
Abstract
Background
Psoriasis is a chronic immune-mediated inflammatory disease associated with heightened cardiovascular risk. Platelets are increasingly implicated in this link through their capacity to amplify vascular inflammation and interact with leukocytes. Circulating leukocyte–platelet aggregates are elevated in psoriasis, although the biological significance of these aggregates remains incompletely understood. We investigated whether increased leukocyte–platelet aggregates is associated with alterations in the platelet transcriptomic profile in psoriasis.
Methods
Leukocyte–platelet aggregate levels were compared between psoriasis patients (n = 42) and healthy controls (n = 29). Psoriasis patients were stratified by the cohort median lymphocyte-platelet aggregate (LyPA) or neutrophil-platelet aggregate (NPA) levels into high vs low aggregate groups. Platelet RNA sequencing was then performed to define transcriptomic differences in high-vs low-aggregate psoriasis.
Results
Psoriasis patients (mean age 46; 60% male; 81% Caucasian) had higher LyPA (P = 0.001) and NPA (P = 0.04) compared with healthy controls (mean age 42; 55% male; 69% Caucasian). Platelet RNA sequencing revealed that psoriasis patients with high LyPA or high NPA had downregulation of platelet inflammatory pathways, including interferon, tumor necrosis factor (TNF), IL-8, and IL-6 signaling.
Conclusion
These findings identify inflammatory platelet transcriptomic alterations associated with elevated lymphocyte-platelet and neutrophil-platelet aggregates in psoriasis and motivate further work to define the functional consequences of leukocyte-platelet aggregates in psoriasis.
Introduction
Psoriasis is a chronic inflammatory skin disease associated with an elevated risk of cardiovascular events.1-3 Psoriasis pathogenesis involves dysregulated immune signaling, particularly in the interferon, tumor necrosis factor (TNF)-α, and interleukin (IL)-17 pathways, which promote systemic and vascular inflammation.4-7 Platelets are key mediators linking psoriasis to cardiovascular disease. 8 Psoriasis patients exhibit increased biomarkers of platelet activation that correlate with disease severity.9-11 We have previously identified a platelet transcriptomic signature in psoriasis patients that correlated with biomarkers of cardiovascular risk. 8 Beyond their role in atherogenesis and arterial thrombus formation, 12 activated platelets modulate immune responses by directly and indirectly interacting with circulating leukocytes, thereby amplifying systemic and vascular inflammation in psoriasis.13-16
An important immunomodulatory function of platelets is their ability to form aggregates with circulating leukocytes. Upon activation, platelets express P-selectin, which binds to P-selectin glycoprotein ligand-1 (PSGL-1) on leukocytes, promoting the formation of leukocyte-platelet aggregates with downstream effects on leukocyte activation and inflammatory signaling.17-19 Psoriasis patients exhibit increased levels of leukocyte-platelet aggregates.8,20 However, the relationship of leukocyte-platelet aggregates to platelet phenotype and platelet-mediated inflammation in psoriasis remains poorly understood.
Platelets are anucleate cells incapable of de novo transcription; however, they retain megakaryocyte-derived RNA and translational machinery, making platelet RNA sequencing a powerful method to interrogate platelet function and dysregulation in disease states. 21 In this study, we performed platelet RNA sequencing to compare psoriasis patients with high vs low levels of leukocyte-platelet aggregates. We aimed to characterize platelet transcriptomic changes associated with elevated leukocyte–platelet aggregates in psoriasis.
Methods
Study Population
Patients with psoriasis were recruited from New York University (NYU) Langone Health between 2021 and 2024. Eligible participants met one of the following criteria: (1) a history of psoriatic arthritis or moderate to severe psoriasis involving >3% body surface area (BSA) and receiving any systemic therapy (oral or biologic), 22 or (2) active psoriatic disease at the time of enrollment defined as ≥1% BSA of psoriasis confirmed by a board-certified dermatologist or psoriatic arthritis ≥1 swollen/tender joint confirmed by a board-certified rheumatologist. Healthy controls were recruited from the community through general advertising.
Exclusion criteria for both psoriasis patients and healthy controls included a history of prior clinical cardiovascular or cerebrovascular events, known obstructive coronary artery disease at enrollment, other autoimmune diseases beyond psoriasis or psoriatic arthritis, active infection within the past month, non-dermatologic malignancy within the past ten years, chronic kidney disease (creatinine clearance <30 mL/min), active pregnancy, or a history of intravenous drug use. Participants were also excluded if they had poorly controlled hypertension, defined as a systolic blood pressure >160 mmHg at enrollment or requiring treatment with two or more anti-hypertensive medications, or if they had significant burden of diabetes, defined as a hemoglobin A1c >7.0% or requiring insulin therapy. Recruitment methodology has previously been published in detail.8,13
Study Protocol
Participants presented to the Clinical and Translational Research Center at NYU after an overnight fast. Medical history was obtained, and a physical exam was performed by a licensed physician. Clinical data collected included age, sex, race/ethnicity, and self-reported medical history, which was verified through review of the NYU Langone Health medical record and International Classification of Diseases, Tenth Revision coding. Resting blood pressure, height, weight, and waist circumference were measured using standardized protocols. Cardiovascular risk was estimated using the 2013 American College of Cardiology/American Heart Association Pooled Cohort Equation to calculate the 10-year risk of a cardiovascular event. 23 Psoriasis diagnosis and severity were confirmed by a board-certified dermatologist, based on the percent BSA affected and Psoriasis Area and Severity Index (PASI), while psoriatic arthritis diagnosis and severity were assessed by a board-certified rheumatologist using the Classification Criteria for Psoriatic Arthritis (CASPAR).
