
Abstract
Introduction:
Tolerance and dependence are known to occur with prolonged cannabis use. Few animal experiments on spontaneous withdrawal from delta-9-tetrahydrocannabinol (THC), the primary psychoactive compound in cannabis, have been conducted. The experiments that have been conducted use extremely high doses and report relatively mild symptoms. The objective of the present experiment was to determine the magnitude and duration of spontaneous THC withdrawal in male and female rats using voluntary home cage wheel running as a sensitive, objective, and continuous measure of withdrawal.
Materials and Methods:
Male and female Sprague-Dawley rats were injected with THC (3 mg/kg, s.c.) or vehicle twice daily for 7 days to induce tolerance and dependence. Spontaneous withdrawal was assessed for 5 days beginning 24 h after the day 8 tolerance test in which all rats received a THC injection.
Results:
On day 1, THC profoundly decreased wheel running in male and female rats compared to vehicle-treated rats. On day 8, THC given to vehicle-treated rats produced a significantly greater decrease in running than in rats previously treated with THC, indicating tolerance development. There was no sex difference in the magnitude of the decrease in wheel running caused by THC injections or in the development of tolerance. There were no significant changes in wheel running associated with spontaneous THC withdrawal.
Discussion:
These data suggest that spontaneous THC withdrawal in male and female rats is mild at best despite profound locomotor suppression and tolerance to repeated injections. The lack of spontaneous THC withdrawal contrasts sharply with the pronounced changes in wheel running produced by opioid withdrawal.
Introduction
Prolonged use of high doses of cannabis can lead to cannabis use disorder. This disorder is characterized by spontaneous withdrawal symptoms upon termination of use. In humans these symptoms include irritability, anxiety, depressed mood, and sleep problems.1,2 In contrast to these psychological symptoms, animal research tends to focus on somatic symptoms of withdrawal such as paw tremors and head twitches, although changes in reward, anxiety, and sleep disturbances have been measured. Another major difference between human and animal characterizations of cannabinoid dependence is that most animal experiments use a cannabinoid receptor antagonist to precipitate withdrawal following prolonged cannabinoid agonist administration, even though precipitated withdrawal has limited clinical relevance to the spontaneous withdrawal that occurs in humans. The few animal experiments on spontaneous cannabinoid withdrawal that have been conducted are limited in that objective measures of spontaneous withdrawal are rare, extremely high doses of delta-9-tetrahydrocannabinol (THC) are used, and symptoms are rarely assessed for more than 3 days (72 h) following the last THC injection (Table 1), even though the primary withdrawal symptoms in humans occur from 2 to 6 days. 7
Published Experiments Assessing Spontaneous THC Withdrawal
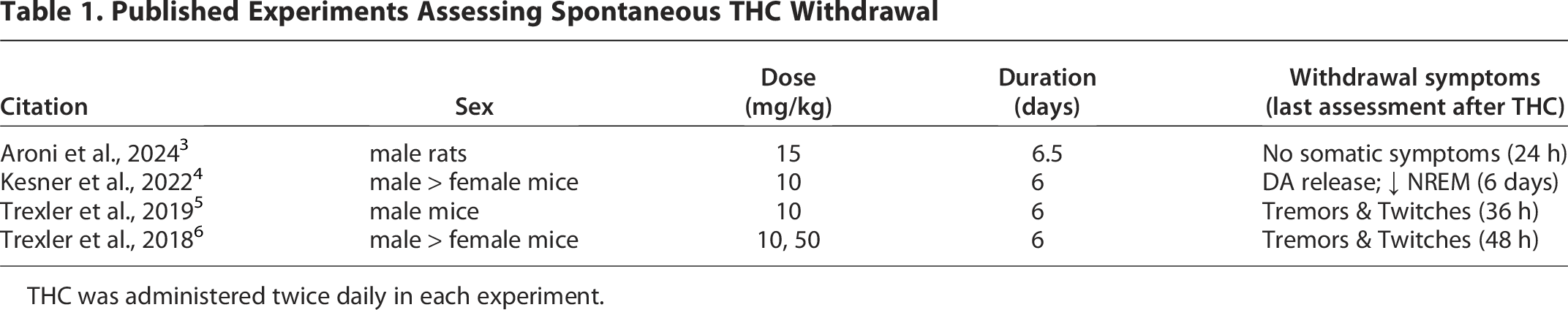
THC was administered twice daily in each experiment.
The two experiments examining sex differences in spontaneous THC withdrawal report more pronounced withdrawal in male compared to female mice. A comprehensive analysis of spontaneous withdrawal from the cannabinoid receptor agonist WIN 55,212-2 showed comparable somatic symptoms in male and female rats at 6–24 h after the last i.v. infusion and inconsistent sex differences at 7 and 15 days. 8 The somatic symptoms of spontaneous withdrawal are difficult to measure both because they are mild relative to symptoms observed during precipitated withdrawal 3 and the relevant time point at which to assess symptoms ranges from 6 h to more than 6 days after the last injection (see Table 1). Continuous assessment of spontaneous THC withdrawal using an objective and sensitive method is needed.
In previous studies, we have successfully used voluntary home cage wheel running to measure the magnitude and duration of spontaneous opioid withdrawal in rats. 9 Wheel running provides several advantages over traditional measures of withdrawal. Given that wheel running is assessed in the home cage, it is not confounded by the stress of moving the rat to a test chamber. Other advantages are that wheel running data are objective, continuous, and sensitive to any disruption in well-being. We have shown that pain, drug side effects, and opioid withdrawal each cause a decrease in voluntary wheel running.10,11 The objective of the current project is to determine the magnitude and duration of spontaneous THC withdrawal in male and female rats using home cage wheel running. This approach will help align preclinical research with the spontaneous withdrawal that occurs in humans.
