
Abstract
Astaxanthin (AST) is a marine-derived xanthophyll carotenoid widely distributed in aquatic organisms and edible seafood. Owing to its unique molecular structure and potent antioxidant properties, AST has attracted considerable attention as both a nutraceutical and a potential therapeutic agent. Increasing evidence indicates that oxidative stress and inflammation are interconnected processes that contribute to the onset and progression of numerous chronic diseases. In this context, AST has emerged as an important modulator of cellular defense mechanisms. This review summarizes current knowledge regarding the effects of AST on two major redox-sensitive transcription factors, nuclear factor erythroid 2-related factor 2 (Nrf2) and nuclear factor kappa B (NF-κB). Nrf2 coordinates the expression of genes involved in antioxidant defense and detoxification, whereas NF-κB regulates inflammatory and immune responses through the induction of cytokines, chemokines, and adhesion molecules. Particular attention is given to the molecular crosstalk between these pathways and the mechanisms through which AST restores redox and inflammatory homeostasis. Furthermore, recent experimental and clinical evidence supporting the therapeutic potential of AST in diseases associated with oxidative stress and chronic inflammation is discussed.
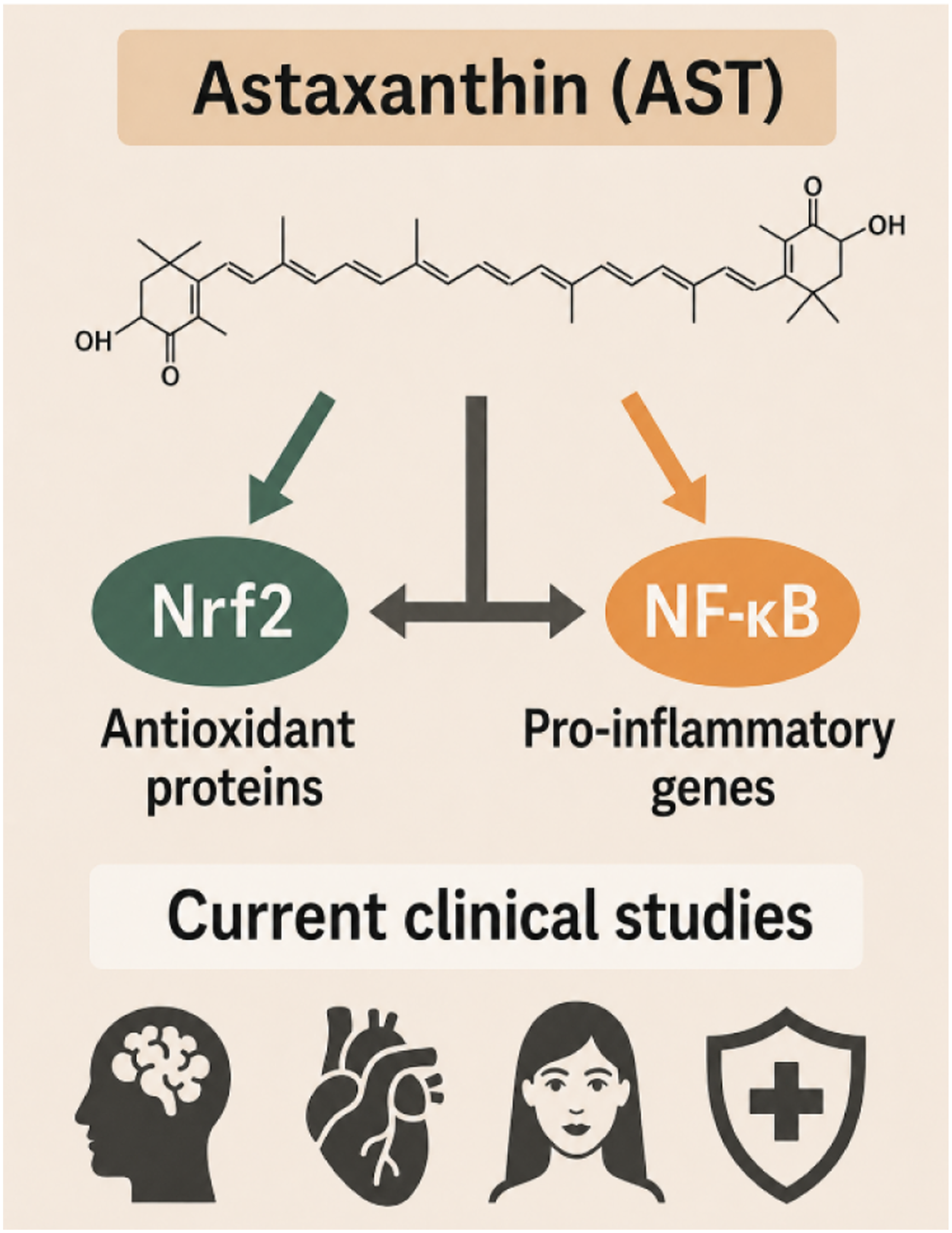
Highlights
Astaxanthin (AST) is a potent dietary antioxidant with broad nutraceutical and pharmaceutical applications.
AST regulates the redox-sensitive transcription factors Nrf2 and NF-κB, which are central to oxidative stress and inflammatory responses.
Nrf2 activation by AST enhances cellular antioxidant defenses, while NF-κB suppression reduces pro-inflammatory gene expression.
AST influences the crosstalk between Nrf2 and NF-κB, modulating redox-inflammatory homeostasis.
Preclinical studies support AST’s cytoprotective effects in chronic diseases involving oxidative stress and inflammation.
Clinical evidence remains limited; further human trials are required to confirm AST’s regulatory impact on Nrf2/NF-κB pathways.
Introduction
Astaxanthin (AST) is a naturally occurring oxygenated carotenoid belonging to the xanthophyll family, widely distributed in aquatic ecosystems and marine organisms, where it contributes to their characteristic pink-to-red pigmentation. 1 It is synthesized primarily by microalgae, phytoplankton, and certain marine bacteria, forming the basis of its accumulation along aquatic food chains and its presence in fish and seafood products. 2 Because of this distribution, AST is considered one of the most important marine-derived carotenoids with relevance for both nutrition and biomedical research.