Laboratory and Platelet Analyses
Blood samples were collected without the use of a tourniquet. Complete blood count and high-sensitivity C-reactive protein (hs-CRP) levels were measured from blood collected into K2 EDTA phlebotomy tubes (BD Vacutainer cat# 367856). High-sensitivity C-reactive protein (hs-CRP) was measured in the NYU clinical laboratory using the Abbott Architect system. Complete blood count was assessed in a research laboratory setting using a Sysmex XN-1000 PR Automated Hematology Analyzer. Research blood samples were processed within 30 minutes of blood draw into appropriate tubes, including 3.2% sodium citrate tubes for platelet studies. Citrated samples were fixed with 1% formalin at a 1:1.2 ratio and processed by hypotonic lysis. Leukocytes were identified by CD45 positive expression (Miltenyi Biotec, item 130-110-638) and characterized into leukocyte subtypes by size, granularity, and, in the case of monocytes, CD14 positivity (Miltenyi Biotec, item 130-110-521). Cells co-expressing CD61 (Agilent Dako, item F0803) were classified as leukocyte–platelet aggregates. Flow cytometry was performed using a Miltenyi MACSQuant Analyzer 10 (Gaithersburg, MD), and results are reported as the percentage of leukocytes positive for adherent platelets, as previously described.8,24
Light Transmission Aggregometry
Light transmission aggregometry (LTA) was performed using a Helena AggRAM light transmission aggregometer (Beaumont, TX) based on the method of Born within 2 hours of blood collection. 25 Platelet aggregation was measured at baseline and following stimulation with collagen (1.0 µg/mL), arachidonic acid (1600 µM) or adenosine diphosphate (ADP) (2.0 µmol/L). Percent aggregation was recorded at 180 and 300 seconds as well as at the point of maximal aggregation. Participants reporting nonsteroidal anti-inflammatory drugs or aspirin use or demonstrating less than 20% aggregation in response to high-dose arachidonic acid (1600 µM) were excluded from platelet aggregation analyses.
Platelet RNA Isolation and Sequencing
Citrated blood was collected and centrifuged to isolate platelet-rich plasma. Acid-citrate-dextrose (ACD) was added at a 1:10 ratio to resting citrated samples, which were then centrifuged at 1000×g for 10 minutes. The resulting pellet was resuspended in EasySep buffer (STEMCELL) and subjected to negative selection using human CD45+ and glycophorin A+. Flow cytometry confirmed a relative platelet-to-leukocyte ratio of approximately 1 × 107. 26 Samples were lysed in TRIzol (Thermo Fisher) and stored at −80°C until RNA extraction. Total RNA was isolated using Direct-zol microspin columns (Zymo Research), and RNA yield, quantity, and integrity were evaluated with an Agilent 2100 Bioanalyzer (Agilent Technologies, Santa Clara, CA). Libraries were generated using the Clontech SMART-Seq HT with Nxt HT kit (Takara Bio USA) and sequenced as single-end reads on an Illumina NovaSeq 6000.
RNA Sequencing Analysis
Sequencing read quality was assessed using FastQC (v0.11.7) 27 and MultiQC (v.1.10.1). 28 Reads were aligned to the human reference genome (hg38) using STAR (v2.6.1d) 29 and read count tables were generated with FeatureCounts (Subread v1.6.3). 30 A relative platelet purity ratio defined by PTPRC/ITGA1B <0.02 was used to verify adequate platelet purity, supported by a principal component analysis demonstrating clear separation driven by combined white blood cell– and platelet-specific biomarkers. Counts were normalized by library size using the DESeq2 R package (v1.34.0). 31 Transcripts with an average log2 normalized count ≥0.75 in at least half the samples were retained for downstream analyses.
Differential expression was determined using DESeq2 while adjusting for age, sex, and biologic therapy use. Multiple hypothesis correction was done using the Benjamini-Hochberg method. Genes with nominal P < 0.01 were considered differentially expressed. Heatmaps of top differentially expressed genes (P < 0.01) were generated with the pheatmap R package (v.1.0.12) 32 on Z-scaled normalized counts, and volcano plots were created with ggplot2 (v3.5.2). 33
Pathway analyses were performed using gene set enrichment analysis (GSEA) 34 implemented in the R fgsea package (v1.20.0), 35 ranking genes by DESeq-derived P-value multiplied by the sign of the log2 fold change. GSEA identified enrichment of annotated pathways using human Molecular Signatures Database (MSigDB) collections including Reactome, Hallmark, and KEGG curated gene sets. Pathways with adjusted P < 0.05 were considered significantly enriched. Upstream cytokine regulators were identified using Ingenuity Pathway Analysis (Qiagen Bioinformatics) using a cutoff P < 0.05.