Materials and Methods
Animals and housing
Sixteen male and 16 female Sprague-Dawley rats (Envigo Laboratories, California) between the ages of 66 and 67 days old at the time of the first THC injection were used. Rats were housed in same-sex pairs for 1 week upon arrival and then housed individually in standard Plexiglas cages (24 × 36 cm), each fitted with a top-mounted stainless steel running wheel (Tecniplast, Starr Life Sciences Corp., Pennsylvania). The diameter of the wheel was 33 cm so each wheel revolution was 1.037 meters. This apparatus allows for continuous, undisturbed measurement of voluntary wheel running in the home cage. Animals were housed eight to a room with 4 males and 4 females in each cohort. The room was maintained on a 12:12-h light-dark cycle (lights off at 10:30 or 11:00 AM depending on the cohort test room) in a temperature- and humidity-controlled room. Food and water were available ad libitum. Cages were changed once per week. This experiment was approved by the Washington State University Institutional Animal Care and Use Committee and conformed to specifications in the Guide for the Care and Use of Laboratory Animals (2011).
Independent variables
The experiment employed a 2 × 2 factorial design with sex and THC administration as between-subjects factors (8 rats/condition). Rats were allowed to habituate to wheel running for 11 days. Although a previous experiment indicates it takes at least 17 days for wheel running to reach a stable daily number, this study indicates that the duration of the baseline period (7 vs. 21 days) did not have a major effect on the result. 12 Sex differences are well established by 11 days of baseline running, and the effects of THC on running are evident even with shorter baseline periods.10,13 The mean of the last 2 days was used as the baseline value.
Half the male and half the female rats received twice-daily injections of THC (3 mg/kg, s.c.) while the other half received injections of vehicle (saline:ethanol:cremophor, 18:1:1) for 7 consecutive days. THC was provided as a gift from the NIDA Drug Supply Program. Injections were administered 5–10 min prior to the beginning of the dark phase and again 6 h into the dark phase. Tolerance to THC was assessed on the 8th day by injecting rats in all four groups (Male/Female × THC/Vehicle) with THC (3 mg/kg, s.c.) 25 min prior to the beginning of the dark phase. Withdrawal was assessed for 5 days beginning 1 day after the last THC injection (i.e., the day following the THC tolerance test). This 1-day delay was necessary because of the direct effects of THC during the tolerance test day.
Dependent variables
The number of wheel revolutions was recorded in 5-min bins 24 h/day for 24 days (11-day baseline, 7-day Veh/THC injection phase, 1-day tolerance assessment, and 5-day spontaneous withdrawal phase). Animals were only disturbed during the hour prior to the beginning of the dark phase (weighing, animal care, and injections) and again 6 h into the dark phase for the 7 days of Veh/THC injections. Body weight was measured every day. Rats were euthanized with isoflurane following the final day of withdrawal assessment.
Statistics
Four aspects of wheel running were analyzed: (1) Number of wheel revolutions during the dark phase; (2) Number of wheel revolutions during the light phase; (3) The percentage of total running that occurred during the dark phase; and (4) Maximum running speed achieved each day, defined as the maximum number of wheel revolutions during a single 5-min bin. Running during the light phase was not distributed normally so is presented as the median, not the mean. All other data are presented as mean ± SEM. Light phase data underwent Aligned Rank Transformation (ART) so data could be analyzed with a repeated measures ANOVA. 14 The other wheel running and body weight data did not require transformation prior to analysis with 2- and 3-way ANOVAs (sex × THC × day). Sex differences in dark-phase wheel running and body weight required converting data to a percent of baseline because baseline values were significantly different in male and female rats. Light-phase wheel running was too low during the baseline period to accurately convert data to percent of baseline (e.g., three rats had fewer than 10 wheel revolutions during 10-h of baseline data collection). Data were only analyzed for the first 10 h of the 12-h light phase because running increased dramatically immediately prior to and while experimenters were in the animal room to provide animal care and weigh rats. The percent of running during the dark phase was determined by dividing the number of wheel revolutions during the dark phase by the number of wheel revolutions averaged over the last 2 days of the baseline period. Average hourly running was used to calculate the percent of running that occurs during the dark phase to control for the 12 and 10 h of data collection during the dark and light phases. The percent of dark running was calculated for each rat and then averaged. Data were analyzed separately for each phase of the experiment: baseline, Veh/THC administration, THC tolerance, and THC withdrawal. Statistical significance was defined as a probability of less than 5%.
Results
Baseline data
A significant increase in wheel running occurred over the first 11 days (day: F(10,300) = 41.467, p < 0.001). During the dark phase of the circadian cycle, when running is highest, female rats were significantly more active than male rats (Fig. 1A) as indicated by a significant main effect of sex (F(1,30) = 25.703, p < 0.001). This sex difference increased from day 1 to 11 as shown by a significant sex × day interaction (F(10,300) = 10.265, p < 0.001). Wheel running during the light phase (Fig. 1B) was low and variable and did not differ significantly between male and female rats (F(1,30) = 0.118, p = 0.734). On day 1, about 80% of daily running occurred during the dark phase (Fig. 1C). As dark-phase running increased over days, the percentage of running that occurred during the dark phase also increased such that by day 8 and beyond, approximately 90% of total running occurred during the dark phase (day: F(10,300) = 10.232, p < 0.001). There was no significant sex difference in the percent of running occurring during the dark phase (F(1,30) = 1.258, p = 0.271). Maximum running speed (Fig. 1D) increased across days (F(10,300) = 108.215, p < 0.001). The mean maximum running speed was significantly higher in female compared to male rats (F(1,30) = 6.659, p = 0.015).
