
Abstract
Background
Alzheimer's disease (AD) is a neurodegenerative disorder characterized by cognitive decline and behavior impairments. Despite recent approvals of anti-amyloid antibodies, there remains a need for disease modifying and easily accessible therapies. Emerging evidence suggests that targeting the endocannabinoid system may hold promise for AD therapy as it plays a crucial role in different physiological processes, including learning, memory and anxiety, as well as inflammatory and immune responses.
Objective
In this study, we investigated the therapeutic potential of the synthetic cannabinoid WIN 55,212-2 on memory deficits in Tg4-42 transgenic mice.
Methods
Tg4-42 mice were assigned to two treatment groups to investigate the preventive effects of WIN 55,212-2 after a prolonged washout period, as well as the therapeutic effects of WIN 55,212-2 on behavior. Furthermore, the effects of WIN 55,212-2 treatment on AD pathology, including inflammation, amyloid-β load, neurogenesis, and brain glucose metabolism, were evaluated.
Results
Therapeutic WIN 55,212-2 treatment rescued recognition memory and spatial reference deficits in Tg4-42 mice. Furthermore, therapeutic WIN 55,212-2 administration improved motor performance. In addition, preventative WIN 55,212-2 treatment rescued spatial learning and reference memory deficits. Importantly, WIN 55,212-2 treatment did not affect anxiety-like behavior. However, therapeutic and preventative WIN 55,212-2 treatment resulted in an increase locomotor activity and swimming speed in Tg4-42 mice. WIN-treatment reduced microgliosis in the hippocampus of preventively treated mice and rescued brain glucose metabolism in therapeutically treated Tg4-42 mice.
Conclusions
Our findings emphasize the therapeutic promise of the synthetic cannabinoid WIN 55,212-2 in alleviating behavioral and cognitive deficits linked to AD.
Introduction
Alzheimer's disease (AD) is a neurodegenerative condition that is characterized by gradual cognitive decline and neurodegeneration. The range of deficits includes memory loss, confusion, personality changes, language difficulties, motor coordination deficits, and gradual loss of independence. The disease typically initiates with memory complaints. These problems can involve episodic memory and speech production, including naming or semantic difficulties, as well as visual and spatial orientation.1–4
While the precise etiology of AD remains incompletely understood, evidence indicates that amyloid-β (Aβ) peptides are pivotal in its development. 5 In addition to full-length Aβ, various truncated and modified forms of Aβ are known to contribute significantly to the pathogenesis and progression of AD.6,7 Among these variants, Aβ4-42 was one of the earliest documented N-terminally truncated forms. 8 This specific Aβ isoform is detected in abundance in familial and sporadic AD brains, as well as in aged control subjects.9–11
Despite the recent approval of two anti-amyloid monoclonal antibodies lecanemab (Leqembi®) and aducanumab (Aduhelm®), in the US there is still no universal and easily assessable treatment for AD available.12–14 In recent years, there has been increasing interest in exploring the therapeutic potential of modulating the endocannabinoid system (ECS) as a possible treatment approach for various conditions, including AD.15–20 The ECS is a lipid signaling system that consists of cannabinoid receptors, endogenous cannabinoids, and enzymes involved in the synthesis and degradation of endocannabinoids.21,22 Physiologically, the ECS is involved in the homeostasis of numerous body functions, including anxiety, inflammation, neurogenesis, cardiovascular activity, and immune function.23,24 Furthermore, the ECS has a substantial impact on regulating learning and memory functions.25,26
The therapeutic potential of the ECS in AD is based on its potential to impact various processes linked to AD pathogenesis, including inflammation, mitochondrial dysfunction, and excitotoxicity, as well as Aβ and tau metabolism. 24 Both natural and synthetic cannabinoids display anti-inflammatory, antioxidative, analgesic, and anti-proliferative characteristics.27–29 It has been shown that WIN 55,212-2, a synthetic cannabinoid acting as an agonist for CB1 and CB2 receptors, exhibits neuroprotective effects and reduces tau and Aβ pathology as well as neuroinflammation both in vitro and in rodent in vivo studies.30–34
In the current study, we examined how chronic WIN 55,212-2 treatment affects the behavior of Tg4-42 mice. Tg4-42 mice overexpress Aβ4-42 without simultaneously overexpressing human amyloid-β protein precursor (AβPP), nor do they carry mutations associated with autosomal dominant forms of AD. 35 In Tg4-42 mice, the intracellular accumulation of Aβ4-42 is accompanied by microglial and astroglial activation, with the most prominent effects observed in the hippocampus. Additionally, this model demonstrates age-dependent synaptic dysfunction and neuronal loss, despite the absence of plaque formation.36–38 Furthermore, Tg4-42 mice exhibit age-dependent cognitive and behavioral impairments, starting at six months of age. These deficits include deficits in object recognition memory, spatial memory, and motor skills and are associated with a decrease in anxiety-related behavior.37,39–42 In this study, we addressed whether a cannabinoid agonist could ameliorate the cognitive deficits observed in Tg4-42 mice. Furthermore, a preventive treatment strategy was compared to a therapeutic approach.
Methods
Animals and drug treatment
The generation of the Tg4-42 mouse line has been previously described. 35 In brief, Tg4-42 mice express human Aβ4-42 fused to the murine thyrotropin-releasing hormone (TRH) signal peptide under the control of the neuronal Thy-1 promoter. 40 Mice were bred and maintained on a C57Bl6/J genetic background. For this study, homozygous Tg4-42 mice, evenly distributed between males and females, were utilized.
Mice were kept in individually ventilated cages (IVC, 32 × 16 × 14 cm; Tecniplast, Hohenpeißenberg, Germany) in a controlled environment with a 12/12 h light/dark cycle, randomly grouped in up to five. Ad libitum access to water and food was provided. All animals were handled according to the German guidelines and EU legislations for animal care and the experiments were approved by the local authorities (Niedersächsisches Landesamt für Verbraucherschutz und Lebensmittelsicherheit, Röverskamp 5, 26203 Oldenburg, Germany, [16/2364] and Landesamt für Gesundheit und Soziales LAGeSo Darwinstr. 15, 10,589 Berlin [65/18]). All experiments followed the recommendations of the ARRIVE guidelines.
The CB1/CB2 receptor agonist (R)-(+)-[2,3-dihydro-5-methyl-3-(4-morpholinyl-methyl) pyrrolo-[12,3-d,e]-1,4-benzoxazin-6-yl]-1-naphthalenyl-methanone (WIN 55,212-12, Sigma-Aldrich, St Louis, MO, USA) was dissolved in a vehicle solution containing 5% Tween 80 (Carl Roth GmbH, Karlsruhe, Germany), 5% ethanol (100%), and 90% NaCl (0.9%). 22 To analyze the preventive effects of Win 55,212-2, treatment was administered at 3 months before the onset of neuron loss or behavior deficits. 37 In addition, to analyze the therapeutic effects of WIN 55,212-2, Tg4-42 mice were treated with Win 55,212-2 starting at 5 months after the onset of behavior deficits and neuron loss in the hippocampus. 37 Both treatment approaches lasted for six weeks. Mice were allocated randomly to groups receiving either WIN 55,212-2 or vehicle and received daily intraperitoneal injections at a volume of 10 ml/kg. The drug was administered at a dosage of 0.2 mg/kg, and behavioral assessments began for all mice at the age of 6 months. The experimental design is illustrated in Figure 1.
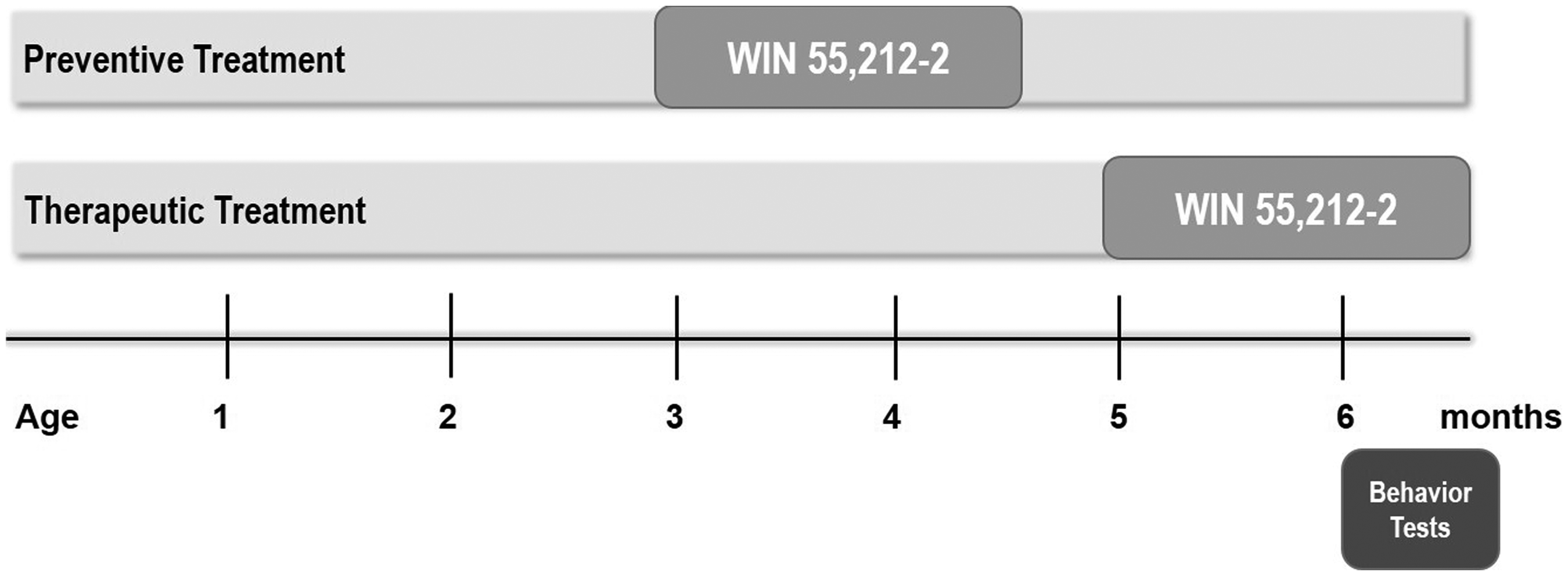
Schematic illustration of the experimental setup. Tg4-42 mice received treatment with WIN 55,212-2 or a vehicle solution for 42 consecutive days, starting at 3 months (preventive) or 5 months (therapeutic), respectively. Behavioral assessments started at 6 months.
