptosis rate dropped significantly (Figure 5C), and the levels of pro-inflammatory cytokines were substantially reduced (Figure 5D). Concomitantly, indices of cellular injury (LDH release), oxidative stress (MDA content), and metabolic imbalance (lactate accumulation) all declined markedly (Figure 5E–G). Functional rescue experiments validated the NORAD/miR-22-3p/PTEN axis in LPS-induced injury. (A) Co-transfection of si-PTEN effectively suppressed PTEN expression in cells with NORAD knockdown and miR-22-3p inhibition, confirming efficient PTEN silencing. (B–F) PTEN knockdown reversed the detrimental effects of miR-22-3p inhibition on NORAD-silenced cells, as evidenced by restored cell proliferation (B), reduced apoptosis (C), attenuated inflammatory cytokine release (D), and decreased levels of LDH (E), MDA (F), and lactate (G), collectively demonstrating that PTEN was a critical downstream effector in this regulatory axis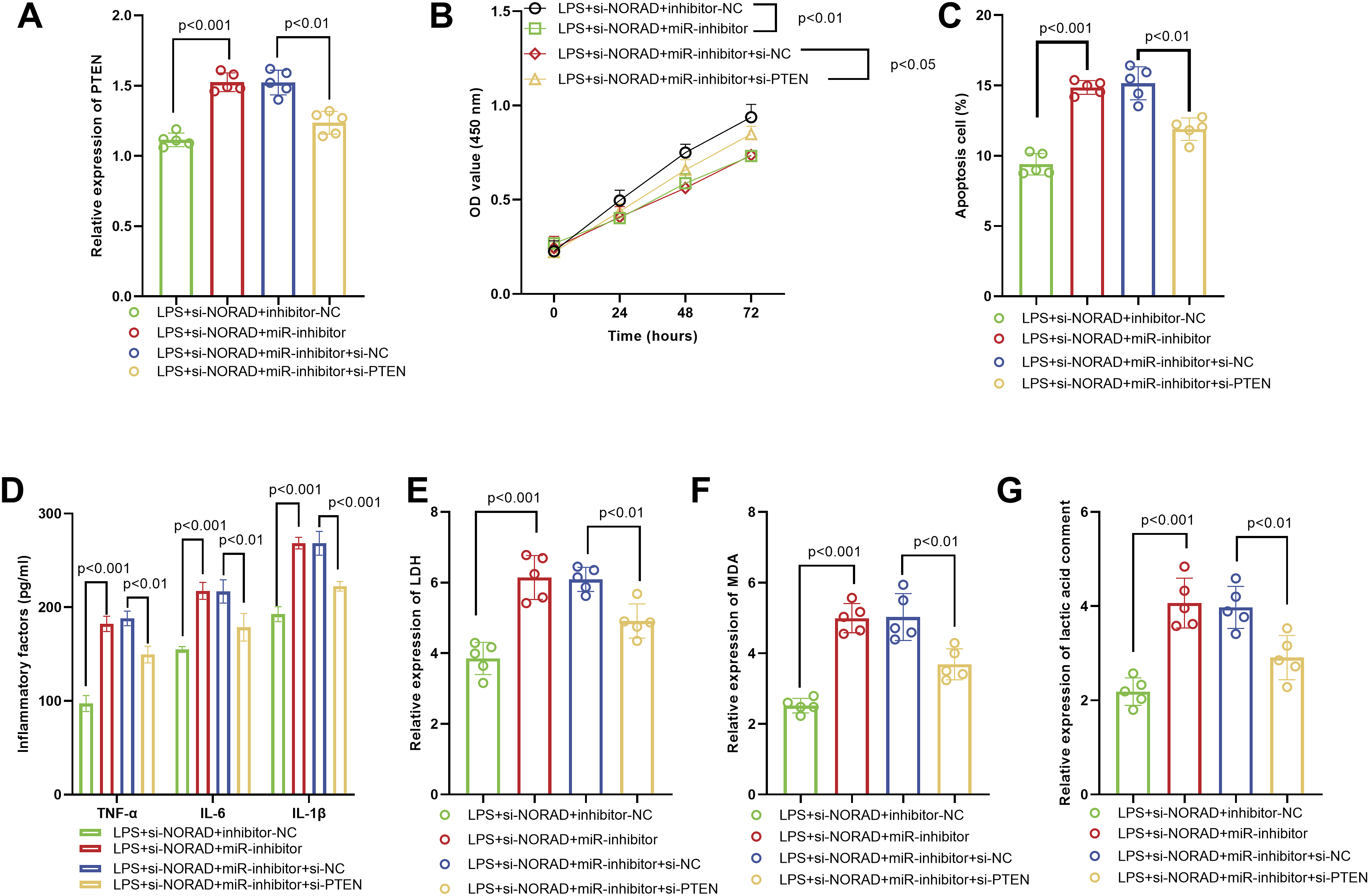
Discussion
SCI is a severe traumatic disease of the central nervous system, 31 and so far, there is still no ideal treatment. 32 It is worth noting that NORAD plays a pro-inflammatory and pro-injury role in a variety of diseases,33-35 but its function and regulatory network in SCI are still unclear. Through the analysis of clinical samples and the construction of SCI rats and cell models, this study demonstrated that the NORAD expression was up-regulated in SCI patients and SCI models. Mechanistically, our work newly identified that NORAD could remove its inhibition on PTEN by adsorbing miR-22-3p and aggravate LPS-induced cell damage, apoptosis, inflammatory response, and oxidative stress.
NORAD competes with miR-28-3p for binding through E2F2, promoting the progression of lung cancer (LC). 36 In diabetic nephropathy, knockdown of NORAD can inhibit the inflammation of human mesangial cells (HMCs), 37 and the upregulation of NORAD participates in the apoptosis, inflammation, and oxidative stress of ulcerative colitis through the nuclear factor-κB signaling pathway. 17 It is worth noting that our study was consistent with previous research. Our study found that NORAD played a pro-injury role in the pathological process of SCI, and its high expression may further worsen neurological function by exacerbating inflammatory responses and oxidative stress, aggravating cell damage, and metabolic imbalance. Specifically, the NORAD expression in SCI patients was significantly increased, and it had a high diagnostic