Natural sources of AST, particularly the microalga Haematococcus pluvialis, have attracted major industrial and scientific interest due to their high accumulation capacity under stress conditions. 3 Beyond its application in animal nutrition, AST has been approved for human use in dietary supplements following safety evaluations, which has significantly expanded its biomedical relevance. 4 In aquaculture, AST supplementation remains widely used to enhance pigmentation in farmed fish and crustaceans, reflecting its well-established role as a feed additive.5–7
Structurally, AST is distinguished from other carotenoids by the presence of terminal keto and hydroxyl groups, which contribute to its amphipathic nature and strong antioxidant capacity. 5 These structural features enable efficient interaction with lipid bilayers and reactive oxygen species, supporting its role in protecting cellular components against oxidative damage. 6
The biological significance of these properties becomes particularly evident in the context of oxidative stress and inflammation. Chronic oxidative stress is now recognized as a key driver of inflammatory signaling, and together these processes contribute to the pathogenesis of a wide range of diseases, including cardiovascular disorders, metabolic dysfunction, neurodegeneration, and cancer. 7
The molecular interactions underlying the balance between oxidative stress and inflammation, particularly the crosstalk between Nrf2 and NF-κB signaling pathways, are schematically illustrated in Figure 1. Oxidative stress–inflammation network and regulatory crosstalk between Nrf2 and NF-κB signaling pathways. Reactive oxygen species (ROS) act as central mediators of cellular stress, triggering activation of NF-κB–dependent inflammatory responses. In parallel, oxidative stress induces Nrf2 activation, leading to transcription of antioxidant and cytoprotective genes. The bidirectional interaction between NF-κB and Nrf2 illustrates a tightly regulated feedback system in which inflammatory and antioxidant responses counterbalance each other to maintain redox homeostasis.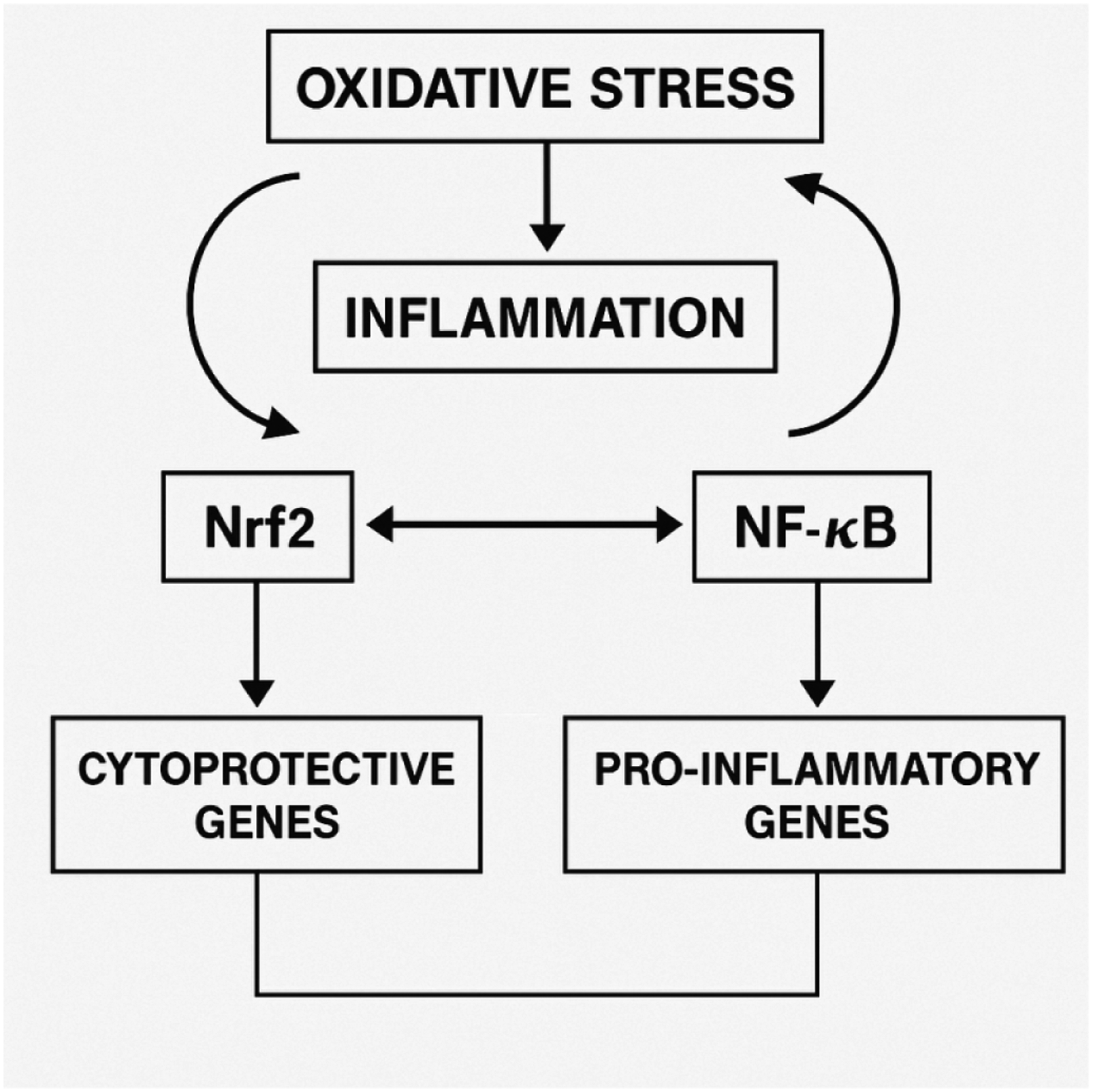
In this context, AST has been extensively studied for its ability to modulate oxidative and inflammatory responses at the cellular level. 8
A growing body of evidence suggests that AST exerts its biological effects, at least in part, through the regulation of redox-sensitive transcription factors. Among these, nuclear factor erythroid 2-related factor 2 (Nrf2) plays a central role in maintaining cellular redox homeostasis by inducing the expression of antioxidant and cytoprotective genes via binding to antioxidant response elements (AREs). 9 Activation of the Nrf2 pathway is therefore considered a major endogenous defense mechanism against oxidative injury. 10
In parallel, nuclear factor kappa B (NF-κB) represents a key transcriptional regulator of inflammatory and immune responses, controlling the expression of cytokines, chemokines, adhesion molecules, and other mediators involved in inflammation and stress responses. 11 Dysregulation of NF-κB signaling is strongly associated with chronic inflammatory diseases and pathological immune activation. Importantly, increasing evidence highlights a functional interplay between Nrf2 and NF-κB signaling pathways, which together regulate the balance between oxidative stress and inflammation. 12 This molecular crosstalk is particularly relevant in aging and chronic disease progression, where persistent redox imbalance drives sustained inflammatory activation. Consistent with these mechanisms, comparative studies have demonstrated that AST exhibits higher antioxidant activity than several other carotenoids, including β-carotene and zeaxanthin, particularly in quenching singlet oxygen and inhibiting lipid peroxidation processes.13,14
AST has emerged as a compound capable of modulating both pathways simultaneously. Experimental studies suggest that AST enhances Nrf2-dependent antioxidant defenses while suppressing NF-κB-mediated inflammatory signaling, thereby contributing to the restoration of cellular homeostasis under oxidative stress conditions. 13 This dual regulatory activity positions AST as a promising bioactive molecule in the context of diseases driven by oxidative and inflammatory stress.
