
Abstract
CRISPR-based gene drives represent a powerful new technology for limiting disease transmission and controlling invasive populations. These systems rely on homology-directed repair (HDR) to “drive” a genetic element through a population. However, mammals tend to favor non-homologous end joining (NHEJ), which generates mutations that halt further drive propagation. Here, we describe the experimental characterization of a putative target locus for a gene drive system targeting the haploinsufficient spermatogenesis gene Klhl10 in the laboratory mouse. Using a newly designed “coding sequence cassette,” we introduce downstream guide RNAs within the gene, ensuring that sperm undergoing NHEJ are selectively removed from the population. As a proof of principle, we demonstrate that targeting Klhl10 with constitutively expressed LbCas12a results in strong selection against frameshift-containing sperm, validating the core purification mechanism required for this drive strategy. Unexpectedly, we also observed that female offspring lacked most frameshift mutations, suggesting a previously unrecognized role for Klhl10 in oogenesis or early embryonic development.
Introduction
Engineered gene drive is a powerful genetic technology designed to propagate specific genetic traits through populations at rates exceeding Mendelian inheritance. First conceptualized in the early 2000s and experimentally realized through CRISPR-Cas systems in the 2010s, gene drive systems can function by converting germline cells in heterozygous individuals into homozygotes for the drive allele, resulting in preferential transmission to offspring.1–4 This “super-Mendelian” inheritance has profound implications for ecological management, disease control, and evolutionary biology.
In wild populations, gene drives could control disease vectors like malaria-transmitting mosquitoes or preserve endangered organisms through genetic rescue, while localized forms might directly control populations of invasive species or pests.2,5,6 Initial successes in Anopheline mosquitos demonstrated high transmission rates exceeding 95% in both sexes in some laboratory studies. 7 However, the efficiency of gene drives varies considerably across species, with technical implementation challenges including localization, the evolution of resistance alleles, and importantly, differences in DNA repair pathway preferences. 8 The latter challenge is particularly pronounced in mammals, where it has proven difficult to bias double-strand break repair toward homology-directed repair (HDR) compared with non-homologous end joining (NHEJ).9–11 While HDR faithfully copies the drive element, NHEJ often creates indels that render the drive nonfunctional and generate resistance alleles.
The highest reported rates of HDR gene conversion in Mus musculus, 72% in females and 11% in males, are likely too low for efficient gene drive.9,10 In Rattus norvegicus, the highest reported rates of reusable HDR gene conversion are 67% in females and 0.9% in males. 12 This strong preference for NHEJ in rodents, especially in males, has severely limited gene drive efficiency in these organisms (depicted in Fig. 1A).