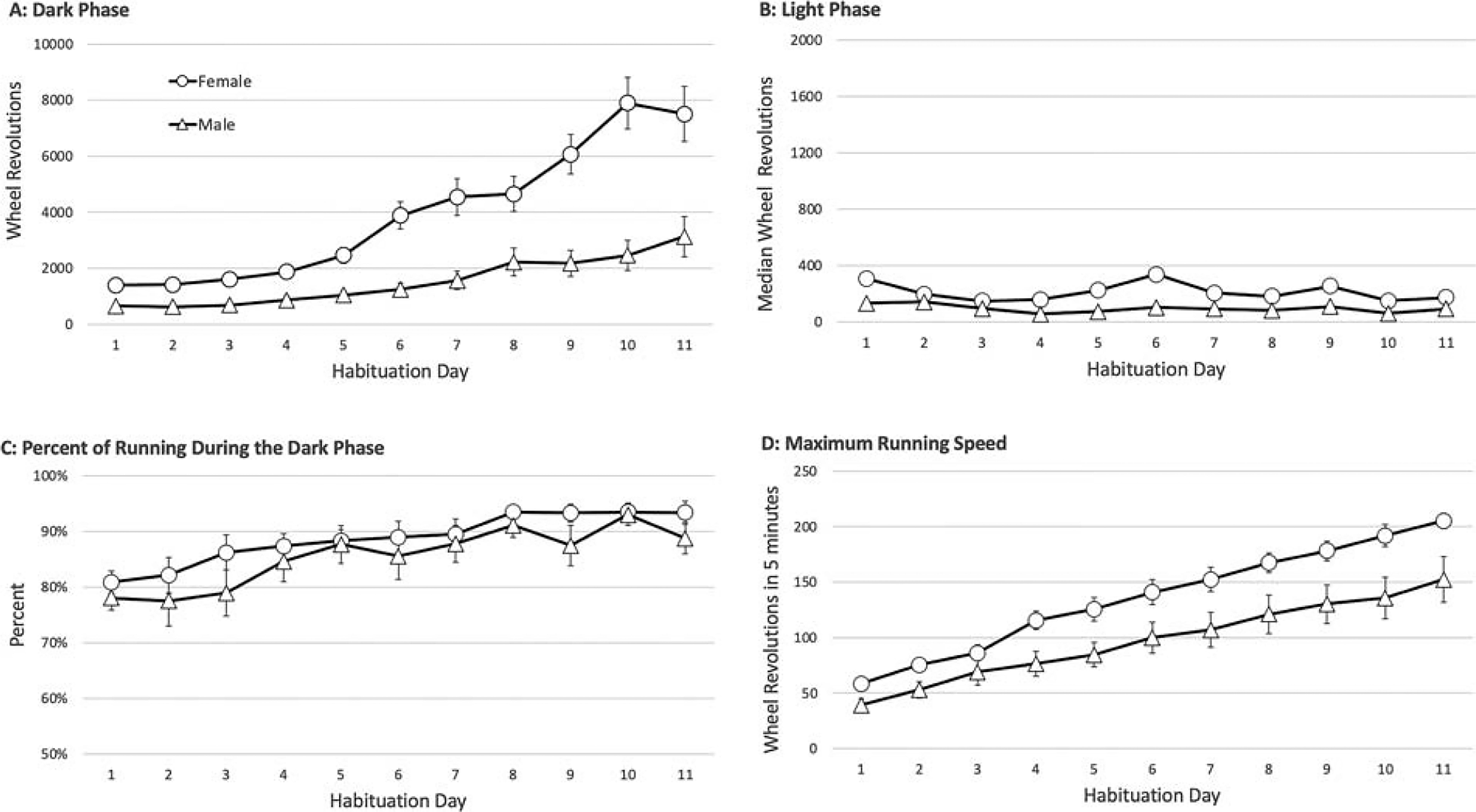
Baseline wheel running is greater in female compared to male rats.
THC injections
Twice daily injections of THC suppressed wheel running for the entire 12 h dark phase. Figure 2A shows that baseline running is highest during the first 3 h of the dark phase and gradually decreases across the dark phase for both female and male rats. This pattern is maintained in rats injected with vehicle, but not in those injected with THC (Fig. 2B). The first two injections of THC caused an almost complete suppression of wheel running for the entire dark phase in female (F(1,14) = 44.271, p < 0.001) and male (F(1,14) = 10.829, p = 0.005) rats. Some running was evident at the beginning of the dark phase because of the slow onset of effects following subcutaneous administration of THC. The baseline pattern of wheel running was reinstated during THC withdrawal (Fig. 2C).
