
Abstract
Adult-onset hearing loss is associated with cognitive decline, although the mechanisms underlying this relationship remain unclear. Emerging evidence suggests that the early stages of hearing loss can alter brain network dynamics, which may be detectable before other neurocognitive markers. This systematic review aims to present evidence for altered brain connectivity in age-related hearing loss, and its subsequent relationship with cognitive performance. The review was preregistered in the Prospective Register of Systematic Reviews and conducted in accordance with the Preferred Reporting Items for Systematic Reviews and Meta-Analyses guidelines. We screened 203 articles from 4 databases, of which 25 cohort and case-control studies published between 2019 and 2025 met the inclusion criteria. According to the Newcastle–Ottawa Scale, 76% of the included studies were classified as high methodological quality. The findings show that older adults with age-related hearing loss exhibit changes in brain connectivity. A novel finding is the disruption of higher-order networks, such as the Default Mode Network, and its association with reduced performance in cognitive control functions, particularly shifting. This may shed light on the association between hearing loss and cognitive decline, although causality has yet to be established. These findings suggest that brain connectivity may serve as an early neural correlate of cognitive vulnerability and inform future research into interventions targeting cognitive decline.
Introduction
Population aging is occurring at an increasingly rapid rate; in 2020, the number of individuals over 60 years of age surpassed that of children under 5 years of age (World Health Organization, 2024). Many chronic noncommunicable diseases are age-related, including hearing loss and cognitive impairment. The prevalence of hearing loss in people over 60 years of age is estimated to be 65%, with 25% experiencing moderate or greater levels of hearing loss (Haile et al., 2021). According to the Global Burden of Disease study, the prevalence of Mild Cognitive Impairment (MCI) in individuals over 50 years of age is estimated to be 19.7% (Song et al., 2023). The average annual conversion rate from MCI to dementia is approximately 12.2%, nearly three times higher than that observed in the general population (Ghanem et al., 2024). Every year, there are 10 million newly reported cases of dementia worldwide (World Health Organization, 2025).
There is a well-documented association between adult-onset hearing loss and cognitive decline. The Lancet Standing Commission reported that hearing loss is a potentially modifiable risk factor for dementia and that, if it is causal, it could account for 7% of all cases worldwide (Livingston et al., 2024). A systematic review with a meta-analysis found that each 10 dB worsening in age-related hearing loss (ARHL) is associated with a 16% increase in the risk of developing dementia (Yu et al., 2024). Additionally, a longitudinal study that followed participants for eight years reported a faster rate of cognitive decline in individuals with hearing loss (Samelli et al., 2025). However, Machado-Fragua et al. (2025) reported lower hazard ratios (1.14 and 1.12 across two databases), corresponding to approximately a 12-14% increase in risk, which is comparable to estimates from studies on midlife hearing impairment. In any case, Dawes and Munro (2024) reviewed the evidence and concluded that there is currently no good quality evidence to prove, or disprove, that hearing loss causes dementia or that treating hearing loss will reduce the risk.
Several hypotheses have been proposed regarding the possible direction of causality between ARHL and cognitive decline. The review by Wayne and Johnsrude (2015) outlines four main hypotheses and an alternative framework. The Cognitive Load on Perception Hypothesis suggests that cognitive decline leads to audiometric hearing loss by reducing the cognitive resources available for auditory perception (Lindenberger & Baltes, 1994). In contrast, the Information Degradation Hypothesis proposes that cognitive decline is a consequence of compensatory efforts made in response to a degraded auditory signal and may be potentially reversible (Schneider & Pichora-Fuller, 2000). Similarly, the Sensory Deprivation Hypothesis posits that hearing loss leads to a more permanent cognitive decline, possibly through neuroplastic changes that adversely affect overall cognition (Lindenberger & Baltes, 1994). The Common Cause Hypothesis argues that a single underlying mechanism is responsible for both auditory and cognitive changes (Baltes & Lindenberger, 1997). The alternative framework combines the Information Degradation and Common Cause Hypotheses, proposing that neurodegenerative processes lead to hearing decline, sensory deficits, and increased demands on a reduced pool of cognitive resources. Sensory deficits resulting from both central nervous system damage and age-related inner ear pathology exacerbate perceptual challenges, requiring additional cognitive effort and ultimately leading to functional cognitive impairments (Wayne & Johnsrude, 2015).
The nature of the relationship between adult-onset hearing loss and dementia is not clear-cut (Munro & Dawes, 2025). Recent research has therefore shifted towards identifying neurobiological mechanisms that may underlie this association. Prior systematic reviews have focused primarily on structural neuroanatomy and molecular biomarkers in ARHL, reporting greater gray matter atrophy in temporal regions and increased tau accumulation (Lasica et al., 2025; Slade et al., 2022), findings linked to early cognitive decline and Alzheimer’s disease (Cody et al., 2024; Jack et al., 2018; van de Mortel et al., 2021). Disrupted brain connectivity—defined as alterations in the neuronal circuits and connections within the brain (Friston, 1994)—has been reported in cognitively normal individuals with abnormal cerebrospinal fluid amyloid and tau levels (Al-Ezzi et al., 2024), and has also been observed in at-risk populations prior to overt cognitive impairment or detectable volumetric changes.(Bidelman et al., 2019; Fitzhugh & Pa, 2022; Mai et al., 2024).
Network analyses suggest that the organization of the brain’s structural connections provides the anatomical scaffold that supports efficient information processing and complex brain functions (Park & Friston, 2013). Within this framework, functional connectivity is not a direct reflection of structural connectivity but rather emerges from the dynamic interactions of neuronal populations operating within an anatomically constrained system (Deco et al., 2013). This distinction has important implications, as functional and effective connectivity may be particularly sensitive to early network-level reorganization, even in the absence of detectable structural alterations.
In this context, brain connectivity is commonly described in terms of three complementary dimensions: Structural Connectivity, which refers to anatomical links; Functional Connectivity (FC), which indicates undirected statistical dependencies; and Effective Connectivity (EC), which represents directed causal relationships among distributed responses (Friston, 1994). The organizational principles of these interactions are often examined within the framework of intrinsic connectivity networks (ICNs), which extend the concept of resting-state networks to encompass large-scale, functionally connected brain systems detectable during both resting-state and task-based neuroimaging data (Laird et al., 2011). These networks are organized around highly connected hub nodes that facilitate efficient global communication across brain systems (van den Heuvel & Sporns, 2011). Prominent ICNs include the Default Mode Network (DMN), Salience Network (SN), and Frontoparietal Network (FPN), which together form the triple network model (Menon, 2023), among others.
Within this framework, functional connectivity can be further characterized according to its temporal properties: while traditional functional connectivity assumes temporal stationarity (Friston, 2011), dynamic functional connectivity captures time-varying coupling between brain regions (Hutchison et al., 2013). At the network level, static functional network connectivity (sFNC) refers to time-averaged statistical dependencies among large-scale networks, typically estimated as correlations between independent component (ICA) time courses across the entire scan (Jafri et al., 2008). In addition, dynamic functional network connectivity (dFNC) extends this framework by explicitly modelling time-varying reconfiguration of interactions among large-scale brain networks, including their transitions over time (Calhoun et al., 2014).
Emerging evidence suggests that early stages of hearing loss are associated with alterations in brain network dynamics. Functional reorganization occurs between auditory and language-related regions (Price et al., 2019), and disruptions involving the DMN and auditory networks have been proposed as candidate neural mechanisms linking ARHL and cognitive vulnerability (Gao et al., 2023; Li et al., 2023). Changes in EC within the DMN have been observed up to nine years before the clinical onset of dementia (Ereira et al., 2024). Although most available evidence is derived from cross-sectional studies and therefore does not allow causal or prognostic inferences, alterations in brain connectivity may represent early neural correlates of cognitive vulnerability in ARHL. Moreover, the current evidence does not rule out a mediating role for these connectivity changes in the association between ARHL and cognitive decline, underscoring the need for studies using alternative research designs to address these questions.
No comprehensive synthesis has focused explicitly on connectivity-based correlates of cognitive change in ARHL. Therefore, the aim of this systematic review was to address this gap by aggregating and analysing studies of brain connectivity in older adults with ARHL and their relationship with cognitive performance. Specifically, this review aims to describe and contextualize patterns of functional and effective connectivity reported in association with cognitive outcomes in ARHL. We hypothesise that specific disruptions in brain connectivity—particularly within the auditory network and the DMN—are associated with early network-level alterations linked to cognitive vulnerability in individuals with ARHL. By highlighting consistent patterns of connectivity disruption, this work may help inform future research aimed at developing early-stage diagnostic and intervention strategies for dementia prevention.
