
Abstract
Endophytic fungi associated with the mangrove plant Ceriops tagal are a promising source of antibacterial compounds. This study aimed to isolate and identify endophytic fungi from C. tagal and evaluate the antibacterial activity of their secondary metabolites. Fungi were isolated from roots, branches and leaves using a direct plating method and cultured in potato dextrose broth. Crude extracts were prepared with ethyl acetate and tested against Escherichia coli, Staphylococcus aureus and Candida albicans using the disk diffusion method. Ten isolates were obtained, and all inhibited at least one test microorganism with inhibition zones ranging from 6.6 ± 0.55 mm to 19.1 ± 0.05 mm. Three isolates, ECT52, ECT57, and ECT63, showed strong activity with inhibition zones above 10 mm. Molecular identification based on ITS and LSU sequences identified them as Neopestalotiopsis saprophytica, Schizophyllum commune and Muyocopron laterale. Extracts of S. commune ECT57 showed the strongest activity against S. aureus with an MIC of 62.5 ± 0.96 µg/mL. LC-HRMS analysis of ECT57 extract revealed the presence of several dominant compounds, especially Cyclo(phenylalanyl-prolyl), 6-Quinolinecarboxylic acid and Caffeic acid. These findings indicate that C. tagal contains endophytic fungi that can synthesize antimicrobial compounds, which may have potential relevance for antibiotic development.
Keywords
Introduction
The rising incidence of antimicrobial resistance (AMR) has emerged as a critical global health issue, representing a substantial threat to public health worldwide. 1 AMR is defined as the capacity of microorganisms to survive exposure to antimicrobial agents, resulting in an increased frequency and severity of infections caused by pathogenic bacteria and fungi.2,3 Methicillin-resistant Staphylococcus aureus (MRSA) and third-generation cephalosporin-resistant Escherichia coli are two of the most common killer combinations; in 2019, bacterial AMR was expected to be responsible for 1.27 million deaths worldwide and linked with 4.95 million.4 In addition, among fungal pathogens, Candida albicans has been designated as a critical priority organism for the development of novel antifungal therapies. 5 These alarming trends have intensified the demand for new and effective strategies, particularly those based on natural products, to address infections caused by drug-resistant microorganisms. Consequently, the search for novel antimicrobial agents has increasingly targeted natural resources. 6 Nevertheless, the excessive use of plant-derived antimicrobial compounds may pose risks to plant conservation and biodiversity. 7 As a sustainable alternative, endophytic fungi associated with host plants have gained considerable attention as potential sources of antimicrobial agents. 8 Endophytic fungi are capable of producing secondary metabolites that are identical or structurally related to those of their host plants and offer several advantages, including rapid growth and suitability for large-scale production of bioactive compounds.9,10
Endophytic fungi inhabiting mangrove plants are regarded as a promising alternative source of antimicrobial compounds with potential for antibiotic development, due to their distinctive ecological traits and the extensive diversity of secondary metabolites they produce. 11 The mangrove plant Ceriops tagal (generally known as the spurred mangrove or “tengar”) is a mangrove species belonging to the family Rhizophoraceae. 12 C. tagal (Perr.) C.B. Rob is a hardy mangrove species with great salinity tolerance, and it is extensively found in intertidal coastal zones of tropical and subtropical climates, particularly in Africa, South Asia, the South Pacific, and southern China. 13 C. tagal has a wide spectrum of secondary metabolites, including alkaloids, flavonoids, polyphenols, cardiac glycosides, saponins, and steroids.14,15 Several of these substances were identified to have potential antibacterial action. 16
Several studies have reported antimicrobial potential from endophytic fungi isolated from C. tagal. An endophytic fungus, Hypoxylon mangrovei, isolated from C. tagal collected at Ngurah Rai Grand Forest Park, Bali, demonstrated antibacterial activity against S. epidermidis. 17 Endophytic Aspergillus fumigatus isolated from the Sundarbans mangrove ecosystem exhibited antimicrobial activity. 18 Antibacterial assays of talarocyclopenta B, a phenolic ether derivative produced by Talaromyces assiutensis JTY2 associated with C. tagal leaves from southern China, revealed potent antibacterial activity against several pathogenic bacteria. 19 In addition, Aspergillus spp. isolated from C. tagal have been reported to possess antibiotic activity against various microorganisms. 20 Furthermore, the metabolite citrinin is known to exhibit strong antibacterial activity, 21 while seiricardine D produced by Cytospora sp. has demonstrated antifungal activity against Magnaporthe oryzae. 12
The urgent need to create new and efficient natural-based solutions to fight drug-resistant pathogenic microbial has increased due to the rapid rise in AMR. Therefore, it is crucial to identify new antimicrobial medications. Therefore, the goal of this work was to separate endophytic fungi from the West Sumatra, Indonesian mangrove plant C. tagal and determine their ability to produce antimicrobial compounds effective against pathogenic microbes.
Materials and Methods
Identification of Sample Material
Plant material of C. tagal was collected from the mangrove ecosystem of Batang Tomak, Sungai Beremas District, West Pasaman Regency, West Sumatra, Indonesia, at the geographical coordinates 0°14′00.0″N and 99°17′58.5″E. The plant samples were characterized and confirmed at the Herbarium of Andalas University (ANDA) under voucher specimen number 299/K-ID/ANDA/IV/2024. Endophytic fungi were isolated from healthy, infection-free plant parts such as roots, branches, and leaves (Figure 1). The leaves used came from the third position from the branch apex. In contrast, branches were selected from branches bearing healthy leaves, and relatively young roots were collected. 20 All collected samples were transported to the laboratory for additional testing in a cooler box kept at 4°C and processed within 8 hours of collection. 22
The Pictures of Ceriops tagal Leaves (A) and Roots (B).

Sample Surface Sterilization and Isolation of Endophytic Fungi
Plant samples consisting of leaves, branches, and roots were surface-sterilized and directly inoculated onto growth media following a previously described method. 20 The samples were first cleaned with distilled water for 5 minutes to remove dirt particles and surface-associated bacteria. Surface sterilization was performed by immersing the samples in 70% ethanol for 2 minutes before treating them with sodium hypochlorite for 1 minute. The samples were then cleaned three times with sterile distilled water for 2 minutes each before drying on sterile filter paper. Sterilized tissues were aseptically sliced into 2 cm segments with a sterile scalpel and placed on potato dextrose agar (PDA) medium with 50 mg/L chloramphenicol to inhibit bacterial growth. The inoculation plates were incubated for 3 to 7 days at 27°C–30°C. 20 Endophytic fungi from plant explants were isolated, purified, and maintained by successive subcultures. The efficiency of the surface sterilization technique was tested using the final rinse water as a control. An aliquot (0.1 mL) of the last sterile distilled water washing was plated onto PDA; the absence of microbial growth indicated effective surface sterilization. 23
Cultivation and Extraction
