
Abstract
Introduction:
The endocannabinoid system regulates a broad range of psychological and physiological processes throughout the body, and its dysregulation has been implicated in numerous pathological conditions. As a result, components of this system are common targets in diagnostic and therapeutic research. Given the challenges associated with obtaining tissue samples from human participants, current research commonly relies on the quantification of circulating endocannabinoids in blood as a surrogate for tissue-level endocannabinoid activity. However, this approach presents limitations due to the invasive nature of blood collection and its potential to induce pain or stress-related activation of the endocannabinoid system. New methods to study the activity of the endocannabinoid system in humans are therefore needed.
Methods:
This narrative review examines the current literature investigating the measurement, origin, and significance of the endocannabinoid ligands N-arachidonoylethanolamide (AEA) and 2-arachidonoylglycerol (2-AG) in human saliva, a non-invasive and easily accessible biological matrix.
Results:
Mass spectromic analysis have demonstrated that AEA and 2-AG levels are dynamic and can be influenced by physiological challenges, including fear, stress, and exercise. Salivary endocannabinoid profiles have also been shown to reflect longer-term traits, such as metabolic status, and can distinguish between patients and healthy individuals in conditions like orofacial pain.
Discussion:
Collectively, salivary endocannabinoids reflect both acute and chronic aspects of endocannabinoid system activity and may represent a promising non-invasive matrix for assessing endocannabinoid function. However, further understanding of the mechanisms underlying endocannabinoid production in the oral cavity is required for accurate interpretation of salivary measurements in future studies.
Introduction
N-arachidonoylethanolamide (AEA) 1 and 2-arachidonoylglycerol (2-AG) 2 are bioactive lipids that serve as endogenous ligands of the endocannabinoid system. The endocannabinoid system also encompasses the two G protein-coupled receptors, cannabinoid receptor 1 (CB1) and cannabinoid receptor 2 (CB2),3,4 as well as a network of enzymes that modulate endocannabinoid concentrations across various tissues (Fig. 1). Although both comprised of from arachidonic acid, AEA and 2-AG have distinct metabolic pathways. AEA is primarily synthesized and degraded upon the expression of N-acyl-phosphatidylethanolamine-specific phospholipase D (NAPE-PLD) 5 and fatty acid amide hydrolase (FAAH), 6 respectively, while 2-AG is synthesized under the expression of diacylglycerol lipase (DAGL) α and β, 7 and degraded upon the expression of monoacylglycerol lipase. 