Materials and Methods
This systematic review was performed in accordance with the Preferred Reporting Items for Systematic Reviews and Meta-Analyses (PRISMA) (Page et al., 2021). The protocol was registered at the PROSPERO international prospective register of systematic reviews (PROSPERO ID: CRD420251075156).
Eligibility Criteria
The eligibility criteria for the studies were defined based on the PICOS framework, encompassing participants, interventions or assessments, comparators, outcomes, and study designs.
Participants
Adults aged ≥ 50 years with a pure-tone average > 25 dB HL measured by audiometry, bilateral sensorineural hearing loss consistent with age-related hearing loss (ARHL), and who were non–hearing aid users. Studies that included other types of hearing loss not typical of ARHL, such as unilateral or conductive hearing loss, were excluded. Studies were required to assess performance in at least one cognitive domain in addition to language (e.g., general cognition, memory, executive function, attention, or processing speed). Studies relying solely on self-reported measures to determine hearing loss were excluded, due to their low sensitivity for mild hearing loss (Mukari & Hashim, 2018). Studies involving animals were also excluded.
Interventions or Assessments
Assessment of brain connectivity using functional Magnetic Resonance Imaging (fMRI), Electroencephalography (EEG), or functional Near-Infrared Spectroscopy (fNIRS).
Comparators
Studies that included a control group of age-matched participants with normal hearing, defined as a pure-tone average (PTA) < 25 dB HL, based on audiometry.
Outcomes
A description of brain connectivity between regions of interest (ROIs), intra- and inter-network connectivity (connectivity within the same brain network and between different brain networks, respectively), and the relationship between connectivity and cognitive performance. The effect was determined by analyzing correlations or statistically significant differences between groups.
Study Designs
Analytical observational studies, such as case-control and cohort studies, were included. Reviews, case reports, and conference proceedings were excluded.
Information Sources
Studies were identified through a systematic search of the following databases: PubMed, EBSCO, Scopus, and Web of Science. Only articles written in English were considered, and no restrictions were applied regarding the date of publication. The search for studies was conducted in May 2025 and updated in January 2026.
Search Strategy
The following keywords, classified as MeSH terms, were utilized: Hearing Loss, Aging, and Presbycusis. Additionally, other related keywords were included, such as age-related hearing loss, functional connectivity, effective connectivity, brain connectivity, resting state and Intrinsic Connectivity Networks. The search expression used was: ((“Hearing Loss” AND “Aging”) OR (“Presbycusis” OR “age-related hearing loss”)) AND (“Functional Connectivity” OR “Effective Connectivity” OR “Brain Connectivity” OR “Resting State” OR “Intrinsic Connectivity Networks” OR “Network Organization” OR “Network Dynamics”).
Data Management
Articles were exported from the databases into Rayyan (https://www.rayyan.ai/), which was used to screen studies and record decisions. Duplicate records were removed within the platform prior to the initial screening. Reasons for exclusion were documented using the tools provided within the Rayyan platform for all excluded articles.
Selection Procedure
Studies were screened based on their titles and abstracts to assess their compliance with the inclusion and exclusion criteria. The full text of the selected articles was reviewed to examine in detail whether they included cognitive assessment methods and whether they considered results on brain connectivity. The first two stages were conducted independently by two reviewers: an initial title and abstract review, followed by a full-text review of the selected articles. Disagreements occurred in less than 3% of cases at both stages and were resolved by a third independent reviewer.
Data Collection Process and Data Items
Microsoft Excel was utilized for data extraction and management. The following information was gathered from the included articles: author and year, country, objective, population sample size, demographic characteristics, hearing measures, cognitive measures, and neuroimaging measures. The aim of the review was to provide a qualitative synthesis of the findings across studies. Consequently, no quantitative subgroup analyses were conducted.
Quality of Individual Studies
To evaluate the quality of the selected studies, we utilized the Newcastle-Ottawa Scale (NOS) for assessing nonrandomized studies in meta-analysis (Wells et al., 2011). This scale employs a point system to assess studies based on three main criteria: the selection of study groups, the comparability of these groups, and the ascertainment of either the exposure or outcome of interest for case-control or cohort studies, respectively. The maximum score, reflecting low risk of bias, is 9 points. Good quality: 3 or 4 points in the selection domain, and 1 or 2 points in the comparability domain, along with 2 or 3 points in the exposure/outcome domain. Regular quality (Acceptable): 2 points in the selection domain, and 1 or 2 points in the comparability domain, along with 2 or 3 points in the exposure/outcome domain. Poor quality: 0 or 1 point in the selection domain, or 0 points in the comparability domain, or 0 or 1 point in the exposure/outcome domain. The automated NotebookLM tool created by Google Labs was first used to classify article quality according to the NOS scale criteria. The results were then compared with the assessment of an independent reviewer. Eight studies showed discrepancies, which were resolved by a second independent reviewer.
Results
Search and Selection of Studies