Endophytic fungi were kept static in 500 mL of liquid potato dextrose broth (PDB) at room temperature (20°C–25°C) for 7 days. 24 Each culture was macerated with ethyl acetate (EtOAc) at a ratio of 1:1 (v/v). 25 The resultant extracts were concentrated using a rotary evaporator to obtain crude extracts, which were then used for antibacterial activity tests. 20
Antimicrobial Activity Screening
Screening for antimicrobial activity was performed using the Kirby–Bauer disk diffusion method 26 against the test microorganisms, including the Gram-negative bacterium E. coli American Type Culture Collection (ATCC) 25922, the Gram-positive bacterium S. aureus ATCC 25923, and the yeast C. albicans ATCC 10231. Inoculum preparation and incubation conditions were aligned with basic Clinical and Laboratory Standards Institute (CLSI) procedural recommendations. Microbial suspensions were adjusted to 0.5 McFarland turbidity and inoculated onto nutrient agar (for bacteria) or Sabouraud dextrose agar (SDA) plates (for yeast). Sterile paper disks (6 mm diameter) were impregnated with 10 µL of a 5% (w/v) solution of endophytic fungal extract in DMSO, resulting in about 0.5 mg of dried extract per disk, and placed on the inoculated agar surfaces. 25 Positive controls for antibacterial and antifungal efficacy were chloramphenicol (Oxoid→; 30 µg/disk) and nystatin (Oxoid→; 100 IU/disk), whereas negative controls were DMSO-containing disks. The plates were incubated at 35°C for yeast and 37°C for bacteria for 24–48 h. Following incubation, the inhibition zones were inspected and quantified to determine the diameter of microbial growth inhibition. 27 For each fungal extract producing an inhibition zone larger than 10 mm, the mean inhibition diameter and standard deviation were calculated. Extracts exhibiting the strongest activity were further evaluated by determining the minimum inhibitory concentration (MIC) using a broth microdilution assay conducted following CLSI methodological principles for antimicrobial susceptibility testing. Two-fold serial dilutions were prepared in appropriate broth medium in 96-well microplates to obtain final concentrations ranging from 1.95 to 1,000 µg/mL. Chloramphenicol (for bacteria) and nystatin (for yeast) were included as positive controls, while broth containing 0.5% DMSO served as the negative control. All assays were performed in triplicate. The MIC was defined as the lowest concentration that completely inhibited visible microbial growth. 17
Preliminary Phytochemical Testing of Extract from Endophytic Fungi
To discover bioactive compound classes, secondary metabolite and phytochemical analyses were performed on EtOAc extracts of an endophytic fungi exhibiting antimicrobial activity and inhibitory zones greater than 10 mm. The screening aimed to identify the presence of terpenoids and alkaloids, phenolic compounds, and steroids using standard qualitative methods.27,28 Terpenoids were tested using the Liebermann-Burchard reagent, with a positive result indicated by a color change of the extract to green, blue, or purple. Dragendorff’s reagent was used to identify alkaloids, which resulted in an orange tint. Phenolic compounds were detected using specialized reagents, such as ferric chloride, and a characteristic color shift indicated the presence of phenolic ingredients.27,28
Characterization and Identification of Endophytic Fungi
Endophytic fungi with antimicrobial activity were observed under a light microscope at 100× magnification. Microscopic examination was performed using the slide culture technique with the addition of one drop of lactophenol cotton blue stain. Microscopic characteristics were evaluated based on hyphal structures, spore types, spore morphology, and other distinctive features. The observed characteristics were compared with those described in fungal identification references. 29
Molecular Identification
Molecular techniques were used to identify endophytic fungal isolates with the highest antibacterial activity at the species level. Molecular characterization was performed at the Research Center for Applied Microbiology, National Research and Innovation Agency (BRIN), Indonesia, while PCR products were then delivered to Genetika Science Indonesia for DNA sequencing. Genomic DNA was isolated using the Quick-DNA™ Fungal/Bacterial Miniprep Kit (Zymo Research, D6005) as per the manufacturer’s instructions. Amplification of the internal transcribed spacer (ITS) region was performed by polymerase chain reaction (PCR) using (2×) MyTaq™ HS Red Mix (Bioline, BIO-25048) and the universal primers ITS1 (5′-TCCGTAGGTGAACCTGCGG-3′) and ITS4 (5′-TCCTCCGCTTATTGATATGC-3′). 30 For isolates that could not be identified through ITS analysis, the D1/D2 domain of the large subunit (LSU) was examined using primers NL1 (5′-GCATATCAATAAGCGGAGGAAAAG-3′) and NL4 (5′-GGTCCGTGTTTCAAGACGG-3′). The resultant nucleotide sequences were matched to reference sequences from the GenBank database using the NCBI’s Basic Local Alignment Search Tool (BLAST). MEGA version 11 was used to perform multiple sequence alignments, with the CLUSTAL W algorithm. Phylogenetic relationships were determined using the Neighbor-Joining method and 1000 bootstrap repetitions. Bootstrap support values are expressed as percentages and indicated adjacent to the corresponding nodes. The phylogenetic tree was constructed to scale, with branch lengths reflecting the number of nucleotide substitutions per site and representing the evolutionary distances applied in the analysis. 31
Phytochemical Profiling Using LC-HRMS
The crude extract was characterized using UHPLC (Thermo Scientific) with Binary Pump (Germering, Germany) equipped with Orbitrap™ Exploris 240 HRMS (Bremen, Germany) detector, Optamax™ NG Heated Electrospray Ionization (H-ESI) ion source and Thermo Scientific™ Accucore™ Phenyl Hexyl column (100 mm length, 2.1 mm ID, 2.6 μm particle size). The separation parameters were as follows: mobile phase water with 0.1% formic acid (mobile phase A) and acetonitrile with 0.1% formic acid (mobile phase B); gradient elution with 5% B and increased gradually to 90% in 16 min, hold at 90% for 4 min and back to 5% B until 25 min; flow rate 0.3 mL/min; column temperature 40°C; injection volume 5 μL. Chemical profiling was implemented using Thermo Scientific Compound Discoverer 3.3 (San Jose, USA).
Statistical Analysis
Antimicrobial activity was measured by measuring the inhibition zone diameter (mm) and expressed as the mean ± SD of three independent replicates. Differences between treatments were examined using one-way ANOVA, followed by Dunnett’s post hoc test to compare each treatment to the positive control. A p value of <.05 was judged statistically significant.
Result
Isolation of Endophytic Fungi from Leaves, Branches, and Roots of C. tagal
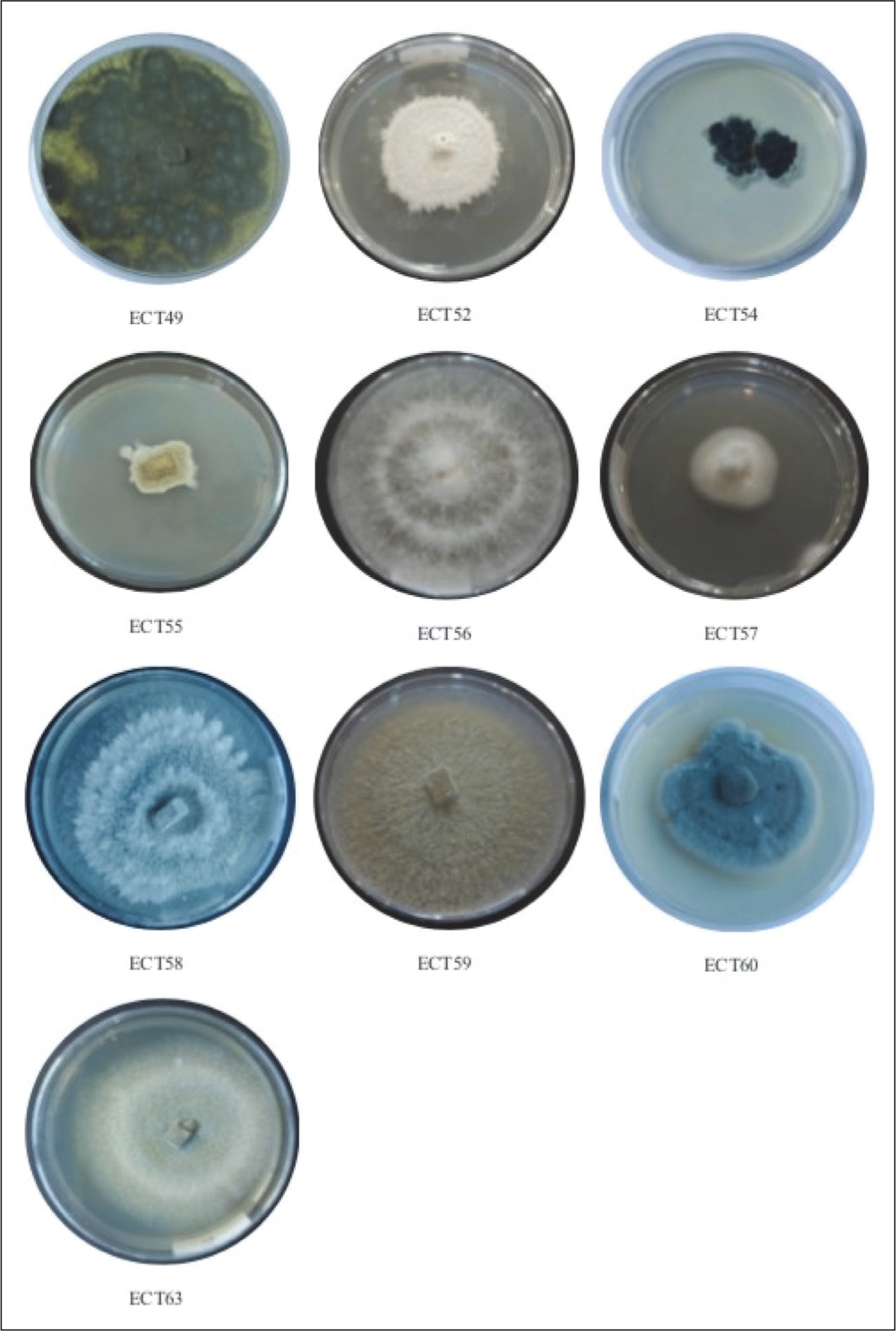
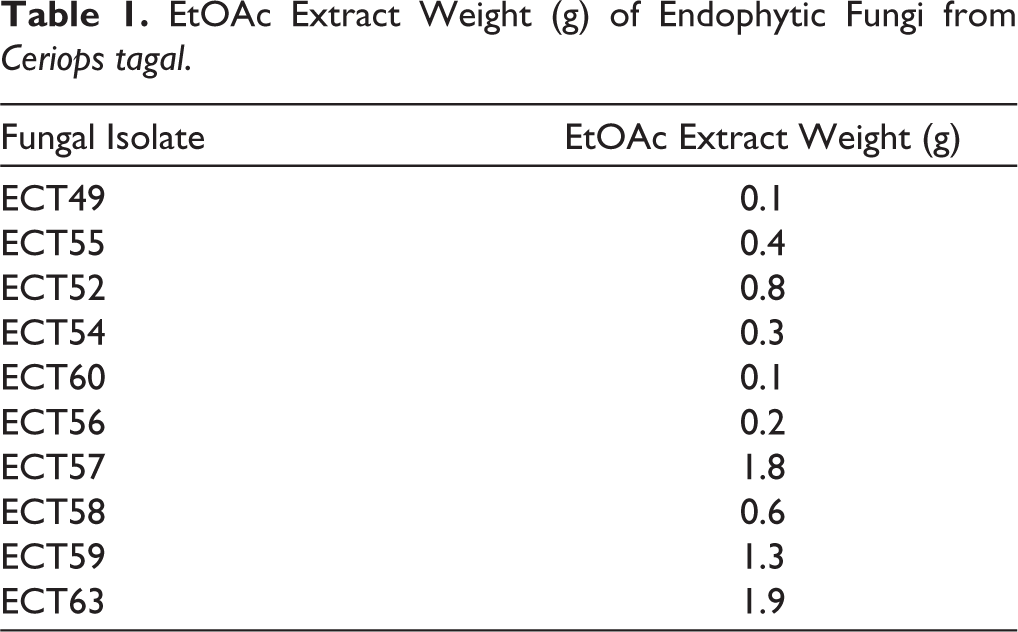
Ten endophytic fungal isolates were successfully isolated from the roots, leaves, and branches of C. tagal. All isolates were identified with the code ECT. Two isolates were obtained from the roots and coded as ECT49 and ECT55, three isolates were obtained from the leaves and coded as ECT52, ECT54, and ECT60, and five isolates were obtained from the branches and coded as ECT56, ECT57, ECT58, ECT59, and ECT63 (Figure 2). The fungal isolates were grown in liquid PDB and incubated at 20°C–25°C for 7 days. Secondary metabolites were extracted with EtOAc, and the resulting extracts were concentrated under reduced pressure to obtain crude extracts. The EtOAc extract was tested for antimicrobial activity. The weight of EtOAc extract ranges from 0.1 to 1.9 g (Table 1).