8 However, the regulation of endocannabinoid signaling is complex, involving not only these primary metabolic routes but also other enzymes such as cyclooxygenase-2, N-acylethanolamine acid amidase, and α/β-hydrolase domain enzyme-6. 9 Furthermore, endocannabinoid-like ligands, such as oleoylethanolamide (OEA) and palmitoylethanolamide (PEA), are also often co-synthesized with AEA. These N-acylethanolamines (NAEs) potentiate endocannabinoid effects without binding to the classical cannabinoid receptors. 10
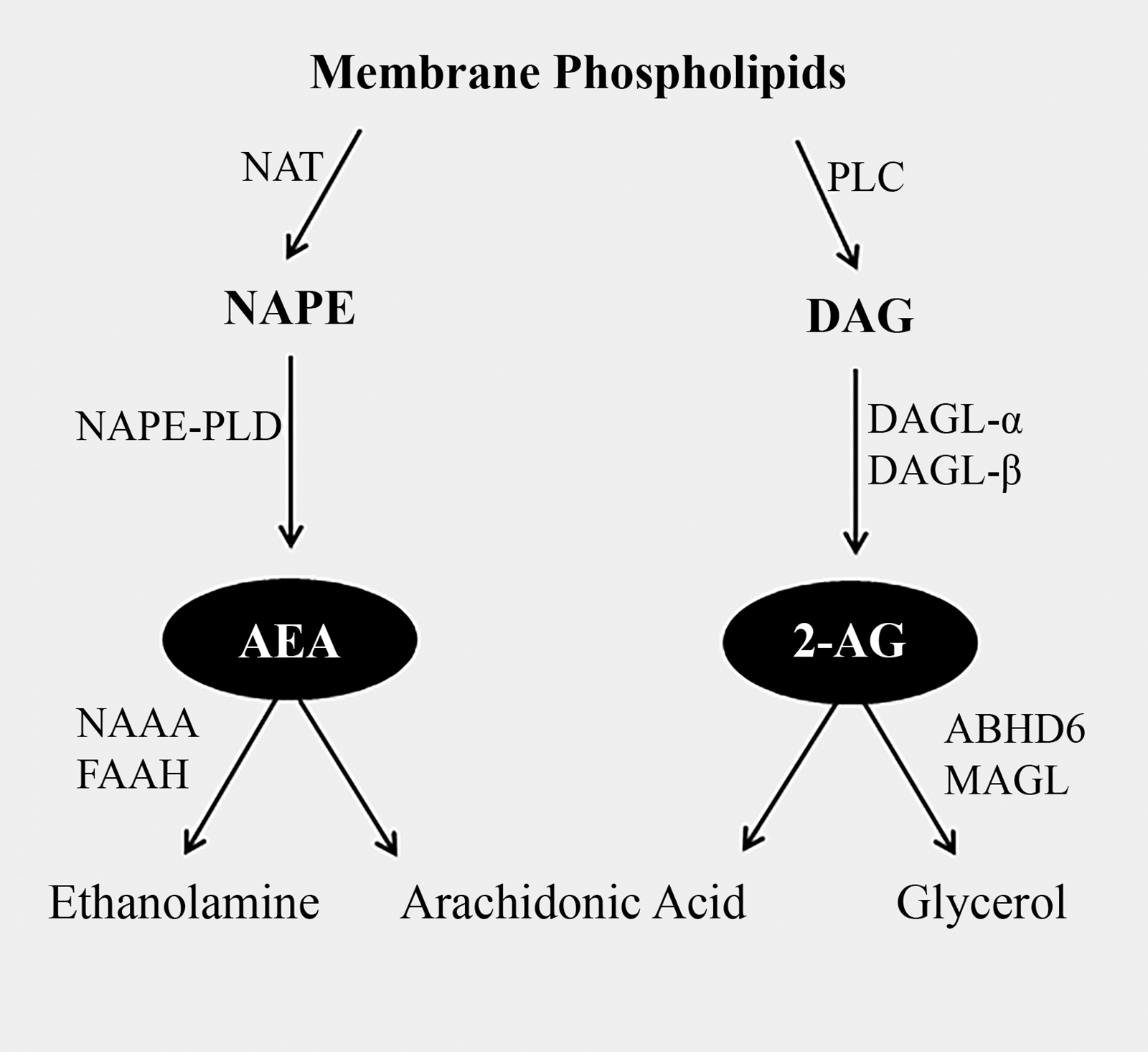
Classical enzymatic pathway for the synthesis and degradation of N-arachidonoylethanolamide (AEA) and 2-arachidonoylglycerol (2-AG). DAG, diacylglycerol; DAGL-α, diacylglycerol lipase alpha; DAGL-β, diacylglycerol lipase beta; FAAH, fatty acid amide hydrolase; MAGL, monoacylglycerol lipase; NAPE, N-acylphosphatidylethanolamine; NAPE-PLD, N-acyl phosphatidylethanolamine phospholipase D; NAT, N-acyltransferase; PLC, phospholipase C.
CB1 receptors are abundantly expressed on the presynaptic membranes of neurons throughout the central nervous system (CNS). Here, binding of AEA or 2-AG inhibits the release of both excitatory and inhibitory neurotransmitters by modulating intracellular calcium ion signaling. 11 Through this mechanism, endocannabinoids regulate a range of psychological functions, including emotion, 12 learning and memory,13,14 as well as reward and addiction 15 (Fig. 2). In addition to the CNS, endocannabinoid receptors are also expressed in various peripheral tissues, where receptor activation initiates distinct intracellular signaling pathways depending on the localization.16,17 The endocannabinoid system therefore also plays a vital role in regulating numerous physiological processes, including energy homeostasis and metabolism, 18 pain modulation, 19 inflammation, 20 the stress response,21–23 sleep regulation, 24 and cardiovascular function. 25 Due to its critical role in regulating these diverse processes, dysregulation of the endocannabinoid system is often found in association with many pathological conditions. This makes components of the endocannabinoid system potentially valuable as therapeutic targets or diagnostic tools.