Statistical Analysis
Data are presented as mean ± standard deviation for normally distributed variables or median [interquartile range, IQR] for non-normally distributed variables. Categorical variables were compared using Fisher’s exact or Chi-square tests, and continuous variables were compared using two-sample t-tests or Mann-Whitney U tests, as appropriate. Comparisons among more than two groups were performed using either ANOVA or the Kruskal-Wallis test. Platelet aggregation on light transmission aggregometry measured at multiple time points were compared using linear mixed-effects models to account for repeated measures of platelet aggregation. Separate models were fit for each agonist. Mixed-effects modeling was performed using R with the lme4 (v 1.1.38) and lmerTest (v 3.2.0) packages. The significance of between-group differences was assessed using ANOVA.
Graphs display individual measurements as boxplots with overlaid jittered points. A two-sided P-value <0.05 was considered statistically significant unless stated otherwise. All statistical analyses were performed in RStudio (version 4.3.0).
Results
Baseline Demographic and Clinical Characteristics of Psoriasis Patients and Healthy Controls
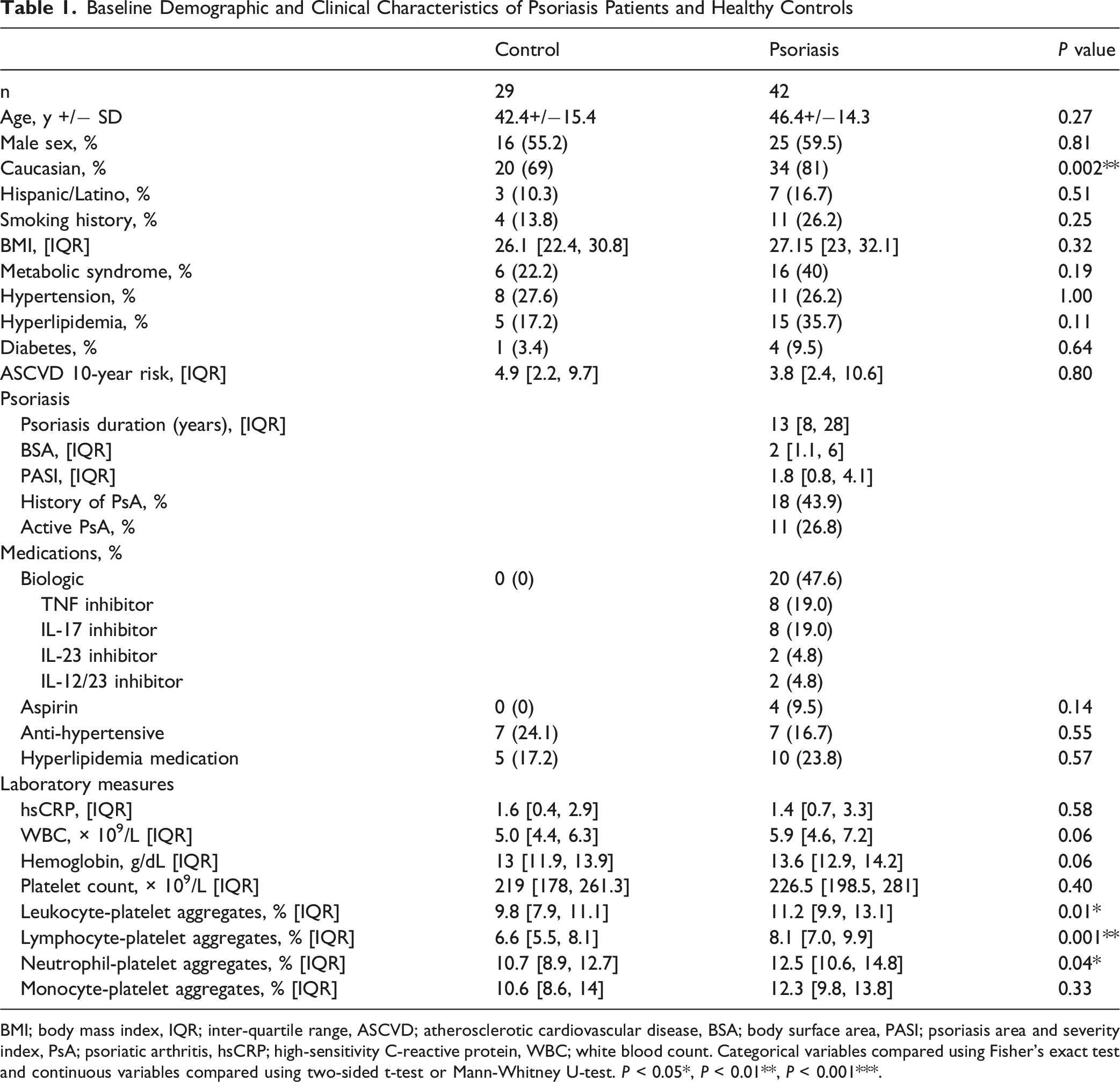
BMI; body mass index, IQR; inter-quartile range, ASCVD; atherosclerotic cardiovascular disease, BSA; body surface area, PASI; psoriasis area and severity index, PsA; psoriatic arthritis, hsCRP; high-sensitivity C-reactive protein, WBC; white blood count. Categorical variables compared using Fisher’s exact test and continuous variables compared using two-sided t-test or Mann-Whitney U-test. P < 0.05*, P < 0.01**, P < 0.001***.