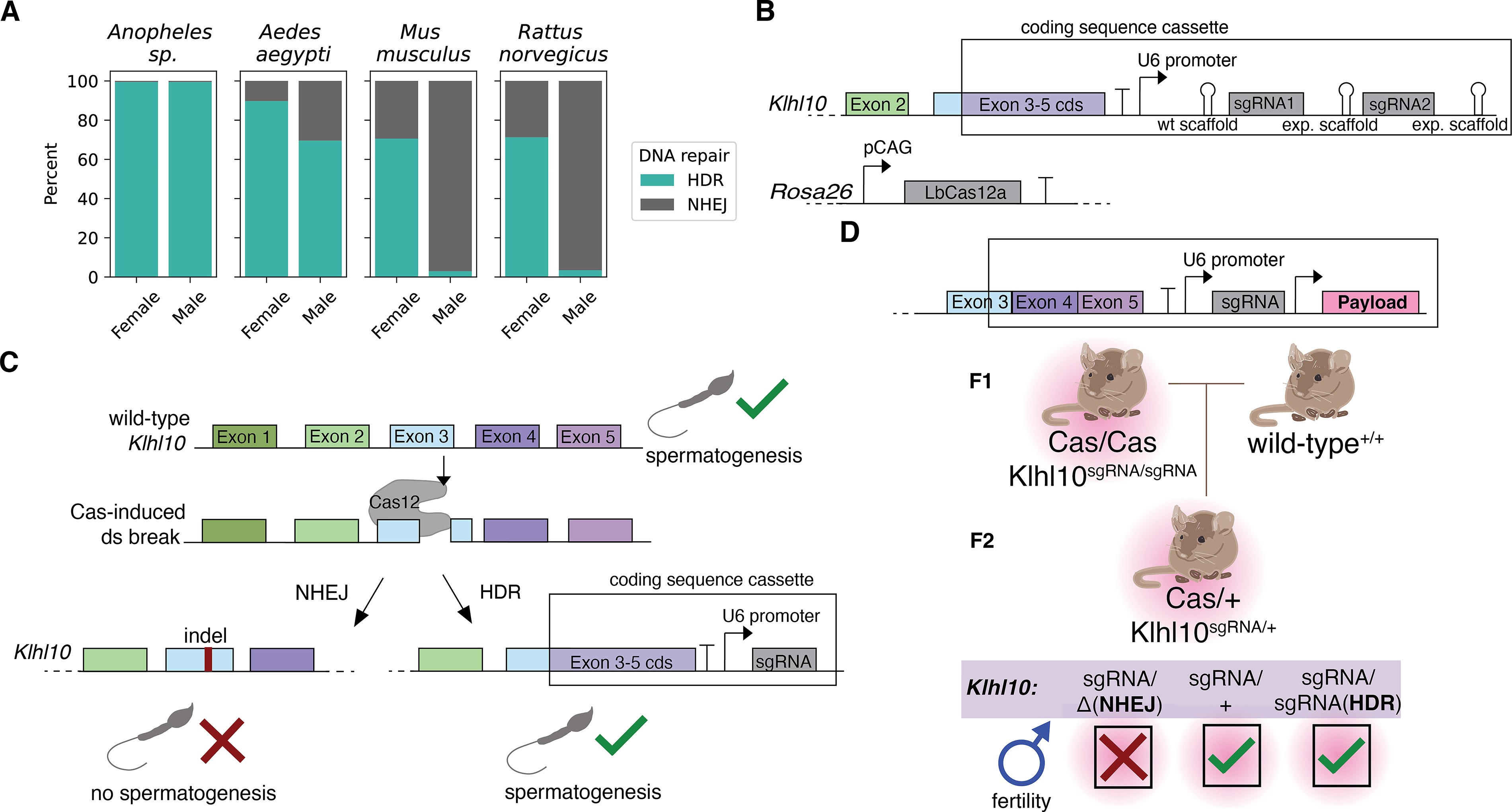
A proposed system of gene drive to compensate for low rates of HDR in males.
Our approach examines a potential solution to the low gene drive homing efficiency in male mice by targeting Klhl10, a gene that reportedly exhibits haploinsufficiency for spermatogenesis in mice. 15 Rather than attempting to increase HDR rates directly, we exploit the natural consequences of NHEJ at this locus: frameshift mutations render sperm nonfunctional, effectively ensuring that only drive-carrying or uncut sperm can produce offspring. This strategy eliminates NHEJ alleles at the abundant sperm stage, effectively decreasing the inheritance rate of mutated, drive-resistant alleles.
Our experimental system targets Klhl10 using the RR-variant of the Lachnospiraceae bacterium (Lb) variant of Cas12a (LbCas12a), which we selected for its multiplexing potential and targeting versatility. To investigate its utility, we generated transgenic mice expressing LbCas12a and guide RNAs targeting the Klhl10 locus in a split-drive system. Our analysis focused on understanding LbCas12 cutting efficiency, mutation patterns, and the feasibility of using this system for gene drive applications.
Methods
Mouse model generation and husbandry
Transgenic mice were generated via CRISPR/Cas-mediated pronuclear injection by the Division of Comparative Medicine MIT Core using donor plasmids. For the LbCas12a mice, SpCas9 ribonucleoprotein (RNP) was generated from commercial SpCas9 protein obtained from Integrated DNA Technologies (Coralville, IA, USA) and Rosa26-targeting crRNA (5′-ACTCCAGTCTTTCTAGAAGA-3′) from Synthego (Redwood City, CA, USA). The donor plasmid contained Rosa26 homology arms of 820 bp, upstream, and 530 bp, downstream, flanking a knock-in cassette of chicken beta-actin promoter (pCAG)-nuclear localization sequence (NLS)-LbCas12aRR-NLS-βA(polyA), PGK promoter-NLS-enhanced green fluorescent protein(EGFP)-NLS-βGH(polyA). For the Klhl10sgRNA mice, LbCas12a RNP was generated from purified LbCas12aRR obtained from the QB3 MacroLab at UC Berkeley (Berkeley, CA, USA) and the Klhl10-targeting crRNA (5′-GCCACCAATAGCGAATAGGATGG-3′) from Synthego (Redwood City, CA, USA). The donor plasmid contained Klhl10 homology arms of 500 bp, upstream, and 912 bp, downstream, flanking a knock-in cassette of Klhl10(CDS)-wtDR-crRNA1-mtDR1-crRNA2-mtDR2. The sequences for these elements is as follows: wtDR: 5′-TAATTTCTACTAAGTGTAGAT-3′, crRNA1: 5′-GCAGTGTTGAGACGCACGTAGCC-3′, mtDR1: 5′-TAATTTCTACTTAGTGTAGAT-3′, crRNA2: 5′-GCCACCAATAGCGAATAGGATGG-3′, mtDR2: 5′-TAATTTCTACTAATAGTAGAT-3′. Injected embryos were implanted into pseudopregnant females. Of 39 pups born from the Rosa26-LbCas12a injection, one pup carried the full-length knock-in. Of the 22 pups born from the Klhl10-Klhl10sgRNA injection, three pups carried the full-length knock-in. Additional sequencing confirmed correct integration at the target locus in one mouse. Mice were generated and maintained on a C57BL/6NTac background. Food and water were available ad libitum. One mouse was determined to be infertile after harem breeding for 17 weeks produced no offspring.