The selection process is shown in the PRISMA flow diagram (Figure 1). The search identified a total of 203 articles: 49 articles in PubMed, 47 articles in EBSCO, 54 articles in SCOPUS and 53 articles in Web of Science. After removing duplicates, 126 articles were eliminated from consideration. The titles and abstracts of the remaining articles were reviewed. Out of 77 articles, 39 met the initial inclusion/exclusion criteria. After full-text review, 14 articles were excluded: 11 studies lacked cognitive measurements, 1 study did not include hearing measurements, and 2 studies focused on other types of hearing loss. In total, 25 articles were included in the analysis. Prisma flow diagram of the selection process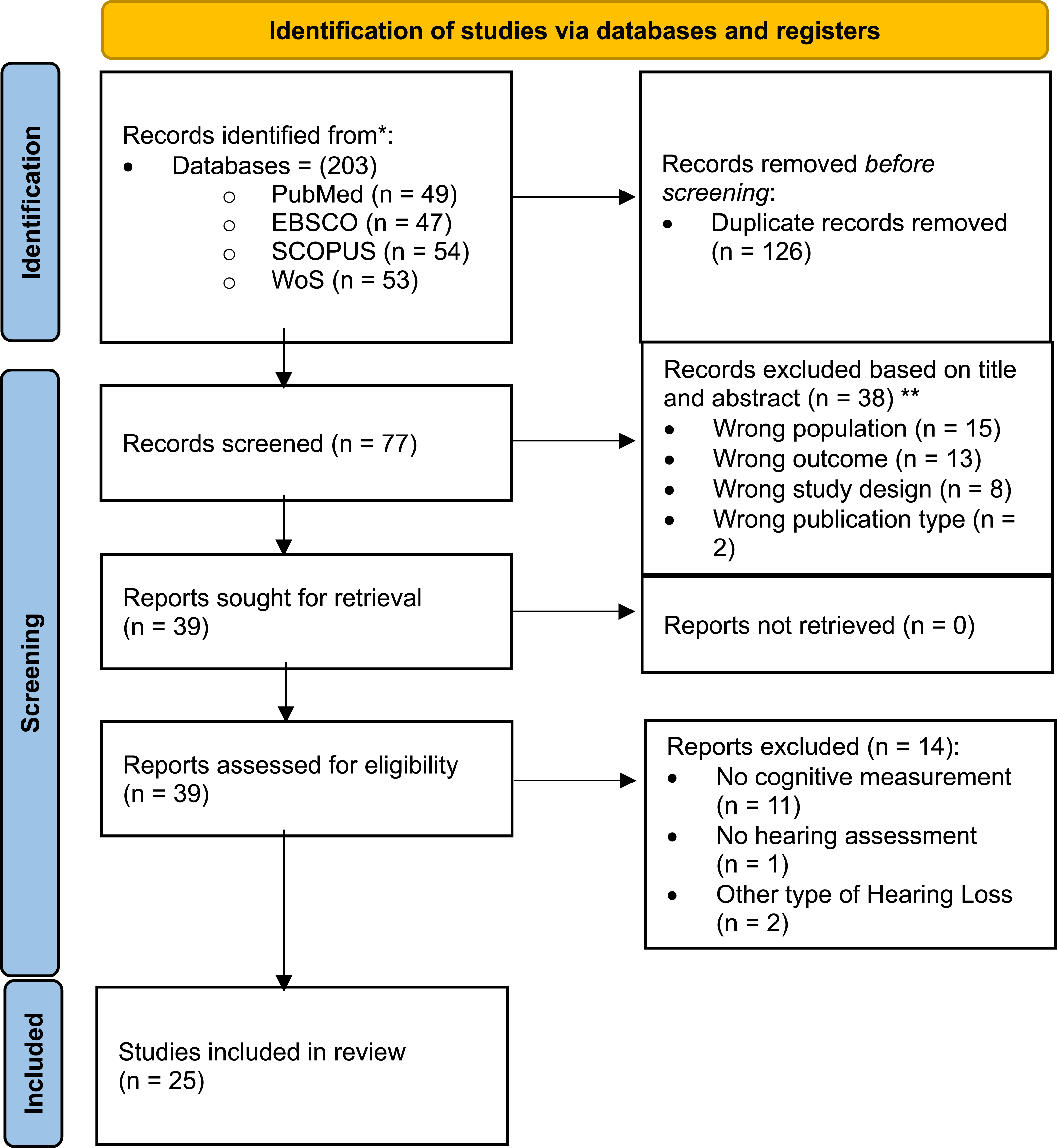
Study Characteristics
Overview of the Studies Included in the Systematic Review
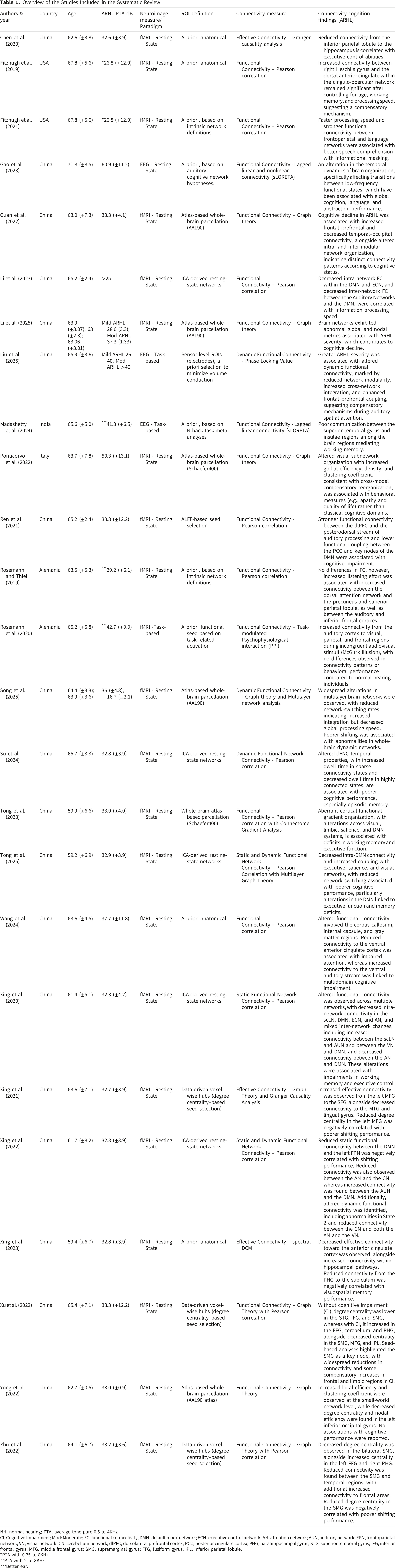
NH, normal hearing; PTA, average tone pure 0.5 to 4KHz.
CI, Cognitive Impairment; Mod: Moderate; FC, functional connectivity; DMN, default mode network; ECN, executive control network; AN, attention network; AUN, auditory network; FPN, frontoparietal network; VN, visual network; CN, cerebellum network; dlPFC, dorsolateral prefrontal cortex; PCC, posterior cingulate cortex; PHG, parahippocampal gyrus; STG, superior temporal gyrus; IFG, inferior frontal gyrus; MFG, middle frontal gyrus; SMG, supramarginal gyrus; FFG, fusiform gyrus; IPL, inferior parietal lobule.
*PTA with 0.25 to 8KHz.
**PTA with 2 to 8KHz.
***Better ear.
Study Characteristics, Reported Effect Sizes, and Quality Assessment Using the Newcastle–Ottawa Scale (NOS)

Quality was assessed using the Newcastle–Ottawa Scale (NOS), with total scores ranging from 0 to 9 points (Selection: 0–4; Comparability: 0–2; Exposure/Outcome: 0–3); NH, normal hearing; CI, Cognitive Impairment.
Two pairs of studies appear to share the same participant sample, as they report identical sample sizes and mean ages, although they pursued different research objectives. Li et al. (2023) examined intra and inter network FC in conjunction with glutamate and GABA levels, whereas Ren et al. (2021) applied Amplitude of Low-Frequency Fluctuation (ALFF) analyses slow-4 and slow-5 frequency bands to explore changes in low-frequency oscillation amplitudes. Similarly, Fitzhugh et al. (2019) explored resting state functional connectivity changes in the primary auditory cortex, while Fitzhugh et al. (2021) assessed predictors of speech-in-noise comprehension under informational and energetic masking conditions.
Participant Characteristics
The mean age of participants in all studies exceeded 60 years, except for Xing et al. (2023), which reported a mean age of 59.4 years. Regarding hearing loss classification, most studies used the pure tone average (PTA) criterion between 0.5 and 4 kHz. Exceptions included Fitzhugh et al. (2019), who used PTA between 0.25 and 8 kHz, and Rosemann (Rosemann et al., 2020; Rosemann & Thiel, 2019), who applied PTA between 2 and 8 kHz. According to the WHO (Olusanya et al., 2019) classification of hearing loss severity, most studies included participants with a mean hearing loss in the mild range (26–40 dB HL). Most studies calculated hearing thresholds using PTA across both ears; however, one study (Madashetty et al., 2024) defined hearing loss based on the PTA of the better ear. Three studies (Madashetty et al., 2024; Ponticorvo et al., 2022; Rosemann et al., 2020) involved participants with moderate hearing loss (41–60 dB HL). The only study classified within the severe hearing loss range (61–80 dB HL), based on its mean threshold (60.9 dB HL), also had the highest mean participant age (Gao et al., 2023). An exception is the cohort studies by Fitzhugh (2019 and 2022), which reported a mean hearing level of 26 dB HL (SD = 12), with many participants below 25 dB HL.
Region of Interest (ROI) Selection
There was substantial variability in ROI selection methods across studies. The most common approach was atlas-based whole-brain parcellation ROIs, used in 24% of studies (Guan et al., 2022; Li et al., 2025; Ponticorvo et al., 2022; Song et al., 2025; Tong et al., 2023; Yong et al., 2022). ICA-derived resting-state network ROIs were reported in 20% of studies (Li et al., 2023; Su et al., 2024; Tong et al., 2025; Wang et al., 2024; Xing et al., 2022), followed by a priori anatomical ROIs in 16% of cases (Chen et al., 2020; Fitzhugh et al., 2019; Wang et al., 2024; Xing et al., 2023). Data-driven voxel-wise hub ROIs identified using graph-theoretical metrics accounted for 12% of studies (Xing et al., 2021; Xu et al., 2022; Zhu et al., 2022). Less frequently used approaches included a priori intrinsic network–based ROIs in 8% of studies (Fitzhugh et al., 2021; Rosemann & Thiel, 2019), a priori task-based ROIs also in 8% (Madashetty et al., 2024; Rosemann et al., 2020), a priori auditory–cognitive ROIs in 4% (Gao et al., 2023), ALFF-based seed ROIs in 4% (Ren et al., 2021) and sensor-level ROIs (electrodes) selected a priori to minimize volume conduction effects in 4% of studies (Liu et al., 2025).
Methods for Assessing Brain Connectivity
Most studies used fMRI (88%) to assess brain connectivity, while three studies employed EEG (12%) (Gao et al., 2023; Liu et al., 2025; Madashetty et al., 2024). No included studies used fNIRS. Regarding the type of brain connectivity, Functional Connectivity (FC) was the most commonly investigated modality, accounting for 68% of the included studies (Fitzhugh et al., 2019, 2021; Gao et al., 2023; Guan et al., 2022; Li et al., 2023, 2025; Madashetty et al., 2024; Ponticorvo et al., 2022; Ren et al., 2021; Rosemann et al., 2020; Rosemann & Thiel, 2019; Tong et al., 2023, 2025; Wang et al., 2024; Xu et al., 2022; Yong et al., 2022; Zhu et al., 2022). Effective connectivity (EC) was examined in 12% of the studies (Chen et al., 2020; Xing et al., 2021, 2023). In addition, within the broader functional connectivity framework, some studies employed dynamic or network-based approaches. Dynamic Functional Network Connectivity (dFNC) was investigated in 12% of articles (Su et al., 2024; Tong et al., 2025; Xing et al., 2022), Static Functional Network Connectivity (sFNC) was also reported in 12% (Tong et al., 2025; Xing et al., 2020, 2022), and ROI-based dynamic functional connectivity analyses were applied in 8% of studies (Liu et al., 2025; J. Song et al., 2025).