Therefore, the aim of this review is to critically analyze the molecular mechanisms through which AST regulates Nrf2 and NF-κB signaling pathways, to explore their functional interaction, and to summarize current experimental and clinical evidence supporting its potential therapeutic applications in oxidative stress- and inflammation-related diseases.
Search strategy
A structured literature search was conducted to evaluate the effects of AST on oxidative stress and inflammatory processes, with a focus on the Nrf2 and NF-κB signaling pathways.
Relevant studies were identified through electronic databases including PubMed/MEDLINE, Scopus, Web of Science, ScienceDirect, and Google Scholar. The search strategy used combinations of keywords such as “astaxanthin,” “Nrf2,” “NF-κB,” “oxidative stress,” and “inflammation,” combined using Boolean operators to optimize retrieval.
The search was limited to peer-reviewed articles published between 2014 and 2024. Eligible studies included in vitro, in vivo, and clinical investigations reporting original data on the biological effects of AST in oxidative stress and inflammation-related conditions. Only English-language publications were considered. Studies were excluded if they were duplicates, non-peer-reviewed, conference abstracts, or lacked sufficient methodological detail. Titles and abstracts were screened initially, followed by full-text evaluation for eligibility. The selected studies were synthesized thematically, focusing on: (i) modulation of Nrf2-mediated antioxidant pathways, (ii) regulation of NF-κB-driven inflammatory signaling, (iii) molecular crosstalk between these pathways in disease contexts characterized by oxidative stress and chronic inflammation.
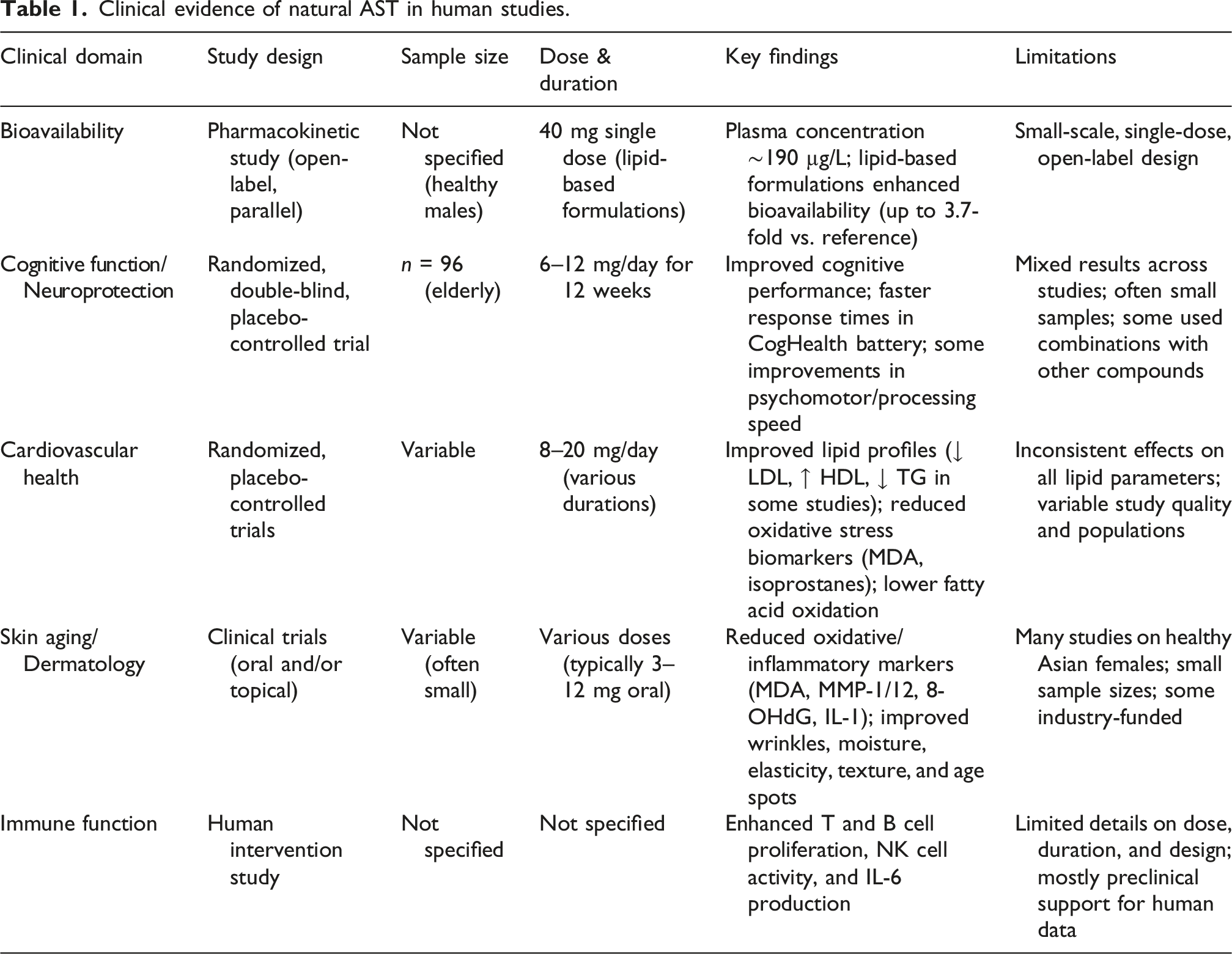
Clinical evidence of natural AST in human studies.
Structural and physicochemical properties of astaxanthin
AST is a xanthophyll carotenoid characterized by a C40 polyene chain with conjugated double bonds and polar terminal ionone rings containing hydroxyl (–OH) and keto (C=O) functional groups. 14 This unique structural arrangement distinguishes AST from other carotenoids and is responsible for its distinctive physicochemical and biological properties.
Chemical structure and electronic properties
The extended structure of conjugated double bonds along the central polyene chain enables efficient delocalization of π-electrons, which is critical for the molecule’s strong antioxidant activity.12–14 This conjugation allows AST to stabilize unpaired electrons and quench reactive oxygen species (ROS), including singlet oxygen and peroxyl radicals, through both physical and chemical mechanisms. 15
The presence of electron-withdrawing keto groups and electron-donating hydroxyl groups at the terminal rings introduces intramolecular polarization, enhancing the redox reactivity of the molecule. 16 These functional groups contribute to a unique charge distribution that facilitates electron transfer reactions and radical scavenging efficiency in lipid-rich environments.
Amphipathic character and membrane orientation
A defining feature of AST is its amphipathic nature, resulting from the combination of polar terminal groups and a hydrophobic polyene chain. 17 This structural configuration enables AST to span lipid bilayers, with the polar ends anchored at the membrane surfaces while the nonpolar chain remains embedded within the hydrophobic core.