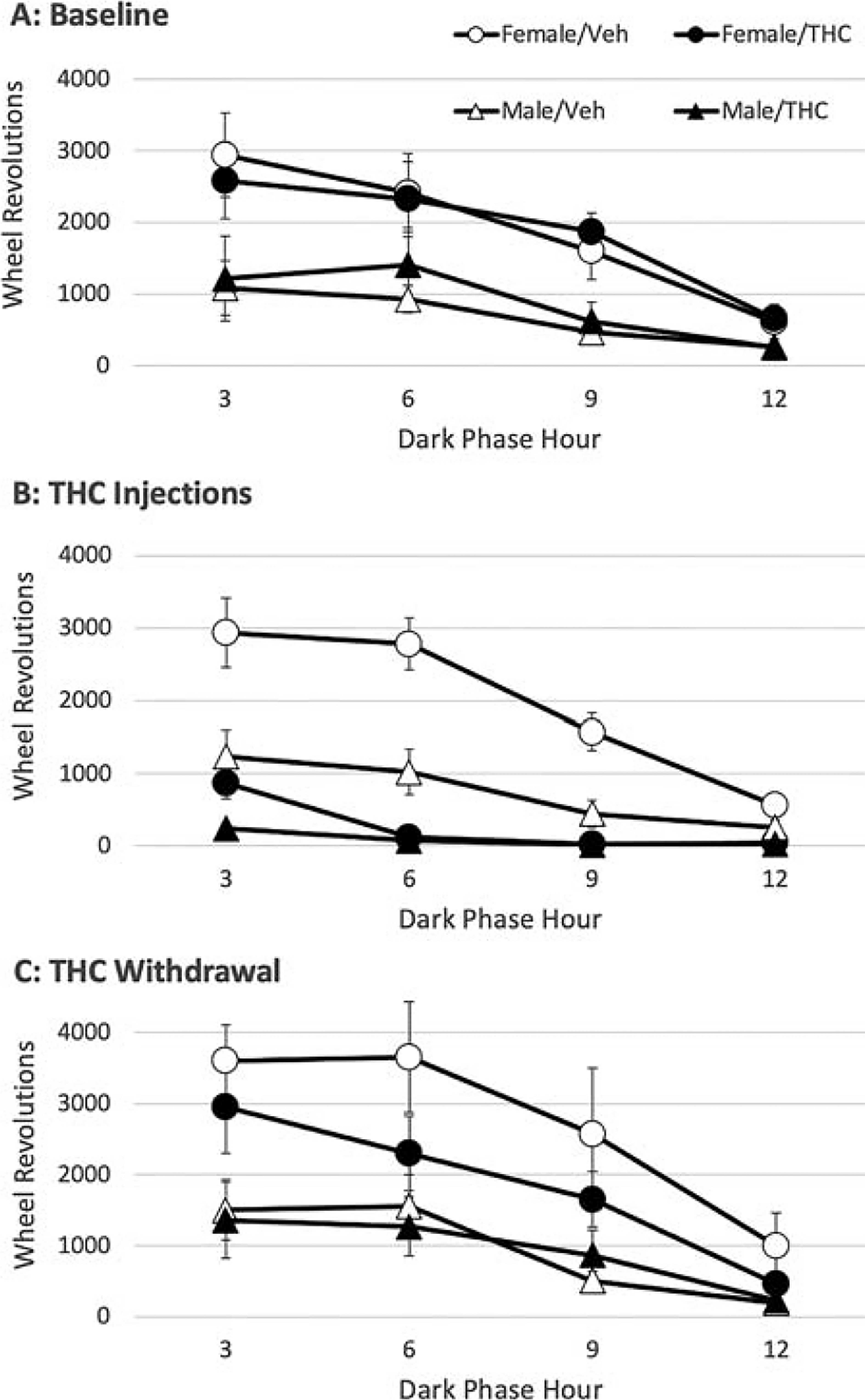
THC injections disrupt the time course for wheel running.
The decrease in dark phase running caused by THC administration compared to vehicle-injected controls was evident on all 7 days (Fig. 3A) (THC: F(1,28) = 53.389, p < 0.001). The decrease in running caused by THC injections was comparable in female and male rats, as evident by the lack of a significant main effect of sex (F(1,28) = 0.004, p = 0.950). THC did not significantly affect running during the light phase (F(1,28) = 0.907, p = 0.349), which began 6 h after the second THC injection (Fig. 3B). The large decrease in dark phase running caused by THC resulted in a significant decrease in the percent of running that occurred during the dark phase, to 80% and lower (Fig. 3C) (F(1,28) = 14.821, p < 0.001). This decrease in percent of dark phase running did not significantly differ between male and female rats (F(1,28) = 2.366, p = 0.135) and was relatively consistent across the 7 days of treatment (THC × day: F(6,168) = 1.572, p = 0.158). Administration of THC also significantly reduced maximum running speed (Fig. 3D) (F(1,28) = 23.7676, p < 0.001). Analysis of running speed as a percent of baseline revealed no significant sex difference (F(1,28) = 0.178, p = 0.676).
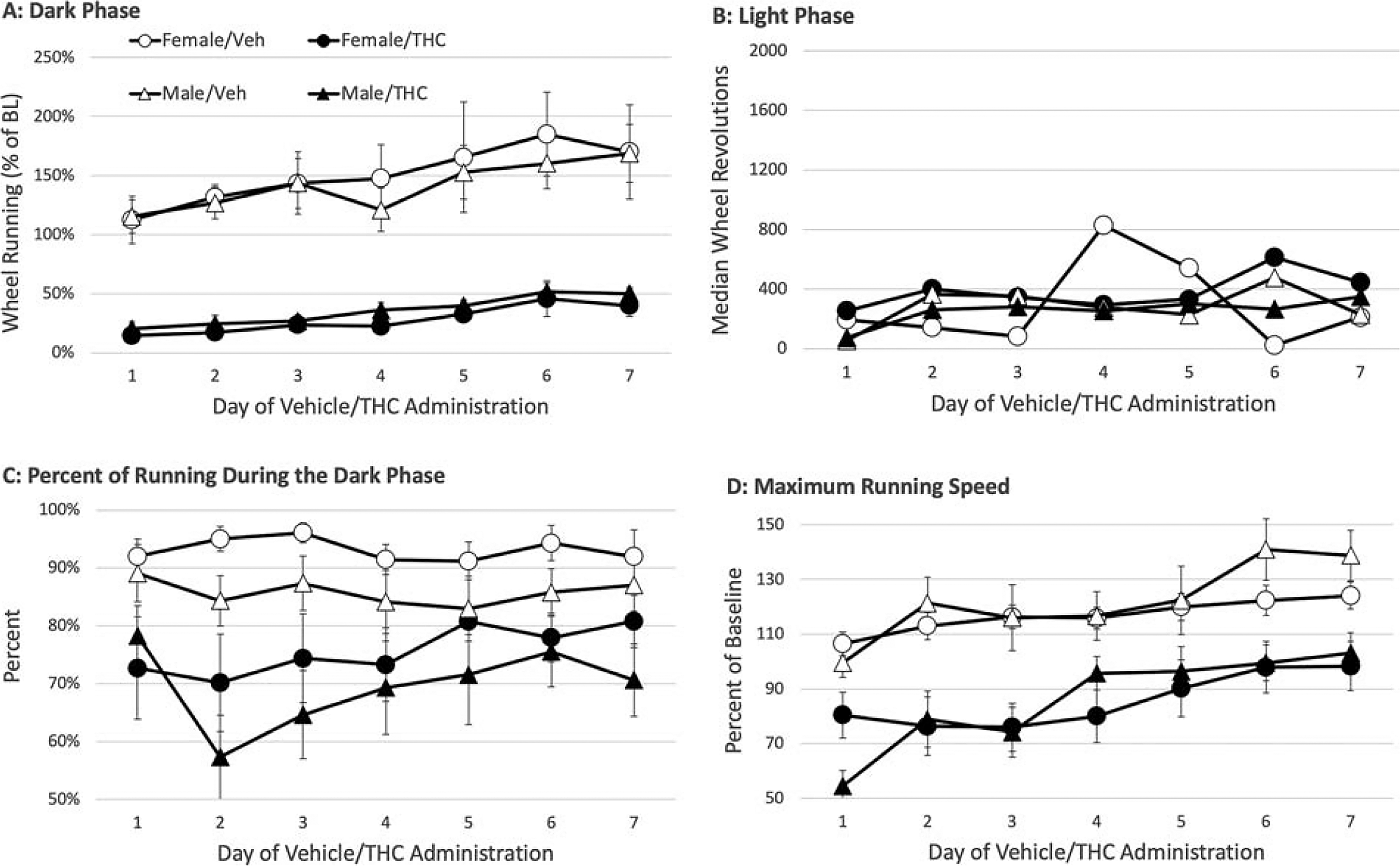