Platelet counts did not differ significantly between psoriasis subjects and healthy controls (Figure 1A). However, psoriasis patients had significantly higher levels of total leukocyte-platelet aggregates (11.2% [IQR 9.9%, 13.1%] vs 9.8% [IQR 7.9%, 11.1%], P = 0.01; Figure 1B). No differences were observed in monocyte-platelet aggregates in psoriasis vs controls (12.3% [IQR 9.8%, 13.8%] vs 10.6% [IQR 8.6%, 14.0%], P = 0.33; Figure 1C), but both lymphocyte-platelet aggregates (LyPA) and neutrophil-platelet aggregates (NPA) were significantly elevated in psoriasis patients compared to healthy controls (LyPA: 8.1% [IQR 7.0%, 9.9%] vs 6.6% [IQR 5.5%, 8.1%], P = 0.001; Figure 1D; NPA: 12.5% [IQR 10.6%, 14.8%] vs 10.7% [IQR 8.9%, 12.7%], P = 0.04; Figure 1E). Cardiovascular risk factors and atherosclerotic cardiovascular disease (ASCVD) 10-year risk scores were similar between the two groups. Leukocyte-platelet aggregates in psoriasis patients vs healthy controls. Comparison between psoriasis (n = 42) and control groups (n = 29) in (A) platelet count, (B) leukocyte-platelet aggregates, (C) monocyte-platelet aggregates, (D) lymphocyte-platelet aggregates with high and low LyPA groups denoted, and (E) neutrophil-platelet aggregates with high and low NPA groups denoted. Data displayed as box-and-whisker plots displaying median, boxes displaying interquartile range (IQR), and whiskers displaying 1.5 IQR. Comparisons were performed using Mann-Whitney U test. P < 0.05*, P < 0.01**, P < 0.001***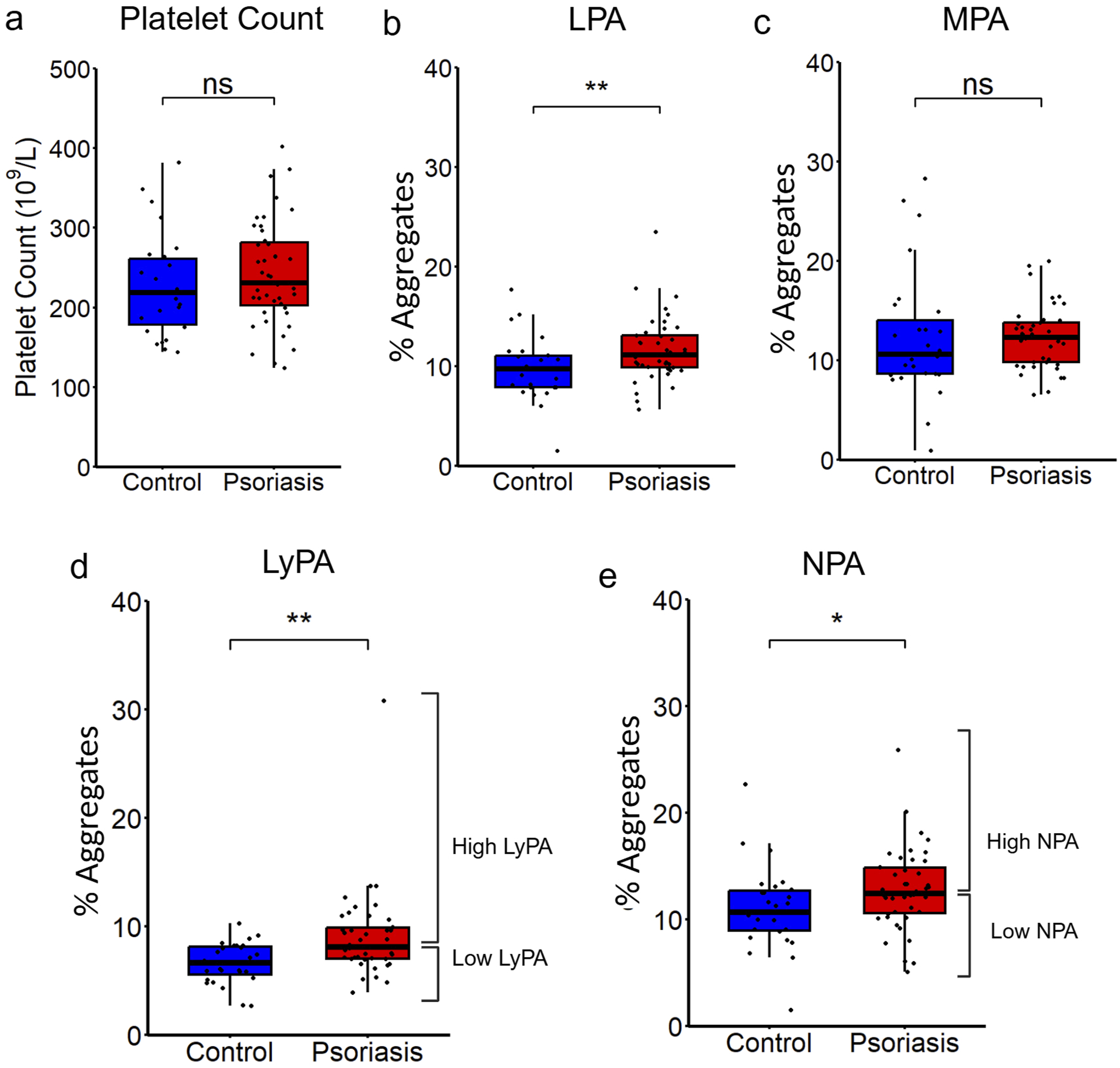
We investigated the effect of systemic biologic therapy on platelet phenotype in patients with psoriasis. Point estimates of leukocyte-platelet aggregate levels were unaffected by biologic therapy use among psoriasis patients (Supplemental Figure S1A-D). Platelet aggregation was measured on light transmission aggregometry (LTA) with adenosine diphosphate (ADP) and collagen stimulation. There was no difference in platelet aggregation to ADP or collagen stimulation in patients on biologic therapy compared to patients off biologic therapy (Supplemental Figure S1E-F).