Endophytic Fungi Isolated from Ceriops tagal were Cultured on PDA.
EtOAc Extract Weight (g) of Endophytic Fungi from Ceriops tagal.
Antimicrobial Activity
The antimicrobial activity assay demonstrated that endophytic fungi isolates from C. tagal could inhibit the growth of E. coli, S. aureus, and C. albicans. The findings revealed disparities in inhibitory zone widths across the isolates, indicating differences in their ability to manufacture antimicrobial chemicals. The antimicrobial activity of EtOAc extracts of individual fungal isolates was tested. The results of antimicrobial activity screening of the fungus extract with a concentration 5% are presented in Table 2. The differential responses observed among the three test microorganisms reflect the antimicrobial spectrum of the isolates against E. coli, S. aureus, and C. albicans. These findings indicate that secondary metabolites produced by endophytic fungi associated with C. tagal have antimicrobial activity. The differences in inhibition zone diameters observed in the disk diffusion assay further indicate differences in the capacity of the isolates to produce and secrete secondary metabolites with antimicrobial activity.
Antimicrobial Activity of Endophytic Fungal Extract from Ceriops tagal.
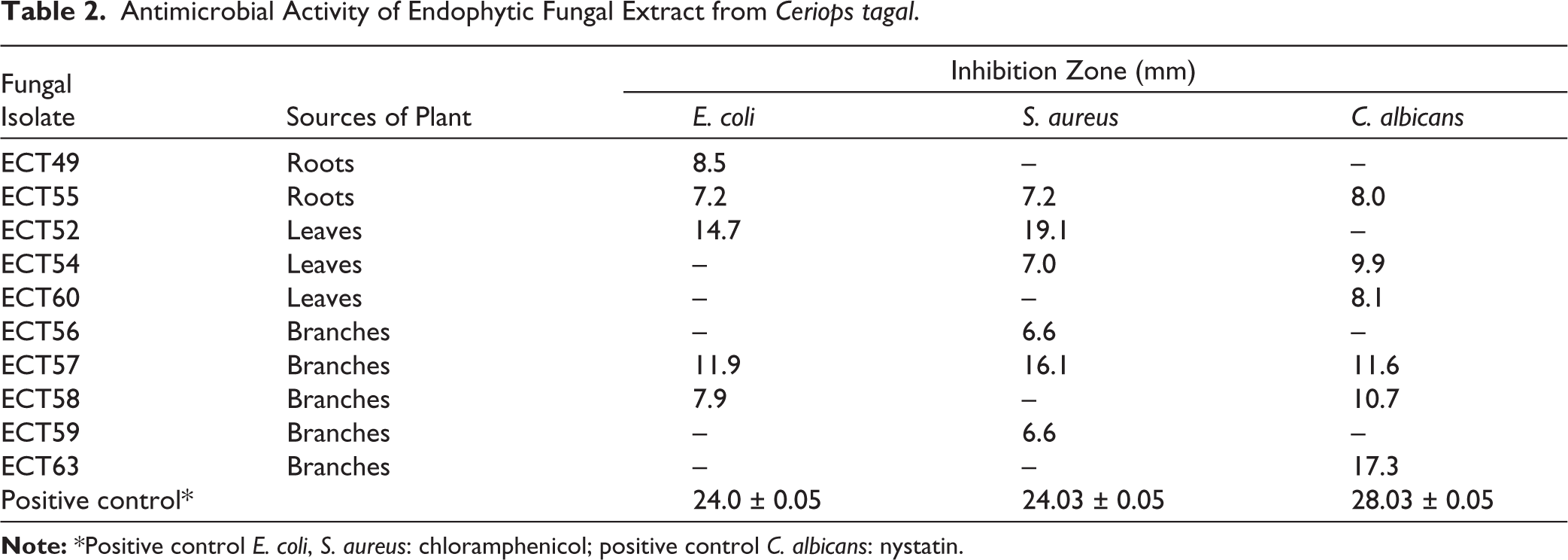
At a concentration of 5%, three fungal isolates showed significant inhibition against the test microorganisms, with inhibition zone diameters exceeding 10 mm (Table 3). Isolate ECT52 showed higher antibacterial activity against S. aureus with an inhibition zone diameter of 19.06 ± 0.15 mm, and activity against E. coli of 14.76 ± 0.30 mm. Isolate ECT57 showed significant activity against S. aureus (16.06 ± 0.15 mm) and C. albicans (11.8 ± 0.20 mm), while ECT63 stood out against C. albicans with a diameter of 17.83 ± 0.05 mm. Isolates ECT52 and ECT57 showed a greater ability to inhibit the growth of Gram-positive bacteria (S. aureus) compared to Gram-negative bacteria (E. coli). This can be explained by differences in cell wall structure, where Gram-positive bacteria have simpler cell walls than Gram-negative bacteria, so they are more susceptible to antimicrobial compounds.
Antimicrobial Activity of Potential Endophytic Fungi.

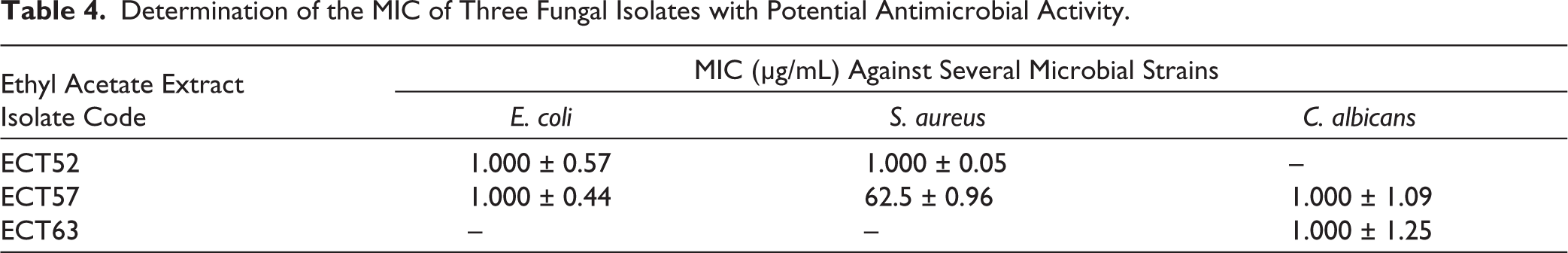
Data analysis with mean ± standard deviation showed that several isolates had significant differences compared to the positive control. Isolates ECT52 and ECT57 showed statistically significant growth suppression of S. aureus, while ECT63 had primary activity against C. albicans. These results indicate that endophytic fungi from C. tagal have the potential to be a source of secondary metabolites with antimicrobial activity. To determine the MIC of each bacterial and fungal strain, MIC testing was carried out (Table 4). The results showed that isolates ECT52, ECT54, and ECT63 had potential antimicrobial activity against the test microbes, with MIC values ranging from 62.5 ± 0.96 μg/mL to 1,000 ± 1.25 μg/mL. Isolate ECT57 showed the lowest MIC against S. aureus, indicating stronger antimicrobial activity among the other isolates.