DNA extraction and genotyping
Genomic DNA was extracted from mouse ear tissue by overnight incubation of 12.5 µL of a 20 mg/mL stock proteinase K and 500 µL of “tail buffer” (10 mM 1M Tris pH 8.0, 100 mM 5M NaCl, 0.5M Ethylenediaminetetraacetic acid [EDTA] pH 8.0, 0.5% sodium dodecyl sulfate [SDS]) at 55°C in 1.7 mL microcentrifuge tubes. After overnight incubation, 167 µL of 5M NaCl was added and mixed for 5 min, then centrifuged on a benchtop centrifuge at 16,000 x g for 10 min. 500 µL of supernatant was removed and replaced with 500 µL of isopropanol and inverted until a precipitate formed. DNA was then extracted using phenol–chloroform and purified via ethanol precipitation.
For F3, mice were initially genotyped using Transnetyx (Cordova, TN) to determine which were homozygous or heterozygous for the engineered allele. For mice that were determined to be heterozygous, a follow-up DNA extraction, polymerase chain reaction (PCR), and sequencing were used to determine genotype.
Klhl10 sanger and NGS sequencing
For Sanger sequencing and Synthego ICE analysis, a pair of primers (oWK1307: 5′-TACCACCTCCACAAAACAAAACCATCTAT-3′, oWK1308: 5′-GAACTGTCTCAAATAAGTAAATGGGGGCT-3′) was used to amplify the targeted gDNA fragment with Primestar GXL DNA polymerase (Takara Bio USA, San Jose, CA). PCR products were purified and Sanger sequenced using primer pair (oWK1414: 5′-TATCAAAAATACAGTATAGCAATTAACAGTG-3′, oWK1149: 5′-AAGCAAACTAAGGGGCTGG-3′). For next-generation sequencing (NGS) analysis, an adaptor-ligated amplicon was generated using primer pair (oWK1837-R1: 5′-TCCCTACACGACGCTCTTCCGATCTGTTAAGCGTTTTGACCCTGTGAAGAAAAC-3′, oWK1838-R2: 5′-GTTCAGACGTGTGCTCTTCCGATCTCCTGGTTCTTTACCTTCCCATAGAGTGTT-3′) and was sent to Quintara Bio (Cambridge, MA) for Illumina MiSeq sequencing. Fastq reads were then analyzed using CRISPResso2 using default settings for paired-end reads. For both Synthego ICE and CRISPResso2 analyses, we analyzed indels from the LbCas12-RR guide sequence (5′-GCAGTGTTGAGACGCACGTAGCC-3′). While the wildtype scaffold showed the expected activity, the experimental scaffold failed to produce detectable cutting at its target site in vivo. Downstream LbCas12a cutting analyses were therefore only conducted using the wildtype scaffolded guide.
Population modeling
Gene drive dynamics were simulated using a discrete-generation Wright-Fisher model with random mating, a carrying capacity of 10,000, and a survival rate of 0.95 per generation. Drive parameters and purification efficiencies are described in Results. Simulations were run for 30 generations with 100 D/D individuals introduced at generation 1. All modeling was performed in R (v4.4.1).
Institutional ethics and regulatory approval
This work has obtained regulatory approval from the relevant institutions. All activities involving recombinant DNA and biosafety were carried out in accordance with guidelines set forth by the MIT Environment, Health, Safety committee. Furthermore, all animal-related work adhered to the guidelines established by the MIT Institutional Animal Care and Use Committee (IACUC), following approved protocols (Protocol 2304000512).
Results
A system of drive targeting haploinsufficient spermatogenesis genes
Our design targets the Klhl10 gene, reportedly haploinsufficient for spermatogenesis, 15 with the LbCas12a(RR) endonuclease to overcome low mammalian HDR rates through strong selection against NHEJ-containing sperm (Fig. 1A). Because initial attempts to express LbCas12a from the meiosis-specific SPO11 locus did not yield appreciable cutting rates (data not shown), our experimental design tested the feasibility of sperm-specific selection for HDR by constitutively expressing LbCas12a with the CAG promoter from known safe harbor locus ROSA26 (Fig. 1B). Given the constitutive CRISPR activity outside the meiotic window and the apparent lack of a significant majority of offspring inheriting the sgRNA allele, we did not expect nor observe HDR. Instead, we sought to experimentally test the efficacy of targeting spermatogenesis gene Klhl10 with CRISPR to select against alleles modified by NHEJ.
As previous research has shown that male mice require two functioning alleles of Klhl10 for fertility, 15 we hypothesized that in the context of a drive system, NHEJ repair will produce indels that will disrupt spermatogenesis, while HDR introduces a “coding sequence cassette” containing the remaining protein sequence needed for Klhl10 function (Fig. 1C). Importantly, this system is amenable to driving a genetic payload downstream of the guide targeting the spermatogenesis gene. The payload (Fig. 1D) could be another guide RNA or a protein such as an antibody. In order to test a design with multiple guide RNAs, which can increase genetic stability, we evaluated a dual-guide system incorporating a wild-type crRNA scaffold with a guide RNA targeting exon 3 and an experimental scaffold with a guide RNA targeting a downstream site in exon 3 (Fig. 1B).