Behavior assessment
To identify potential cognitive and behavioral benefits of WIN 55,212-2 administration in Tg4-42 mice, a series of anxiety and memory tests were performed. Additionally, wild-type (WT) mice treated with the vehicle solution were tested as controls. Equal numbers of male and female mice were used, and no statistically significant differences between the sexes were observed in any of the tests. All mice underwent behavior analysis at 6 months of age, with testing extending over a period of 15 days (n = 12-16). Mice were maintained under a 12 h/12 h reversed light cycle, with all behavior experiments occurring during the dark phase from 7 a.m. to 7 p.m. 22
Rotarod
Motor coordination, balance and motor learning were assessed using the rotarod (TSE Systems GmbH, Bad Homburg, Germany). 43 The testing protocol involved four trials daily for two consecutive days, with inter-trial intervals lasting 10 to 15 min as previously described. 22 Each mouse was placed individually on the rod, which accelerated from 4 to 40 revolutions per minute (rpm) within a maximum trial duration of 300 s. The duration the mouse spent on the rod (latency to fall) was recorded as a measure of motor performance, and trials concluded either when animals fell off or reached the maximum time limit. 39 Between trials, the apparatus was cleaned with 70% ethanol to remove odor cues.
Open field
Exploratory behavior and spontaneous motor activity were tested in the open field (OF) test 44 as previously described. 22 Mice were observed for a duration of 5 min within a 50 × 50 cm arena with 38 cm high walls. ANY-Maze video tracking software (Stoelting Co, Wood Dale, IL, USA) was used to document both the distance covered by the mice and the duration of time spent in the central area of the arena. To minimize odor cues, the maze was cleaned with 70% ethanol between animals.
Elevated plus maze
The elevated plus maze (EPM) was used to evaluate exploratory behavior and responses related to anxiety.44,45 The apparatus consisted of four arms (5 cm width × 15 cm length) extending at 90° angles from a central zone (5 cm width × 5 cm length) and elevated to a height of 75 cm above a padded surface. 46 The maze consisted of two open arms facing each other and two closed arms surrounded on three sides by a 15 cm high transparent plastic wall. Mice were positioned in the center of the maze, facing an open arm, and allowed to explore freely for 5 min as previously described. 22 Their movements and time spent in each arm were tracked and recorded using ANY-Maze tracking software. Anxiety-like behavior was assessed based on the time spent in the open arms, with longer durations indicating lower anxiety levels. 47 To prevent odor cues, the EPM was cleaned with 70% ethanol between mice.
Dark light box
The dark light box (DLB) served as a tool to assess potential anxiolytic-like or anxiogenic-like effects in mice. 48 This test relies on the natural light aversion and spontaneous exploratory behaviors typically observed in rodents. 44 Conducted within a gray plastic arena measuring 73 cm × 25 cm × 32 cm, the test featured two distinct sections: a smaller dark area (31 cm × 25 cm) and a larger light area (42 cm × 31 cm). A narrow opening (5 cm × 5 cm) in the dividing wall allowed mice to move freely between the two compartments. Each mouse was introduced into the illuminated area, positioned facing the wall, and allowed to explore the space freely for 5 min. ANY-Maze video tracking software (Stoelting Co., Wood Dale, IL, USA) was utilized to document the time spent in each area, the latency to enter the dark area, and the number of crossings between sections. 22
Novel object recognition test
In order to analyze recognition memory and novelty preference, the novel object recognition test (NOR) was executed within a gray plastic open-field arena measuring 50 × 50 cm, as previously described. 22 Twenty-four hours following the OF test, the Novel Object Recognition (NOR) test was conducted in the same arena, featuring two identical objects during the training phase. Mice were given 5 min to freely explore the box. During the testing phase on the subsequent day, mice were reintroduced to the same arena for 5 min, where a familiar object was placed alongside a new object. The new object was similar in height but differed in shape and appearance. Mouse movements were tracked using an automated video tracking system (ANY-maze, Stoelting Co, USA), recording both the duration of exploration for each object and the distance traveled. Between each mouse, objects and the arena were cleaned with 70% ethanol to eliminate lingering scents.
The percentage of time spent exploring the novel object was determined using the following calculation:
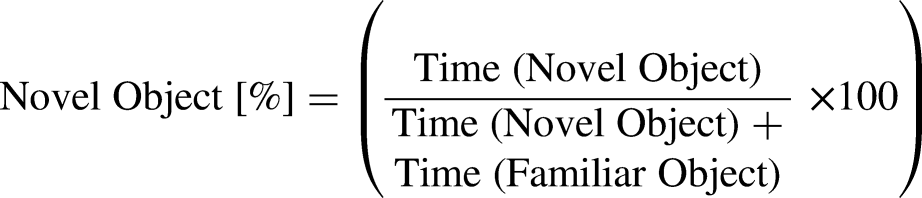
Morris water maze
To assess spatial reference memory, the Morris water maze test was used as previously described.22,35,40 The test uses spatial cues to find a hidden submerged platform (10 cm in diameter) positioned within a circular pool (110 cm in diameter) containing non-transparent tap water. The pool was divided into four distinct virtual quadrants, defined according to their spatial orientation relative to the platform: left (L), right (R), opposite (O), and target (T) quadrant, with the latter containing the goal platform.
Initially, mice were subjected to 3 days of cued training to assess their swimming capability and eyesight. Throughout the cued training, a triangular flag was employed to indicate the platform's position and mice were given 60 s to locate the submerged platform. Notably, both the starting point and platform location changed between the trials. Each day consisted of four cued training trials with an average interval of 15 min between trials.
Twenty-four hours following the completion of the cued training, mice underwent 5 days of acquisition training, consisting of 4 trials per day. During this training, proximal visual cues were affixed to the outside of the pool, and the flag indicating the platform location was removed. The platform remained fixed in place throughout the acquisition training, with mice placed into the pool from one of four predetermined entry points. 40
In the concluding probe trial, the platform was removed from the pool, allowing mice to swim freely for 1 min. ANY-Maze video tracking software (Stoelting Co., Wood Dale, USA) was used to record swimming speed, escape latency, and quadrant preference.
Immunohistochemistry
Following fixation of the right hemisphere in 4% buffered formalin at 4°C, the tissue was embedded in paraffin. Immunohistochemistry was conducted on 4 μm sagittal paraffin sections as previously described. 49 In brief, the sections were first deparaffinized in xylene and then rehydrated through a graded ethanol series (100%, 95%, 70%). Endogenous peroxidase activity was blocked by treating the sections with 0.3% H2O2 in PBS. Antigen retrieval was performed by boiling the sections in 0.01 M citrate buffer (pH 6.0), followed by a 3-min incubation in 88% formic acid. To prevent non-specific binding, the sections were treated with skim milk and fetal calf serum in PBS before the application of the primary antibodies: pan-Abeta 24,311 (rabbit-anti-Aβ, 1:500 35 ), GFAP (guinea pig, 1:1000, Synaptic Systems, Germany), and IBA1 (guinea pig, 1:500, Synaptic Systems, Germany). The primary antibody incubation took place overnight at room temperature in a humid chamber. Biotinylated anti-rabbit antibodies were used as secondary antibodies (1:200, Jackson ImmunoResearch, USA). The staining was visualized using the ABC method with a Vectastain kit (Vector Laboratories, Burlingame, USA) and diaminobenzidine (DAB) as the chromogen. Hematoxylin was used for counterstaining for the Aβ staining.
A semi-quantitative analysis of immunoreactivity was performed using images of the CA1 pyramidal layer at 20× magnification on three sections per mouse, with each section spaced at least 30 μm apart. The analysis was conducted using an Olympus BX51 microscope equipped with a digital camera (MoticamPro 282B, Motic, Germany). The images were converted to 8-bit black-and-white images, and a fixed intensity threshold was applied to define the DAB signal. The IBA1- and GFAP-immunoreactivity were then calculated as the percentage of positive DAB staining within the defined region of interest. The Aβ load was manually quantified at 10× magnification, and the experimenter was blinded to the treatment of the mice. The results were normalized to the Vehicle group.