Comparison of psoriasis patients (n = 30) vs active psoriatic arthritis patients (n = 11, Supplemental Table S1) showed no difference between the groups in leukocyte-platelet aggregates and platelet aggregation to LTA (Supplemental Figure S2).
Psoriasis patients were stratified into high and low LyPA groups based on the psoriasis cohort median. Participants in the high LyPA group had significantly elevated LyPA compared to controls (9.9% vs 6.6%, P < 0.001), while those in the low LyPA group showed no difference from controls (7.0% vs 6.6%, P = 0.97; Supplemental Figure S3A). A similar stratification by median NPA was performed in psoriasis patients such that the high NPA group had significantly elevated NPA compared to controls (14.9% vs 10.7%, P < 0.001), whereas the low NPA group did not differ significantly from controls (10.5% vs 10.7%, P = 0.32; Supplemental Figure S3B). There was a significant association between LyPA and NPA status: individuals with high LyPA were more likely also to have high NPA (P = 0.01; Supplemental Figure S3C).
Demographic and clinical characteristics of psoriasis patients stratified by LyPA and NPA status, as well as healthy controls, are summarized in Supplemental Tables S2 and S3. Among psoriasis participants, those in the high LyPA and high NPA groups had higher platelet counts (high LyPA vs low LyPA: 264 vs 205 × 109/L, P < 0.001; high NPA vs low NPA: 261 vs 212 × 109/L, P < 0.001) and lower rates of hypertension (high LyPA vs low LyPA: 9.5% vs 42.9%, P = 0.03; high NPA vs low NPA: 9.5% vs 42.9%, P = 0.03). No other significant differences were observed between high and low aggregate groups in cardiovascular risk factors, ASCVD risk score, inflammatory markers, psoriasis severity, or disease duration. Patients in the high LyPA and high NPA groups demonstrated a trend of increased platelet aggregation in response to ADP (P = 0.063) and collagen (P = 0.045), respectively (Supplemental Figures S4-S5)
We next performed platelet RNA sequencing on samples from 24 psoriasis patients. Psoriasis patients were stratified by LyPA status: 9 in the high LyPA group and 15 in the low LyPA group (Supplemental Table S4). Differential expression analysis was performed.