LbCas12a in mice
To robustly measure the in vivo cutting efficiency of LbCas12a, we analyzed the genetics of Klhl10 in F2 mice (Fig. 2) by performing targeted Sanger sequencing followed by trace decomposition analysis of ear tissue samples. 16 Out of 21 initial offspring, 8 (38%) exhibited detectable mosaicism (Fig. 2B). Of the 8 mosaic mice, edits were observed in 4% of cells to 87% of cells, with an average of 36% of cells containing indels (Fig. 2C). Three male mice and four female mice were selected for breeding. We were able to detect a wide range of indels in these mosaics, from a 17 base pair deletion to a 14 base pair insertion (Fig. 3A). Previous work using SpCas9 expressed from the same locus under the same promoter yielded 94 − 100% mosaicism in offspring mice.9,11
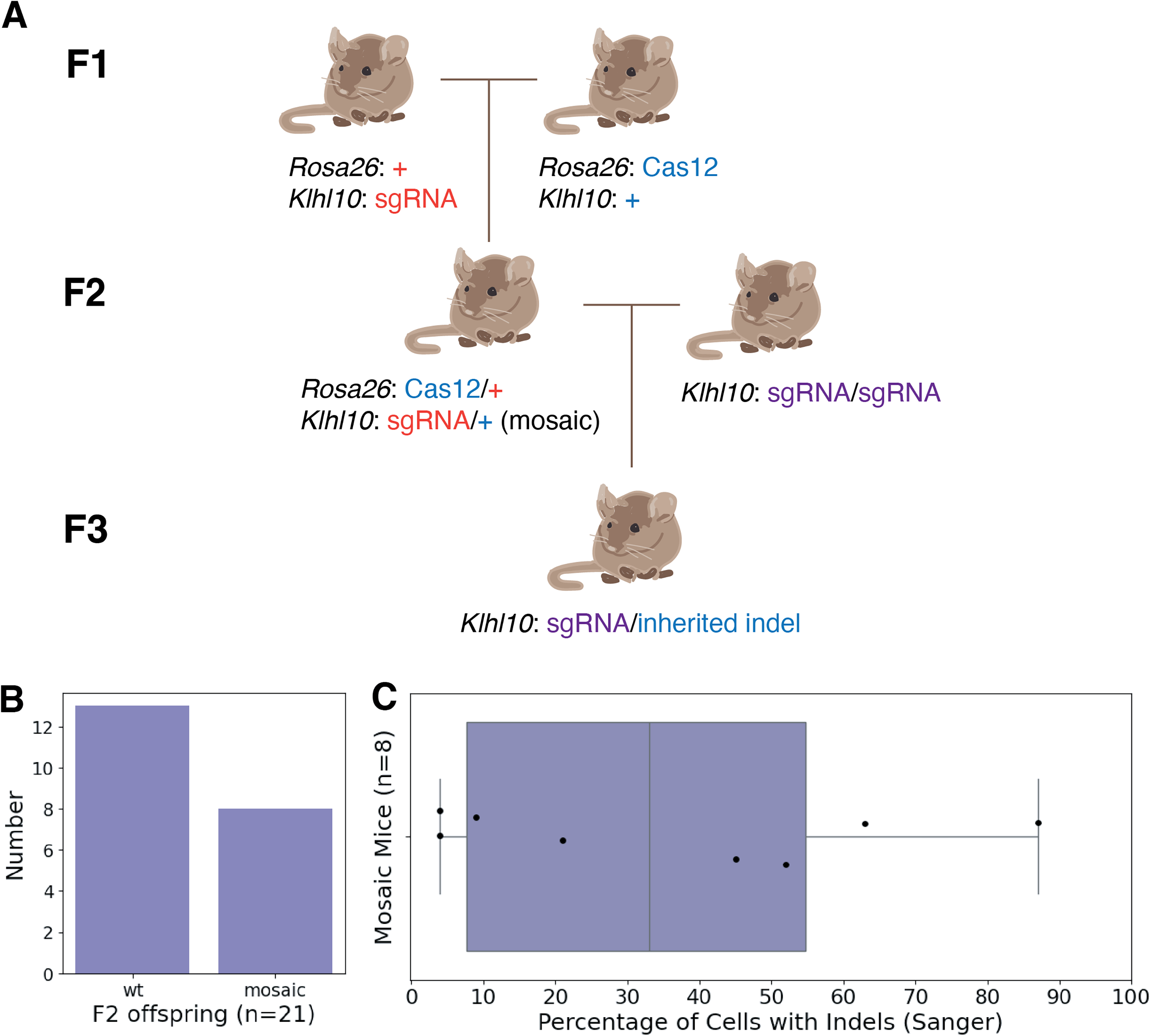
LbCas12a has moderate, varied cutting efficiency in vivo.