To quantify the number of newborn neurons, a free-floating staining protocol was employed on every 10th coronal frozen section of the left brain hemisphere, each 30 µm thick. Neurogenesis was assessed using the doublecortin (DCX) as a marker for cell proliferation. Therefore, the sections were rehydrated in PBS, and endogenous peroxidase activity was blocked by immersing the sections in PBS containing 0.3% hydrogen peroxide for 30 min. The sections were then washed in PBS with Triton X-100, and non-specific antibody binding was prevented by incubating them in PBS containing 10% fetal calf serum (FCS) and 4% low-fat dry milk powder. The primary antibody against DCX (1:4000, Synaptic Systems, Germany) was incubated overnight, followed by incubation with a biotinylated secondary antibody. Staining was visualized using the ABC method with a Vectastain kit (Vector Laboratories) and DAB as the chromogen. The images were analyzed using an Olympus BX51 microscope equipped with a motorized stage. The total number of newborn neurons in the subgranular zone (SGZ) of the dentate gyrus (DG) was counted using the meander scan option of StereoInvestigator 7.0 (MBF Bioscience) to quantify all DCX-positive cells in each section. The final neuron count was multiplied by 10 to estimate the total number of newborn neurons. 50
Quantification of neuron numbers using unbiased stereology
Neuron numbers in the CA1 region of the hippocampus were quantified using unbiased stereology, as previously described. 49 Seven-month-old mice were anesthetized and transcardially perfused with 4% paraformaldehyde. The left brain hemispheres were then fixed in 4% paraformaldehyde, cryoprotected in 30% sucrose, frozen, and sectioned frontally into 30-μm thick slices using a cryostat (Microm HM550, Germany). Every tenth section was systematically sampled and stained with cresyl violet. Stereological analysis was conducted on the hippocampal CA1 cell layer (Bregma −1.22 to −3.80 mm) utilizing a stereology workstation [Olympus BX51 with a motorized specimen stage for automatic sampling, StereoInvestigator 7 (MicroBrightField, Williston, VT, USA)].
18F-FDG PET/MRI
To evaluate brain glucose metabolism in the hippocampus of 7-month-old Tg4-42 mice therapeutically treated with WIN or vehicle, 18F-fluoro-deoxy-glucose positron emission tomography/magnetic resonance imaging (18F-FDG-PET/MRI) acquisition and analysis were conducted as previously described. 36 Female Tg4-42 mice were fasted overnight, and blood glucose levels were measured. 18F-FDG was administered intravenously into a tail vein with a maximum volume of 200 μl. During the scans, mice were anesthetized with isoflurane supplemented with oxygen, while they were awake during the uptake period. PET images were acquired 45 min after 18F-FDG administration on a small animal 1 Tesla nanoScan PET/MRI system (Mediso, Hungary). Mice were placed on a heated bed maintained at 37°C, and their respiration rate was continuously monitored during the 20-min PET scan. MRI-based attenuation correction was applied (matrix 144 × 144 × 163 with a voxel size of 0.5 × 0.5 × 0.6 mm³, TR: 15 ms, TE: 2.032 ms, flip angle: 25°), and PET images were reconstructed with a matrix of 136 × 131 × 315 and a voxel size of 0.23 × 0.3 × 0.3 mm³. Image analysis was performed using PMOD v3.9 (PMOD Technologies, Switzerland) with a predefined mouse brain atlas template to analyze the hippocampus region (HI). The corresponding PET images were aligned with the MRI, and statistics within the volume of interest (VOI) in kBq/cc was generated. For semi-quantitative analysis, the standardized uptake value (SUV) was calculated [SUV = tissue activity concentration average (kBq/cc) × body weight (g)/injected dose (kBq)], and SUV values were corrected for blood glucose levels [SUVGlc = SUV × blood glucose level (mg/dl)].
Statistical analysis
Statistical analyses were conducted using GraphPad Prism Version 9 (GraphPad Software, San Diego, CA, USA). Group differences were assessed using unpaired t-tests, one-way analysis of variance (ANOVA) followed by Bonferroni multiple comparisons, or two-way ANOVA followed by Bonferroni multiple comparisons, as specified. Data are shown as mean +/- standard deviation. Significance levels are given as follows: *p < 0.05; **p < 0.01; ***p < 0.001, #p < 0.05; ##p < 0.01; ###p < 0.001.
Results
Prolonged WIN 55,212-2 treatment effects motor performance
The motor performance of Tg4-42 mice, subjected to either preventive or therapeutic treatment with WIN 55,212-2, was assessed using the accelerating rotarod task and compared to WT controls. The rotarod task is commonly used to assess motor skill learning and coordination.39,43 Overall, all Tg4-42 mice performed worse than WT control mice, as previously described, 39 regardless of the treatment start (Figure 2; two-way repeated measures ANOVA followed by Bonferroni comparisons, preventative: WT versus Tg4-42Vehicle: p < 0.001; WT versus Tg4-42WIN 55,212-2: p < 0.001; therapeutic: WT versus Tg4-42Vehicle: p < 0.001; WT versus Tg4-42WIN 55,212-2: p < 0.001). However, all Win- and Vehicle-treated Tg4-42 mice improved their ability to stay on the rod over the eight training trials (Figure 2; two-way repeated measures ANOVA followed by Bonferroni comparisons, trial 1 versus 8: WIN preventive: p < 0.05; Vehicle preventive: p < 0.001; WIN therapeutic: p < 0.001; Vehicle therapeutic: p < 0.001). Furthermore, therapeutic WIN-treated Tg4-42 performed significantly better than control mice (Figure 2(c); two-way repeated measures ANOVA: treatment therapeutic: F(1,26) = 4.675, p = 0.040; Bonferroni comparisons: trial 1: p < 0.05; trial 2: p < 0.05). In contrast, preventive Tg4-42 mice performed significantly worse on day 7 (Figure 2(a); Bonferroni comparisons: trial 7: p < 0.05).
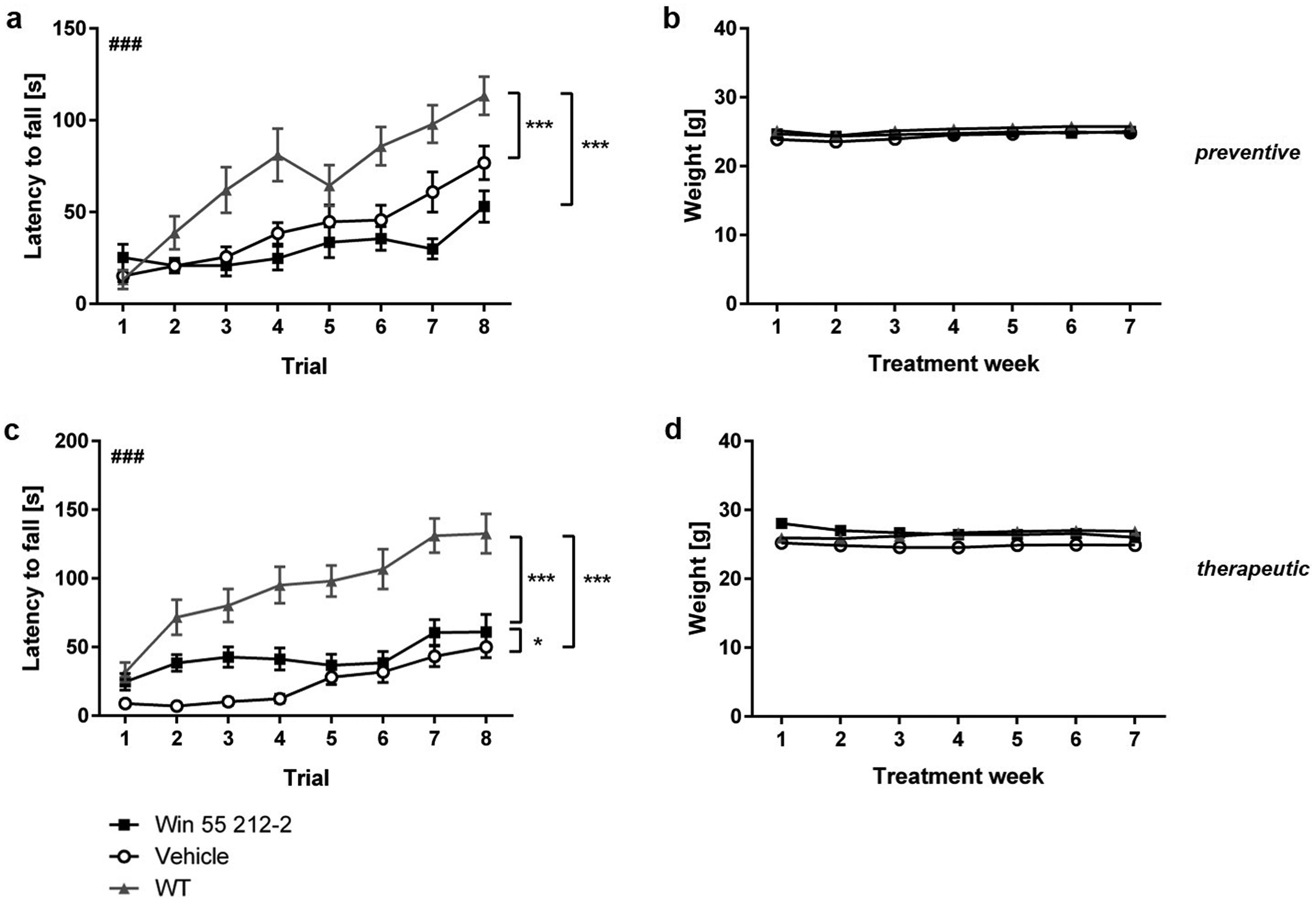
Influence of prolonged WIN treatment on motor performance of Tg4-42 mice. (a) Tg4-42 mice that received preventive WIN 55,212-2 performed worse than control mice on day 7. (c) In contrast, preventively with WIN 55,212-2 treated mice displayed a significantly improved motor performance in the rotarod task. (b,d) The administration of WIN 55,212-2 did not impact the body weight of Tg4-42 mice. Two-way ANOVA followed by Bonferroni multiple comparisons; n = 13-16. Data presented as mean ± S.E.M; *p < 0.05.
Prolonged WIN 55,212-2 treatment does not affect body weight of Tg4-42
Mice treated preventively or therapeutically with WIN 55,212-2 exhibited comparable weights to control Tg4-41 and WT mice over the 7 weeks of treatment (Figure 2(b), (d); two-way repeated measures ANOVA, treatment preventive: F(2,44) = 0.2593, p = 0.7728; treatment therapeutic: F(2,41) = 0.8608, p = 0.4303).