Determination of the MIC of Three Fungal Isolates with Potential Antimicrobial Activity.
Phytochemical Analysis
Extracts from isolates ECT52, ECT57, and ECT63 were analyzed for phytochemicals to evaluate the presence of bioactive compounds, particularly those with antibacterial activity. The results revealed that phenolic compounds, terpenoids, and flavonoids were present in the extracts of ECT52 and ECT57, while alkaloids were detected only in the extract of ECT57. Flavonoids were also identified in the extract of ECT63, as shown in Table 5.
Phytochemical Screening of EtOAc Extract from Ceriops tagal.

Characterization and Identification of Endophytic Fungi
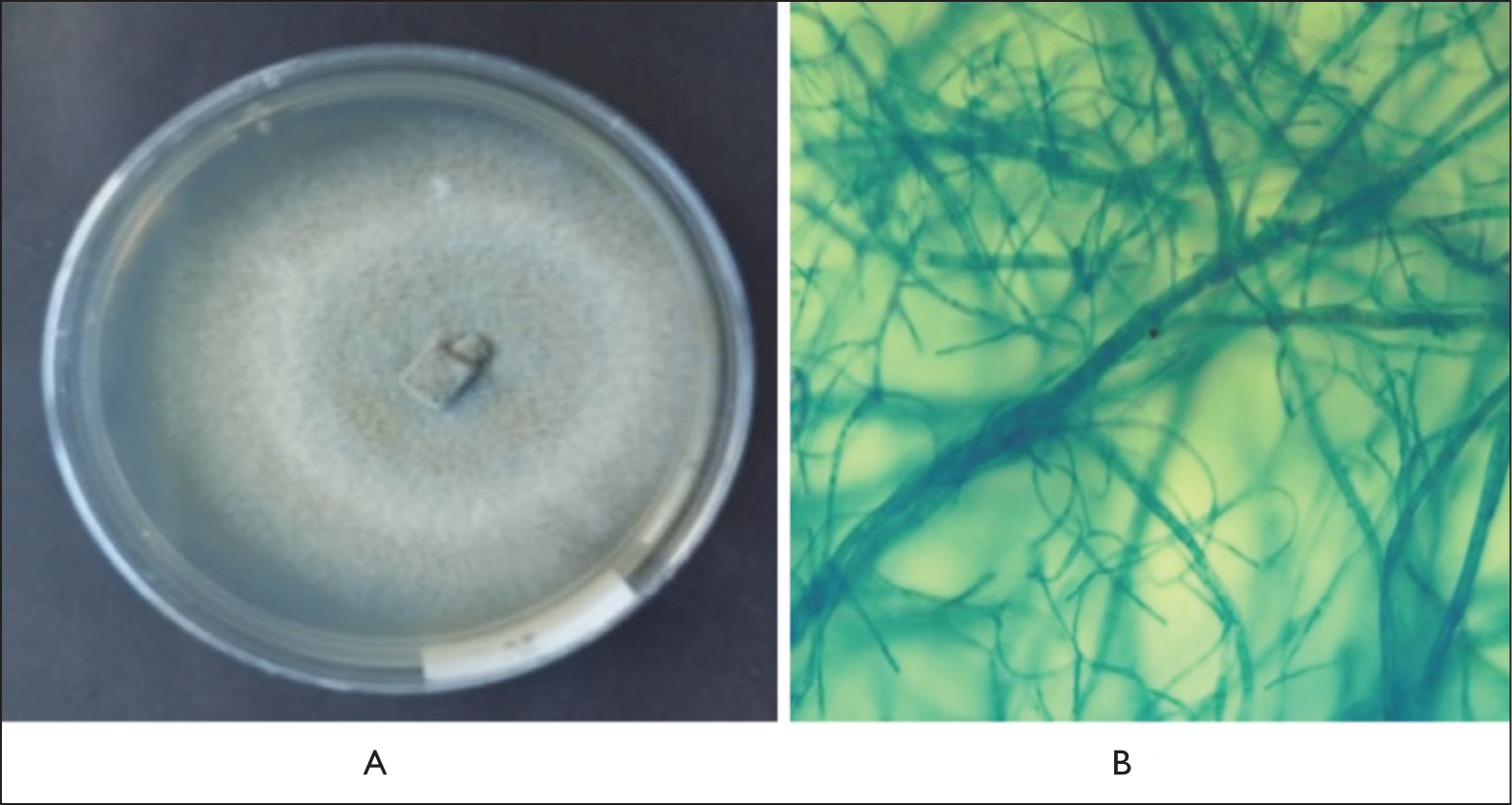
Isolate ECT52 exhibited colonies with a rough surface. The obverse side of the colony was grayish white, while the reverse side appeared yellowish white. The colonies were irregular in shape, with a convex elevation and filamentous margins. Microscopically, the conidia were ellipsoidal with four transverse septa (five cells), consisting of two hyaline terminal cells and three brown-pigmented median cells (Figure 3). Isolate ECT57 displayed colonies with a smooth, cottony surface. Both the obverse and reverse sides of the colonies were white. The colonies were regular in shape, with a flat elevation and filamentous margins. Microscopically, septate and branched hyphae were observed, along with the presence of clamp connections at the hyphal septa (Figure 4). Isolate ECT63 exhibited colonies with a smooth, velvety surface. The obverse side of the colony was gray with white margins, while the reverse side was black with yellow margins. The colonies were regular in shape, with a raised central elevation and filamentous margins. Microscopically, the isolate showed brown-pigmented, septate hyphae with irregular branching (Figure 5).
Macroscopic (A) and Microscopic (B) Pictures of Fungal Isolate ECT52 Showing Ellipsoid-shaped Conidia with Four Septa, Consisting of Two Hyaline Terminal Cells and Three Brown-pigmented Median Cells.

Macroscopic (A) and Microscopic (B) Pictures of Fungal Isolate ECT57 Showing Branched, Septate Hyphae with the Presence of Clamp Connections at the Hyphal Septa.
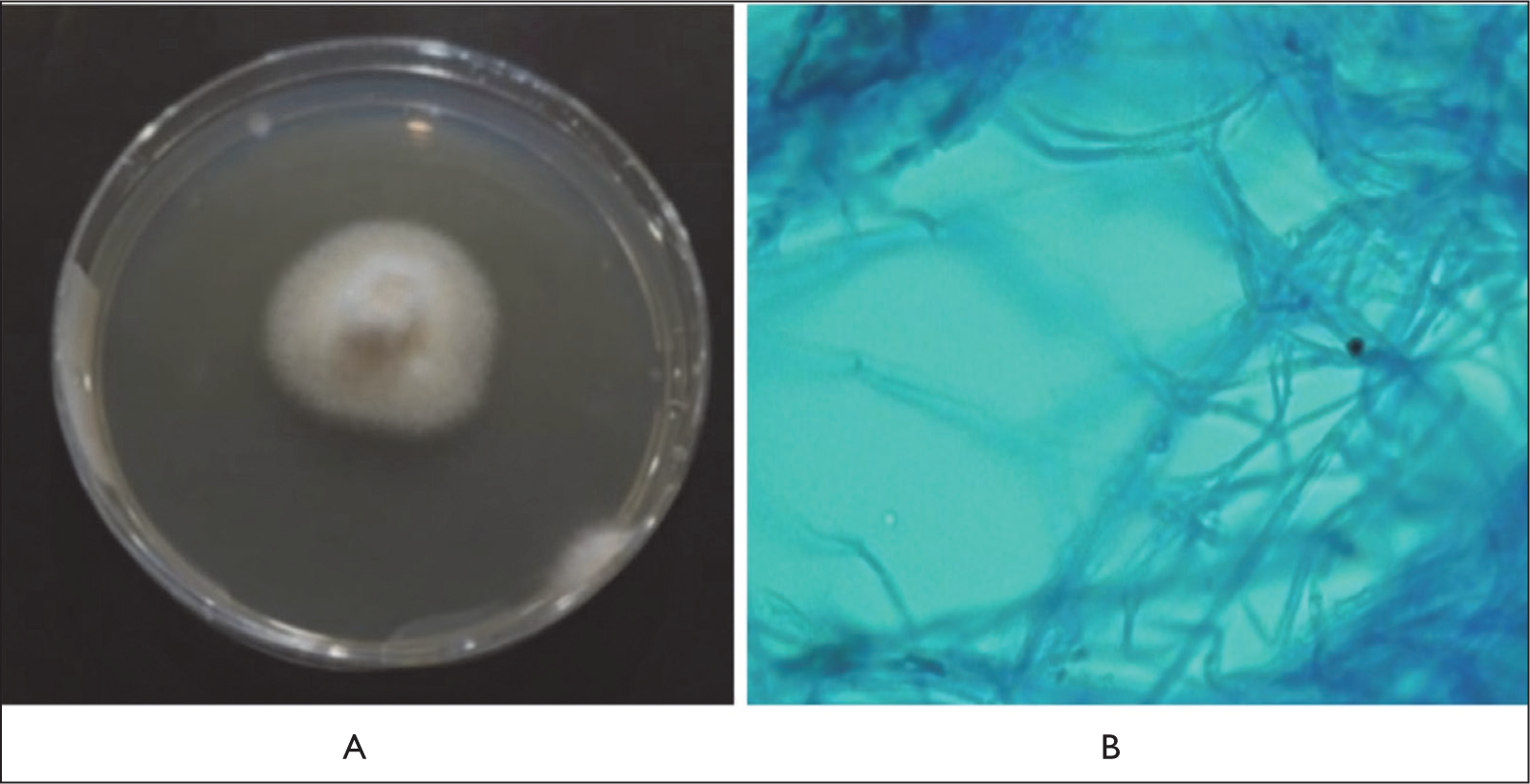
Macroscopic (A) and Microscopic (B) Pictures of Fungal Isolate ECT63 Showing Brown-pigmented, Septate Hyphae with Irregular Branching.