THC administration produced similar reductions in wheel running in male and female rats compared to vehicle treated rats. THC was injected twice daily, once immediately prior to the dark phase and again 6 h into the dark phase for 7 consecutive days.
Tolerance to THC
Tolerance was assessed by injecting all rats with THC immediately prior to the dark phase on the day after the last day of twice-daily THC or vehicle injections (two female rats in each condition did not receive this injection and were excluded from this analysis). In drug-naïve rats (i.e., rats previously injected with vehicle), THC administration reduced wheel running for the entire 12 h dark phase in both male and female rats (Fig. 4A). An overall analysis of dark phase running shows that THC decreased running to less than 20% of baseline levels in drug-naïve rats (Fig. 4B). THC suppressed running much less in rats previously treated with THC, resulting in a significant main effect of THC on wheel running (F(1,24) = 19.010, p < 0.001). Tolerance to THC was comparable in male and female rats, as indicated by the lack of a significant main effect of sex (F(1,24) = 1.630, p = 0.214).
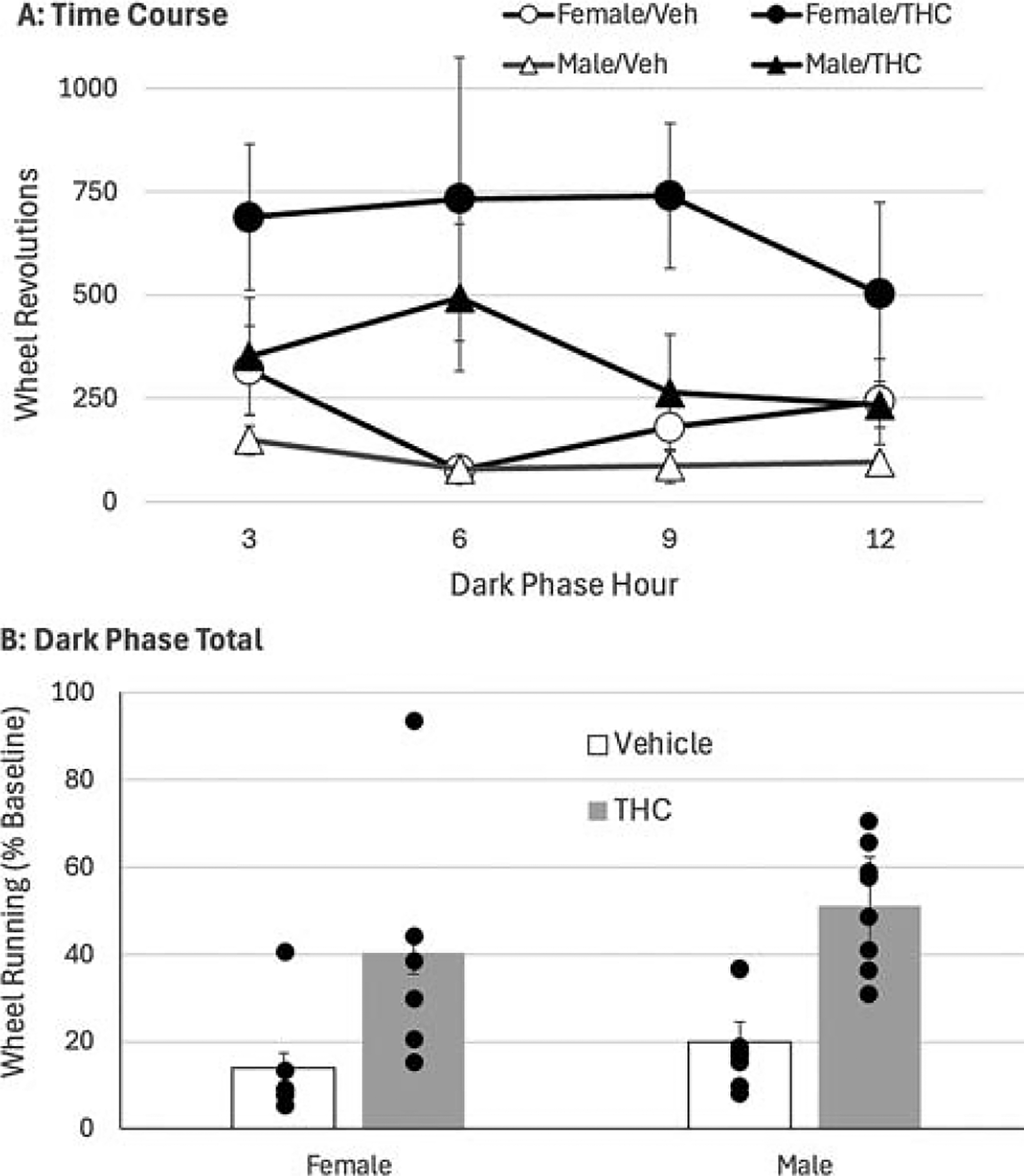
Repeated THC injections produce comparable tolerance to the locomotor effects in male and female rats. All rats were injected with THC (3 mg/kg) on the day following 7 days of twice daily injections of THC or vehicle.