Therapeutic WIN 55,212-2 treatment rescues recognition memory in Tg4-42 mice
Recognition memory of Tg4-42 was assessed using the NOR. Vehicle-treated Tg4-42, independent of the treatment start, demonstrated impaired recognition memory as they were not able to differentiate between the novel (N) and familiar (F) object (Figure 3(c),(g). Similarly, preventive WIN-treated Tg4-42 exhibited impaired recognition memory, evident from their absence of preference for the novel object (Figure 3(c)).

Influence of prolonged WIN treatment on novel object recognition memory in Tg4-42 mice. (a,e) All mice spent an equal amount of time interacting with the two identical objects (O1, O2) during the training phase. During the testing phase, only (g) therapeutically treated Tg4-42 displayed a significant preference for the novel object (N). Control Tg4-42 and (c) preventive WIN-treated Tg4-42 were unable to differentiate between the novel (N) and the familiar object (F). Distance traveled, regardless of treatment start, was not affected by WIN treatment during the (b,f) training and testing phase (d,e). The dashed line represents the 50% chance level. Two-way ANOVA followed by Bonferroni multiple comparisons; n = 11-16. Blue dots represent male mice, while red dots represent female mice. Data presented as mean ± S.E.M; ***p < 0.001.
In the initial exploration phase on the first day, preventive WIN-treated, control Tg4-42 and WT mice allocated equal time to explore both objects (Figure 3(g); one-way repeated measures ANOVA, treatment preventive: F(5,78) = 1.105, p = 0.3645). Subsequently, during the recognition memory test conducted 24 h later, both WIN-treated and control mice spent an equivalent duration exploring the familiar and novel object (Figure 3(c); one-way repeated measures ANOVA, treatment preventive: F(5,78) = 2.691, p = 0.0269; Bonferroni comparisons: vehicle N versus F: p > 0.05; WIN N versus F: p > 0.05). This observation suggests an inability to discriminate between the two objects in both groups. In contrast, WT mice exhibited an intact memory, as indicated by a clear preference for the novel object (WT N versus F: p < 0.001). There was no variation in the distance traveled between the treatment groups on either day (Figure 3(b),(d); one-way repeated measures ANOVA, day 1: F(2,39) = 0.4353, p = 0.6502; day 2: F(2,39) = 1.005, p = 0.3752).
In addition, Tg4-42 therapeutically treated with WIN 55,212-2 or the vehicle solution explored the two identical objects during in exploration phase equally, similar to the WT controls (Figure 3(e); one-way repeated measures ANOVA, treatment therapeutic: F(5,66) = 0.9399, p = 0.4612). On the testing day, Tg4-42 mice treated with WIN demonstrated the ability to differentiate between the novel and familiar objects, suggesting intact recognition memory similar to that of WT controls. (Figure 3(g); one-way repeated measures ANOVA, treatment therapeutic: F(5,66) = 6.916, p < 0.001; Bonferroni comparisons: WIN N versus F: p < 0.01; WT N versus F: p < 0.001). In contrast, vehicle control mice showed no significant preference for the novel object (WIN N versus F: p > 0.05). The distance traveled did not differ between mice treated therapeutically with WIN, those treated with the vehicle, and WT controls (Figure 3(f),(h); one-way repeated measures ANOVA, therapeutic: day1 F(2,33) = 2.258, p = 0.1204; day 2: F(2,33) = 2.233, p = 0.1232).
Prolonged WIN 55,212-2 treatment rescues spatial reference memory deficits in Tg4-42 mice
At 6.5 months of age, Tg4-42 mice displayed spatial reference memory deficits in the Morris water maze (MWM) task, consistent with previous descriptions.37,40 Testing began with three days of cued training to eliminate possible motor or sensory impairments that could affect the mice's performance. Over these three days of cued training, all groups displayed decreasing escape latencies and successfully completed the test (data not shown, two-way repeated measures ANOVA, treatment preventive: F(2,41) = 3.126, p = 0.0550; treatment therapeutic: F(2,38) = 3.123, p = 0.0552).
During the subsequent acquisition training, Tg4-42 mice showed longer escape latencies than WT mice, as previously described 37 (Figure 4(a); two-way repeated measures ANOVA, treatment preventive: F(2,41) = 25.07, p < 0.001; Bonferroni comparisons: WT versus Tg4-42Vehicle: p < 0.001; WT versus Tg4-42WIN 55,212-2: p < 0.01). However, WIN-treated Tg4-42 mice performed significantly better than vehicle control animals. Across the 5 days of acquisition training, preventively WIN-treated mice showed faster escape latencies compared to control mice (Figure 4(a); Bonferroni comparisons: WIN versus Vehicle: p < 0.01). Similarly, Tg4-42 mice therapeutically treated with WIN 55,212-2 displayed faster escape latencies compared to control mice, performing similarly to WT mice (Figure 4(e); two-way repeated measures ANOVA, treatment therapeutic: F(2,38) = 15.24, p < 0.001; Bonferroni comparisons: WT versus Tg4-42Vehicle: p < 0.001; Tg4-42Vehicle versus Tg4-42WIN 55,212-2: p < 0.01; WT versus Tg4-42WIN 55,212-2: p > 0.05;). In addition, WIN-treated mice, regardless of treatment start, swam significantly faster than control mice over the five days of acquisition training (Figure 4(b),(f); two-way repeated measures ANOVA, treatment preventive: F(2,41) = 6.165, p = 0.0046; Bonferroni comparisons: WT versus Tg4-42Vehicle: p > 0.05; Tg4-42Vehicle versus Tg4-42WIN 55,212-2: p < 0.01; WT versus Tg4-42WIN 55,212-2: p < 0.05; treatment therapeutic: F(2,38) = 7.198, p = 0.0022; WT versus Tg4-42Vehicle: p > 0.05; Tg4-42Vehicle versus Tg4-42WIN 55,212-2: p < 0.01; WT versus Tg4-42WIN 55,212-2: p > 0.05).

Influence of prolonged WIN 55,212-2-treatment on spatial reference memory in Tg4-42 mice. (a) Preventive and (e) therapeutic WIN-treated mice exhibited significantly shorter latencies to reach the goal platform compared to control mice in the acquisition training. (b) Preventive and (f) therapeutic WIN-treated mice demonstrated significantly higher swimming speeds than vehicle-treated animals throughout the acquisition training and probe trial (d,h). During the probe trial, WIN-treated mice demonstrated a significant preference for the target quadrant (c,g), while vehicle-treated Tg4-42 mice displayed no preference for the target quadrant. The dashed line signifies the chance level. Two-way (a,b,e,f) and one-way (c,g,d,h) ANOVA followed by Bonferroni multiple comparisons; n = 12-15. Blue dots represent male mice, while red dots represent female mice. Data presented as mean ± S.E.M; * p < 0.05, ** p < 0.01, *** p < 0.001.
Twenty-four hours following the acquisition training, a probe trial was carried out to evaluate spatial reference memory. Both control groups of Tg4-42 mice displayed no preference for the target quadrant, indicating deficiencies in spatial reference memory (Figure 4(g); one-way repeated measures ANOVA followed by Bonferroni multiple comparisons, preventive: vehicle: F(3,52) = 1.584, p = 0.2043; therapeutic: vehicle: F(3,44) = 1.075, p = 0.3695). In contrast, Tg4-42 mice treated preventively with WIN demonstrated a notably stronger preference for the target quadrant, as evidenced by the proportion of time spent in different quadrants of the pool (Figure 4(c); one-way repeated measures ANOVA followed by Bonferroni multiple comparisons, preventive: WIN: F(3,56) = 26.48, p < 0.001; Bonferroni for target quadrant versus left, versus right and versus opposite quadrant: p < 0.001). Similarly, Tg4-42 mice treated therapeutically with WIN also spent significantly more time in the target quadrant (Figure 4(c); one-way repeated measures ANOVA followed by Bonferroni multiple comparisons, therapeutic: WIN: F(3,52) = 6.023, p = 0.0013; Bonferroni for target quadrant versus right and versus opposite quadrant: p < 0.01). WIN 55,212-2 Tg4-42 performed similar to WT mice in the probe trial (Figure 4(c),(g); one-way repeated measures ANOVA followed by Bonferroni multiple comparisons, preventive: WT: F(3,56) = 20.76, p < 0.001; Bonferroni for target quadrant versus right versus left versus opposite quadrant: p < 0.001; therapeutic: WT: F(3,52) = 59.81, p < 0.001; target quadrant versus right versus left versus opposite quadrant: p < 0.001).
Additionally, WIN-treated mice swam at a significantly faster pace than control mice, regardless of the treatment initiation (Figure 4(d),(h); preventive: one-way repeated measures ANOVA, F(2,41) = 5.464, p = 0.0079; Bonferroni multiple comparisons: WT versus Tg4-42Vehicle: p > 0.05; Tg4-42Vehicle versus Tg4-42WIN 55,212-2: p < 0.01; WT versus Tg4-42WIN 55,212-2: p > 0.05; treatment therapeutic: F(2,34) = 4.510, p = 0.0183; WT versus Tg4-42Vehicle: p > 0.05; Tg4-42Vehicle versus Tg4-42WIN 55,2122: p < 0.05; WT versus Tg4-42WIN 55,212-2: p < 0.05).
In summary, WIN treatment successfully mitigated spatial learning and spatial reference deficits in Tg4-42 mice.