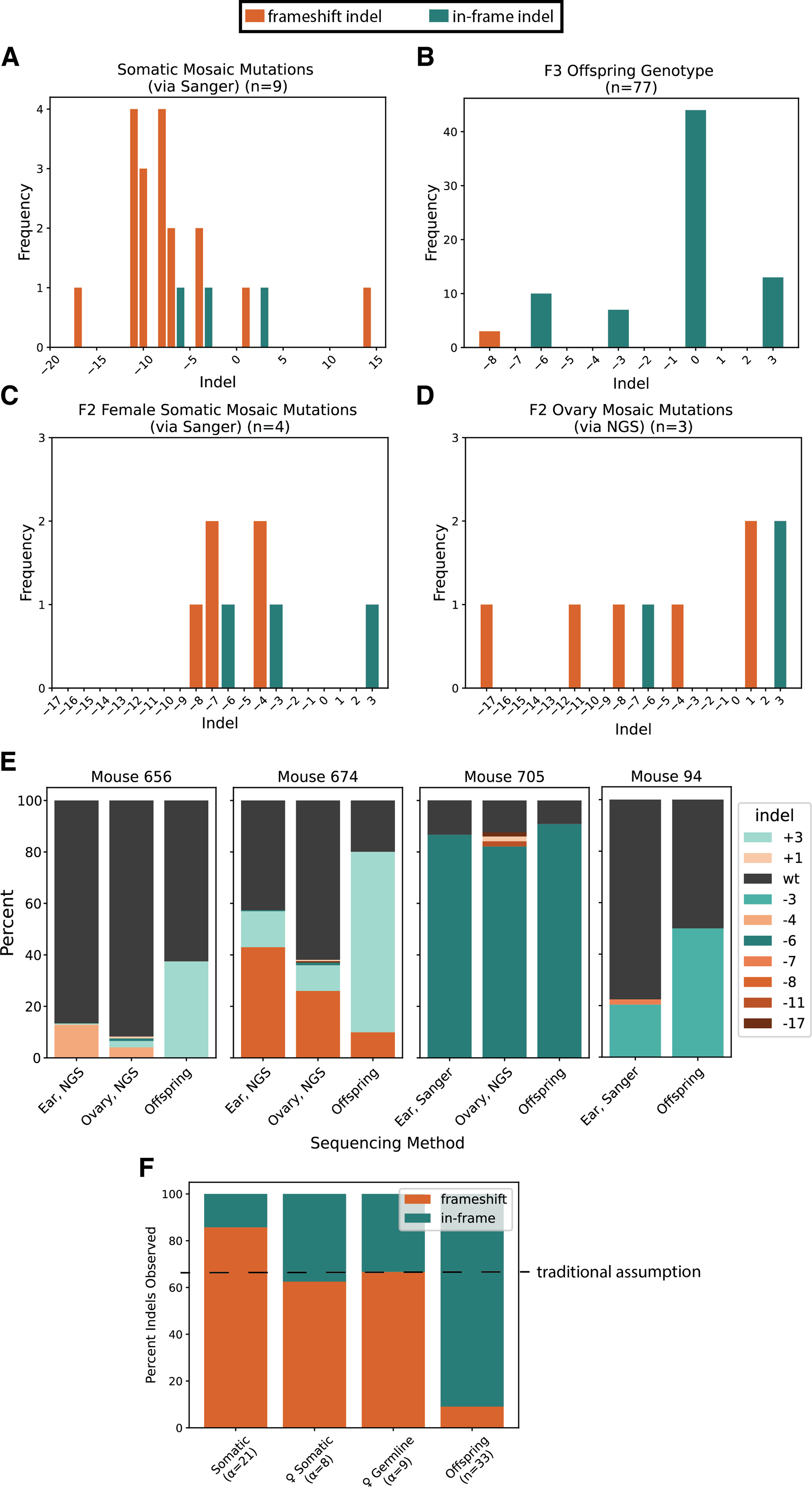
Selection for in-frame mutations purifies Klhl10 alleles.
Male mice with mosaic edits either produced offspring without indels (n = 2) or no offspring (n = 1). This is consistent with previous research that found that low-percentage Klhl10 chimeric males were able to produce wild-type offspring, but that high-percentage chimeric males were infertile. 15 Notably, in our experiment, males with a lower frequency of mosaicism (4% and 50% somatic tissue mosaicism) produced offspring, whereas the infertile male featured 63% knock-out mutations in somatic tissue and 91% in germline tissue (Supplementary Table S1). In establishing and maintaining the mouse line, we observed that male mice that were heterozygotic or homozygotic for this knock-in cassette were able to produce typical numbers of offspring that successfully inherited our engineered allele, confirming that the “coding sequence cassette” restores function of the gene and that there are no obvious fertility defects associated with our design (Supplementary Table S1). Given the constitutive CRISPR activity outside the meiotic window and the apparent lack of a significant majority of offspring inheriting the sgRNA allele, we did not observe HDR.