With respect to connectivity analysis methods, Pearson correlation–based analyses were the most frequently used approach, reported in 52% of the studies (Fitzhugh et al., 2019, 2021; Li et al., 2023; Ren et al., 2021; Rosemann & Thiel, 2019; Su et al., 2024; Tong et al., 2023, 2025; Wang et al., 2024; Xing et al., 2020, 2022; Xu et al., 2022; Zhu et al., 2022). Graph theory-based connectivity analyses, incorporating nodal and network-level metrics, were reported in 36% of the articles (Guan et al., 2022; Li et al., 2025; Ponticorvo et al., 2022; Song et al., 2025; Tong et al., 2025; Xing et al., 2021; Xu et al., 2022; Yong et al., 2022; Zhu et al., 2022). Lagged linear and nonlinear connectivity analyses (Gao et al., 2023; Madashetty et al., 2024) and Granger causality analysis (Chen et al., 2020; Xing et al., 2021) were each reported in 8% of the studies. More specialised analytical approaches were each reported in a single study (4% each), including psychophysiological interaction (PPI) (Rosemann et al., 2020), connectome gradient analysis (Tong et al., 2023), spectral Dynamic Causal Modelling (DCM) (Xing et al., 2023), phase locking value (Liu et al., 2025) and multilayer network analysis (Song et al., 2025). Percentages are not mutually exclusive, as some studies employed multiple analytical approaches.
Connectivity Between Specific Regions
Summarizes Findings on Functional and Effective Connectivity Between the Most Frequently Reported Brain Regions Across Studies
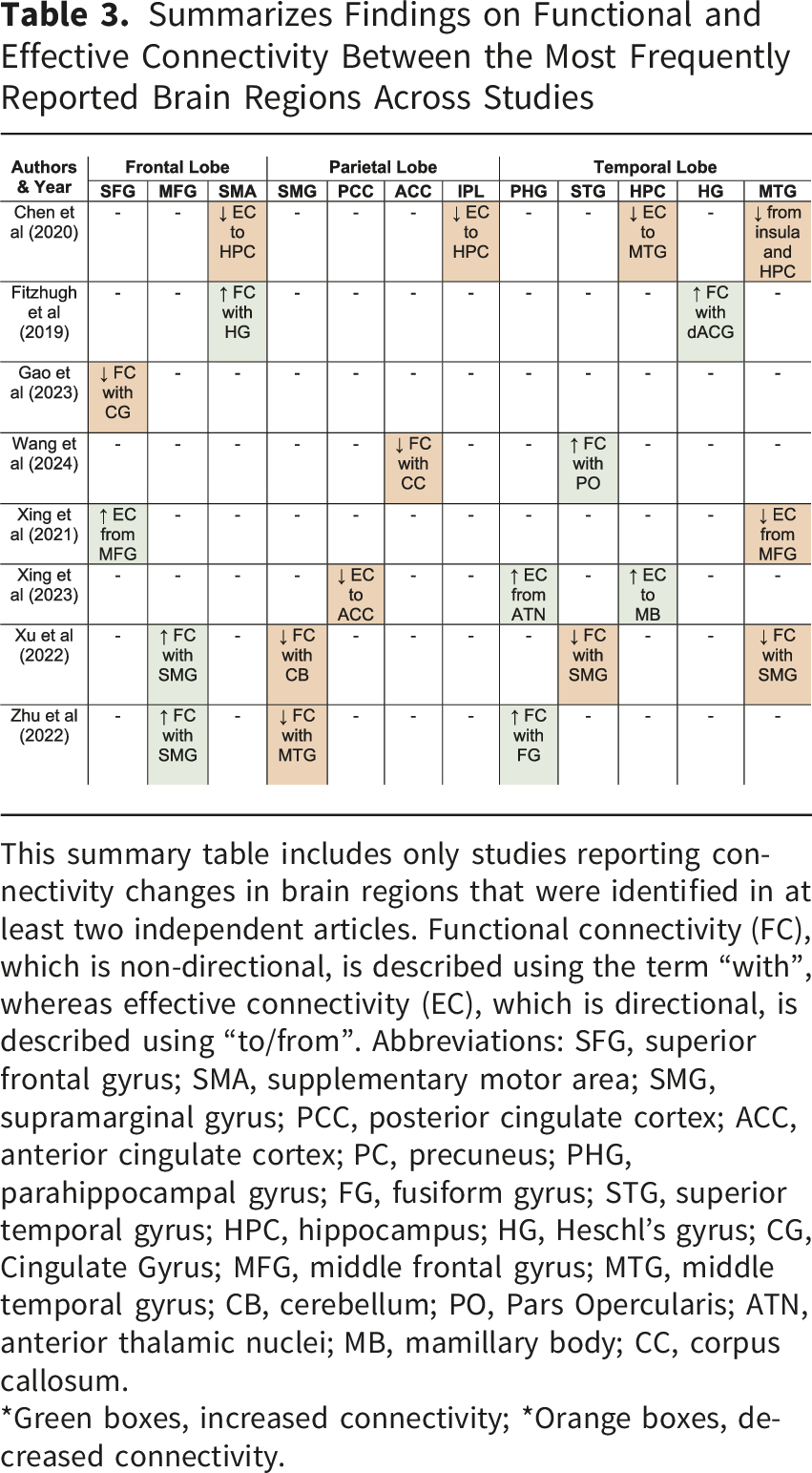
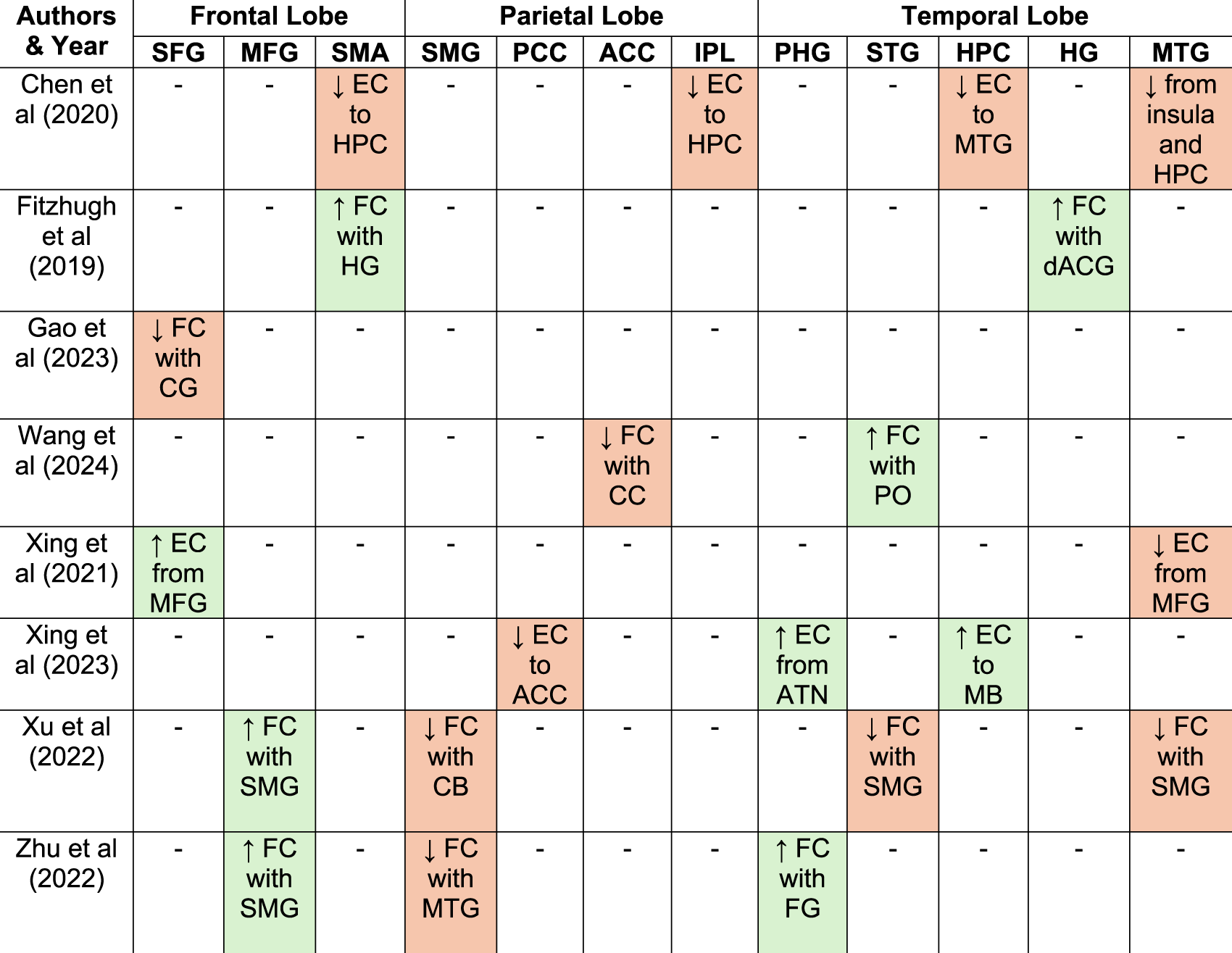
This summary table includes only studies reporting connectivity changes in brain regions that were identified in at least two independent articles. Functional connectivity (FC), which is non-directional, is described using the term “with”, whereas effective connectivity (EC), which is directional, is described using “to/from”. Abbreviations: SFG, superior frontal gyrus; SMA, supplementary motor area; SMG, supramarginal gyrus; PCC, posterior cingulate cortex; ACC, anterior cingulate cortex; PC, precuneus; PHG, parahippocampal gyrus; FG, fusiform gyrus; STG, superior temporal gyrus; HPC, hippocampus; HG, Heschl’s gyrus; CG, Cingulate Gyrus; MFG, middle frontal gyrus; MTG, middle temporal gyrus; CB, cerebellum; PO, Pars Opercularis; ATN, anterior thalamic nuclei; MB, mamillary body; CC, corpus callosum.