value. Subgroup analysis showed that the expression level of NORAD might be closely related to the severity of SCI. It was speculated that NORAD may participate in the pathological injury process after SCI. To define the biological role of NORAD in SCI, we established both rat and cell models of SCI. Silencing NORAD markedly improved locomotor recovery (elevated BBB scores) and concurrently reduced oxidative stress (MDA), cellular damage (LDH), metabolic derangement (lactate), and the pro-inflammatory cytokines TNF-α, IL-6, and IL-1β.
The classic “ceRNA” mechanism of lncRNAs involves sponging miRNAs to relieve their inhibitory effects on downstream mRNAs, thereby exerting regulatory functions.38,39 Bioinformatics predictions, RIP, and dual-luciferase assays all verified the direct targeting relationships between NORAD and miR-22-3p, as well as between miR-22-3p and PTEN. Previous studies have confirmed that NORAD promotes the progression of myocardial infarction by targeting the miR-22-3p/PTEN axis, 40 which is consistent with our findings. This study found that NORAD may target and adsorb miR-22-3p through the ceRNA mechanism, thereby regulating the expression of the downstream target gene PTEN, forming the NORAD/miR-22-3p/PTEN regulatory axis, which participates in the pathological progression of SCI. Specifically, in SCI patients, the expression of miR-22-3p was significantly decreased while the expression of PTEN was increased. Moreover, NORAD was significantly negatively correlated with miR-22-3p, and miR-22-3p was negatively correlated with PTEN. It is worth noting that miR-22-3p inhibits uric acid sodium-induced gout inflammation by targeting NLRP3. 41 Enrichment of miR-22 accelerates axonal regeneration and remyelination, fostering neurological recovery after SCI. 42 Moreover, the elevated miR-22-3p promotes the M2 polarization of macrophages, inhibits tissue inflammation by suppressing IRF5, and thereby alleviates spinal cord I/R injury. 23 Our study also demonstrated that miR-22-3p exerted a protective role in SCI. Functional rescue experiments further confirmed that miR-22-3p inhibitors could partly reverse the protective effects of NORAD silencing on SCI-related pathological damage, while PTEN silencing could counteract the antagonistic effect of miR-22-3p inhibitors. These results clarify that NORAD exacerbated post-SCI inflammation, oxidative stress, and cellular damage by inhibiting miR-22-3p expression and relieving its targeted suppression of PTEN.