Prolonged WIN 55,212-2 treatment does not alter anxiety behavior in the EPM or OF
The EPM was utilized to assess anxiety-related behavior subsequent to WIN treatment. Tg4-42 mice treated preventively or therapeutically with WIN 55,212-2 did not exhibit a significant difference in the duration of time spent in the open arms compared to control animals (Figure 5(a),(e); one-way repeated measures ANOVA, preventive: F(2,42) = 2.880 p = 0.0673; Bonferroni multiple comparisons: WT versus Tg4-42Vehicle: p < 0.05; Tg4-42Vehicle versus Tg4-42WIN 55,212-2: p > 0.05; WT versus Tg4-42WIN 55,212-2: p < 0.05; one-way repeated measures ANOVA, therapeutic: F(2,39) = 1.414 p = 0.2553). However, preventively treated Tg4-42 animals spend significantly more time in the open arms of the maze compared to WT mice (p = 0.057). Furthermore, preventively and therapeutically WIN-treated Tg4-42 mice traveled significantly more than the control animals (Figure 5(b),(f); one-way repeated measures ANOVA, preventive: F(2,42) = 1.793 p = 0.1790; Bonferroni multiple comparisons: WT versus Tg4-42Vehicle: p > 0.05; Tg4-42Vehicle versus Tg4-42WIN 55,212-2: p < 0.05; WT versus Tg4-42WIN 55,212-2: p < 0.05; therapeutic: F(2,39) = 0.6920 p = 0.5071).
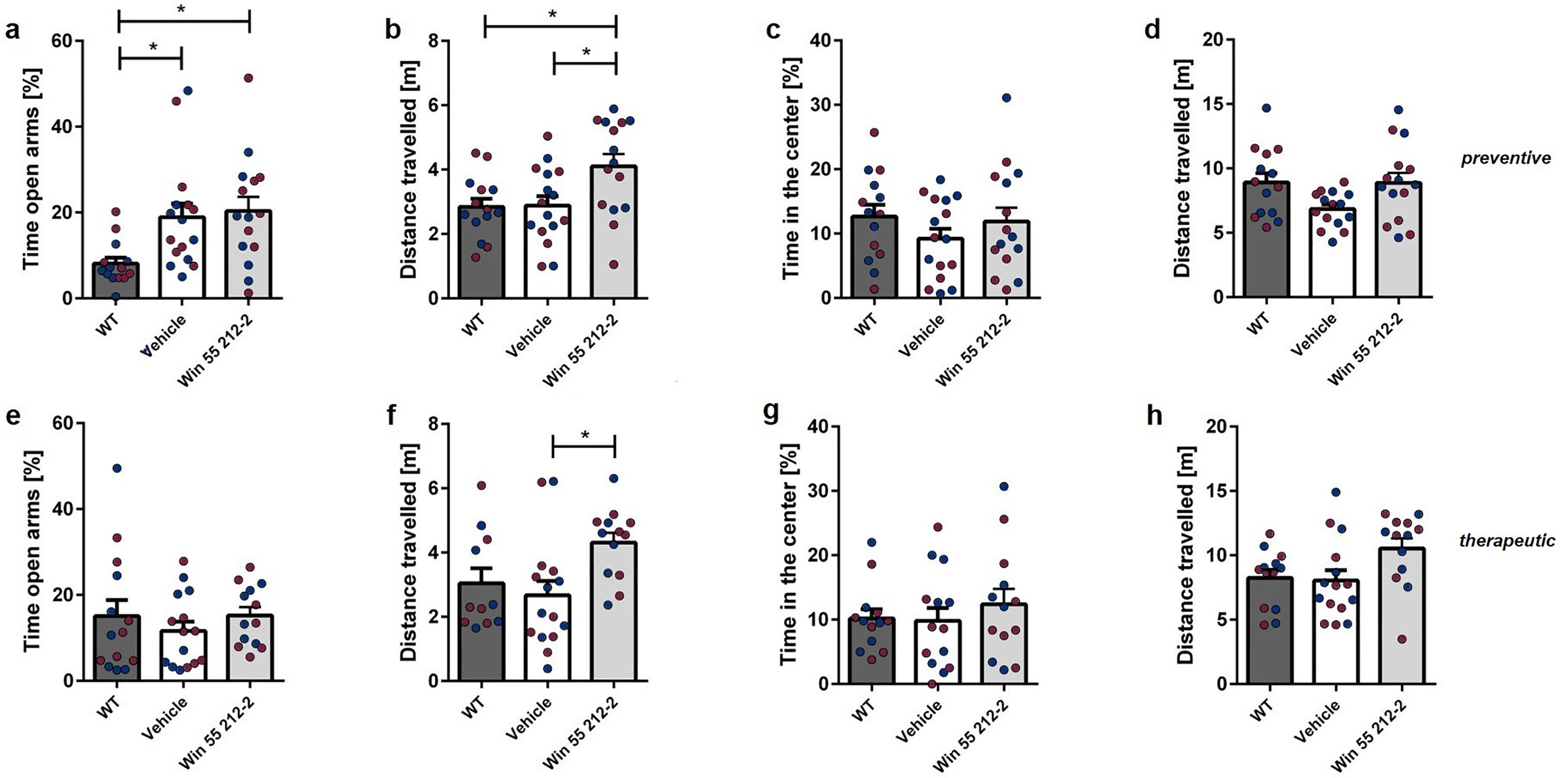
Influence of prolonged WIN 55,212-2-treatment on exploratory behavior and anxiety-related behavior in the elevated plus maze and open field in Tg4-42 mice. (a,e) The duration spent in the open arms of the EPM remained unchanged after WIN treatment, irrespective of the treatment start. (b,f) Preventive and therapeutic WIN-treated mice covered a significantly greater distance in the EPM compared to control mice. The time spent in the central area of the open field (c, g) did not exhibit a significant difference between WIN-treated and control mice, regardless of treatment initiation. (d,h) Both groups treated with WIN 55,212-2 travelled a similar distance in the OF as the control mice. One-way ANOVA followed by Bonferroni multiple comparisons, n = 13-16. Blue dots represent male mice, while red dots represent female mice. Data presented as mean ± S.E.M; *p < 0.05, **p < 0.01.
The OF test was used to assess the exploratory and spontaneous locomotor activity of mice. Regardless of the treatment time, there were no notable differences found between WIN-treated and control mice in terms of the time spent in the center (Figure 5(c),(g); one-way repeated measures ANOVA, preventive: F(2,42) = 3.505 p = 0.0393; therapeutic: F(2,39) = 1.722 p = 0.1927). Moreover, there was no variance in the distance covered between the WIN-treated group and the control mice (Figure 5(d),(h); one-way repeated measures ANOVA, preventive: F(2,42) = 2.507 p = 0.0939; therapeutic: F(2,39) = 0.3748 p = 0.6900).
Preventive WIN 55,212-2 treatment has anxiolytic effects in the DLB
The DLB was used to evaluate the anxiety status of mice. Tg4-42 mice treated preventively with WIN 55,212-2 spent significantly more time exploring the light area compared to vehicle-treated mice, performing similarly to WT controls (Figure 6(a); one-way repeated measures ANOVA, preventive: F(2,42) = 6.439 p = 0.0036; Bonferroni multiple comparisons: WT versus Tg4-42Vehicle: p < 0.05; Tg4-42Vehicle versus Tg4-42WIN 55,212-2: p < 0.05; WT versus Tg4-42WIN 55,212-2: p > 0.05). In contrast, therapeutic treatment with WIN 55,212-2 did not improved the anxiety-like phenotype of Tg4-42 mice. (Figure 6(d); one-way repeated measures ANOVA, therapeutic: F(2,39) = 0.6714 p = 0.0102; Bonferroni multiple comparisons: WT versus Tg4-42Vehicle: p < 0.01; Tg4-42Vehicle versus Tg4-42WIN 55,212-2: p > 0.05; WT versus Tg4-42WIN 55,212-2: p > 0.05). There was no difference in the time taken to enter the dark area between the treatment groups (Figure 6(b),(e); one-way repeated measures ANOVA, preventive: F(2,42) = 2.730 p = 0.0771; therapeutic: F(2,39) = 2.634 p = 0.0852). Tg4-42 mice displayed significantly fewer transitions between the areas. However, there was no effect on the number of transitions between the light and dark areas following WIN-treatment (Figure 6(c),(f); one-way repeated measures ANOVA, preventive: F(2,42) = 1.500 p = 0.0044; Bonferroni multiple comparisons: WT versus Tg4-42Vehicle: p < 0.05; Tg4-42Vehicle versus Tg4-42WIN 55,212-2: p > 0.05; WT versus Tg4-42WIN 55,212-2: p < 0.01; therapeutic: F(2,39) = 1.027 p = 0.0046; WT versus Tg4-42Vehicle: p < 0.01; Tg4-42Vehicle versus Tg4-42WIN 55,212-2: p > 0.05; WT versus Tg4-42WIN 55,212-2: p > 0.01).