*Green boxes, increased connectivity; *Orange boxes, decreased connectivity.
On the other hand, studies that found increases in connectivity activity included Wang et al. (2024), which found an increase in FC from the STG to the pars opercularis. Fitzhugh et al. (2019) noted an increase in FC from the right Heschl’s gyrus (HG) to the SMA and the dorsal anterior cingulate gyrus (ACG). Gao et al. (2023) reported increases in gamma PSD in the MTG. Xing et al. (2023) found increased EC from the parahippocampal gyrus (PHG) to anterior thalamic nuclei (ATN) and from the hippocampus to the mamillary body (MB). Xu et al. (2022) found that the FC increased between SMG and MFG, a finding consistent with that reported by Zhu et al. (2022). Rosemann et al. (2020) observed increased cross-modal connectivity from the auditory cortex to visual, parietal, and frontal regions during incongruent audiovisual stimuli (McGurk illusion), although no differences in connectivity patterns or behavioral performance were observed compared to normal-hearing individuals.
Intra and Inter-network Connectivity
Included Studies Reporting Altered Intra/Inter-Network Functional Connectivity in ARHL
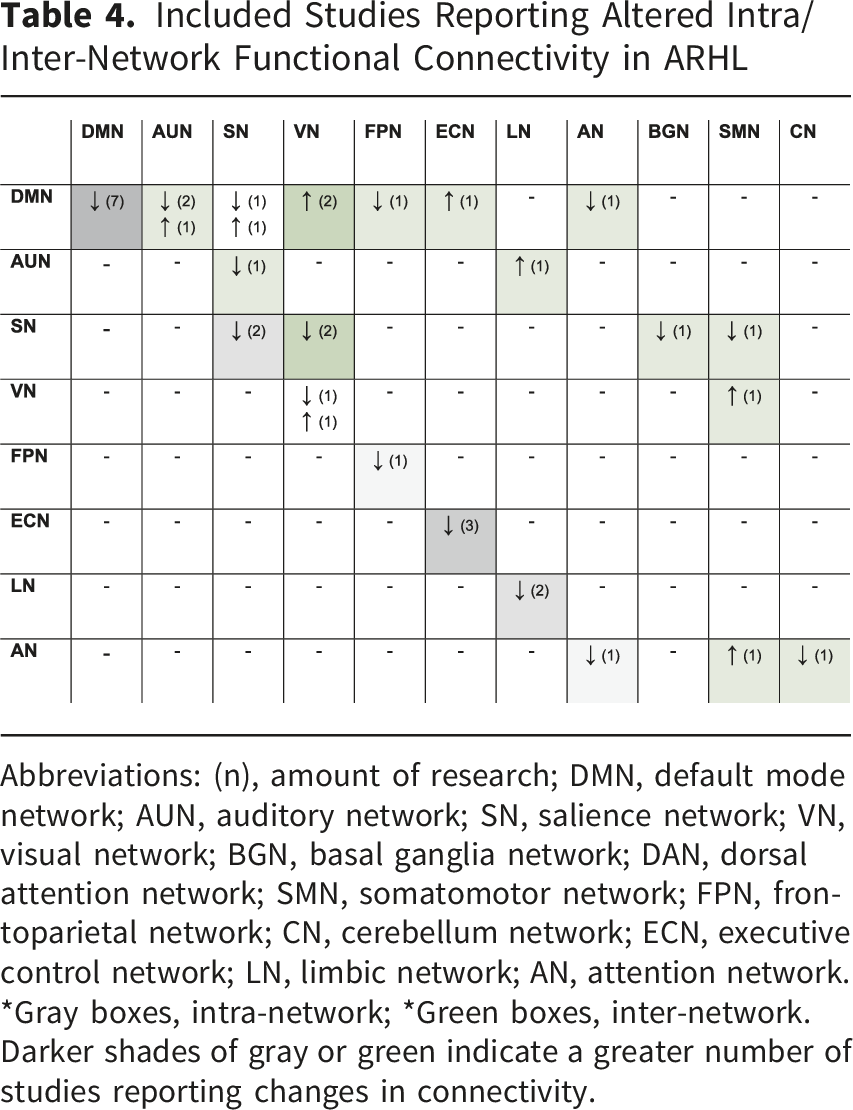
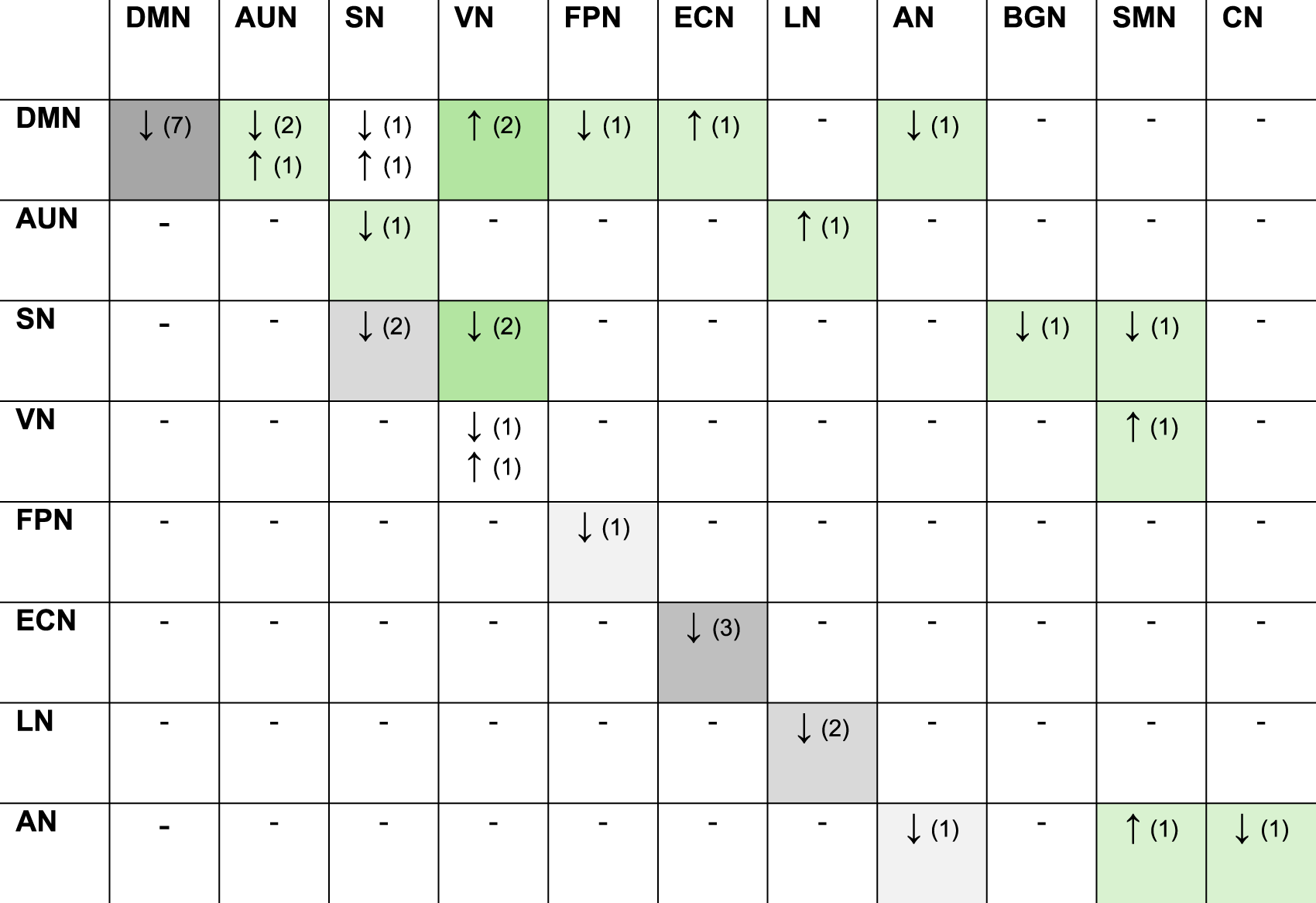
Abbreviations: (n), amount of research; DMN, default mode network; AUN, auditory network; SN, salience network; VN, visual network; BGN, basal ganglia network; DAN, dorsal attention network; SMN, somatomotor network; FPN, frontoparietal network; CN, cerebellum network; ECN, executive control network; LN, limbic network; AN, attention network.