Female mosaic mice (n = 4) produced typical numbers of offspring (Supplementary Table S1). However, we were surprised to see that virtually all of these offspring contained non-frameshift mutations (Fig. 3B). Given that our initial Sanger sequencing of the mosaic parents yielded a wide variety of edits (Fig. 3A), we investigated the consequences of Klhl10 mutations for viability. We first confirmed that our analysis was accurate using NGS; we found similar results from the trace decomposition analysis and NGS/CRISPResso2 analysis (Supplementary Fig. S1).16,17 We then dissected mice to compare the somatic and germline tissues, given that various tissues can have different ratios or kinds of indels, to explore the possibility of previously undescribed developmental functions of Klhl10 (Fig. 3C–E).
Klhl10 gene analysis
Our 4 mosaic female mice had 55 total offspring. Initial analysis of the mosaic mothers indicated that 86% of observed mutations in parent somatic tissue were frameshifts, whereas only 9% of offspring inherited frameshift mutations (Fig. 3F). In some breeding pairs (mouse 656 and 674), a mutation that was a minority mutation in the somatic and germline tissue was heavily favored in offspring, suggesting some form of selection (Fig. 3E).
The only frameshift mutation that was inherited by offspring mice was a single −8 deletion. Given the assumption that ⅔ of mutations will be frameshift mutations and ⅓ of mutations will be in-frame mutations in the absence of selection, we calculated the probability of this occurrence using a cumulative binomial distribution. We observed 33 mice that inherited indels from 4 mosaic mothers; of those, 30 of them inherited in-frame mutations (+3, −3, −6), whereas 3 inherited a frameshift mutation (−8). Of 33 offspring, where P(frameshift) = 2/3, the cumulative binomial probability of 3 or fewer mice containing frameshift mutations by chance is
To ensure that our results were not biased by differential somatic and germline editing, we performed a similar analysis on the distribution of indels present in the parent mouse ovarian tissue (Fig. 3D). There were nine types of indels detected in the ovaries of three females, of which three were in-frame mutations, suggesting that there is no selection against somatic editing (Fig. 3F). However, these same females produced 23 in-frame offspring and 3 frameshift offspring; therefore, given 26 offspring and P(frameshift) = (2/3), the cumulative binomial probability of 3 or fewer such offspring is
We went on to successfully breed a heterozygote female mouse with the −8 deletion, observing no inheritance bias (Supplemental Fig. 2B). We similarly bred F3 heterozygote males and females with in-frame mutations (+3 and −6) and did not observe detectable inheritance biases between the mutations and the engineered allele (Supplementary Fig. S2A). Therefore, our targeted mutation site purifies most frameshift mutations in both males and females but not the −8 frameshift or the observed in-frame mutations. As there is no crystal structure of the Klhl10 protein, we used AlphaFold to examine the predicted structure and visualize our mutation target (Supplementary Fig. S3A). 18 All indels occur in an exterior loop, and none of them are predicted to alter the overall structure of the protein (Supplementary Fig. S3B). The reason that the −8 frameshift mutation does not impair female development, unlike all other possible frameshifts, is unclear.
Modeling spermatogenic selection
To evaluate the population-level dynamics of our purifying gene drive system, we adapted a previous modeling framework for mouse populations. 19 We parameterized the model using the sex-specific germline activity observed in females in Grunwald et al and in males in Weitzel et al: female homing efficiency of 37.6% (calculated as pC(1−pN) where pC = 0.552 and pN = 0.319) and a male homing efficiency of 3.0% (pC = 0.289, pN = 0.895).9,10 The model simulated a population of 10,000 mice following a one-time release of 100 D/D homozygous drive-carrying individuals. Purification is modeled as occurring during germline activity in each breeding cycle in both female and male drive carriers. When drive-carrying individuals produce gametes, NHEJ-derived resistance alleles (r2) are subject to purification with a specified efficiency. The remaining gametes are then renormalized, effectively increasing the transmission bias toward drive and wild-type alleles.
In the absence of purification (Fig. 4A), both the drive allele and the NHEJ-derived resistance alleles (r2) reached equilibrium at approximately 43% each within 30 generations. We expected purification to occur exclusively in male gametes, based on Klhl10’s only previously identified function in male gametogenesis (Fig. 4B). 15 To our surprise, about 83% of frameshift mutations were eliminated in female offspring, since only one of the six frameshift mutations identified in female ovaries was passed onto offspring (Fig. 4C). Lastly, we looked at the genotype composition dynamics under high purification. We observe progressive waves of the top three genotypes, with D/WT heterozygotes peaking around generation 15 as drive-carriers mate with wild-type individuals, then decline as the population transitions to predominately D/D homozygotes by generation 30. Targeting Klhl10 outperforms targeting a haploinsufficient lethal gene as well as a recessive lethal gene (Supplementary Fig. S4).