THC withdrawal
There was no significant effect of THC withdrawal on any of the running parameters (Fig. 5A). Although there was a slight decrease in dark phase running, an overall ANOVA on the main effect of THC shows that this decrease did not reach statistical significance (F(1,28) = 2.071, p = 0.161). Moreover, there was no significant difference in dark phase running between male and female rats across the 5 days of withdrawal assessment (sex: F(1,28) = 2.264, p = . 144). THC withdrawal had no effect on wheel running during the light phase (Fig. 5B) (THC: F(1,28) = 0.747, p = 0.395). Likewise, THC withdrawal had no effect on the percent of running that occurred during the dark phase (F(1,28) = 0.043, p = 0.838). Ninety percent of running occurred during the dark phase for the majority of rats (Fig. 5C). Finally, withdrawal from THC did not significantly affect maximum running speed (Fig. 5D) (THC: F(1,28) = 0.469, p = 0.499). THC withdrawal appears to cause a decrease in maximum running speed on day 2, but even this decrease did not reach statistical significant (F(1,28) = 3.655, p = 0.066).
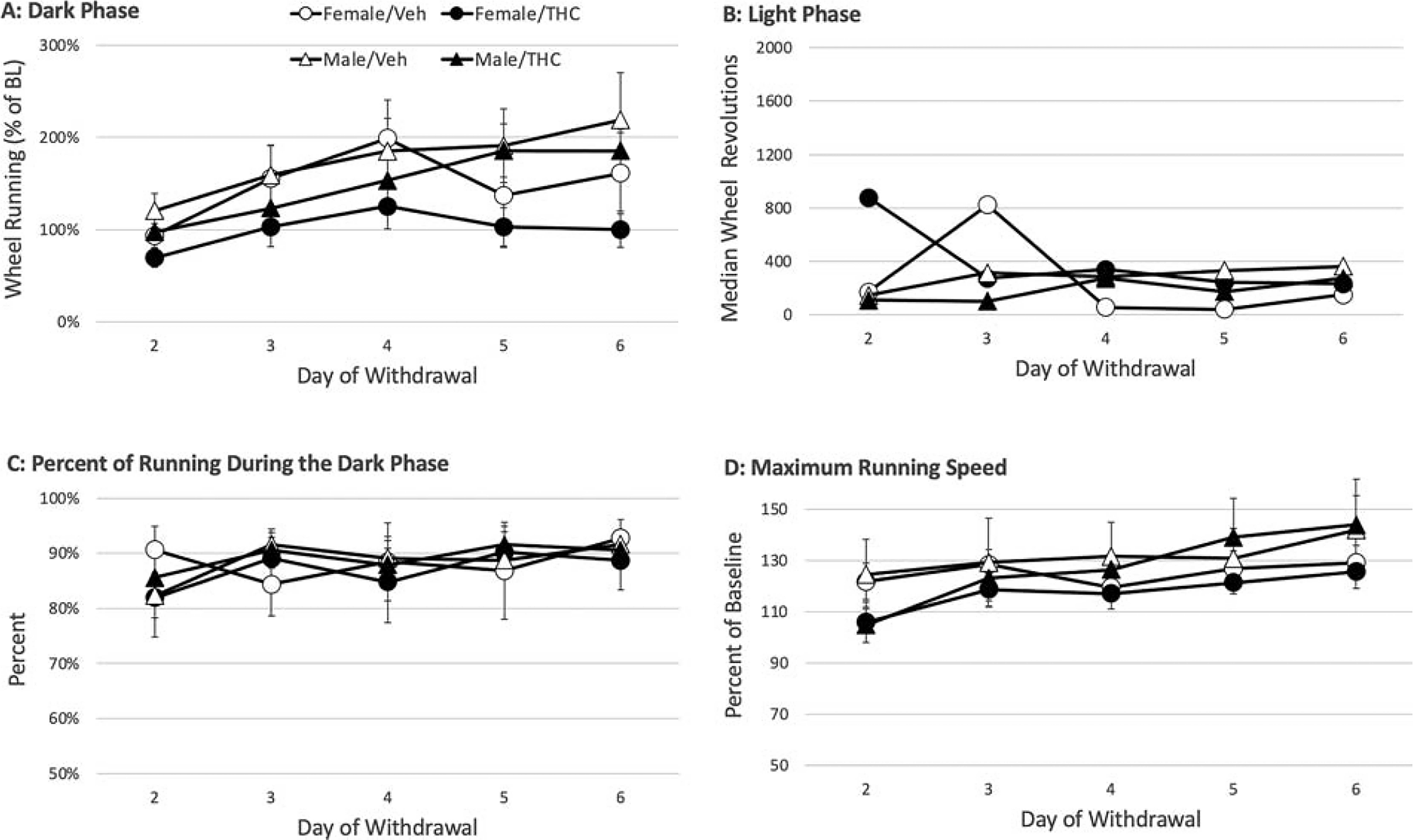
Assessment of spontaneous THC withdrawal using home cage wheel running. THC withdrawal had no consistent effect on dark
Body weight
Baseline body weight was significantly higher in male (266 ± 3.1 g) compared to female rats (186 ± 2.3 g) (t(30) = 20.609, p < 0.001). Thus, sex comparisons were based on analysis of percent of baseline data. Body weight significantly increased across the 7 days of injections (day: F(6,168) = 15.109, p < 0.001), but weight gain was reduced in rats receiving repeated THC injections compared to those receiving vehicle injections (THC: F(1,28) = 10.842, p = 0.003) (Fig. 5A). Body weight loss caused by THC administration was delayed by a day, resulting in a significant day × THC interaction (F(6,168) = 6.290, p < 0.001). Although THC-induced body weight decreases were greater in male compared to female rats (Fig. 6A), this difference did not reach statistical significance (Sex: F(1,28) = 2.531, p = 0.123).