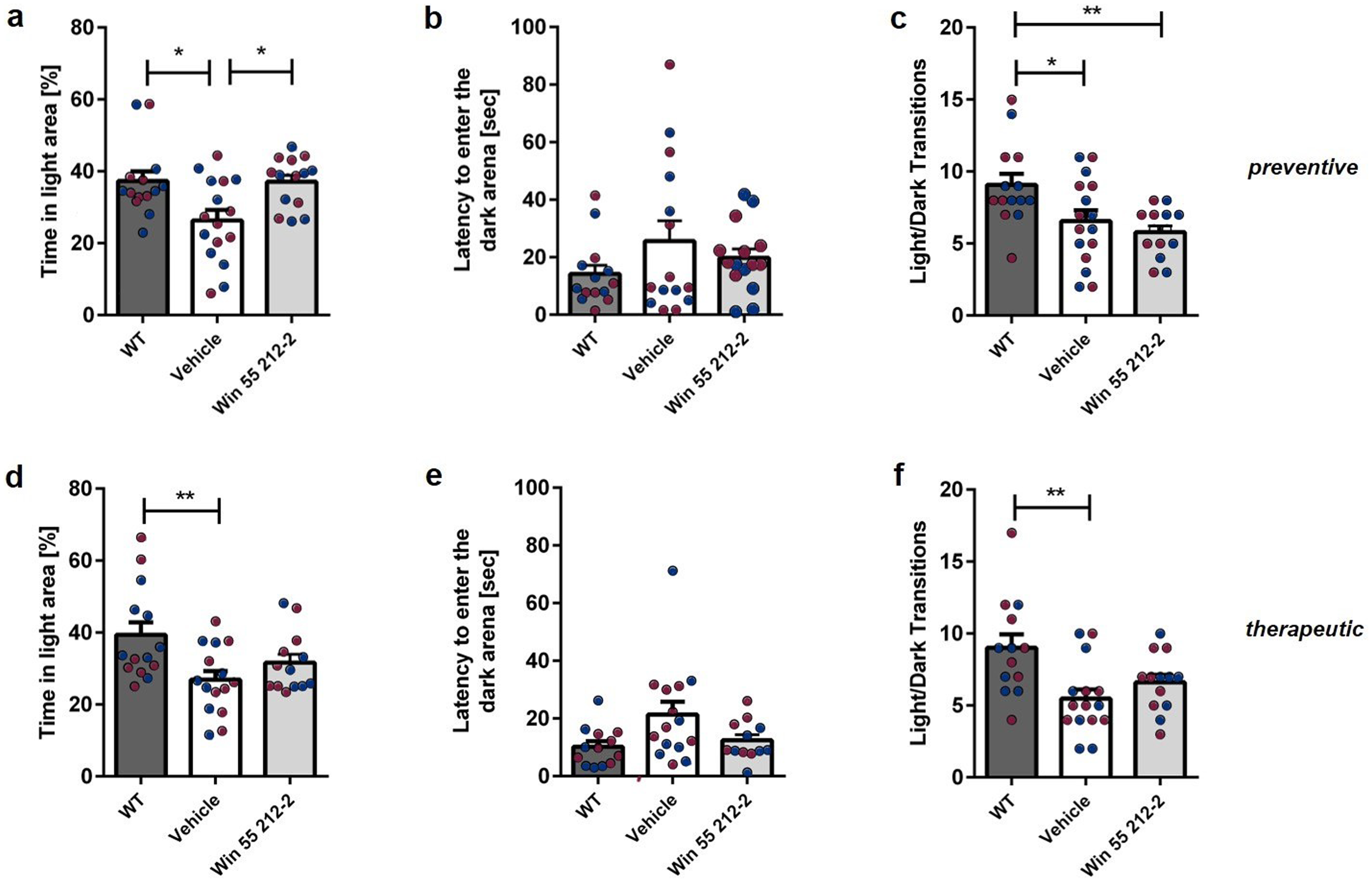
Influence of prolonged WIN 55,212-2-treatment on anxiety-related behavior in the Dark-Light Box in Tg4-42 mice. (a) Preventive WIN-treated Tg4-42 mice spent significantly more time in the light area than control mice. (d) No significant distinction in the duration spent in the illuminated area between mice treated therapeutically with WIN and the control group. (b,e) The time taken to enter the dark area was comparable between WIN- and vehicle-treated mice, irrespective of the treatment start. (c,f) The number of light/dark transitions, serving as an indicator of mobility, did not exhibit any significant differences between the treatment groups. One-way ANOVA followed by Bonferroni multiple comparisons, n = 13-16. Blue dots represent male mice, while red dots represent female mice. Data presented as mean ± S.E.M.
Prolonged WIN 55,212-2 treatment has no influence on neuron number or Aβ expression in the CA1 region of Tg4-42 mice
Homozygous 7-month-old Tg4-42 mice show a 58% neuron loss in the CA1 region of the hippocampus compared with WT mice. 37 Aβ4-42 peptides preferentially accumulate in the CA1 region of Tg4-42 mice.35,36 Therefore, we examined whether prolonged WIN 55,212-2 treatment could provide neuroprotective benefits or affect Aβ accumulation. However, WIN 55,212-2, independent of the treatment start, did not ameliorate the neuron loss in the hippocampus (Figure 7(d),(i); Supplemental Figure 1: one-way repeated measures ANOVA, treatment preventive: F(2,24) = 113.2, p < 0.001; Bonferroni comparisons: WT versus Tg4-42Vehicle: p < 0.001; WT versus Tg4-42WIN 55,212-2: p < 0.001; Tg4-42Vehicle versus Tg4-42WIN 55,212-2: p > 0.05; treatment therapeutic: F(2,32) = 175, p < 0.001; Bonferroni comparisons: WT versus Tg4-42Vehicle: p < 0.001; Tg4-42Vehicle versus Tg4-42WIN 55,212-2: p > 0.05; WT versus Tg4-42WIN 55,212-2: p < 0.001). Furthermore, WIN 55,212-2 did not affect Aβ4-42 accumulation in the CA1 region of the hippocampus (Figure 7(e),(j); Supplemental Figure 1: unpaired t-test, preventive: F(3,3) = 1.112, p = 0.9218; therapeutic: unpaired t-test: F(3,3) = 1.805, p = 0.7592).

Effects of prolonged WIN 55,212-2 treatment on AD pathology in Tg4-42 mice. (a) Preventive WIN treatment reduced microgliosis, while (f) therapeutic treatment had no effect on IBA1 expression in Tg4-42 mice. (b,g) GFAP expression remained unaltered after WIN treatment. (e,j) WIN had no impact on Aβ load in the hippocampus of Tg4-42 mice. Additionally, (c,h) WIN treatment did not affect neurogenesis in the hippocampus or the (d,i) number of neurons in the CA1 region. (k) Therapeutic WIN treatment resulted in increased 18F-FDG uptake in the hippocampus. (a-d; f-i) One-way ANOVA followed by Bonferroni multiple comparisons and (e,j,k) unpaired t-test. Blue dots represent male mice, while red dots represent female mice. Data presented as mean ± S.E.M.
In addition, neurogenesis (Supplemental Figure 1) is impaired in aged Tg4-42 mice compared to WT mice. 51 WIN 55,212-2 treatment did not affect the number of newborn neurons in the dentate gyrus of Tg4-42 mice (Figure 7(c),(h): one-way repeated measures ANOVA, treatment preventive: F(2,33) = 36.23, p < 0.001; Bonferroni comparisons: WT versus Tg4-42Vehicle: p < 0.001; WT versus Tg4-42WIN 55,212-2: p < 0.001; Tg4-42Vehicle versus Tg4-42WIN 55,212-2: p > 0.05; treatment therapeutic: F(2,28) = 58,72, p < 0.001; Bonferroni comparisons: WT versus Tg4-42Vehicle: p < 0.001; WT versus Tg4-42WIN 55,212-2: p < 0.001; Tg4-42Vehicle versus Tg4-42WIN 55,212-2: p > 0.05).
Prolonged WIN 55,212-2 treatment reduces microgliosis in the hippocampus of Tg4-42 mice
To assess whether prolonged WIN 55,212-2 treatment impacts disease-associated microgliosis (Supplemental Figure 1), a key aspect of neuroinflammatory pathology in Tg4-42 mice, 35 ionized calcium-binding adaptor molecule 1 (Iba1) was used as a marker for activated microglia. 52 Preventative treated Tg4-42 mice showed a reduced microgliosis in the hippocampus (Figure 7(a): one-way repeated measures ANOVA, treatment preventive: F(2,22) = 15.05, p < 0.001; Bonferroni comparisons: WT versus Tg4-42Vehicle: p < 0.001; Tg4-42Vehicle versus Tg4-42WIN 55,212-2: p < 0.001, WT versus Tg4-42WIN 55,212-2: p > 0.05). In contrast, therapeutic WIN 55,212-2 treatment did not affect microgliosis in the hippocampus of Tg4-42 animals (Figure 7(f): one-way repeated measures ANOVA, treatment therapeutic: F(2,22) = 15,81, p < 0.001; Bonferroni comparisons: WT versus Tg4-42Vehicle: p < 0.01; Tg4-42Vehicle versus Tg4-42WIN 55,212-2: p > 0.05; WT versus Tg4-42WIN 55,212-2: p < 0.001).
To evaluate the effects of WIN 55,212-2 treatment on astrogliosis, reactive astrocytes were quantified using glial fibrillary acidic protein (GFAP) as a marker (Supplemental Figure 1). 53 No differences in GFAP signal intensities were detected in the hippocampus after WIN 55,212-2 treatment, regardless of when the treatment was started (Figure 7(b),(g): one-way repeated measures ANOVA, treatment preventive: F(2,21) = 0.2033, p = 0.8176; treatment therapeutic: F(2,22) = 1.334, p = 0.2840).
Therapeutic WIN 55,212-2 treatment increases metabolic activity in the hippocampus of Tg4-42 mice
18F-FDG-PET/MRI was used to assess cerebral glucose metabolism in therapeutically treated Tg4-42 mice. 18F-FDG-PET is an established method for clinically diagnosing AD in patients 54 and for assessing neuronal dysfunction in AD mouse models, including Tg4-42. 36
Significant beneficial effects of WIN 55,212-2 on glucose metabolism in Tg4-42 mice were observed, as WIN 55,212-2 treatment resulted in increased 18F-FDG uptake in the hippocampus compared to control mice (Figure 7(k): unpaired t-test, therapeutic: unpaired t-test: F(3,3) = 8.969, p = 0.0181).
Discussion
Over the last years, cannabinoids have gained attention as potential therapeutic agents for treating AD. 55 The potential of the endocannabinoid system (ECS) in AD treatment is based on cannabinoids’ ability to address various processes involved in the pathogenesis of AD. These processes include Aβ and tau metabolism, inflammation, mitochondrial dysfunction, and excitotoxicity.22,55–57 In the current study, we examined the impact of extended WIN 55,212-2 treatment on Tg4-42 AD mice and its consequences on memory deficits, anxiety behavior, motor learning, and AD pathology. WIN 55,212-2 is a synthetic, exceptionally potent cannabinoid receptor agonist, exhibiting higher potency as a CB1/CB2 agonist compared to the phytocannabinoid tetrahydrocannabinol (THC).58,59 Here, we demonstrate for the first time that WIN 55,212-2 rescues spatial reference and recognition memory deficits and improves motor performance in Tg4-42 mice. Importantly, WIN treatment does not significantly affect anxiety-like behavior in Tg4-42 mice.