*Gray boxes, intra-network; *Green boxes, inter-network. Darker shades of gray or green indicate a greater number of studies reporting changes in connectivity.
Regarding inter-network relationships, the DMN displayed altered connectivity with several other networks. There was decreased connectivity with the FPN (Xing et al., 2022) and AN (Xing et al., 2020) but increased connectivity with the ECN (Tong et al., 2025) and the VN (Tong et al., 2025; Xing et al., 2020). The SN also exhibited significant changes in its relationships with other networks, showing decreased connectivity with the VN (Li et al., 2023; Su et al., 2024), the BGN (Li et al., 2023), and the SMN (Su et al., 2024). The auditory network (AUN) has been shown to exhibit decreased connectivity with the SN (Tong et al., 2025) and increased connectivity with the LN (Xing et al., 2020). Additionally, the VN and AN exhibited increased FNC with the somatomotor network (SMN) (Su et al., 2024).
Neuropsychological Assessments Used Across Studies
A comprehensive battery of neuropsychological tests was utilized across all selected studies. Among studies that performed statistical comparisons between ARHL and normal-hearing groups, older adults with ARHL consistently performed poorer than their normal-hearing peers across measures of general cognition, as assessed by the Montreal Cognitive Assessment (MoCA) (Gao et al., 2023; Guan et al., 2022; Li et al., 2023; Su et al., 2024; Wang et al., 2024). They also exhibit poorer performance in specific cognitive domains, such as: attention with Trial Making Test (TMT) part A (Li et al., 2023; Ren et al., 2021; Su et al., 2024; Tong et al., 2025; Wang et al., 2024), shifting with TMT part B (Y.-C. Chen et al., 2020; Li et al., 2023; Ren et al., 2021; Su et al., 2024; Tong et al., 2025; Wang et al., 2024; Xing et al., 2020, 2021, 2022, 2023Xu et al., 2022; Yong et al., 2022; Zhu et al., 2022), inhibitory control with the Stroop (Li et al., 2023; Ren et al., 2021; Su et al., 2024; Wang et al., 2024), attention and information processing abilities with the Symbol Digit Modalities Test (SDMT) (Li et al., 2023; Su et al., 2024; Wang et al., 2024) and working memory with the Digit Span Test (DST) (Tong et al., 2023, 2025; Xing et al., 2020).
Association Between Brain Connectivity and Cognitive Performance
One of the most consistently reported findings across studies was the association between performance on the Trail Making Test-B (TMT-B), a measure of executive function that indexes cognitive set-shifting, and alterations in brain connectivity in adults with ARHL compared to age-matched peers without ARHL. Chen et al. (2018) found a positive correlation between TMT-B performance and both ALFF and ReHo in the left MFG. In contrast, Chen et al. (2020) reported negative correlations between TMT-B scores and directed connectivity, specifically reduced EC from the left IPL to the left hippocampus (r = −0.423, p = 0.025) and from the right IPL to the right hippocampus (r = −0.516, p = 0.005). Similarly, Tong et al. (2025) identified a negative correlation between TMT-B performance and sFNC within the DMN. Wu et al. (2024) observed that poorer TMT-B performance in individuals with ARHL was associated with increased activation in both the left and right premotor cortex (PMC). Additionally, Xing et al. (2022) reported a negative correlation between connectivity within the DMN and the lateral frontoparietal network (LFPN) and TMT-B performance. Finally, Zhu et al. (2022) found that lower degree centrality (DC) in the bilateral SMG was correlated with worse TMT-B outcomes.
As for other cognitive domains, Gao et al. (2023) reported that transitions between EEG microstates, specifically from state C to B (DeltaTM), were significantly related to PTA, general cognition MoCA scores, and language performance. Ren et al. (2021) found a negative correlation between Stroop scores (inhibitory control and interference) and ALFF in the left inferior parietal gyrus (IPG), as well as between TMT-B scores and ALFF in the left inferior temporal gyrus (ITG); in contrast, Symbol Digit Modalities Test (SDMT) scores (processing speed) showed a positive correlation with ALFF in the right SMG. Similarly, regions such as the dlPFC and PCC that showed altered ALFF also exhibited frequency-dependent changes in functional connectivity, which were associated with attention and executive control (Ren et al., 2021). In addition, Li et al. (2023) showed that frontal intra-network connectivity was positively related to processing speed, with stronger connectivity of the left superior frontal cortex associated with better SDMT performance (β = 4.213, p = 0.001, R2 = 0.571). In contrast, increased inter-network connectivity between the AN and the DMN was associated with poorer executive–attentional performance, as reflected by higher TMT-A scores (β = −41.399, p = 0.003, R2 = 0.614).
Xing et al. (2020) reported a positive correlation between Digit Span Test (DST) (working memory) performance and connectivity between the AUN and the DMN (ρ = 0.327, p = 0.040), while Xing et al. (2023) also linked EC from the PHG to the subiculum with scores on the Complex Figure Test–Delayed Recall (CFT-delay) (visuospatial memory) (rho = −0.259, p = 0.044). Fitzhugh et al. (2019) found that associations between functional connectivity of Heschl’s gyri and age, working memory, or processing speed were no longer significant after controlling for hearing ability, suggesting that connectivity changes in these regions were primarily driven by hearing loss rather than by these variables. Xu et al. (2022) reported distinct connectivity patterns from the SMG in individuals with presbycusis, depending on cognitive status. When comparing cognitively impaired and unimpaired individuals, the MFG, inferior frontal gyrus, PHG, rolandic operculum, and cerebellum emerged as key regions differentiating connectivity profiles. Using an auditory spatial selective attention paradigm, Liu et al. (2025) reported reduced network modularity and a shift from right parietal–frontal to frontal–prefrontal coupling, consistent with compensatory frontal engagement. Similarly, Song et al. (2025) found that higher network-switching rates were positively associated with cognitive performance in individuals with ARHL, underscoring the relevance of dynamic network reconfiguration.
Discussion
Most of the included studies were rated as good methodological quality according to the Newcastle–Ottawa Scale, reflecting adequate control of key confounders and acceptable analytical rigor in the assessment of brain connectivity. Although sample size and effect size reporting were heterogeneous, and effect size measures were not consistently reported across studies, the overall quality of the evidence provides a reasonable basis for interpreting convergent patterns of altered connectivity in adults with ARHL.