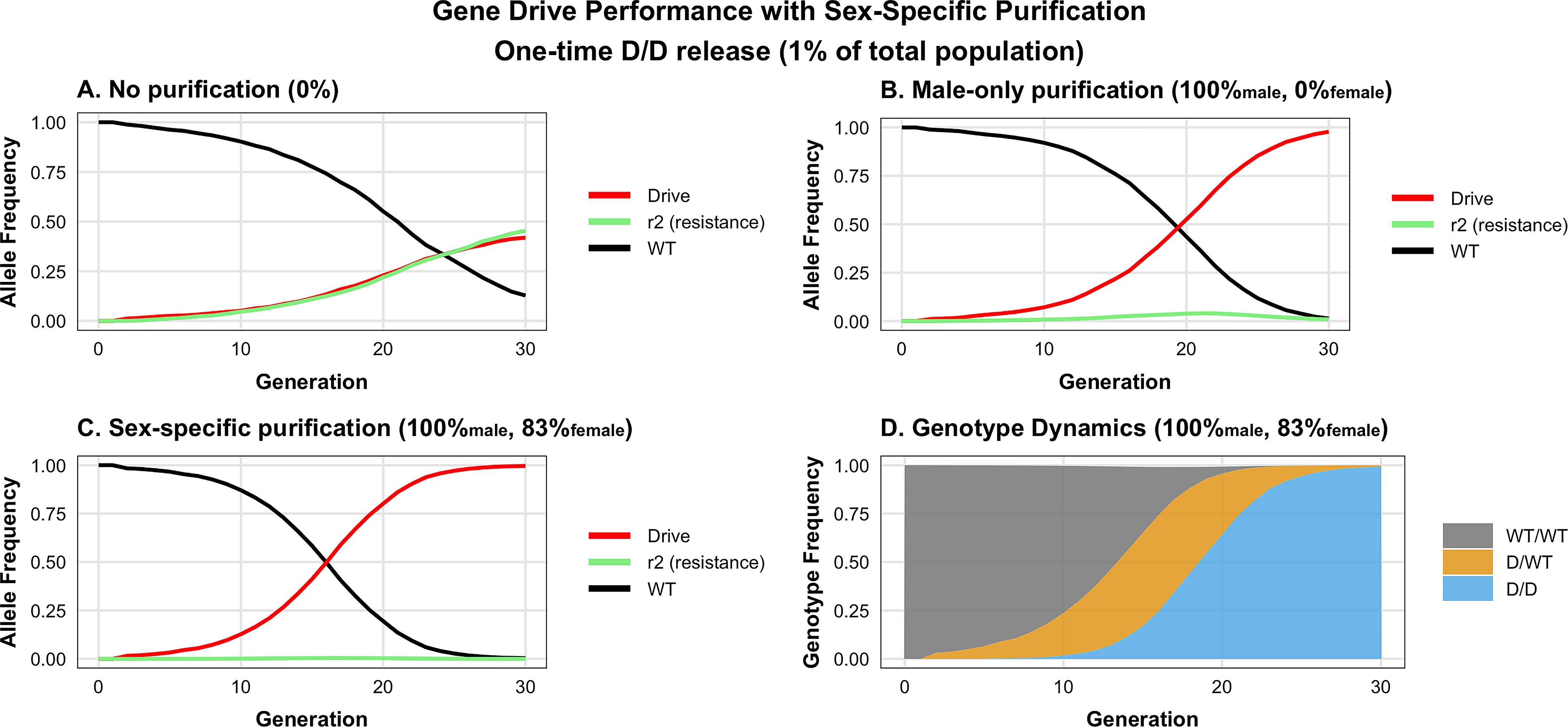
Modeling this system indicates enhanced purification of resistant alleles and drive efficiency.
These modeling results demonstrate that purification of NHEJ-derived resistance alleles could be a critical component of successful gene drive propagation given modest homing rates in rodents. The model predicts that our experimental approach of targeting haploinsufficient Klhl10 should provide sufficient purifying selection to enable drive spread, provided purification efficiency can be maximized through optimal guide selection and targeting.
Discussion
This work provides a proof of principle for targeting a haploinsufficient spermatogenesis gene as a means of purifying undesired repair outcomes, suggests previously unknown functions of Klhl10 in female fertility or oocyte development, and characterizes the effectiveness of LbCas12a in a mammalian gene drive context.
It is important to delineate what this study demonstrates from what remains to be shown. We validated that frameshift mutations at Klhl10 are effectively excluded from male reproduction: a male with high frameshift mosaicism (63% somatic/91% germline) was infertile, while the “coding sequence cassette” rescued fertility in heterozygous and homozygous knock-in males. This confirms that the purification mechanism central to our proposed drive strategy, the elimination of NHEJ outcomes via spermatogenic selection, functions as predicted. However, because constitutive Cas expression precludes HDR, we did not observe or measure drive-mediated gene conversion. A gene drive system would use a germline promoter to increase HDR. Fortunately, Klhl10 is expressed exclusively during spermiogenesis, well after the meiotic window in which germline-restricted Cas would act. 15 This temporal separation suggests that precut protein accumulation is unlikely to rescue NHEJ-bearing spermatids, and the established haploinsufficiency of Klhl10 confirms that intercellular bridge-mediated sharing is insufficient to compensate for loss of function. Regardless, a full demonstration of this gene drive system will require germline-restricted expression during meiosis to enable HDR while maintaining the spermatogenic selection we characterize here.