Progressive memory impairments are the central aspects of AD symptomatology. 60 Similarly, Tg4-42 mice display age-dependent memory deficits with severe impairments in spatial and recognition memory by the age of six months.36,40 The ECS plays a crucial role in the regulation of learning and memory processes25,26 and is involved in various memory processes including consolidation, destabilization, and forgetting. 61 In this study, we demonstrated that extended WIN treatment prevents spatial memory deficits in Tg4-42 mice in the MWM especially when administered preventively. The observed effects can possibly be attributed to the observed reduction in neuroinflammation in the hippocampus, possible mediated by the CB2 receptor. 62 For the first time, we were able to show that preventive cannabinoid treatment effectively counteracted the well-known Aβ-induced microglial activation in Tg4-42 mice. 35 These observations are in line with a study by Ramírez and colleagues, who demonstrated that intracerebroventricular administration of WIN55,212-2 to rats prevented Aβ-induced microglial activation and cognitive impairments. 62 Furthermore, Fakhouri et al. (2012) described a WIN-induced reduction in the expression of pro-inflammatory markers, including TNF-α, activated caspase-3, and nuclear NFκB, in the hippocampus of rats with Aβ-induced neurodegeneration. 34 Martin-Moreno et al. (2012) also reported a reduced expression of the inflammatory markers TNF-α and COX-2 in the hippocampus of transgenic APP mice following treatment with a synthetic cannabinoid. 30
Notably, the observed memory improvement following WIN treatment cannot be attributed to a reduction in Aβ levels in the hippocampus, as Aβ pathology remained unchanged. Aβ4–42 leads to intracellular Aβ accumulation in the brains of these mice, albeit without forming plaques. 41 Similar to our findings, prolonged oral administration of WIN did not reduce cortical Aβ1-40 levels in Tg APP mice in a study by Martín-Moreno and colleagues. However, the authors reported a reduction in Aβ1-42 without altering Aβ release in these mice. 30 In contrast to Tg4-42 mice, Tg APP mice display parenchymal Aβ plaques.63,64 It should be noted that only intraneuronal Aβ in the hippocampus of Tg4-42 mice was measured. Therefore, future studies should investigate the effects of WIN 55,212-2 on total and soluble Aβ in the brain.
Importantly, WIN treatment in the Tg4-42 mouse model resulted in only slight improvements in the therapeutic paradigm and notable improvements in the preventive strategy with respect to spatial memory performance. Thus, the age of Tg4-42 mice at the beginning of therapy appears to be critical in preserving the cognitive abilities in the MWM. The MWM test heavily relies on hippocampal function, and the spatial memory deficits observed in aged Tg4-42 mice can be attributed to significant neuron loss and increased inflammation in the hippocampus.35–37 Although neuronal dysfunction and glucose metabolic impairments have been described in 3-month-old Tg4-42 mice, no neuronal loss has been reported at this age. In contrast, at the start of the therapeutic treatment, Tg4-42 mice already demonstrate severe neuron loss exceeding 40% and severe inflammation. 37 Therefore, initiation of WIN treatment after the onset of severe neuronal loss appears to be too late to fully resolve the cognitive deficits in the MWM.
Consistent with our findings, positive effects of WIN 55,212-2 in the MWM were also demonstrated by Martin-Moreno et al. (2011) and Ramirez et al. (2005) using similar WIN dosages in AD mice and rats, respectively. Interestingly, both studies employed pharmacological models of AD, where Aβ peptides were intraventricularly injected.62,65 In addition, Bilkei-Gorzo et al. (2017) demonstrated that chronically administered low-dose THC, a CB1/CB2 receptor antagonist, could restore spatial reference memory in aged WT mice. 66
Furthermore, Martin-Moreno et al. (2012) examined the effects of orally administered 0.2 mg/kg WIN 55,212-2 and JWH-133, a selective CB2 receptor agonist, on eleven-month-old transgenic APP mice in the Novel Object Recognition Task. Interestingly, only JWH-133 showed a positive effect on the reference memory deficits in APP mice. Similarly, preventative WIN treatment had no effect on the reference deficits in Tg4-42 mice as they were unable to differentiate between a new and familiar object. In contrast, prolonged therapeutic WIN treatment prevented the observed reference memory deficits in Tg4-42 mice.
The observed effects on memory can possibly be attributed to the neuroprotective role of the CB1 receptor, which enhances synaptic plasticity. 62 While WIN treatment did not affect the number of neurons, we observed an increased FDG uptake in therapeutically treated mice, which might indicate heightened neuronal activity. Cannabinoids have previously been described as having neuroprotective effects across various experimental conditions. Activation of cannabinoid receptors has been shown to protect hippocampal and granule cerebellar neurons from excitotoxicity67–69 as well as from hypoxia and glucose deprivation. 70 Additionally, in vivo studies demonstrate that cannabinoids reduce hippocampal neuronal loss and infarct volume following cerebral ischemia.70–72 In addition to the hippocampus, the perirhinal cortex is essential for object recognition memory. The ability to recognize a previously encountered item as familiar depends on the integrity of the medial temporal lobe.73–78 Therefore, further research is needed to analyze the effects of WIN on different brain regions. Furthermore, it cannot be ruled out that the observed hypermetabolism may be influenced by improvements in vascular function, as previous studies have shown that WIN 55,212-2 and other cannabinoids can ameliorate vascular dysfunction in AD mouse models. 79 Vascular function and glucose metabolism are indirectly related, as improved vascular function can enhance glucose and oxygen delivery, potentially increasing metabolic activity detectable by FDG-PET. 80 However, further investigation using vascular MR imaging or specific PET tracers targeting blood flow is needed to directly assess potential improvements in vascular function and validate this hypothesis.81,82
The endocannabinoid system is known to play a crucial role in neurogenesis during juvenile and embryonic stages. Additionally, the ECS can modulate neurogenesis in adult individuals through cannabinoid receptors.83,84 However, in this study, WIN treatment had no effect on neurogenesis in Tg4-42 mice, whether administered preventively or therapeutically, compared to control animals. Our findings align with those of Abboussi et al. (2014), who also observed no impact of WIN on neurogenesis in adult animals. 85 In contrast, Marchalant et al. (2009) demonstrated an enhancement of age-related reduced neurogenesis in 23-month-old WT rats using a dose of 2 mg/kg WIN administered over four weeks. 86 The lack of effect in our study may be attributed to the much lower dose of WIN (0.2 mg/kg) used.
The ECS plays a role in the regulation of various physiological processes, including appetite, metabolism, and body weight. The activation of CB1 receptors typically stimulates appetite and leads to an increase food intake, while CB1 receptor antagonists suppress food intake and decrease weight in animals and humans.87–89 In a placebo-controlled crossover study conducted by Volicer and colleagues (1997), Dronabinol was effectively used to counteract weight loss in AD patients who refused food intake. 90 Sinnayah et al. (2008) demonstrated that the injection of 1 mg/kg of WIN 55,212-2 increased food intake in the hours following the injection in WT mice. 91 In contrast, chronic administration of 30 mg/kg of THC resulted in weight reduction in Sprague-Dawley rats. 92 Previous studies have indicated that THC exhibits a biphasic effect on rodent food intake, where moderate doses led to increased consumption, while higher doses resulted in reduced consumption.93–95 In the current study, prolonged WIN treatment did not influence the body weight of Tg4-42, independent of the treatment start, indicating that the selected dose was well tolerated by the animals in terms of metabolism.
In AD patients, particularly in the advanced stages of the disease, motor impairments, and agitation are well documented.96–98 In line with clinical observations, various mouse models of AD, including the Tg4-42 model, exhibit motor impairments.99–102 The impairments in Tg4-42 mice are age-dependent with motor learning and coordination deficits starting at 6 months of age. 39
It has been demonstrated that the endocannabinoid system effects motor function, synaptic plasticity, and motor learning.103,104 Noteworthy, CB1 and CB2 receptors have also been identified in brain regions crucial for motor control, including the cerebellum and basal ganglia. 105 Additionally, evidence of a biphasic effect of phytocannabinoids, synthetic and endogenous cannabinoids on motor activity has been described.106–108 Thereby, high doses of cannabinoids have been described to have hypoactive effects, even leading to catalepsy, while low doses have been associated with hyperactive effects.109,110 For example, the consumption of edible THC negatively affected subsequent drug-free rotarod performance in adolescent male C57BL/6J mice. 111 However, there are limited studies on the effects of WIN 55,212 on motor function. After spinal cord ischemia, Huo et al. demonstrated that WIN 55,212-2 enhanced motor function in male Sprague-Dawley rats as measured by the 14-point motor deficit index (MDI) score. 112 Furthermore, Arévalo-Marin et al. (2003) examined the effects of 10 mg/kg of WIN 55,212-2 on motor behavior in a mouse model of multiple sclerosis. While mice injected with WIN 55,212-2 exhibited a reduced locomotor response compared to controls, they did not display typical signs of catalepsy and performed significantly better in the rotarod test. The authors attributed these findings to a direct impact of WIN 55,212-2 on motor neurons, along with a reduction in inflammation in the central nervous system of the animals. 113 In line with these findings, therapeutic treatment with WIN 55,212-2 enhanced motor performance of Tg4-42 mice on the rotarod task.