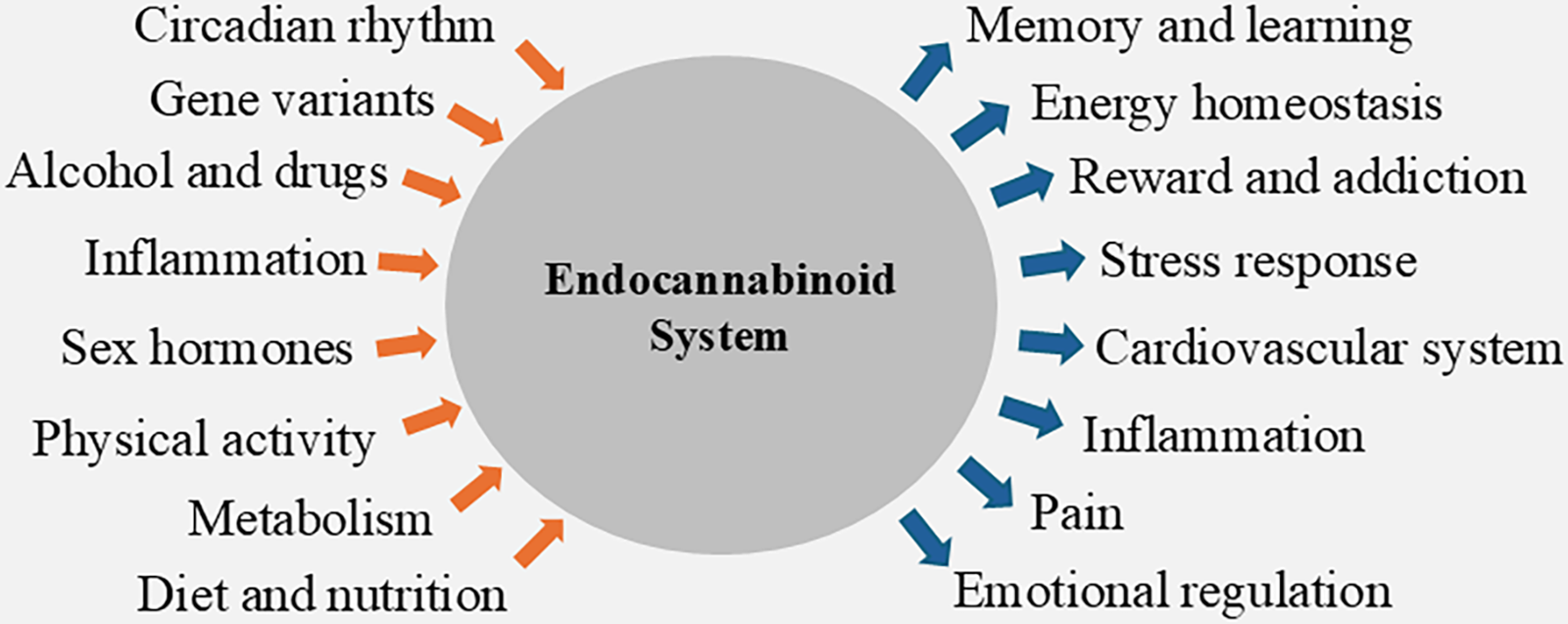
Factors modulating (orange arrows) and modulated by (blue arrows) the endocannabinoid system.