In high LyPA compared to low LyPA, 195 genes were differentially expressed after adjusting for age, sex, and biologic therapy (nominal P < 0.01), with 53 genes upregulated and 142 downregulated (Figure 2A-B). Gene set enrichment analysis (GSEA) identified 61 significantly downregulated pathways and no significantly upregulated pathways (adjusted P < 0.05), including the downregulation of key inflammatory pathways such as interferon (IFN) signaling, tumor necrosis factor (TNF) signaling, IL-8, and IL-2 signaling (Figure 2C; Supplemental Table S5). Upstream cytokine regulator analysis predicted reduced activity of IFN-γ, TNF, IL-1β, CD40 ligand and IL-6 (Figure 2D; Supplemental Table S6). Platelet RNA sequencing in psoriasis patients stratified by LyPA. (A) Volcano plot of differentially expressed genes (P < 0.01) in high LyPA (n = 9) vs low LyPA (n = 15). (B) Heatmap of differentially expressed genes in high LyPA vs low LyPA (P < 0.01). Heatmap color displays expression Z-score. (C) Select top differentially expressed pathways in high LyPA vs low LyPA (adj. P < 0.05). (D) Select altered cytokine regulators (P < 0.05) measured by Ingenuity Pathway Analysis in high LyPA vs low LyPA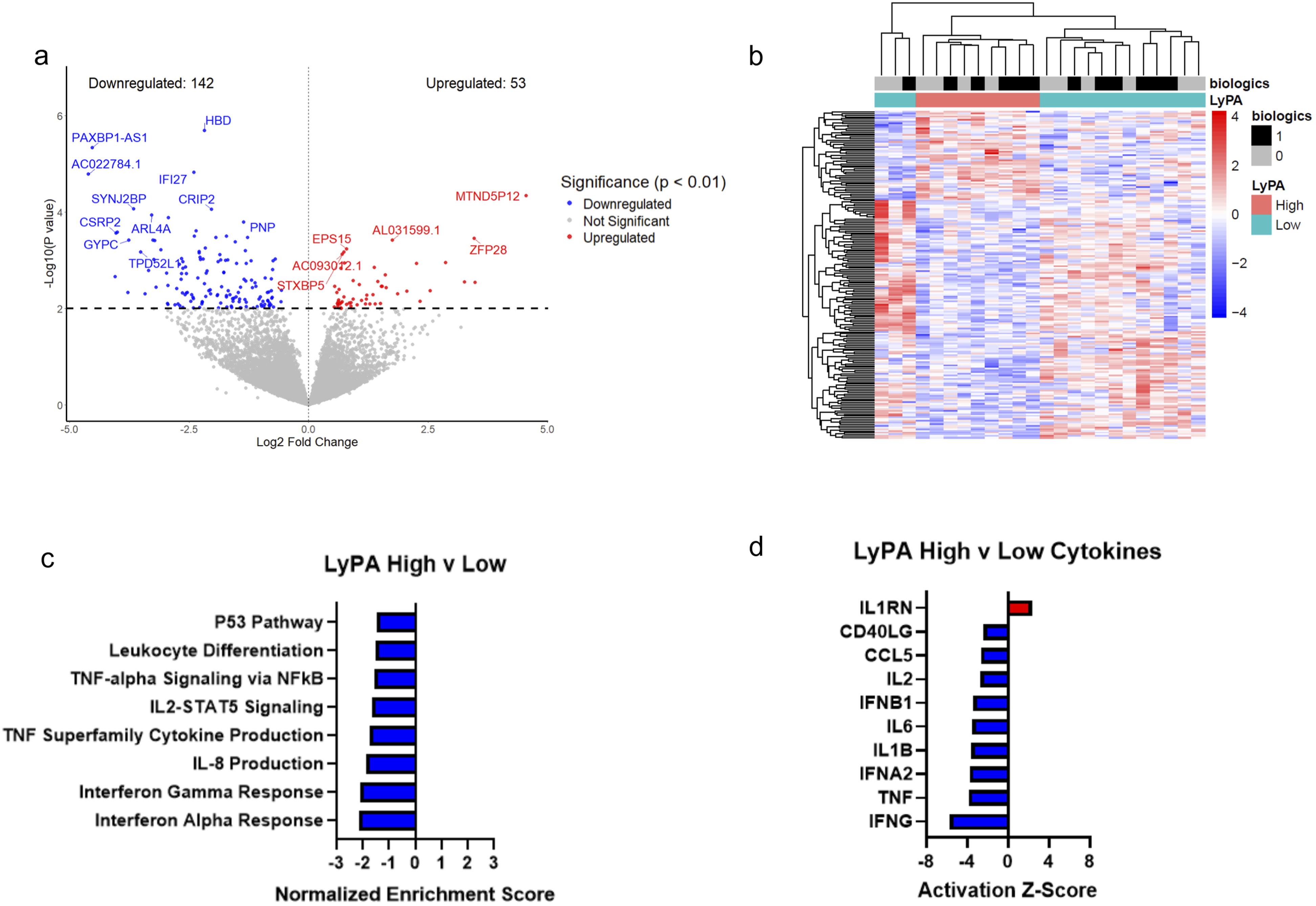
We then performed analogous comparisons for NPA status. In high NPA subjects (n = 8) vs low NPA subjects (n = 16; Supplemental Table S7), 208 genes were differentially expressed (35 upregulated, 173 downregulated; Figure 3A-B). Pathway analysis identified 83 differentially expressed pathways, with 80 downregulated and 3 upregulated pathways. High NPA demonstrated significant downregulation of IFN and TNF signaling pathways as well as IL-8 and IL-6 signaling (Figure 3C; Supplemental Table S8). Upstream regulator analysis predicted reduced activity of IFN-γ, TNF, IL-1β, CD40 ligand, and IL-6 (Figure 3D; Supplemental Table S9). Platelet RNA sequencing in psoriasis patients stratified by NPA. (A) Volcano plot of differentially expressed genes (P < 0.01) in high NPA (n = 8) vs low NPA (n = 16). (B) Heatmap of differentially expressed genes in high NPA vs low NPA (P < 0.01). Heatmap color displays expression Z-score. (C) Select top differentially expressed pathways in high NPA vs low NPA (adj. P < 0.05). (D) Select altered cytokine regulators (P < 0.05) measured by Ingenuity Pathway Analysis in high NPA vs low NPA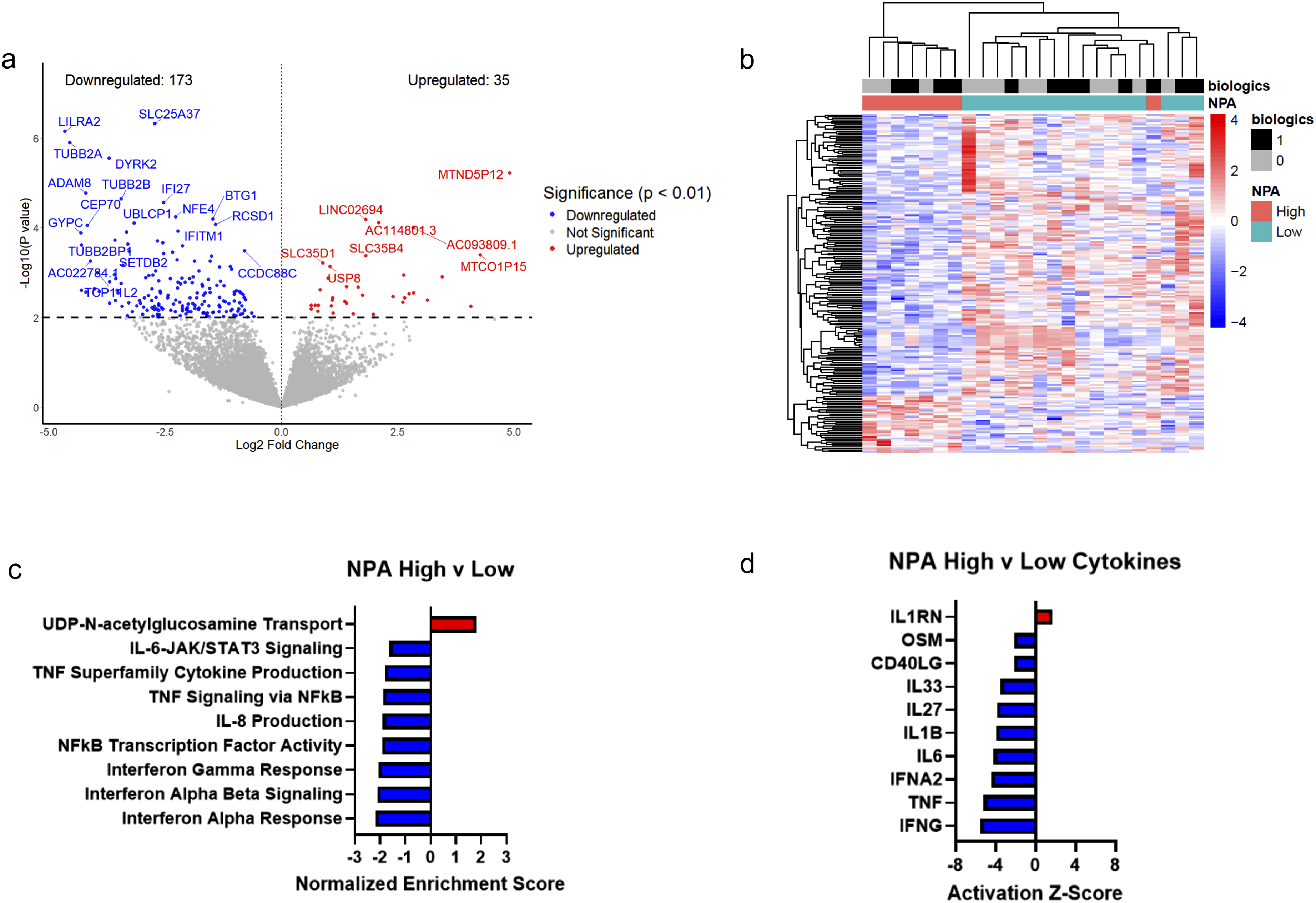