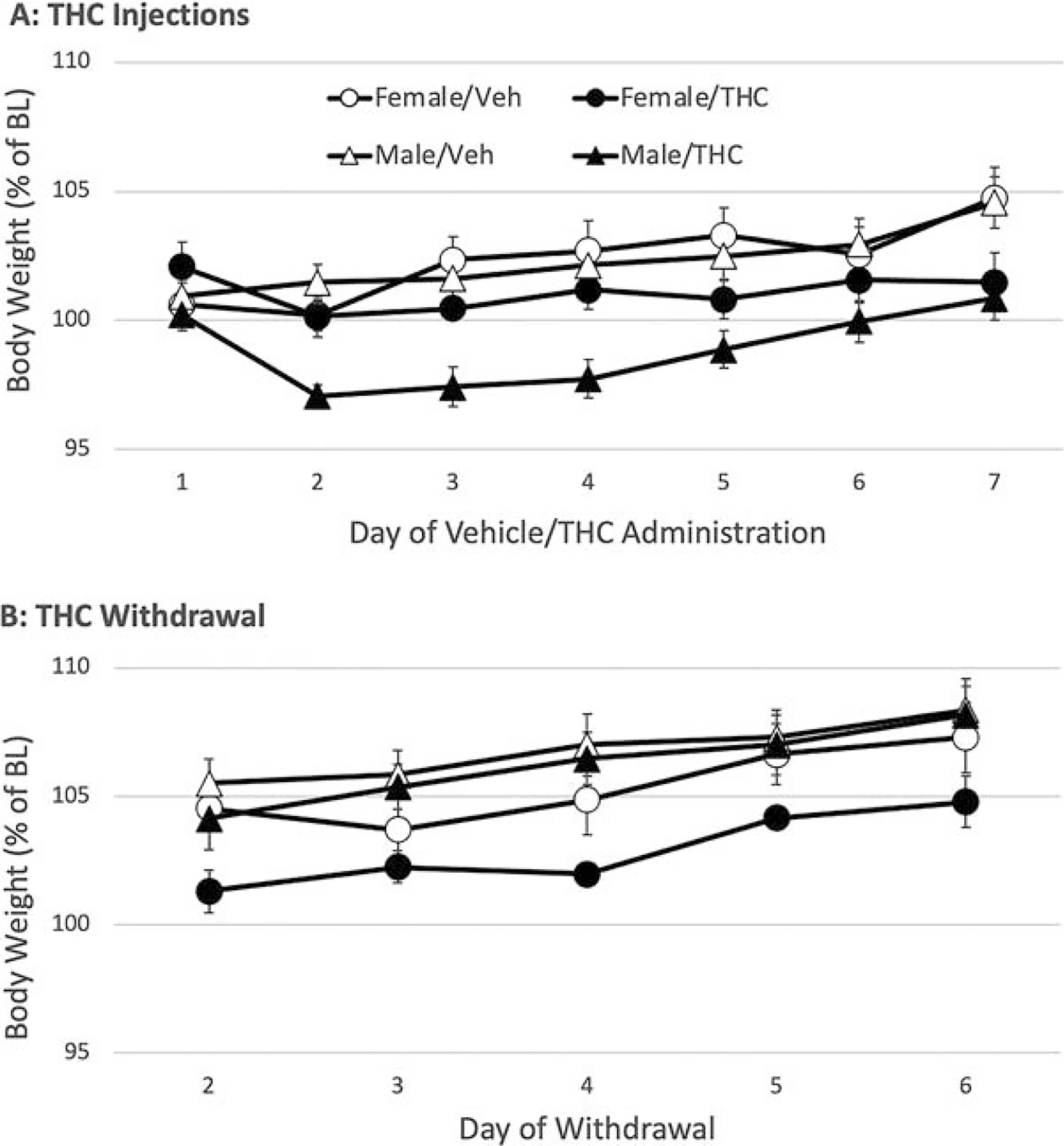
Effect of repeated THC administration and spontaneous withdrawal on body weight.
Spontaneous withdrawal from THC had no significant effect on body weight (Fig. 6B) (THC: F(1,28) = 2.558, p = 0.121). Male rats gained significantly more weight than female rats (Sex: F(1,28) = 6.043, p = 0.02), but this increase was independent of prior THC administration.
Discussion
The three main findings of this experiment are: (1) administration of THC profoundly suppressed wheel running; (2) tolerance developed to this suppression; and (3) spontaneous withdrawal from THC had no obvious effect on wheel running. These effects were similar in male and female rats.