This “transmembrane bridge” orientation enhances membrane stability by reducing lipid peroxidation both at the surface and within the inner bilayer region. Consequently, AST is particularly effective in protecting biological membranes against oxidative damage induced by free radicals and inflammatory mediators. 18
Antioxidant mechanisms
Astaxanthin exerts antioxidant activity through multiple complementary mechanisms. First, it directly scavenges reactive oxygen and nitrogen species via electron donation or radical adduct formation. Second, it physically quenches singlet oxygen by dissipating excess energy as heat without undergoing structural degradation. 19
Compared with other carotenoids such as β-carotene and lutein, AST demonstrates superior antioxidant capacity due to its more extensive conjugated system and polar functional groups, which enhance its stability and reactivity in biological systems.12–14
Stereochemistry and natural versus synthetic forms
Astaxanthin exists in several stereoisomeric forms, including (3S, 3′S), (3R, 3′R), and meso (3R, 3′S) configurations. 20 Natural AST, primarily derived from Haematococcus pluvialis, is predominantly composed of the (3S, 3′S) isomer and is often esterified with fatty acids, which may enhance its stability and bioavailability. 21 In contrast, synthetic AST is produced as a racemic mixture of stereoisomers and is typically non-esterified. 22 These structural differences may influence intestinal absorption, tissue distribution, and biological activity, although definitive clinical comparisons remain limited.
Bioavailability and formulation aspects
Astaxanthin is lipophilic and exhibits limited aqueous solubility, which affects its oral bioavailability. Its absorption is strongly influenced by dietary lipids and formulation strategies. 23 Lipid-based delivery systems, including emulsions and softgel formulations, have been shown to significantly enhance systemic uptake. 24 Additionally, esterified forms of AST may undergo enzymatic hydrolysis in the gastrointestinal tract, further influencing absorption kinetics and bioactivity. These factors are critical for translating in vitro potency into in vivo efficacy.
Functional implications
The combination of structural features—extended conjugation, amphipathic character, and stereochemical variability—explains the broad biological activity of AST. These properties allow AST to interact efficiently with cellular membranes, reduce lipid peroxidation, and modulate redox-sensitive signaling pathways. 25 Importantly, these physicochemical characteristics provide the mechanistic basis for astaxanthin’s ability to influence intracellular signaling cascades such as Nrf2 and NF-κB, which will be discussed in the following sections.
Astaxanthin and NRF2 signaling pathway
AST has been widely investigated for its ability to modulate cellular redox homeostasis through activation of the nuclear factor erythroid 2-related factor 2 (Nrf2) signaling pathway, a central regulator of antioxidant and cytoprotective responses. 26 Under basal conditions, Nrf2 is sequestered in the cytoplasm by Kelch-like ECH-associated protein 1 (Keap1), which promotes its ubiquitination and proteasomal degradation. Upon exposure to oxidative stress or electrophilic compounds, conformational changes in Keap1 allow Nrf2 stabilization and nuclear translocation, where it binds antioxidant response elements (AREs) to initiate transcription of detoxifying and antioxidant genes. 27
Astaxanthin has been reported to interfere with this regulatory system through multiple upstream signaling pathways, leading to enhanced Nrf2 activation and downstream cytoprotective responses. One of the primary effects of AST is the reduction of intracellular reactive oxygen species (ROS), which indirectly modulates the Keap1–Nrf2 interaction by altering the cellular redox state. 28
Activation of Nrf2 downstream antioxidant genes
Once activated, Nrf2 induces the expression of a wide range of phase II detoxifying and antioxidant enzymes, including heme oxygenase-1 (HO-1), NAD(P)H quinone dehydrogenase 1 (NQO1), glutamate-cysteine ligase catalytic subunit (GCLC), and glutathione S-transferases (GSTs). 29 These enzymes collectively contribute to the maintenance of intracellular glutathione levels, detoxification of electrophiles, and suppression of oxidative damage.
Experimental evidence suggests that AST enhances the expression of HO-1, which plays a crucial cytoprotective role through the degradation of heme into biliverdin, free iron, and carbon monoxide, molecules with antioxidant and anti-inflammatory properties. 30 HO-1 induction has also been associated with attenuation of inflammatory signaling and modulation of immune responses, linking Nrf2 activation directly to anti-inflammatory outcomes.
Upstream signaling pathways involved in Nrf2 activation by astaxanthin
Astaxanthin-mediated activation of Nrf2 is not limited to direct redox modulation but also involves multiple upstream kinase signaling cascades. Among these, the phosphoinositide 3-kinase/protein kinase B (PI3K/Akt) pathway has been shown to facilitate Nrf2 nuclear translocation by inhibiting GSK-3β-mediated Nrf2 degradation. 31
Similarly, mitogen-activated protein kinase (MAPK) pathways, including ERK, JNK, and p38 MAPK, have been implicated in AST-induced Nrf2 activation through phosphorylation events that enhance Nrf2 stability and transcriptional activity. 32 Additionally, AMP-activated protein kinase (AMPK), a key metabolic sensor, may contribute to Nrf2 regulation by linking energy homeostasis with redox signaling. 33
Evidence from experimental models
Multiple in vitro and in vivo studies support the role of AST in activating Nrf2 signaling under oxidative stress conditions. In neuronal models, AST has been shown to attenuate oxidative injury and mitochondrial dysfunction through activation of the Nrf2/ARE pathway. 34 Similarly, in models of hepatic and renal injury, AST administration leads to upregulation of Nrf2-dependent antioxidant enzymes and suppression of oxidative damage markers. 35
In diabetic and metabolic disease models, AST improves insulin sensitivity and reduces oxidative stress by enhancing Nrf2 signaling and restoring glutathione homeostasis. 36 In retinal and neuronal cells exposed to hyperglycemic or oxidative conditions, AST activates the PI3K/Akt/Nrf2 axis, resulting in increased expression of endogenous antioxidant defenses. 37
Nrf2-mediated anti-inflammatory effects
Beyond its antioxidant role, Nrf2 activation exerts significant anti-inflammatory effects. Nrf2 can suppress inflammatory gene expression indirectly by reducing ROS levels and directly through interference with NF-κB signaling pathways.