Molecular Identification
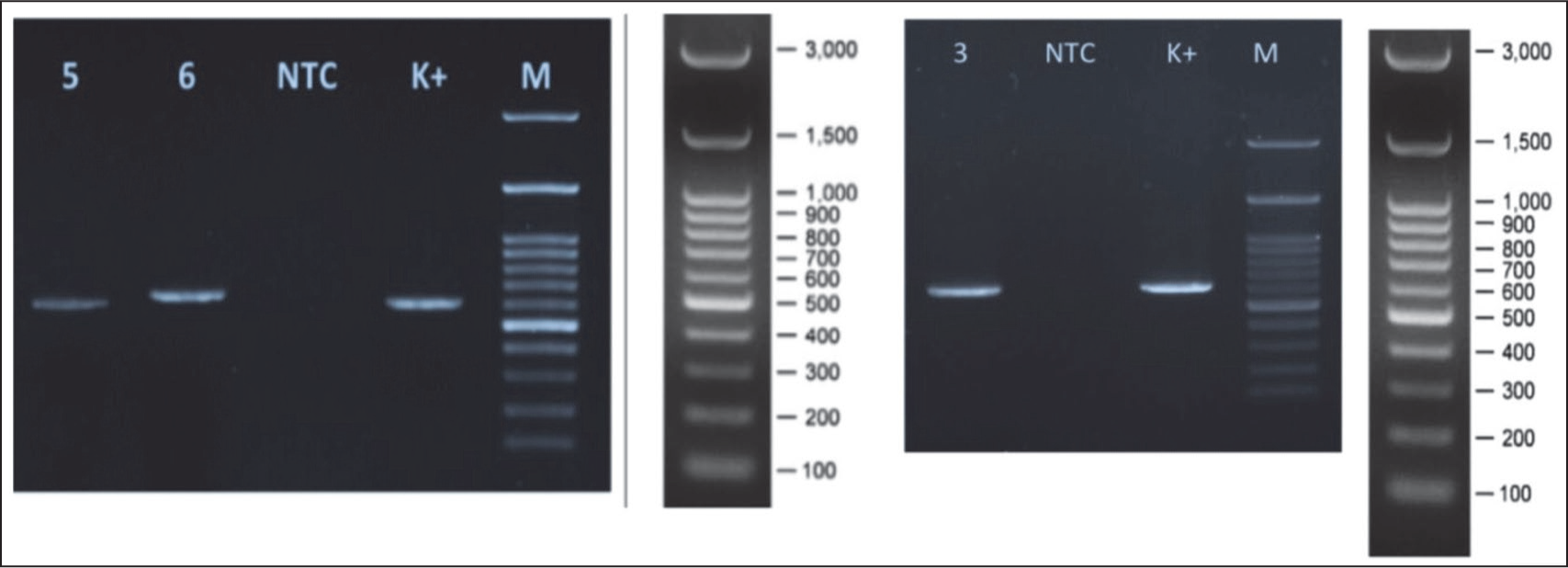
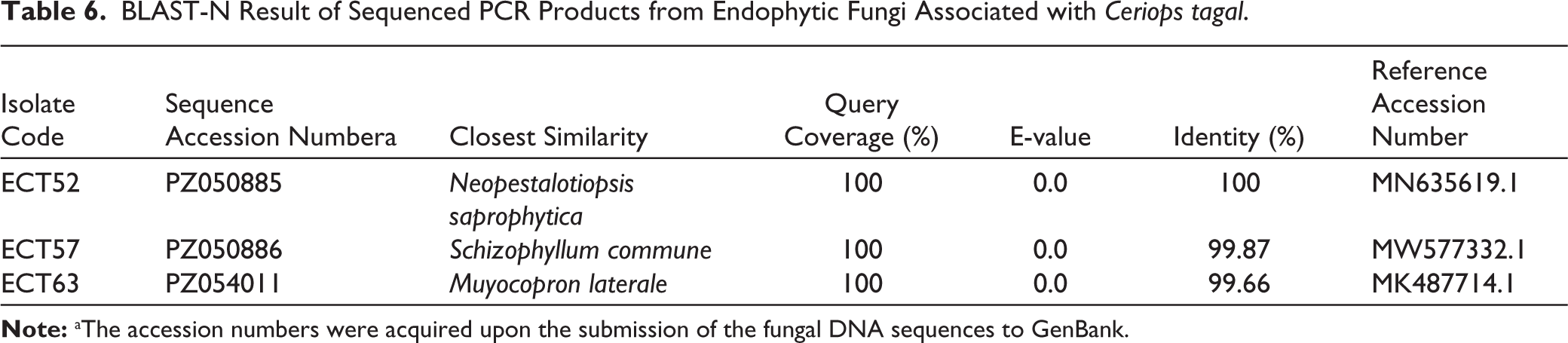
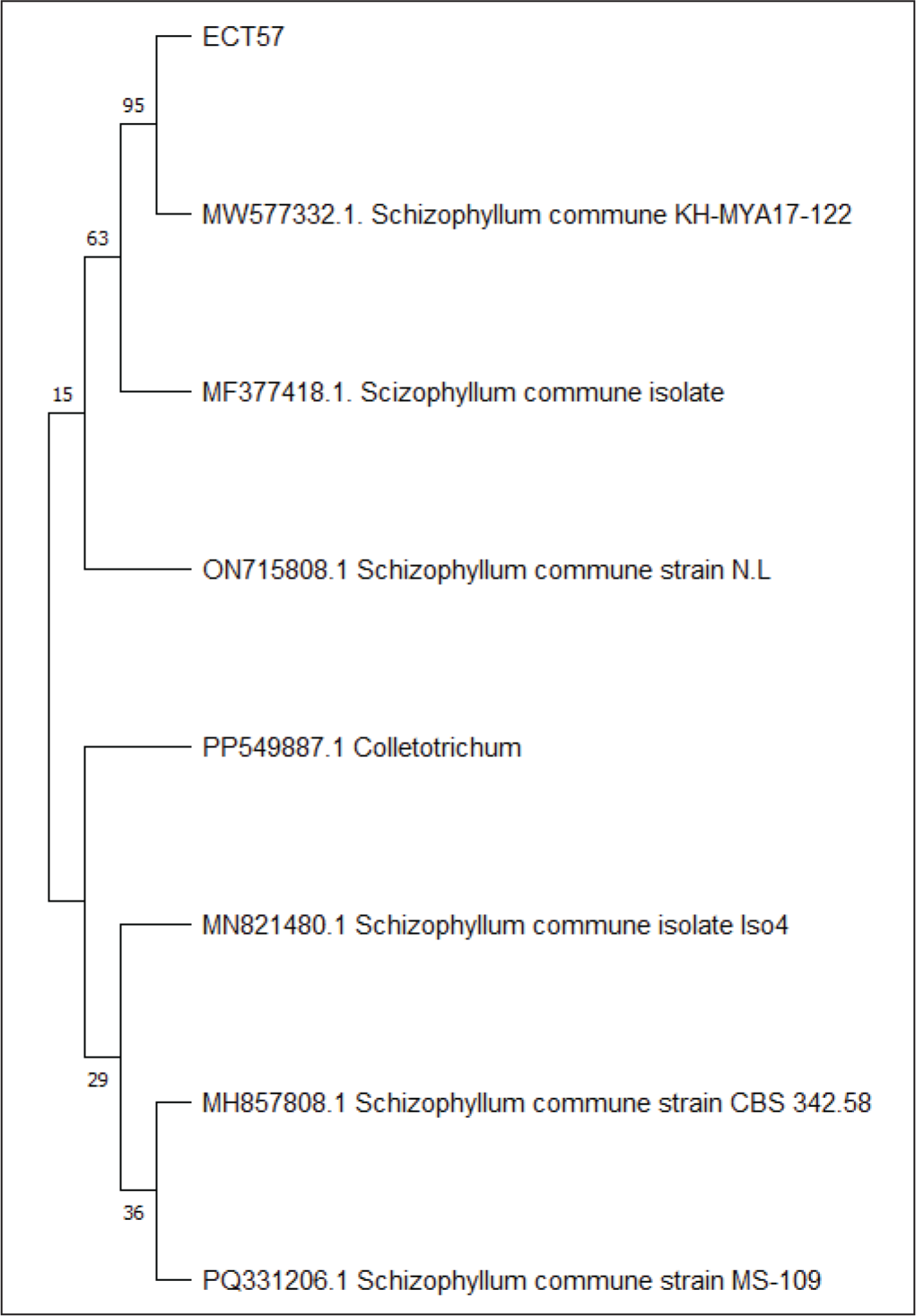
Endophytic fungi species were identified by comparing ITS1 and ITS4 rDNA sequences. Since no amplified DNA bands were found in gel electrophoresis after PCR in isolate ECT63 ITS region, the fungal isolate was further identified by studying the D1/D2 domains of the LSU region. Amplification of the ITS region resulted in discrete DNA bands of around 500–600 bp, as seen in agarose gel electrophoresis (Figure 6). To predict the taxon of each endophytic fungus, the acquired sequences were compared with fungal sequences from the GenBank database using the NCBI BLAST nucleotide algorithm. The results demonstrated sequence homology with three different species, with similarity percentages (<99%) and E values (Table 6). Furthermore, the creation of a phylogenetic tree revealed that each isolate lineage was divided into three major clades with strong bootstrap support, as shown in Figures 7–9.