We were surprised to find a potential implication of Klhl10 in female fertility/embryogenesis when previously Klhl10 has only been associated with fertility in males. 15 Klhl10 belongs to the kelch-repeat-containing family, and proteins in this family are known to have varied roles, including cellular communication, ubiquitination, and activities involved in cellular structure.20,21 Of note, in Drosophila, kelch has a role in maintaining the actin organization of developing oocytes to facilitate cytoplasmic entry. 21 In addition, mouse embryonic transcriptomic analyses have identified that Klhl10 is expressed at the eight-cell and blastocyst stages. 22 Further research could examine the potential for this protein to have a role in mouse development and could further elucidate the candidacy of this gene for gene drive technology.
Notably, male and female selection against Klhl10 frameshifts appear to operate through distinct mechanisms with differing stringency. Male selection is absolute: high frameshift loads result in infertility, consistent with the established haploinsufficiency of Klhl10 in spermatogenesis. Female selection, while strong, permits some frameshift inheritance—including the −8 deletion, which was tolerated in females but would presumably contribute to male infertility. This asymmetry suggests that Klhl10 may play a less dosage-sensitive role in oogenesis or early embryogenesis than in spermatogenesis or that selection occurs at a developmental stage where mosaicism allows some affected cells to be compensated by unaffected neighbors. Regardless of mechanism, the critical observation for gene drive applications is that male purification—the primary bottleneck this strategy addresses—appears to be complete.
We assume functional sperm remain sufficiently abundant after purification to maintain fertility.23,24 This assumption warrants scrutiny: if sperm function requires a minimum threshold of viable cells, high cutting efficiency combined with low male HDR rates could compromise fertility. Our high-mosaicism infertile male (∼91% germline knock-out) is consistent with this concern, though it is equally consistent with simple haploinsufficiency of individual sperm. However, mice are polyandrous and subject to sperm competition, which should favor substantial overproduction of sperm relative to the minimum required for fertilization. 25 The extent to which this expected redundancy buffers against the loss of NHEJ-affected sperm remains unclear and may influence optimal drive parameters in practice. In addition, our model assumes monogamous mating. On average, multiple paternity rates in wild house mouse populations are around 25%. 26 Under polyandry, drive-carrying males with reduced sperm counts following NHEJ purification could be at a competitive disadvantage, though unlike systems where sperm function is intrinsically compromised, the remaining sperm from drive males should be fully competitive. The practical impact remains to be determined empirically.
We selected LbCas12a for its theoretical advantages: alternative protospacer adjacent motif (PAM) requirements, reduced recombination risk from scaffold-based array processing, and potential for iterative editing at previously cut sites.27,28 However, our results do not support its use in mammalian gene drives. The experimental mutant scaffold failed to produce detectable cutting at its target site. Overall editing efficiency was moderate (38% of offspring mosaic), consistent with recent Cas12a mouse studies but substantially lower than published SpCas9 systems in comparable contexts.9–11,29 One offspring showed possible evidence of iterative editing atop a parental +3 indel (Supplementary Fig. S5), but this theoretical advantage does not compensate for reduced cutting efficiency. We recommend future mammalian gene drive designs prioritize SpCas9 or similarly robust nucleases.
Our results indicate that targeting haploinsufficient reproductive genes can provide strong selection pressure against NHEJ outcomes in both sexes. The selection pressure against NHEJ outcomes arising from targeting a haploinsufficient spermatogenesis gene is comparable with one targeting a haploinsufficient essential gene and stronger than targeting a recessive lethal gene (Supplementary Fig. S4). It should be noted that the target site location may have influenced our results. PAM requirements for Cas12a led us to target exon 3, an area we have identified as a loop in a series of beta sheets. Due to this target site, it is possible that our system was more permissible to in-frame mutations. Targeting a structural section of the Klhl10 beta barrel sheets, especially upstream sequences, could provide stronger selection against in-frame mutations. The mechanism by which the −8 frameshift escapes female selection remains unclear; multiplexed guides targeting distinct sites could provide redundancy against such allele-specific exceptions.
Conclusion
In sum, we validate a potential gene drive target locus that could select against NHEJ mutations and enhance the effectiveness of a drive strategy: frameshift mutations at haploinsufficient spermatogenesis gene Klhl10 are excluded from male reproduction, while the “coding sequence cassette” rescues gene function without detectable fertility cost. Unexpectedly, female reproduction also selected against most frameshift alleles, suggesting additional developmental functions for Klhl10. These results support further development of this approach using germline-restricted CRISPR expression to enable efficient HDR-mediated gene drive.
Authors’ Contributions
C.L.P.: Conceptualization, investigation, formal analysis, writing, and methodology. W.S.K.: Conceptualization, investigation, formal analysis, writing, and methodology. J.F.: Investigation and mouse husbandry. Z.J.H.: Investigation. K.M.E.: Conceptualization, methodology, and funding acquisition.
Footnotes
Conflict of Interest
The authors declare no conflicts of interest.
Funding Information
This study was funded by the following grants: NIH: DP2 6937117 to KME NSF: NSF Career 262872.
Supplemental Material
Supplemental Material
Supplemental Material
Supplemental Material
Supplemental Material
Supplemental Material
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.