38
This dual function positions Nrf2 as a central mediator of redox-inflammatory balance. A schematic overview of the dual regulatory role of AST in modulating Nrf2 and NF-κB signaling pathways is shown in Figure 2. Mechanistic overview of astaxanthin-mediated modulation of inflammation through NF-κB and Nrf2 signaling pathways. Astaxanthin regulates inflammatory responses by inhibiting NF-κB activation while simultaneously enhancing Nrf2-dependent antioxidant defense mechanisms. This dual regulatory effect contributes to reduced oxidative damage and suppression of pro-inflammatory gene expression. The diagram also illustrates the role of these pathways in different stages of carcinogenesis, reflecting the multifactorial involvement of redox signaling in disease progression.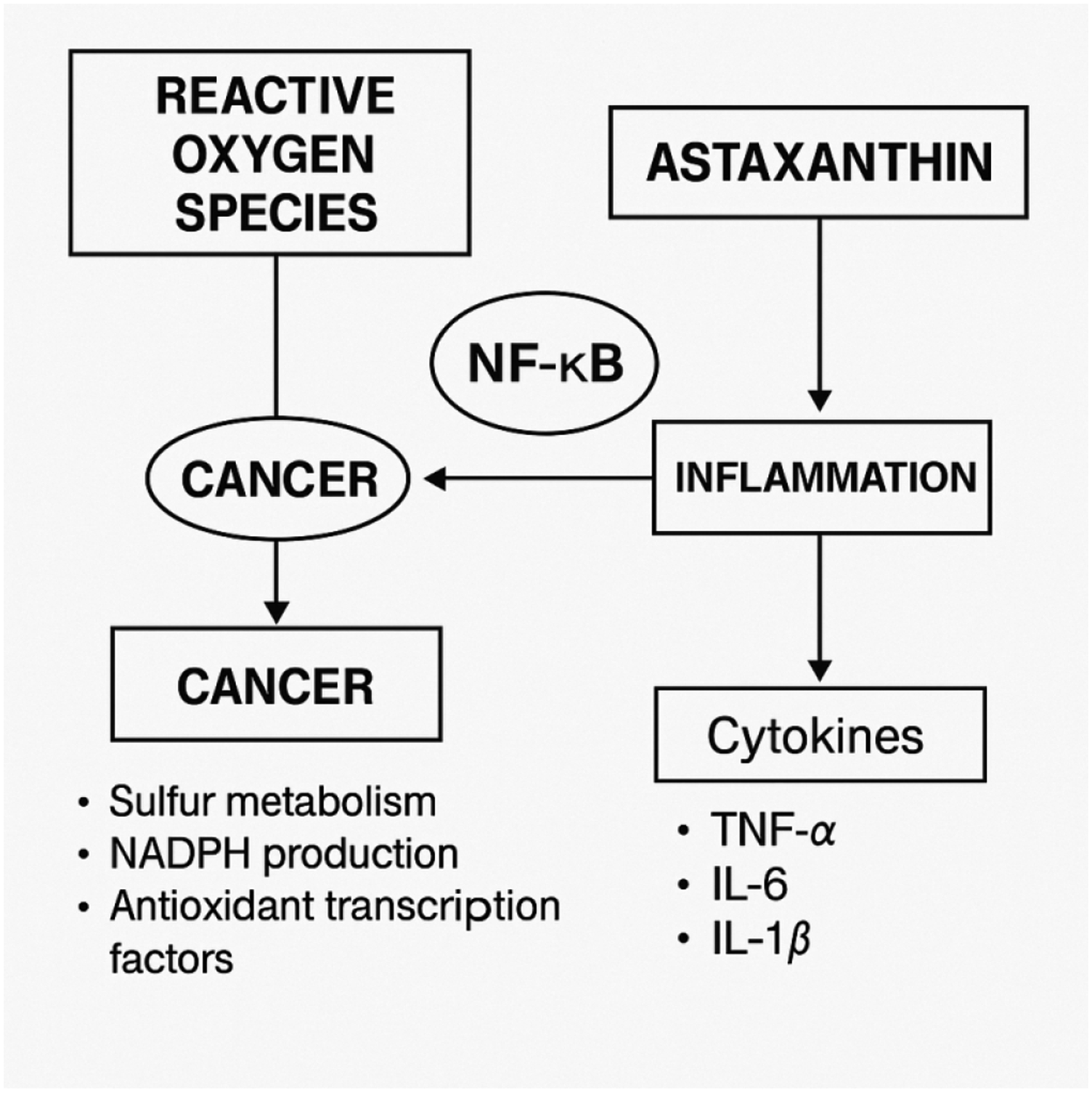
Astaxanthin-induced Nrf2 activation has been associated with reduced production of pro-inflammatory cytokines and attenuation of tissue inflammation in multiple disease models. 39 For example, in lung injury and emphysema models, AST-mediated Nrf2 activation results in decreased inflammatory infiltration and improved tissue integrity. 40
Clinical and translational relevance
The activation of Nrf2 signaling by AST has also been supported by translational and preclinical evidence suggesting protective effects in cardiovascular, neurodegenerative, and metabolic disorders. In models of atherosclerosis and cardiovascular dysfunction, AST reduces oxidative stress markers and improves endothelial function via Nrf2-dependent mechanisms. 41
In neurodegenerative contexts, Nrf2 activation by AST contributes to reduced neuroinflammation, improved cognitive performance, and protection against oxidative neuronal damage. 42 These findings highlight the potential of AST as a dietary bioactive compound capable of modulating endogenous defense pathways relevant to human health.
Summary of Nrf2 modulation by astaxanthin
Overall, AST regulates the Nrf2 pathway through a combination of mechanisms, including modulation of ROS levels, activation of upstream kinase pathways (PI3K/Akt, MAPK, and AMPK), stabilization of Nrf2 protein, and induction of downstream antioxidant genes. This coordinated activation results in enhanced cellular defense capacity and suppression of oxidative stress-induced damage.
Importantly, the Nrf2 pathway does not act in isolation but is functionally interconnected with inflammatory signaling networks, particularly NF-κB, which will be discussed in the following section.
Astaxanthin and NF-ΚB signaling pathway
Nuclear factor kappa B (NF-κB) is a central transcription factor that regulates inflammatory, immune, and stress-responsive gene expression. 43 In most resting cells, NF-κB dimers (commonly p65/p50) are retained in the cytoplasm through interaction with inhibitor of κB (IκB) proteins. Upon stimulation by pro-inflammatory signals, IκB is phosphorylated by the IκB kinase (IKK) complex, leading to its ubiquitination and degradation, thereby allowing NF-κB translocation into the nucleus and activation of target genes. 44
These target genes include a broad spectrum of inflammatory mediators such as tumor necrosis factor-alpha (TNF-α), interleukin-6 (IL-6), interleukin-1β (IL-1β), chemokines, adhesion molecules, cyclooxygenase-2 (COX-2), and inducible nitric oxide synthase (iNOS), all of which contribute to the amplification and maintenance of inflammatory responses. 45
Oxidative stress as an upstream activator of NF-κB
Reactive oxygen species (ROS) act as key upstream activators of NF-κB signaling. Oxidative stress can promote IKK activation and enhance NF-κB nuclear translocation, thereby linking redox imbalance directly to inflammatory gene expression. 46 This ROS–NF-κB axis is particularly relevant in chronic inflammatory diseases where oxidative stress and inflammation form a self-sustaining loop. 47
Astaxanthin, due to its strong antioxidant capacity, interrupts this loop by reducing intracellular ROS levels, thereby indirectly attenuating NF-κB activation. 48 This represents one of the primary mechanisms through which AST exerts anti-inflammatory effects at the transcriptional level.