PTEN silencing combined with chABC overexpression in adipose-derived stem cells promotes functional recovery of SCI in rats. 43 Additionally, PTEN has been demonstrated to be upregulated in the spinal cord tissues of mice with SCI, which may deactivate the PI3K-Akt signaling pathway and promote the progression of SCI. 28 It is worth noting that knockdown of miR-429 inhibits the PI3K/Akt pathway by upregulating PTEN and participates in the regulation of neuronal apoptosis after SCI. 44 This study also suggested that the NORAD-related pathway involved the PI3K-Akt signaling pathway through pathway enrichment analysis. It was speculated that the NORAD/miR-22-3p/PTEN axis may participate in the pathological process of SCI by regulating this pathway. This mechanism needs to be further verified by subsequent experiments.
Collectively, the novel contributions of this study are threefold. First, it established the clinical relevance of lncRNA NORAD in human SCI, correlating its elevated expression with disease severity and identifying its diagnostic potential. Second, it defined a previously unreported pathogenic signaling axis in SCI—the NORAD/miR-22-3p/PTEN positive feedback loop—which drove oxidative stress, neuroinflammation, and neuronal apoptosis. Third, it provided functional preclinical validation that targeting this axis, particularly by silencing NORAD, mitigated functional impairment and pathology in vivo. While NORAD’s pro-inflammatory role was recognized in other diseases, this study uniquely delineates its mechanism and therapeutic relevance specifically for SCI, offering a new target for modulating the secondary injury cascade. This study still has certain limitations. The diagnostic efficacy of NORAD and its correlation with clinical prognosis need to be confirmed by further expanding the sample size. NORAD may participate in the pathological process of SCI by regulating other miRNAs or signaling pathways, and its complete regulatory network needs to be further explored. Future research will conduct long-term animal experiments to further explore NORAD and other possible target miRNAs and other downstream genes, and further improve the regulatory pathway of NORAD in SCI.
Conclusion
NORAD promoted the progression of SCI by sponging miR-22-3p and relieving its inhibitory effect on PTEN.
Footnotes
Ethical Considerations
The study was performed in line with the principles of the Declaration of Helsinki. Approval was granted by the Ethics Committee of Zhucheng People’s Hospital before the study began (No. 202106). This experiment was conducted with approval of the Animal Ethics Committee of Zhucheng People’s Hospital (No. 202114). All institutional and national guidelines for the care and use of laboratory animals were followed. Appropriate measures were taken to minimize the number and suffering of animals.
Consent to Participate
The written informed consent has been obtained from the participants involved.
Authors Contributions
Conceptualization, C.Z.,L.C., Q.W., Y.C., H.X.,Y.Z.; Data curation, Y.P., S.Y., Y.C., H.X.; Formal analysis, C.Z., Q.W., Y.C., H.X.; Funding acquisition, C.Z.; Investigation, Y.C.; Methodology, Y.P., S.Y.; Project administration, C.Z.,L.C.; Resources, Y.C.; Software, C.Z.,L.C.; Supervision, H.X.; Validation, Y.C.; Visualization, C.Z., Q.W.; Roles/Writing - original draft, C.Z., L.C.; Writing - review & editing, Y.Z.
Funding
The authors disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This work was funded by Wuhan Municipal Health Commission Research Fund Support (WX23Z29).
Declaration of Conflicting Interests
The authors declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Data Availability Statement
The datasets used and/or analysed during the current study are available from the corresponding author on reasonable request.