Discussion
In this study, we found that psoriasis patients have elevated levels of LyPA and NPA compared to healthy controls, notably within a psoriasis cohort with well-controlled disease activity. Using platelet RNA sequencing, we identified a transcriptomic profile in psoriasis patients with high LyPA or NPA characterized by downregulation of inflammatory pathways, including IFN and TNF signaling, as well as predicted decreased activity of upstream cytokines including IL-6, TNF, and IL-1β.
We demonstrate an association between high LyPA and high NPA which supports the idea of a shared mechanism driving leukocyte-platelet interactions across leukocyte subtypes in psoriasis. Prior literature demonstrates that leukocyte-platelet aggregates are induced by platelet activation.24,36-38 We show a trend of increased platelet aggregation on LTA in patients with high LyPA and NPA. However, monocyte-platelet aggregates were not different between psoriasis and controls. As monocyte-platelet aggregates are robust biomarkers of platelet activation,17,24 our data suggests that the increased interaction of platelets with lymphocytes and neutrophils in patients with psoriasis may be partially driven by a mechanism independent of platelet activation, consistent with findings in murine models of psoriasis. 39
We and others have previously reported elevated LyPA and NPA in psoriasis.8,20,39 In patients with psoriasis, neutrophils have been shown to form aggregates with platelets, correlating with both skin inflammation and non-calcified coronary plaque burden.39,40 Elevated LyPA has been associated with responsiveness to TNF inhibitor therapy and linked to increased IL-17 production by platelet-bound lymphocytes, suggesting a potential role in modulating disease activity and response to therapy. 20 We extend these findings by demonstrating platelet transcriptional alterations associated with elevated leukocyte-platelet aggregates in psoriasis.