Direct modulation of NF-κB signaling by astaxanthin
Beyond its antioxidant effects, AST has been shown to directly modulate NF-κB signaling components. Experimental studies indicate that AST inhibits phosphorylation and degradation of IκBα, thereby preventing nuclear translocation of NF-κB p65 subunit. 49
In addition, AST interferes with upstream signaling pathways such as MAPK (ERK, JNK, p38) and TLR4/MyD88-dependent signaling, both of which contribute to NF-κB activation in response to inflammatory stimuli. 50 Inhibition of these pathways results in reduced transcription of pro-inflammatory cytokines and mediators.
Suppression of pro-inflammatory mediators
Astaxanthin-mediated inhibition of NF-κB signaling leads to a marked reduction in inflammatory mediators. Studies have demonstrated decreased expression of TNF-α, IL-6, IL-1β, COX-2, and iNOS in various in vitro and in vivo models following AST treatment. 51 For example, in activated microglial cells, AST suppresses NO production and downregulates COX-2 and iNOS expression through inhibition of NF-κB and associated ERK signaling pathways. 52 Similarly, in macrophage models, AST reduces inflammatory cytokine production through NF-κB-dependent and independent mechanisms. 53
Organ-specific anti-inflammatory effects
AST has demonstrated significant anti-inflammatory effects across multiple organ systems. In pulmonary models, AST attenuates acute lung injury and sepsis-induced inflammation through inhibition of MAPK/NF-κB signaling pathways. 54 In renal injury models, AST suppresses TLR4/MyD88/NF-κB axis activation and enhances antioxidant defense via HO-1 induction. 55
In dermatological models, AST reduces inflammatory responses in atopic dermatitis and UV-induced skin inflammation by suppressing NF-κB-mediated signaling cascades. 56 In neurological models, AST protects astrocytes and neuronal cells from inflammation-induced apoptosis through NF-κB inhibition. 57
NF-κB and oxidative stress interplay
NF-κB signaling is tightly interconnected with oxidative stress pathways. ROS can activate NF-κB, while NF-κB can regulate the expression of antioxidant enzymes such as MnSOD and thioredoxin, forming a bidirectional regulatory network. 58 This crosstalk is essential for balancing cellular survival and inflammatory responses.
AST influences this network by simultaneously reducing ROS levels and modulating NF-κB transcriptional activity. This dual action helps restore redox-inflammatory homeostasis, particularly in chronic diseases characterized by persistent oxidative stress. 59
Interaction with Nrf2 signaling
A critical aspect of astaxanthin’s anti-inflammatory mechanism is its ability to modulate the crosstalk between NF-κB and Nrf2 pathways. Activation of Nrf2 leads to increased antioxidant capacity, which suppresses ROS-mediated NF-κB activation. Conversely, inhibition of NF-κB reduces inflammatory stress that would otherwise suppress Nrf2 activity. 60 AST appears to restore this balance by upregulating Nrf2 while simultaneously inhibiting NF-κB signaling, thereby breaking the pathological feedback loop between oxidative stress and inflammation. 61
Clinical and translational evidence
Clinical and preclinical studies support the anti-inflammatory potential of AST in human-relevant systems. Supplementation with AST has been associated with reduced markers of oxidative stress and inflammation in humans. 62 In cardiovascular and metabolic disorders, AST improves lipid profiles and reduces inflammatory burden through NF-κB-related mechanisms. 63 Furthermore, in neurological and cognitive studies, AST supplementation has been linked to improved cognitive performance and reduced neuroinflammatory markers, suggesting NF-κB modulation as a key mechanism.
Summary of NF-κB modulation by astaxanthin
Overall, AST inhibits NF-κB signaling through multiple converging mechanisms: suppression of ROS production, inhibition of IKK activation, stabilization of IκBα, interference with MAPK and TLR4/MyD88 pathways, and modulation of NF-κB–Nrf2 crosstalk. These effects collectively lead to reduced expression of pro-inflammatory genes and restoration of inflammatory homeostasis.
Crosstalk between NRF2 and NF-ΚB signaling pathways
The Nrf2 and NF-κB signaling pathways represent two major regulatory systems that govern cellular responses to oxidative stress and inflammation. Although they were historically studied independently, increasing evidence demonstrates that these pathways are tightly interconnected and functionally interdependent in maintaining cellular homeostasis. 64
A detailed overview of the transcriptional and post-translational mechanisms governing Nrf2–NF-κB crosstalk is shown in Figure 3. Molecular mechanisms underlying Nrf2–NF-κB crosstalk at transcriptional and protein stability levels. The interaction between Nrf2 and NF-κB occurs through multiple regulatory layers, including competition for transcriptional coactivators, modulation of upstream kinases, and redox-dependent control of protein stability. This bidirectional crosstalk contributes to the balance between antioxidant defense and inflammatory signaling, and represents a key target modulated by astaxanthin in redox-associated disease states.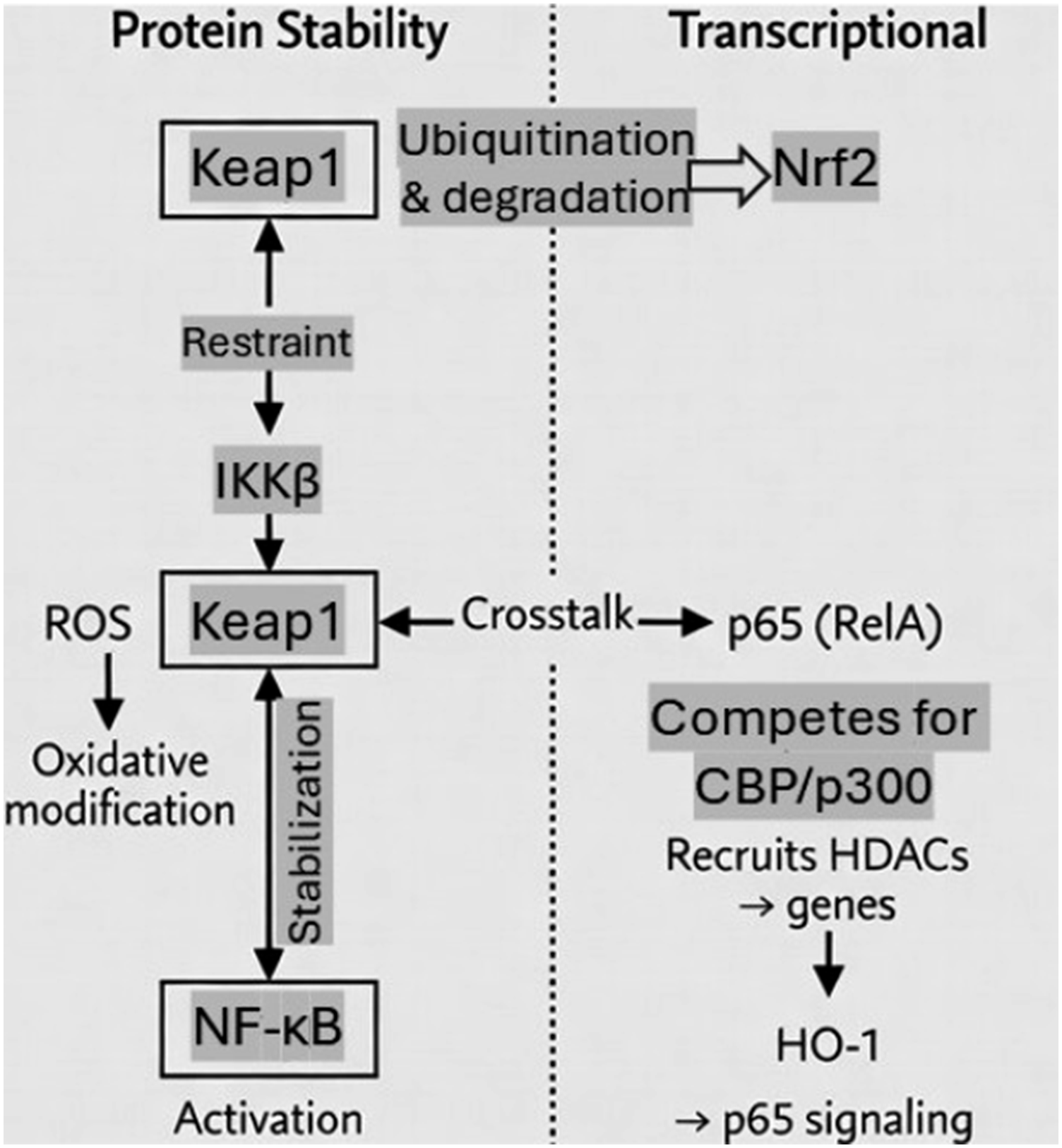