Furthermore, therapeutic and preventative WIN treatment resulted in an increase in locomotor activity and swimming speed in Tg4-42 mice. Similarly, locomotor activity in the open field was increased when rats were treated with 0.6 mg/kg of WIN 55,212-2 but unaltered by higher doses of WIN. 114 In line with these findings, we could recently demonstrate increased locomotor activity and swimming speed in C57BL/6J wildtype after 42 days of WIN treatment. 22 In addition, Schramm-Sapyta et al. (2007) observed an increase in locomotor activity in adolescent rats following THC treatment. 115 In contrast, self-administration of THC has been shown to induce dose-dependent reductions in locomotor activity in healthy mice. 111 Furthermore, acute THC treatment resulted in a dose-dependent reduction in locomotor behavior in mice. 116 Although locomotor activity can potentially confound the evaluation of anxiety or memory performance, it is unlikely to have influenced the observed memory improvements or effects on anxiety behavior.
Repeated cannabinoid exposure can also have long-term consequences on anxiety-like behavior. Thereby, biphasic effects of cannabinoids have been shown in anxiety responses, with both anxiolytic- and anxiogenic-like effects being reported after cannabinoid treatment. 117 However, there is limited information available on the effects of cannabinoids on anxiety behavior in AD mice. Chesworth and colleagues recently showed that extended CBD treatment did not influence anxiety-like behaviors in APPXPS1 mice. 118 Onaivi et al. (1990) reported no effects in mice but noted an anxiety-inducing effect in rats following THC administration during adulthood. 119 O'Shea et al. (2004) did observe an increased anxiety behavior in adolescent rats following prolonged cannabinoid receptor agonist treatment. 120 Repeated THC treatment followed by a washout period resulted in minimal effects on anxiety-like behavior in adolescent and adult mice. 116 In contrast, Sestan-Pesa and colleagues did not observe any significant changes in the anxiety-like behaviors of adult mice following THC exposure during late adolescence. 121 In line with these findings, therapeutic WIN treatment did not affect anxiety behavior in Tg4-42 mice in the current study. Similar to our findings, Mouro et al. (2018) showed that chronic, intermittent administration of WIN 55,212-2 during adolescence had no impact on anxiety-like behavior in wildtype mice. 122 Additionally, we previously demonstrated that exposure to WIN 55,212-2 did not alter anxiety behavior in C57BL/6J mice with or without an extended washout phase. 22
In contrast, preventative WIN treatment resulted in an anxiolytic effect in Tg4-42 mice. Similarly, the administration of 0.25 mg/kg of WIN 55,212-2 resulted in a reduction of anxiety responses in a rat model of post-traumatic stress disorder. 123 Furthermore, Lisboa et al. (2018) observed a reduction in anxiety behavior in a mouse model of psychosocial stress after treatment with 1 mg/kg of WIN 55,212-2. 124 Importantly, the anxiolytic effect in Tg4-42 mice was only observed in the DLB. A review by Mohammad et al. (2016) highlighted that various anxiety tests, including DLB, EPM, and, OF often produce inconsistent results as these tests are influenced by numerous intrinsic and extrinsic factors. 125 Overall, the Tg4-42 WIN treatment did not severely influence anxiety-like behavior in Tg4-42 mice.
It should be noted that a relatively low dosage of WIN 55,212-2 was used in the current study. In in vivo studies, WIN 55,212-2 is commonly administered in doses ranging from 0.01 mg/kg to 6 mg/kg, with doses between 0.1 and 0.5 mg/kg considered low to moderate.30,65,126–130 In the present study, a low dose of 0.2 mg/kg WIN 55,212-2 was selected to minimize potential side effects, especially given the relatively long treatment period. The 0.2 mg/kg dose of WIN 55,212-2 used in this study is based on the positive findings of Martin-Moreno et al. (2012) in APP 2576 mice. The authors observed beneficial effects of WIN 55,212-2 on the pathology of AD mice following a 4-month treatment, with no reported side effects.
According to Tzavara et al. (2003), intraperitoneal administration of WIN 55,212-2 in vivo has activating effects at low doses and inhibitory effects at higher doses on the hippocampus. Furthermore, low doses of cannabinoids can stimulate D1-mediated responses that are more likely to improve cognition than to negatively impact learning and memory.131,132 Furthermore, Ferraro et al. (2001) demonstrated that acute administration of WIN 55,212-2 at low doses enhances cortical glutamate transmission, an effect that diminishes at higher doses. The authors speculated that higher doses of WIN 55,212-2 could trigger various cellular mechanisms involving both excitatory and inhibitory G-protein-coupled pathways, ultimately leading to no overall effect. 133
Importantly, studies have demonstrated that cannabinoids such as WIN 55,212-2 exhibit dose-dependent biphasic effects on locomotion, with low doses enhancing motor activation and high doses causing reduced motor activity and catalepsy.107,108,114 Additionally, high doses of WIN 55,212-2 have been shown to induce significant and prolonged sedation in animals. 131 In the preventative treatment approach, Tg4-42 mice treated with WIN performed significantly worse on the rotarod during the seventh trial, even with the low dosage used in this study. Furthermore, both acute and chronic administration of higher doses of WIN 55,212-2 can disrupt recognition and long-term memory.122,134 Therefore, more severe negative effects, particularly on motor function in Tg4-42 mice, cannot be ruled out if a higher dose of WIN were applied.
Given the potential side effects, especially with prolonged use, high doses of WIN 55,212-2 may be undesirable for treating AD in mice. It is important to note that, although WIN 55,212-2 treatment led to improvements in memory in Tg4-42 mice, it did not fully reverse their deficits, as treated mice generally did not reach WT performance levels in most tests. This partial efficacy suggests that while WIN 55,212-2 may mitigate certain aspects of AD-related pathology, it does not completely restore normal function. Further research is needed to assess whether dose escalation or combination therapies could enhance the therapeutic effects observed here. Nonetheless, WIN has already demonstrated efficacy at a dose of 0.2 mg/kg in the current study with Tg4-42 mice, as well as in a previous study with APP 2576 mice. 30
Importantly, we previously demonstrated that WIN 55,212-2 treatment in WT mice under the same experimental conditions alters cerebral metabolism and induces long-lasting behavioral deficits. 22 These adverse outcomes in WT mice contrast with the beneficial effects observed in Tg4-42 mice, suggesting that WIN 55,212-2's effects are highly context-dependent. In AD models, WIN 55,212-2 appeared to confer neuroprotection, possibly by modulating neuroinflammatory pathways or supporting cognitive functions compromised by AD pathology. Conversely, in WT mice, WIN 55,212-2 led to hypometabolism in brain regions such as the hippocampus, amygdala, and midbrain, and caused spatial and recognition memory deficits, suggesting a disruption of normal brain function in the absence of neurodegeneration. This contrast may reflect the unique neurobiological state in AD, where cannabinoid receptor activation could counteract disease mechanisms, rather than disrupting function as observed in healthy neural systems. These findings highlight a differential impact of WIN 55,212-2 depending on the brain's pathological state, underscoring the need for caution when considering cannabinoids for therapeutic use. While cannabinoid receptor agonists may hold therapeutic potential in neurodegenerative diseases like AD, they may be unsuitable for general use in healthy individuals.
In summary, our findings demonstrate that extended administration of the synthetic cannabinoid WIN 55.212-2 enhances motor learning, recognition memory, and spatial reference memory without significantly affecting anxiety-like behavior of Tg4-42. In conclusion, we described for the first time the positive effects of prolonged WIN treatment on the behavior of Tg4-42 mice. Given that WIN treatment did not result in significant adverse effects on the behavioral measures investigated, this treatment may hold clinical relevance as it demonstrated improvement in several phenotypes relevant to AD. However, future studies should investigate the effects of WIN treatment on brain pathology and the underlying mechanisms in Tg4-42 transgenic mice, thereby addressing the limitations of the current study design.
Supplemental Material
sj-docx-1-alr-10.1177_25424823241306770 - Supplemental material for Chronic exposure to a synthetic cannabinoid improves cognition and increases locomotor activity in Tg4-42 Alzheimer's disease mice
Supplemental material, sj-docx-1-alr-10.1177_25424823241306770 for Chronic exposure to a synthetic cannabinoid improves cognition and increases locomotor activity in Tg4-42 Alzheimer's disease mice by Frederik W Ott, Marius E Sichler, Caroline Bouter, Marzieh Enayati, Jens Wiltfang, Thomas A Bayer, Nicola Beindorff, Maximilian J Löw and Yvonne Bouter in Journal of Alzheimer's Disease Reports
Footnotes
Acknowledgments
We acknowledge the support by the Open Access Publication Funds of the Göttingen University. The Berlin Experimental Radionuclide Imaging Center (BERIC) is supported by Charité 3R—Replace, Reduce, Refine, Charité—Universitätsmedizin Berlin.
Author contributions
Frederik W Ott (Formal analysis; Investigation; Visualization; Writing – original draft); Marius E Sichler (Investigation); Caroline Bouter (Formal analysis; Investigation); Marzieh Enayati (Investigation); Jens Wiltfang (Resources); Thomas A Bayer (Resources); Nicola Beindorff (Investigation); Maximilian J Löw (Investigation); Yvonne Bouter (Conceptualization; Formal analysis; Methodology; Supervision; Writing – original draft).
Funding
The authors disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This study was supported by the “Alzheimer Stiftung Göttingen” to CB and YB. This work was supported in part by the Deutsche Forschungsgemeinschaft (DFG) for PET/MRI use (INST 335/454-1FUGG).
Declaration of conflicting interests
The University Medicine Göttingen holds a patent for the Tg4-42 mouse model, with TB being among the inventors. The remaining authors declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Data availability
The datasets used and/or analyzed during the current study is available from the corresponding author on reasonable request.
Supplemental material
Supplemental material for this article is available online.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.