Electropherogram of Amplified ITS rDNA from (5) ECT52; (6) ECT57; and Amplified LSU Region of (3) ECT63 (NTC) No Template Control; (K+) Positive Amplification Control; (M) 100 bp DNA Ladder.
BLAST-N Result of Sequenced PCR Products from Endophytic Fungi Associated with Ceriops tagal.
The Phylogenetic Tree Inferred Using the Neighbor-joining Method of ITS Sequence of Fungi ECT52 Derived from Ceriops tagal.

The Phylogenetic Tree Inferred Using the Neighbor-joining Method of ITS Sequence of Fungi ECT57 Derived from Ceriops tagal.
The Phylogenetic Tree Inferred Using the Neighbor-joining Method Region of LSU Sequence of Fungi ECT63 Derived from Ceriops tagal.
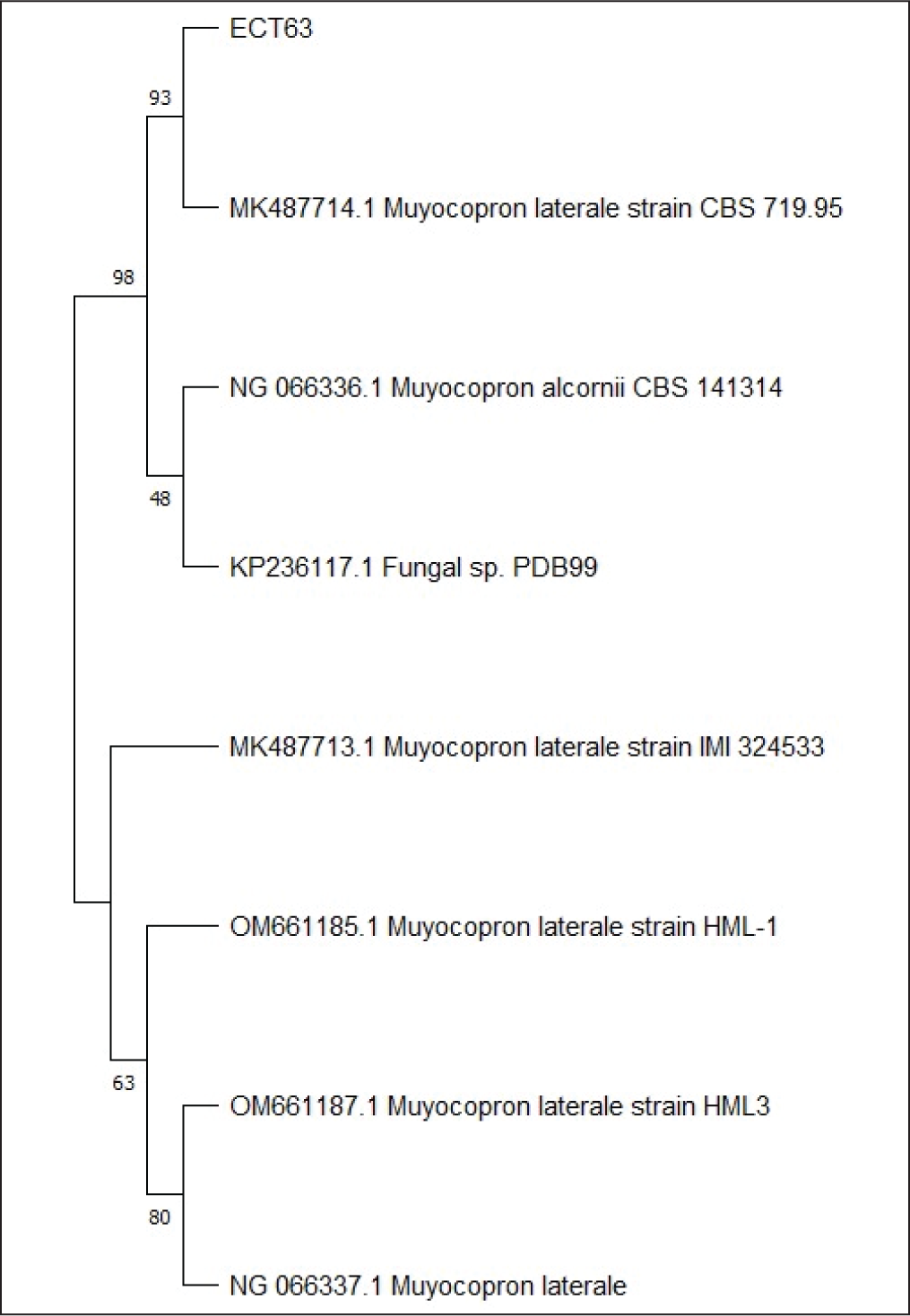
The sequence alignment study revealed that isolate ECT52 belonged to the species Neopestalotiopsis saprophytica, with 100% similarity to this species (MN635619.1), which was consistent with its phylogenetic analysis, which had a high bootstrap support of 100%. Isolate ECT57 was 99.87% similar to Schizophyllum commune (MW577332.1), with 95% bootstrap support in the phylogenetic tree. The bootstrap value for isolate ECT63 (Muyocopron laterale, MK487714.1) has been corrected from 95% to 93%. Please update the manuscript accordingly.