The downregulation of platelet inflammatory pathways is unexpected, as leukocyte-platelet aggregate formation is typically associated with pro-inflammatory activation of both platelets and leukocytes. 37 One plausible explanation is that platelets in the high-aggregate state have increased translation of inflammatory transcripts and secretion of inflammatory mediators, which may result in transcript depletion and apparent downregulation at the RNA level, a phenomenon previously reported with platelet IL-1β in inflammatory states. 41
Alternatively, several recent studies in autoimmune populations have demonstrated that platelet aggregation with T-cells can coincide with reduced T-cell inflammatory signaling.42-44 The effects of leukocyte-platelet aggregation may not be uniformly pro-inflammatory, particularly with lymphocytes. While neutrophil-platelet and monocyte-platelet aggregates are well-studied pro-inflammatory mediators, the effects of lymphocyte-platelet aggregates are poorly understood and may have more complex, context-dependent impacts on leukocyte inflammatory activity. Our finding of downregulated platelet inflammatory pathways in psoriasis patients with high LyPA and NPA may be consistent with prior studies showing that activated platelets in autoimmune contexts aggregate with leukocytes, and platelet aggregation with T-cells results in reduced T-cell inflammatory cytokine signaling including interferon and TNF signaling.43-45 However, our findings raise questions about whether our observed platelet transcriptomic pattern reflects increased translation of inflammatory transcripts or true suppression of platelet inflammatory function, which will require further investigation with targeted functional studies to resolve.
Our study has several limitations. First, the observational and cross-sectional nature of our study precludes causal inference about the directionality of the relationship between platelet transcriptome alterations and platelet-leukocyte aggregation, or if both are driven by a common upstream mechanism. Second, LyPA was measured broadly and did not differentiate among lymphocyte subtypes, preventing us from determining whether the transcriptomic effects are specifically driven by platelet-T-cell interactions. Given the central role of T-cells in psoriasis pathogenesis, future studies using subtype-specific markers will be important to clarify the immunomodulatory role of platelets on T-cells in this context. Finally, we did not fully explore whether our transcriptomic findings were reflected at the protein level.
Overall, our findings suggest alterations in platelet inflammatory phenotype associated with increased LyPA and NPA in psoriasis. Future directions include longitudinal studies of patients with psoriasis over time and over the course of treatment to investigate how platelet-leukocyte aggregates and platelet transcriptomic alterations correspond with treatment and disease course. Studies evaluating inflammatory cytokines and signaling pathways at the protein level are necessary to clarify whether our transcriptomic findings reflect increased translation or true reduction in inflammatory signaling in patients with increased leukocyte-platelet aggregates.
Supplemental Material
Supplemental Material - Lymphocyte-Platelet and Neutrophil-Platelet Aggregates Are Associated With Dysregulated Platelet Inflammatory Pathways in Psoriasis
Supplemental Material for Lymphocyte-Platelet and Neutrophil-Platelet Aggregates Are Associated With Dysregulated Platelet Inflammatory Pathways in Psoriasis by Richard Ni, Isabelle L. Boothman, Filipp Kazatsker, Elliot Luttrell-Williams, Kamelia Drenkova, Florencia Schlamp, Astrid Jara Pernia, Nicole L. Ward, Brittany Weber, Tessa J. Barrett, Jeffrey S. Berger and Michael S. Garshick in Journal of Psoriasis and Psoriatic Arthritis®.
Footnotes
Acknowledgements
The authors wish to thank the NYU Langone Genome Technology Core (GTC) (RRID: SCR_017929) for RNA QC validation, library preparation, and sequencing work. The NYU Langone GTC is partially supported by P30CA016087 (Cancer Center Support Grant). This research is supported in part by a NYU CTSA grant (UL1 TR001445) from the National Center for Advancing Translational Sciences, National Institutes of Health.
Ethical Considerations
This study was approved by the Institutional Review Board of NYU Langone Health (protocol #i20-02001) and conducted in accordance with the principles of the Declaration of Helsinki.
Consent to Participate
Written informed consent was obtained from all participants prior to enrollment.
Author Contributions
Conceptualization: MSG, RN, NLW, JSB; Data Curation: MSG, ELW, FK, KD, FS; Formal Analysis: ILB, RN, FS; Funding Acquisition: MSG, JSB, NLW; Methodology: ILB, ELW, FS; Investigation: RN, ILB, ELW, KD, FS; Visualization: RN, ILB; Resources: MSG, JSB; Supervision: MSG; Writing – Original Draft Preparation: RN; Writing – Review and Editing: ILB, FK, ELW, AJP, KD, FS, NLW, BW, TJB, JSB, MSG
Funding
The authors disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This study was funded in part by awards from the National Institutes of Health, K23HL152013 (MSG), R35HL144993 (JSB), R01HL139909 (JSB), P50AR070590 (NLW), R01AR073196 (NLW), and R01AR084312 (NLW), a non-restrictive Pfizer research grant (MSG), and the National Psoriasis Foundation (MSG, NLW).
Declaration of Conflicting Interests
The authors declared the following potential conflicts of interest with respect to the research, authorship, and/or publication of this article: Dr. Garshick reports consulting fees from BMS, Kiniksa, Argenx, Agepha and unrestricted research support through Pfizer. The other authors have no conflicts of interest to disclose.
Data Availability Statement
All data supporting the findings of this study are available from the corresponding authors upon reasonable request. The RNA sequencing data will be deposited in the Gene Expression Omnibus (GEO) under accession number GSE302674 and will be publicly available upon publication.
Supplemental Material
Supplemental material for this article is available online.
Appendix
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.