Across studies, different analytical frameworks were used to quantify brain connectivity as a shared object of interest, albeit from complementary perspectives. Pearson correlation–based functional connectivity captures undirected temporal covariation between regional signals, whereas effective connectivity approaches (e.g., Granger causality, DCM) aim to model directed influences. Graph-theoretical analyses further summarize functional connectivity matrices in terms of network topology (e.g., modularity, efficiency, hub organization). Dynamic approaches (e.g., dFNC) extend correlation-based measures by characterizing time-varying coupling patterns and state transitions. In EEG studies, lagged connectivity metrics applied in source space seek to mitigate volume conduction by emphasizing time-delayed dependencies.
The review results show that adults with ARHL exhibit altered brain connectivity compared with age-matched peers with normal hearing. Hearing loss not only affects connectivity during auditory signal processing but also induces a global reorganization of the brain, evident both at rest and during non-auditory tasks. A novel finding is the disruption of higher-order networks, such as DMN, and its association with reduced performance in cognitive control functions, such as shifting, which do not directly rely on auditory input. Although this reflects an association rather than a proven causal link, it may reflect an early correlate of neurocognitive vulnerability in ARHL.
Intra and Inter-network Connectivity
There is substantial evidence that individuals with ARHL exhibit altered organization of the intrinsic connectivity network. DMN comprises a collection of widely distributed and interconnected brain regions that are typically suppressed when a person is focusing on an external stimulus. However, when attention is not directed toward an external stimulus, such as at periods of rest, the DMN switches to an “internal focus”, facilitating various processes (Menon, 2023). Five cognitive functions are commonly associated with DMN: self-reference, social cognition, episodic memory, language and semantic memory, and mind-wandering. Inadequate suppression of DMN has been linked to impaired selective attention (Weissman et al., 2006). Ereira et al. (2024) examined alterations in EC and explored the connections between DMN dysconnectivity and major dementia risk factors, revealing strong relationships with polygenic risk for Alzheimer’s disease and social isolation. Alterations in the DMN in individuals with ARHL have also been associated with deficits in executive function and memory (Tong et al., 2025), suggesting that degraded auditory input may contribute to higher-order cognitive impairment.
Some nodes play a critical integrative role by participating in more than one intrinsic connectivity network (see Supplementary Table 1), thereby acting as connector hubs that support communication across large-scale brain systems. Alterations in these shared nodes in ARHL may therefore have consequences that extend beyond local connectivity changes, contributing to a reconfiguration of network-level architecture. For example, the ACC (Wang et al., 2024; Xing et al., 2023) participates in both the DMN and the SN, positioning it as a key hub for coordinating affective regulation and cognitive control (Schimmelpfennig et al., 2023). The superior frontal gyrus (SFG) has been identified as a shared node between the DMN and the FPN (Gao et al., 2023; Xu et al., 2022), suggesting its involvement in both self-referential processes, attention, goal-directed behavior, and cognitive control (Marek & Dosenbach, 2018). The MFG, a key hub of the FPN and the Executive Control Network (ECN), has consistently been linked to individuals with ARHL (Xing et al., 2021; Xu et al., 2022; Zhu et al., 2022), underscoring its role in higher-order cognitive regulation. Additionally, the IPL and SMG are associated with both the FPN/ECN and the FPN/AN, respectively, as reported in several studies (Chen et al., 2020; Xu et al., 2022; Zhu et al., 2022). In particular, the IPL, a core node of the DMN, may contribute to disrupted DMN-related processes and cognitive decline in individuals with ARHL (Chen et al., 2020). Cognitive decline in older adults may also be associated with reduced connectivity between the FPN and the DMN. This pattern aligns with the neural dedifferentiation hypothesis, which posits that neural activity in older adults becomes more diffusely distributed across overlapping populations, resulting in less distinct neural representations (Koshino et al., 2023).
Recent evidence indicates that ARHL is associated with disruptions in the temporal dynamics and stability of functional connectivity patterns, with potential relevance for cognitive performance (Liu et al., 2025; Song et al., 2025). Xing et al. (2022) found that the ARHL group exhibited abnormal dynamic functional network connectivity (dFNC) in State 2, characterized primarily by highly positive connectivity reflecting increased interconnection between brain networks. This group also showed decreased dFNC between the CN, the AUN, and the VN. Furthermore, the ARHL group demonstrated shorter mean dwell time (MDT) and reduced fractional time (FT), suggesting less stable engagement in specific connectivity states. Similarly, Su et al. (2024) observed alterations in the ARHL group, noting a higher presence in State 3, which is associated with a sparsely segregated network configuration. Together, these findings indicate that ARHL is associated not only with altered connectivity strength but also with changes in the temporal organization and stability of large-scale brain networks. Therefore, these temporal dynamics may also reflect frequency-dependent alterations in spontaneous neural activity. In this context, network switching refers to the rate at which a specific brain region transitions between different functional networks (Pedersen et al., 2018). Several studies suggest that this switching rate is linked to the learning of simple motor tasks, attention and executive function (Bassett et al., 2011; Braun et al., 2015; Shine et al., 2016) and predicts performance in higher-order cognitive functions, including working memory, planning, and reasoning (Pedersen et al., 2018). Within this framework, a hierarchical model of network reorganization in ARHL has been proposed (Song et al., 2025). Reduced dynamic flexibility in prefrontal regions may impair DMN suppression, leading to compensatory hyperconnectivity in parietal regions and reduced global integration efficiency. Together, these multilevel changes suggest a transition from localized dysfunction to widespread network reorganization underlying cognitive decline.
Association Between Brain Connectivity and Cognitive Performance
The reduction in intra- and inter-network connectivity associated with ARHL has also been linked to lower cognitive performance, specifically shifting. Shifting has been defined as the ability to adapt behavior in response to changing circumstances quickly and is one of the three core executive functions in Miyake’s model, alongside inhibitory control and updating (Miyake et al., 2000). This function relies on working memory and inhibitory control as prerequisites during problem-solving processes (Uddin, 2021). One of the main paradigms used to study shifting is the TMT (Tombaugh, 2004) which requires connecting numbers in sequence (Part A) and then alternating between linking numbers and letters (Part B). Zhu et al. (2022) found a negative association between connectivity in the SMG and TMT scores, suggesting compensatory over-activation in the context of cognitive decline. Other studies, like Xing et al. (2022) and Wang et al. (2024), further support the idea that both static and dynamic FC alterations in ARHL are associated with performance on cognitive tasks. For instance, reduced FC between the DMN and LFPN correlates with poorer performance on TMT-B, a task involving executive functions, particularly shifting. This is consistent with the notion that efficient shifting depends on the integrity of core executive processes, including working memory, which is required for optimal task performance and has also been reported to be altered in ARHL (Fitzhugh et al., 2019; Madashetty et al., 2024; Tong et al., 2023; Xing et al., 2020).
These findings support the hypothesis that shifting depends on the dynamic integration between brain regions, with the reconfiguration of neural networks serving as a fundamental neuropsychological mechanism underlying executive function (Braun et al., 2015). Frontal integration has been negatively correlated with performance on the TMT-B, suggesting that the ability of the frontal cortex to switch between network modules represents a broader feature of cognitive functioning. This further highlights the critical importance of intersystem connectivity in the successful execution of more cognitively demanding tasks (Braun et al., 2015). Consistent with this dynamic framework, EEG microstate transitions have been associated with global cognition, as well as language and abstraction performance in individuals with ARHL (Gao et al., 2023), supporting the role of dynamic brain reconfiguration in cognitive functioning.
In more ecologically valid contexts, Fitzhugh et al. (2021) highlighted the importance of the FPN for speech comprehension in multi-speaker environments in older adults, suggesting that executive control mechanisms supported by this network play a critical role under complex listening conditions. In line with this, Li et al. (2023) reported decreased intra-network FNC within the DMN and ECN, as well as reduced inter-network FNC between the auditory network and the DMN, with these reductions associated with slower information processing speed. These findings suggest that hearing loss may disrupt both auditory processing and its integration with higher-order cognitive networks. Likewise, Madashetty et al. (2024) reported reduced FC between the STG and insular regions during working memory tasks in individuals with ARHL, an effect that persisted after controlling for age, education, and processing speed. However, not all connectivity alterations in ARHL are directly related to cognitive performance. For instance, Ponticorvo et al. (2022) found that changes in visual network organization were associated with behavioral measures such as apathy and quality of life, rather than with traditional cognitive domains, suggesting a compensatory reorganization of visual networks.