LC-HRMS Analysis
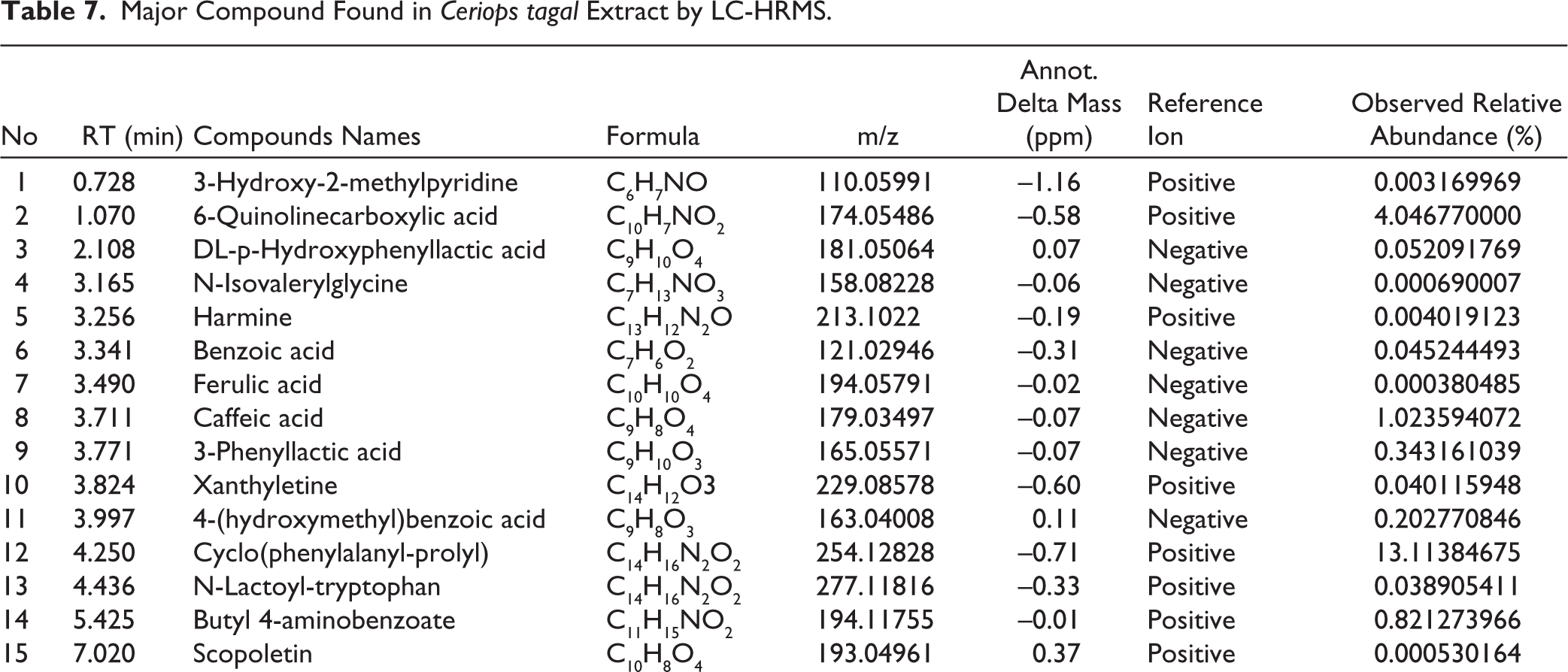
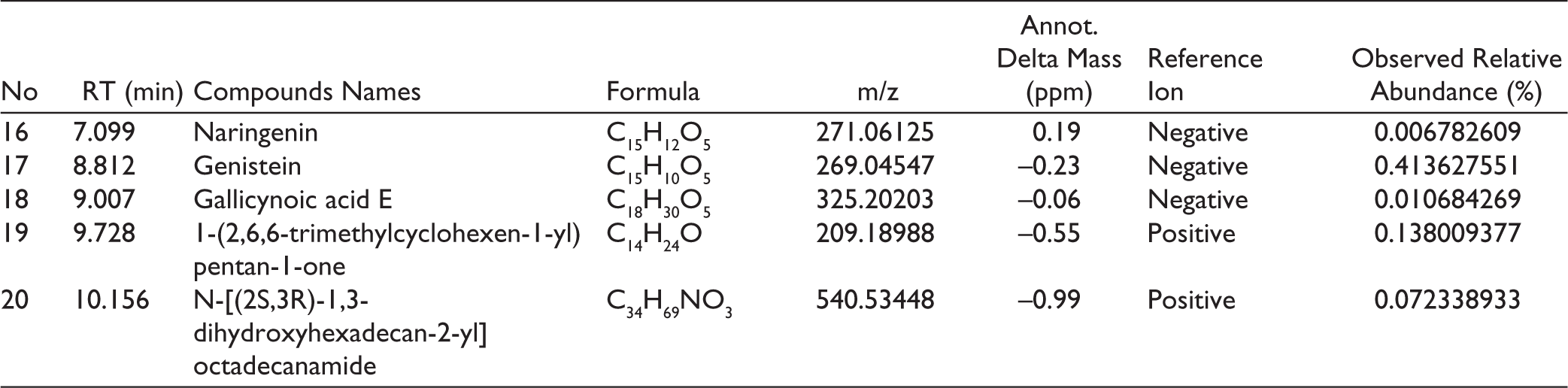
A complex metabolite composition was revealed by LC-HRMS profiling of S. commune EtOAc extract. Of the 433 compounds found, 20 were found to be important contributors based on relative abundance and annotation confidence level. High-resolution mass data with mass precision in the range of ±5 ppm were used for compound identification, enabling precise molecular formula prediction (Table 7). The three most prevalent chemicals in the S. commune extract are Cyclo(phenylalanyl-prolyl), which has an abundance of roughly 13.1%, and C₁₄H₁₆N₂O₂, which has a m/z of 254.1283 [M+H]+. The second, with m/z 174.05486 [M+H]+; C₁₀H₇NO₂, was identified as 6-Quinolinecarboxylic acid with an abundance of around 4.04%, while the third, with m/z 179.03497 [M+H]-; C₉H₈O₄, was identified as Caffeic acid with an abundance of over 1.02%. The chromatogram of the EtOAc extract of S. commune is depicted in Figure 10.
Major Compound Found in Ceriops tagal Extract by LC-HRMS.
Chromatogram of the Endophytic Fungi S. commune Isolated from Ceriops tagal.
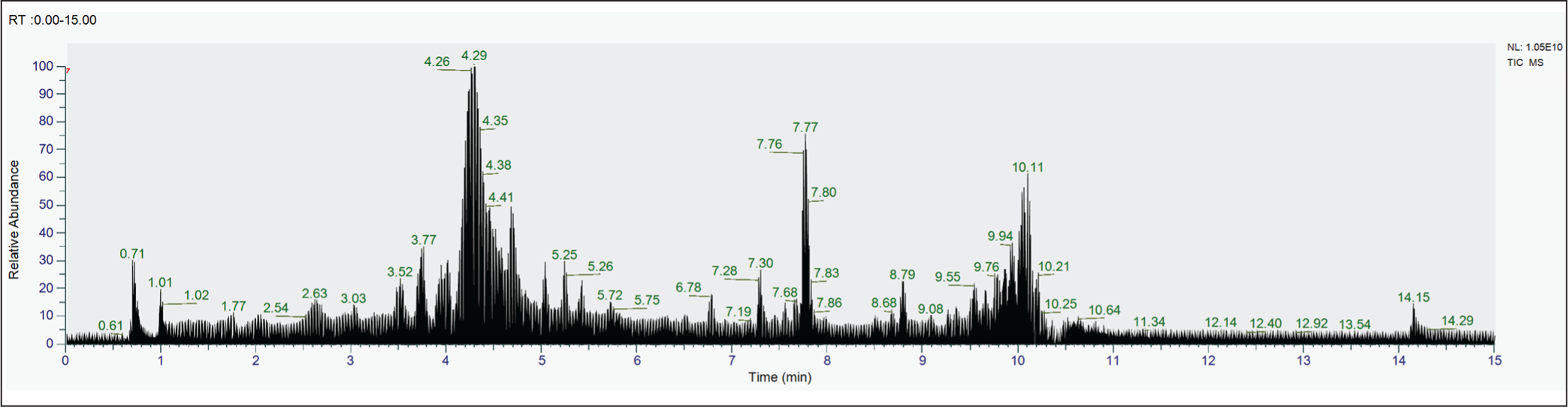
The LC-HRMS profiling of S. commune EtOAc extract revealed a complex mixture of compounds, with Cyclo(phenylalanyl-prolyl), 6-Quinolinecarboxylic acid, and Caffeic acid being the most abundant. These major compounds are likely key contributors to the observed antimicrobial activity, as they have known bioactive properties. Minor compounds, although less abundant, may also enhance the effect through synergistic interactions. This indicates that the antimicrobial activity of the extract results from both the dominant bioactive constituents and the combined action of minor metabolites.
Discussion
Endophytic fungi associated with mangrove plants have developed distinct metabolic adaptations that allow them to persist under harsh environmental conditions, including elevated salinity and temperature, tidal dynamics, oxygen limitation, high moisture levels, and restricted light exposure and air circulation. These adaptive traits enable the fungi to effectively compete for nutrients and ecological niches within the highly competitive mangrove habitat. Such environmental pressures are known to cause the formation of structurally varied secondary metabolites with distinct biological roles and activities. 32 Previous research has shown that mangrove-derived endophytic fungi can produce a wide range of secondary metabolites, including aliphatic chemicals, alkaloids, flavonoids, glycosides, lactones, steroids, and terpenoids. 33 Consequently, mangrove endophytic fungi are regarded as an important source of pharmacologically active compounds, as mangrove ecosystems are recognized for their chemical uniqueness and high metabolite diversity. 34 Several studies have proven the significant antibacterial activity of endophytic fungi isolated from mangrove settings. 35
Recent advances in molecular genetics have significantly enhanced the use of DNA-based techniques in fungal taxonomy. In this study, species-level identification was achieved through molecular characterization based on amplification of the ITS region, which is widely accepted as a reliable barcode for fungal identification. The ITS region is located between the conserved 18S and 28S ribosomal RNA genes33,36 and displays substantial sequence variability among fungal taxa, making it a valuable marker for phylogenetic and taxonomic analyses of filamentous fungi. 37 Phylogenetic reconstruction based on ITS sequence data showed that the isolates were grouped into two main clades corresponding to the genera Neopestalotiopsis and Schizophyllum, while the isolate identified as Muyocopron was classified using analysis of the D1/D2 domain of the LSU region.