Relatively few animal studies have assessed spontaneous withdrawal to THC (see Table 1). The studies that have been conducted report relatively minor withdrawal symptoms. One challenge is that withdrawal from THC can be prolonged—making it difficult to identify the optimal time point at which to examine withdrawal symptoms. In humans, peak withdrawal symptoms occur between 2 and 6 days following the last THC administration. 7 Animal research results seem consistent with this time frame.5,6 Our analysis covered 6 days from the last THC injection and found no consistent changes in wheel running. Both human 2 and animal research (Table 1) indicate that THC withdrawal symptoms are mild when compared to rodent experiments in which withdrawal is precipitated by administration of a cannabinoid receptor antagonist.3,15
One of the advantages of wheel running as a method to assess withdrawal is that it provides a continuous assessment for as many days as desired. Our previous use of wheel running to assess withdrawal from opioids such as fentanyl and morphine revealed a decrease in dark phase running that had a rapid onset and persisted for 2–3 days.11,16,17 THC has a much longer half-life than morphine, so the onset of THC withdrawal would be expected to be delayed. Acute subcutaneous administration of 3 mg/kg of THC produced a decrease in wheel running that persisted for the entire 12 h dark phase, requiring assessment of withdrawal to begin on the second day following the last injection (i.e., beginning 24 h after the injection). The small decrease in maximum running speed on day 2 in male and female rats undergoing THC withdrawal was the only indication that withdrawal might begin 24 h after termination of THC administration. Another advantage is that wheel running provides an objective measure of withdrawal in which data are collected in the rat’s home environment, thereby avoiding the confound of handling.
A potential problem with using wheel running as a measure of withdrawal is that running is rewarding.18–22 Rats may increase running to counteract the negative effects of withdrawal. Our previous research showing dramatic decreases in wheel running in rats undergoing opioid withdrawal suggests that any such compensation is minor if it occurs at all. Moreover, it must be emphasized that rodents are naturally very active, including voluntarily running on a wheel placed in a forest. 23 The fact that most experiments confine rats to a cage that greatly limits movement is probably a bigger confound than allowing rats to run.
Our previous experiments examining opioid withdrawal found two major changes in wheel running. A consistent and large decrease in dark-phase running and an increase in wheel running during the light phase.17,24 The increase in light-phase running corresponded with insomnia, a well-known symptom of drug withdrawal. In contrast, the present experiment found no significant decrease in dark-phase running or increase in the light-phase running during THC withdrawal. It is possible that wheel running is not an appropriate tool to assess THC withdrawal, but this seems unlikely given that wheel running is sensitive to a wide range of conditions that disrupt quality of life.11,25 Increasing the sample size might result in a statistically significant effect on data that is close to statistical significance (e.g., running speed on day 2 of withdrawal; see Fig. 5D), but is unlikely to move the data points. Even if a larger sample size resulted in statistical significance, the present results indicate that the magnitude of spontaneous THC withdrawal is mild at best.
Although there are many potential reasons for the lack of withdrawal seen in the present study, wheel running data are not subject to experimenter bias when compared to experiments in which withdrawal symptoms are rated by a researcher. It is noteworthy that our dose of THC (3 mg/kg) was significantly lower than the doses of 10–50 mg/kg given to mice and rats in previous experiments examining spontaneous withdrawal (Table 1). High THC doses seem to produce more pronounced withdrawal symptoms in humans than lower doses.26,27 The same dose relationship would be expected in animals. However, only in comparison to these other rodent studies would our dose be considered low. A single THC injection of 3 mg/kg on the tolerance test day reduced dark phase running to 20% of baseline levels. Our previous research shows that this dose is 10 times higher than the 0.32 mg/kg dose that does not depress wheel running.10,13
Repeated administration of 3 mg/kg of THC produced tolerance to THC-induced suppression of wheel running. However, tolerance was incomplete in that even after 15 injections, administration of THC still decreased wheel running by over 50%. This decrease was significantly less than the 80% decrease in rats receiving THC for the first time. Tolerance to the antinociceptive effects of THC has been documented in both humans 28 and rodents.29–32 The magnitude of tolerance to the antinociceptive effect of THC has been reported to be greater in female compared to male rats.30–32 This sex difference may be specific to antinociception given that we, and others 32 found no sex difference in tolerance to the THC-induced decrease in locomotion. 32 Of course, it is also possible a sex difference in tolerance was not evident at the dose we used (3 mg/kg), but may have been evident with a full dose-response analysis.
Sex differences in the acute effects of cannabis have been well studied. The rodent literature indicates heightened sensitivity to the antinociception,30,32,33 anxiety, 34 and hypothermic 31 effects of THC. It is not clear how well these effects generalize to humans, where sex differences seem less pronounced, especially at lower doses,35–37 although high doses produce more adverse effects in women than men.36,38 Our finding that THC produced a comparable decrease in voluntary wheel running in male and female rats aligns with much of the human data on acute effects of THC.
In contrast to wheel running, changes in body weight showed trends toward sex differences. Male rats lost more weight than female rats during repeated THC injections. The greater decrease in body weight in male compared to female rats receiving repeated THC administration is consistent with preclinical39,40 and clinical data. 41 In contrast, female rats lost more weight than males during THC withdrawal (see Fig. 6B).
Conclusion
Our data are consistent with previous research showing that acute administration of THC causes a decrease in locomotion and body weight, tolerance with repeated THC injections, and minor or no symptoms of spontaneous THC withdrawal. Although a number of studies report enhanced effects of THC in female compared to male rodents, we found no obvious sex differences in the effects of THC on wheel running to the acute effects, tolerance, or withdrawal.
Authors’ Contributions
C.M.H. contributed to experimental design, data collection, and article writing. C.P., W.G., and A.C. contributed to data collection, editing, and data preparation. M.M.M. contributed to experimental design, data analysis, and article preparation.
Footnotes
Acknowledgments
Thank you to Faith Spencer for assistance with data collection.
Funding Information
Funding provided by Washington State Measure No. 171.
Disclosure Statement
No competing financial interests exist.