Functional antagonism between Nrf2 and NF-κB
A central feature of the Nrf2–NF-κB interaction is their functional antagonism. Activation of Nrf2 enhances cellular antioxidant capacity through induction of cytoprotective genes such as HO-1, NQO1, and glutathione-related enzymes, thereby reducing intracellular reactive oxygen species (ROS) levels. 65 Since ROS are key activators of NF-κB signaling, Nrf2 activation indirectly suppresses NF-κB activation by limiting oxidative stress–driven inflammatory signaling. 66
Conversely, sustained activation of NF-κB can suppress Nrf2 signaling by competing for transcriptional coactivators such as CREB-binding protein (CBP), thereby reducing Nrf2-mediated transcriptional activity. 67 This reciprocal inhibition establishes a tightly regulated balance between antioxidant defense and inflammatory responses.
Molecular mechanisms of pathway interaction
At the molecular level, the crosstalk between Nrf2 and NF-κB occurs through several interconnected mechanisms. One major regulatory node is the Keap1–Nrf2 system, which is highly sensitive to redox changes induced by NF-κB-mediated inflammation. 68 During chronic inflammation, excessive NF-κB activation leads to elevated ROS production, which can initially activate Nrf2; however, prolonged inflammatory signaling may result in Nrf2 dysfunction or depletion of antioxidant capacity. 69
Another important mechanism involves heme oxygenase-1 (HO-1), a downstream target of Nrf2. HO-1 exerts anti-inflammatory effects by inhibiting NF-κB signaling and reducing pro-inflammatory cytokine production. 70 This creates a feedback loop in which Nrf2 activation suppresses NF-κB-driven inflammation, reinforcing cellular protection.
Role of astaxanthin in restoring redox-inflammatory balance
AST plays a dual regulatory role in modulating the Nrf2–NF-κB axis. On one hand, AST activates Nrf2 signaling, enhancing antioxidant defense systems and restoring glutathione homeostasis. On the other hand, AST suppresses NF-κB activation by inhibiting upstream inflammatory signaling pathways, including ROS generation, MAPK cascades, and TLR4/MyD88 signaling. 71 This dual action allows AST to restore the balance between oxidative stress and inflammation. By simultaneously enhancing Nrf2 activity and inhibiting NF-κB signaling, AST effectively breaks the pathological feedback loop that drives chronic inflammatory diseases. 72
Pathophysiological relevance of Nrf2–NF-κB imbalance
Dysregulation of the Nrf2–NF-κB axis is a common feature of many chronic diseases, including cardiovascular disorders, neurodegenerative diseases, metabolic syndrome, cancer, and inflammatory lung diseases. 73 In these conditions, excessive NF-κB activation and insufficient Nrf2 response lead to sustained oxidative stress, persistent inflammation, and progressive tissue damage. Restoration of balance between these pathways has therefore emerged as a promising therapeutic strategy. Natural compounds capable of modulating both Nrf2 and NF-κB, such as AST, are of particular interest due to their multi-targeted mechanisms and favorable safety profiles. 74
Integrated mechanistic model
Based on current evidence, AST can be positioned as a bidirectional regulator of redox and inflammatory signaling. Its effects can be summarized as follows: - Reduction of intracellular ROS levels, limiting NF-κB activation; - Activation of Keap1–Nrf2 dissociation and nuclear translocation; - Induction of antioxidant enzymes (HO-1, NQO1, GSH-related enzymes); - Inhibition of IKK complex activation and IκB degradation; - Suppression of pro-inflammatory cytokine transcription.
Together, these mechanisms converge to restore cellular homeostasis and reduce the chronic inflammatory state associated with oxidative stress–driven diseases.
Summary of pathway crosstalk
The Nrf2 and NF-κB pathways should not be viewed as isolated signaling systems but rather as an integrated regulatory network governing cellular fate under stress conditions. AST exerts its biological effects precisely at this interface, simultaneously enhancing antioxidant defense and suppressing inflammatory signaling, thereby restoring redox equilibrium and reducing tissue injury.
Clinical evidence and potential therapeutic applications of astaxanthin
AST has progressed from a dietary carotenoid of interest to a bioactive compound evaluated in multiple preclinical and clinical settings. Its therapeutic potential is primarily attributed to its ability to modulate oxidative stress and inflammation through coordinated regulation of Nrf2 and NF-κB signaling pathways, which are central to the pathophysiology of many chronic diseases. 75 Although a substantial proportion of evidence originates from experimental models, an increasing number of human studies support its beneficial effects across metabolic, cardiovascular, neurological, dermatological, and ocular conditions.
Cardiovascular and metabolic diseases
Oxidative stress and chronic low-grade inflammation are key drivers of cardiovascular disease and metabolic syndrome. AST supplementation has been associated with improved lipid profiles, including increased HDL-cholesterol and reduced lipid peroxidation markers in human subjects.