All of the taxa discovered in this study had previously been described as endophytic fungi. The results showed that isolate ECT52 (N. saprophytica), isolated from C. tagal leaf tissues, has broad-spectrum antibacterial activity, preventing the growth of E. coli and S. aureus. These findings are consistent with previous publications demonstrating that endophytic Neopestalotiopsis spp. isolated from the fruits of the mangrove Avicennia marina have antibacterial activity. 38 Neopestalotiopsis species have been shown to produce a diverse spectrum of bioactive compounds, including eugenol, which has potent antibacterial activity against pathogenic bacteria. 39 Compounds from Neopestalotiopsis sp. HN-1-6, found in Beibu Gulf marine mud, showed antibacterial action against S. aureus and E. coli. Compounds 3 and 13 had the strongest effects (MIC 64 μg/mL). 40 Furthermore, neopupestalotin B, a metabolite produced by endophytic Neopestalotiopsis sp., has been reported to suppress the growth of the Gram-positive bacteria Bacillus subtilis. 41
The endophytic fungal isolate ECT57 (S. commune), obtained from the branches of C. tagal, was the most promising in this study due to its broad-spectrum antimicrobial activity. This isolate was able to prevent the development of the Gram-negative bacteria E. coli, the Gram-positive bacteria S. aureus, and the pathogenic yeast C. albicans. The occurrence of S. commune in branch tissues is consistent with previous reports describing its ability to exist as an endophyte in woody plants, although it is infrequently reported because this species is more commonly recognized as a saprophytic fungus. 42
Its presence within host tissues highlights the ecological flexibility and opportunistic life strategy of S. commune, allowing transitions between latent endophytic and saprophytic phases depending on environmental conditions and host physiological status. As an efficient wood decomposer, S. commune possesses a highly effective lignocellulolytic enzyme system that enables the utilization of plant tissues. 43 This capability allows S. commune not only to thrive on dead plant material but also to colonize living tissues latently without immediately causing disease symptoms. 44 Previous research has shown that extracts from endophytic S. commune had strong antibacterial activity against S. aureus, Klebsiella pneumoniae, and Vibrio cholerae, with minimum inhibitory and bactericidal doses ranging from 1.25 to 10 mg/mL. 45 In addition, S. commune isolated from the leaves of Coleus amboinicus (a medicinal plant) demonstrated stronger antimicrobial activity against B. subtilis and C. albicans compared to other tested microorganisms. Phytochemical analysis indicated the presence of terpenoid compounds, which were suggested to contribute to the observed antimicrobial activity. 46 Various bioactive metabolites have been reported to be produced by endophytic S. commune. Several novel compounds, such as bisabolane-type sesquiterpenes, polyketides, and terpenoids isolated from S. commune associated with Gastrodia elata, have demonstrated strong antifungal activity. 47 In addition, compounds including mannonerolidol and mannogeranylnerol have been shown to possess antimicrobial effects against a range of pathogenic bacteria, notably B. subtilis and S. aureus. 48 Two active fusidane-type compounds (6 and 7), along with five new compounds (1–5) and nine known compounds (8–16) isolated from S. commune metabolites, demonstrated significant antibacterial activity against Stenotrophomonas maltophilia, with MIC values of 4 and 16 µg/mL, respectively. 49 Taken together, Isolate ECT57’s broad-spectrum antibacterial activity adds to the case for endophytic fungi from mangrove habitats as viable sources of new antibiotic options. The endophytic fungal isolate ECT63 (M. laterale), isolated from the branches of C. tagal, exhibited specific antifungal activity against C. albicans. To date, there have been no reports describing M. laterale as an endophytic fungus associated with mangrove plants. Therefore, this finding represents the first report documenting both the occurrence of M. laterale as a mangrove endophyte and its specific antifungal potential within a mangrove ecosystem. Several species within the genus Muyocopron have previously been reported as endophytic fungi, including M. laterale isolated from the leaves of Centella asiatica 50 and Hydrocotyle verticillata. 51
The results demonstrate that the genus Muyocopron can adapt as an endophyte across a wide range of plant hosts; however, its relationship with mangrove plants, notably C. tagal, has not been recorded before. In terms of secondary metabolite biosynthesis, members of the genus Muyocopron have been documented to produce novel azaphilone derivatives, specifically muyocopronones A and B, which demonstrated weak antibacterial activity against Gram-positive bacteria with a MIC of 128 µg/mL under in vitro conditions. 52 The antifungal activity of M. laterale against C. albicans discovered in this work expands the range of bioactivities known for this genus. Furthermore, these findings reinforce the importance of this work as the first to describe M. laterale as a mangrove-associated endophytic fungus, indicating its potential as a viable source of new antifungal chemicals.
The EtOAc extract of the endophytic fungi S. commune ECT57 exhibited a complicated metabolite profile as determined by LC-HRMS analysis. Compounds detected included phenolic acids, flavonoids, alkaloids, coumarins, amino acid derivatives, lipids, and cyclic peptides (diketopiperazines). In this study, the antimicrobial activity of ECT57 extract is thought to be related to its phytochemical content, such as phenolic acids, flavonoids, alkaloids, coumarins, amino acid derivatives, lipids, and cyclic peptides (diketopiperazines). Phenolic compounds are known to have strong antimicrobial activity due to their flexible structure and susceptibility to chemical modification. Their mechanism of action includes inhibition of cell wall synthesis, DNA replication, and bacterial enzyme production, thereby increasing the susceptibility of bacterial cells to antimicrobial effects. 53 Flavonoids can penetrate bacterial cell walls, increase membrane permeability, and cause damage to intracellular structures such as microsomes and lysosomes. 54 Flavonoids showed greater effectiveness against Gram-positive bacteria, 55 thus contributing to the higher inhibition of Gram-positive bacteria observed in this study. Alkaloids act as antibacterial agents by inhibiting bacterial cell wall synthesis, altering cell membrane permeability, disrupting bacterial metabolism, and inhibiting nucleic acid and protein synthesis. 56
This diversity suggests the existence of a multitarget antibacterial mechanism. The most abundant chemical was Cyclo(phenylalanyl-prolyl). This molecule belongs to the cyclic dipeptide (diketopiperazine) category, which has a variety of biological functions. This chemical serves as a microbial defense molecule, controls bacterial growth, and participates in the quorum-sensing mechanism. In addition to possessing antibacterial and antifungal properties. 57 6-Quinolinecarboxylic acid is a heterocyclic alkaloid derivative. The produced quinoline-3-carbonitrile derivative demonstrated antibacterial effectiveness against both Gram-positive and Gram-negative bacteria without causing considerable harm, with the mechanism presumed to be DNA gyrase inhibition. This chemical has the potential to be a foundation for the creation of broad-spectrum antibacterials. 58 Caffeic acid compound is a hydroxycinnamic acid reported to have antimicrobial activity against bacteria and fungi. 59 Caffeic acid shows antibacterial activity against S. aureus and can increase the effectiveness of several antibiotics when used in combination, so it has the potential to be a supporting agent in the therapy of multi-resistant bacterial infections. 60
Conclusion
Three endophytic fungi, N. saprophytica ECT52, S. commune ECT57, and M. laterale ECT63, have been genetically identified. Among them, the EtOAc extract of S. commune ECT57 showed the lowest MIC (62.5 ± 0.96) μg/mL against S. aureus, indicating stronger antimicrobial activity among the other isolates. LC-HRMS analysis of the extract revealed several prominent compounds, including Cyclo(phenylalanyl-prolyl) (approximately 13.1%), 6-Quinolinecarboxylic acid (4.04%), and Caffeic acid (nearly 1.02%). These findings indicate that C. tagal contains endophytic fungi that can synthesize antimicrobial compounds, which may have potential relevance for antibiotic development. Further studies, including cytotoxicity testing, are needed to evaluate safety and support the potential application of these bioactive compounds.
Footnotes
Acknowledgements
The authors gratefully acknowledge financial support for the research, authorship, and publication of this article from Universitas Andalas through the Doctoral Dissertation Research Scheme for the fiscal year 2025 (SK Number 75/UN16.19/PT.01.03/PDD/2025). The authors also thank all teams who contributed to this research.
Authors’ Contribution
All authors made substantial contributions to conception and design, acquisition of data, or analysis and interpretation of data; took part in drafting the article or revising it critically for important intellectual content; agreed to submit to the current journal; gave final approval of the version to be published; and agree to be accountable for all aspects of the work. All the authors are eligible to be an author as per the international committee of medical journal editors (ICMJE) requirements/guidelines.
Consent to Participate
Not applicable.
Consent for Publication
Not applicable.
Data Availability Statement
All relevant data generated or analyzed during this investigation are contained in the supplementary tables and figures that accompany this article
Declaration of Conflicting Interests
The authors declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Ethical Approval
This study does not involve any animal or human subjects.
Informed Consent
Not applicable.
Publisher’s Note:
All statements made in this article are exclusively those of the authors and do not necessarily reflect those of the publisher, editors, or reviewers. This publication remains neutral on jurisdictional claims in published institutional affiliation.
Use of Artificial Intelligence-assisted Tools:
The authors state that they did not utilize artificial intelligence (AI) techniques to write and revise the text, and that no photos were modified with AI.