Connectivity Between Specific Regions
The differences observed across studies regarding decreased or increased connectivity in individuals with ARHL may vary depending on the specific brain region involved. A neurodegenerative process would be primarily associated with reduced connectivity (Knight & Verkhratsky, 2010), whereas an adaptive process involving compensatory mechanisms would be linked to increased connectivity (Hillary & Grafman, 2017). Consistent with this compensatory framework, Fitzhugh et al. (2019) reported increased connectivity between right Heschl’s gyrus and the cingulo-opercular network, which was specifically driven by hearing loss within the speech frequency range and predominantly in the left ear, suggesting a frequency- and ear-specific reorganization. Cross-modal reorganization may also contribute to these patterns. Increased auditory-to-visual, parietal, and frontal connectivity in ARHL during incongruent audiovisual processing (McGurk illusion) occurs without behavioral differences, suggesting compensatory recruitment (Rosemann et al., 2020). Overall, connections towards frontal regions tend to be increased (Gao et al., 2023; Xu et al., 2022; Zhu et al., 2022), while those toward temporal regions tend to be decreased (Xing et al., 2023; Zhu et al., 2022).
Some of the main regions reported as showing connectivity changes were the SMG, MFG, and MTG, each located in different cerebral lobes—parietal, frontal, and temporal, respectively. The SMG showed increased connectivity with the frontal region, specifically the MFG, while decreased connectivity was observed with the temporal region, particularly the MTG, as well as with the cerebellum. The SMG has been associated with higher-order cognitive functions and is considered a key node in the short-term memory network involved in retaining an abstract representation of serial order information, independently from content information (Guidali et al., 2019).
In contrast, the MTG showed decreased connectivity with the insula, hippocampus, and the SMG. The MTG has been highlighted for its role in multisensory integration supporting the formation of semantic and episodic memory (Chen et al., 2016), and it has been associated with early stages of cognitive decline (Berron et al., 2020). Xing et al. (2021) employed EC analysis and reported reduced directional connectivity from the MFG to the MTG, possibly reflecting diminished top-down frontotemporal interactions that typically facilitate prior predictions in the context of degraded sensory input (Cope et al., 2017). In addition, several studies reported alterations involving limbic structures (Chen et al., 2020; Xing et al., 2023; Xu et al., 2022). These regions are critically involved in memory-related processes, suggesting that ARHL-related connectivity alterations may extend beyond perceptual systems to include memory circuits.
Other regions of interest showing altered connectivity in ARHL include the SFG (Gao et al., 2023; Xing et al., 2021) and the hippocampus (Chen et al., 2020; Xing et al., 2021), as both are considered part of the rich-club organization. Rich-club nodes are defined as the top 10 brain regions (representing the top 12% of all nodes) with the highest average nodal degree, calculated from the group-averaged cortical connectivity network (Guan et al., 2022). This organization is thought to support the brain’s dynamic and diverse functions (Park & Friston, 2013). Disruption of these nodes may lead to disorganization of large-scale brain networks, linking ARHL to aberrant connectivity patterns (Tong et al., 2025) that may underlie its association with cognitive decline. Accordingly, Guan et al. (2022) suggested that rich-club alterations in presbycusis without cognitive decline may reflect compensatory plastic reorganization, whereas the onset of cognitive decline appears to disrupt this balance and contribute to a more maladaptive topological reorganization.
General Findings on Brain Connectivity
Most findings support the presence of brain connectivity changes in ARHL, with results that complement each other across studies by highlighting different brain regions or networks. Only within the intra-network in VN and the inter-network DMN-SN were contradictory findings observed, with one study reporting increased connectivity and another reporting decreased connectivity. In another study by Rosemann and Thiel (2019), categorized as regular quality, no differences in FC were reported between older adults with hearing loss and normal-hearing individuals. Moreover, they found that greater listening effort was significantly associated with reduced FC, specifically reflected in decreased resting-state coupling between the dorsal attention network and the precuneus and superior parietal lobule, as well as between the auditory cortex and the inferior frontal cortex. Therefore, they concluded that it is not hearing loss itself that affects brain connectivity, but rather the individually perceived listening effort. With hearing loss, this increased cognitive and listening effort may elevate cognitive load and reorganize higher-order networks that do not depend on auditory input.
Within this connectivity framework, EC provides more detailed information regarding the directional and causal relationships between brain regions or networks. However, it has been scarcely explored in relation to ARHL and its association with cognitive performance. In this review, only two studies addressed this issue: Chen et al. (2020), who applied the Granger Causality Analysis approach, and Xing et al. (2023), who used spectral Dynamic Causal Modelling (DCM). Friston (2011) emphasizes that measures such as correlation, coherence, or Granger causality reflect statistical dependencies, but do not necessarily indicate changes in the underlying neuronal coupling. Therefore, to make mechanistic inferences about connectivity between specific brain regions, approaches such as DCM are recommended, as they allow for the direct assessment of changes in EC, overcoming the limitations of analyses based solely on correlations (Friston, 2011). A potential clinical application of DCM is the development of models whose parameters have sufficient sensitivity and specificity to accurately identify subgroups of patients characterized by distinct pathophysiological mechanisms (Stephan & Friston, 2010). Therefore, further studies assessing EC in this population are needed to better understand the nature of connectivity patterns.
Altered brain connectivity patterns in ARHL may represent early neural correlates of cognitive vulnerability or risk. In this context, Li et al. (2025) suggest that ARHL may involve broader central nervous system mechanisms, including glymphatic dysfunction and large-scale network reorganization, supporting the view of ARHL as a central nervous system disorder with important implications for dementia prevention. The integration of less-explored metrics, such as EC and dFNC, and their associations with cognitive performance, represents a promising direction for future research. Connectivity disruptions in intra- or inter-network ICNs may also guide target selection for neuromodulation interventions (e.g., transcranial direct current stimulation (tDCS) and transcranial alternating current stimulation (tACS)), auditory training, or cognitive rehabilitation programs in the early stages of ARHL.
Although some patterns in the relationship between connectivity and cognitive performance can be identified across ARHL studies, the evidence remains highly heterogeneous. Such heterogeneity spans cognitive domains, connectivity metrics, and the specific brain regions or networks examined, which limits the ability to establish a consistent framework linking ARHL, connectivity, and cognition. Moreover, the limited number of studies explicitly investigating the underlying mechanisms, together with the lack of control over the temporal sequence of events, further constrains the delineation of specific networks that should be prioritized for future research.
Limitations of the Systematic Review
A limitation of this review is that, although we assessed study quality and risk of bias, we did not explicitly weight them in the interpretation of the results. This decision was based on the small number of studies that met the inclusion criteria and the fact that none were classified as poor quality or at high risk of bias. Additionally, no restrictions were applied regarding the methods used to assess changes in connectivity, which resulted in considerable variability in approaches across the included studies. Finally, this review provides information only on associations between hearing loss, brain connectivity, and cognition, without addressing causality.
Conclusion
A novel finding in ARHL is the disruption of higher-order networks, such as the DMN, and its association with reduced performance in cognitive control functions, particularly shifting, which do not directly rely on auditory input. This association may reflect early network-level alterations linked to neurocognitive vulnerability in ARHL. Consistent with the correlational nature of the available evidence, these findings suggest that brain connectivity may serve as an early neural correlate of cognitive vulnerability and inform future research on interventions targeting cognitive decline.
Supplemental Material
Supplemental Material - Impact of Age-Related Hearing Loss on Brain Connectivity and Cognitive Performance: A Systematic Review
Supplemental Material for Impact of Age-Related Hearing Loss on Brain Connectivity and Cognitive Performance: A Systematic Review by Sebastián Rivera, Andre Gómez-Lombardi, María Francisca Alonso, Steren Chabert, Kevin J Munro and Wael El-Deredy in Trends in Hearing.
Footnotes
Acknowledgements
We acknowledges the financial support of ANID Chile: Fondecyt 1241695 & 13240064, and the centre AC3ECIA250006, and ValER, Valencian Foundation for Excellence in Research, Spain. SR acknowledges the financial support of ANID Chile through the Subdirectorate of Human Capital, National Doctoral Scholarship 2024-21241054. MFA acknowledges the financial support of ANID Chile through Fondequip EQM220060. KJM is supported by the NIHR Manchester Biomedical Research Centre (NIHR203308).
Author Contributions
Conceptualization: SR, AG, WE. Methodology: SC, MA. Validation: SC, KM, Formal analysis: SR, AG Investigation (data/evidence collection): SR, AG, MA. Data curation: SR. Writing – original draft: SR, KM, SC. Writing – review & editing: SR, KM, WE
Funding
The authors received no financial support for the research, authorship and/or publication of this article.
Declaration of conflicting interest
The authors declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Supplemental Material
Supplemental material for this article is available online.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.