In hyperlipidemic individuals, AST intake has been shown to improve adiponectin levels and reduce oxidative stress biomarkers, suggesting a protective role in endothelial function and systemic inflammation. 76 These effects are consistent with inhibition of NF-κB-mediated inflammatory signaling and activation of Nrf2-dependent antioxidant defenses. 77 In metabolic disorders such as insulin resistance and non-alcoholic fatty liver disease (NAFLD), AST has demonstrated the ability to improve glucose metabolism and reduce hepatic inflammation through modulation of oxidative stress and inflammatory pathways.78–86
Neurodegenerative and cognitive disorders
The brain is particularly vulnerable to oxidative damage due to its high oxygen consumption and lipid-rich environment. AST has shown neuroprotective potential in both experimental models and human studies by reducing oxidative stress and neuroinflammation. 87
Clinical evidence suggests that AST supplementation, alone or in combination with other bioactive compounds, may improve cognitive performance in individuals with mild cognitive impairment. These effects are associated with improved cerebral blood flow, reduced oxidative stress markers, and modulation of inflammatory signaling pathways linked to NF-κB. 88 Preclinical studies further indicate that AST may attenuate age-related cognitive decline by enhancing antioxidant defenses and reducing mitochondrial dysfunction in neural tissues. 89
Ocular health and vision protection
AST accumulates in ocular tissues and has been studied for its protective effects against visual fatigue, retinal damage, and age-related eye disorders. Clinical studies have reported improvements in visual acuity and choroidal blood flow following AST supplementation. 90 In retinal cell models, AST protects against oxidative stress-induced damage through activation of the PI3K/Akt/Nrf2 pathway, highlighting its role in maintaining retinal integrity under stress conditions.91–109
Dermatological effects and skin aging
Skin aging is strongly associated with oxidative stress, inflammation, and extracellular matrix degradation. AST supplementation has been shown to improve skin elasticity, reduce wrinkle formation, and decrease expression of matrix metalloproteinases (MMPs). These effects are linked to reduced NF-κB activation and enhanced antioxidant defense mechanisms, which help protect dermal fibroblasts from UV-induced oxidative damage. 110 Clinical studies further support improvements in skin moisture and structural integrity following AST intake. 111
Respiratory and inflammatory diseases
AST has demonstrated protective effects in inflammatory lung conditions, including acute lung injury and airway inflammation. In both experimental and translational models, AST reduces inflammatory infiltration and suppresses NF-κB-mediated cytokine production. 112
In addition, AST attenuates cigarette smoke-induced emphysema through activation of Nrf2 signaling and suppression of oxidative stress–driven inflammation. 113 These findings highlight its potential role in chronic respiratory disease management.
Renal and hepatic protection
Kidney and liver tissues are highly sensitive to oxidative injury. AST has been shown to protect against toxin-induced renal and hepatic damage through activation of the Nrf2/Keap1/HO-1 axis and suppression of NF-κB-mediated inflammatory responses. 114
In models of drug-induced liver injury, AST reduces oxidative stress and apoptosis, suggesting potential hepatoprotective applications. 115 Similar protective effects have been observed in kidney injury models involving oxidative stress and inflammation.116–121
Safety and bioavailability considerations
Clinical studies indicate that AST is generally well tolerated, with a favorable safety profile even at relatively high supplementation doses.122–127 However, its bioavailability remains a key limitation due to its lipophilic nature. Absorption is significantly enhanced when AST is delivered in lipid-based formulations, emphasizing the importance of formulation strategies for clinical efficacy.128–133 Inter-individual variability in absorption may also influence therapeutic outcomes.
Overall clinical perspective
Collectively, current evidence supports AST as a multi-target bioactive compound with potential benefits in diseases characterized by oxidative stress and inflammation. Its ability to modulate both Nrf2 and NF-κB pathways positions it as a promising adjunct in preventive and supportive therapeutic strategies. However, despite encouraging findings, larger randomized controlled clinical trials are still required to establish standardized dosing regimens, long-term efficacy, and disease-specific therapeutic indications.
Conclusions
Astaxanthin is a marine-derived xanthophyll carotenoid with a unique structural configuration that underpins its strong antioxidant and membrane-stabilizing properties. Its amphipathic nature, extensive conjugated double-bond system, and polar terminal functional groups collectively enable efficient interaction with biological membranes and reactive oxygen species, forming the structural basis for its biological activity. Across the evidence summarized in this review, AST consistently demonstrates the ability to modulate two central signaling networks involved in cellular stress responses: the Nrf2 pathway, which governs antioxidant and cytoprotective defenses, and the NF-κB pathway, which regulates inflammatory gene expression. A key mechanistic feature of AST is its dual regulatory action. On one hand, AST activates the Nrf2/Keap1 axis, promoting nuclear translocation of Nrf2 and induction of downstream antioxidant enzymes such as HO-1 and NQO1, thereby restoring redox homeostasis. On the other hand, AST suppresses NF-κB activation through inhibition of upstream oxidative and inflammatory signaling pathways, including ROS-mediated activation, MAPK cascades, and TLR-related signaling. Importantly, the interplay between these two pathways represents a central mechanism in the pathogenesis of many chronic diseases. The mutual antagonism and feedback regulation between Nrf2 and NF-κB establish a dynamic balance between cellular protection and inflammatory activation. AST appears to restore this balance by simultaneously enhancing antioxidant capacity and reducing inflammatory signaling, thereby interrupting the self-amplifying cycle of oxidative stress and inflammation.
From a translational perspective, preclinical and clinical evidence suggests potential benefits of AST in cardiovascular, metabolic, neurodegenerative, ocular, dermatological, renal, and pulmonary disorders. These effects are largely consistent with its molecular actions on Nrf2 and NF-κB signaling pathways, although variability in study design, dosage, and bioavailability remains a limiting factor for direct clinical translation.
Despite the growing body of evidence, several gaps remain. Future research should focus on well-designed randomized controlled clinical trials, standardized dosing strategies, improved delivery systems to enhance bioavailability, and deeper investigation of tissue-specific mechanisms. In particular, further clarification of direct versus indirect effects of AST on redox-sensitive transcriptional networks would strengthen its mechanistic positioning.
In conclusion, AST represents a promising multi-target bioactive compound that bridges antioxidant and anti-inflammatory signaling networks. Its ability to coordinate Nrf2 activation and NF-κB suppression positions it as a relevant candidate for adjunctive strategies targeting chronic diseases characterized by oxidative stress and inflammation.
Footnotes
Authors contributions
Davide Frumento: Conceptualization, methodology, software, validation, formal analysis, investigation, data curation, writing-original draft preparation, writing-review and editing, visualization, funding acquisition. Ștefan Țălu: Resources, software, funding acquisition, validation, project administration. All authors have read and approved the published version of the manuscript.
Funding
The authors received no financial support for the research, authorship, and/or publication of this article.
Declaration of conflicting interests
The authors declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Data Availability Statement
The data used to support the findings of this study are available from the corresponding authors upon request.