Studying endocannabinoid responses in humans is challenging due to the difficulty of obtaining tissue samples, particularly from the CNS. However, endocannabinoid ligands can cross cellular membranes and enter surrounding biological matrices, therefore reflect endocannabinoid tone from nearby tissues.26–28 Among biofluids, venous blood is commonly used to study the endocannabinoid system, 29 and endocannabinoid concentrations in blood can differentiate between patients and controls in those with various diseases. For example, endocannabinoids show different profiles in participants with psychiatric disorders such as post-traumatic stress disorder (PTSD)14,30,31; neurodegenerative diseases, including Alzheimer’s disease 32 ; pathological conditions like hypertension 33 ; as well as pain disorders such as osteoarthritis. 34 However, while many studies report significant differences in endocannabinoid levels, the direction and magnitude of these changes vary between studies.29,35 This inconsistency may be due, in part, to the autocrine and paracrine nature of endocannabinoid signaling, which means that circulating levels likely represent a merged profile of local signaling events from multiple tissues, making it challenging to obtain consistent responses. Blood sample collection also requires venipuncture, which can activate nociceptive and stress systems and may therefore incite endocannabinoid responses,23,36 likely in a highly variable way across participants. 37 Identifying more accessible and localized methods for assessing the endocannabinoid system might therefore be useful for gaining more consistent results.
Recent advancements in highly sensitive liquid chromatography–tandem mass spectrometry (LC-MS/MS) techniques have enabled the quantification of low concentrations of endocannabinoids in saliva.38,39 Saliva sampling offers several advantages over blood sampling, including rapid and noninvasive collection, cost-effectiveness, and suitability for large-scale screening. 40 Several salivary biomolecules are already used as surrogate markers of physiological responses in place of blood-based markers. For example, salivary cortisol is commonly used as a noninvasive marker of hypothalamic–pituitary–adrenal (HPA) axis activity, 41 whereas salivary alpha-amylase serves as an indicator of sympathetic nervous system (SNS) activation. 42 Early evidence suggests that salivary endocannabinoids reflect the recent activity of the endocannabinoid system; therefore, they may provide insight into the functional state of this system and its role in health and disease. However, research in this area is still in its infancy, and many questions remain unanswered.
This review aims to examine the current evidence regarding the origins of salivary endocannabinoids and the physiological processes to which they are responsive. It will also explore factors contributing to variability in their basal concentrations and responses, guiding future studies on considerations necessary for accurate interpretation. Finally, the review will highlight key unanswered questions that must be addressed to fully elucidate the functional relevance of salivary endocannabinoids and assess their potential as biomarkers of endocannabinoid system activity.
Quantification of Salivary Endocannabinoids
Endocannabinoid concentrations in saliva are substantially lower than in blood or tissue; therefore, their quantification has relied on highly sensitive LC-MS/MS methods using triple quadrupole or high-resolution mass spectrometers.39,43 Basal salivary AEA concentrations have been reported in the low pmol/mL range, whereas 2-AG concentrations are typically 100- to 200-fold higher and more variable, falling within the high pmol/mL to low nmol/mL range38,39,43
The quantification of AEA and 2-AG within various biological matrices presents several analytical and pre-analytical challenges,44,45 with similar methodological issues likely to affect salivary measurements. Saliva sampling methods commonly used in biomarker research include the passive drool method,39,43 chew-stimulated collection (e.g., Parafilm®-induced salivation), and swab-based devices (e.g., Salivette®). 46 A single-donor study demonstrated that AEA and PEA concentrations differed between chew-stimulated saliva and saliva collected directly from the parotid gland, 47 while another study reported differences in several NAEs between parafilm-stimulated and unstimulated saliva. 48 Although most studies so far have utilized the passive drool method,39,43 the above studies demonstrate that differences in collection methodology contribute to variability and should be standardized to ensure comparable results across studies.
Post-collection handling of saliva samples has varied across studies, with storage temperatures ranging from −20 to −80°C. Most protocols include a centrifugation step (typically 3000–4000 g), performed either before or after storage, to remove cells, food debris, and other insoluble material prior to downstream processing.39,49 Endocannabinoids are subsequently extracted using liquid–liquid extraction,39,49–52 protein precipitation,38,53 or solid-phase extraction, using a variety of organic solvents, including methanol, acetone, ethyl acetate, and chloroform.47,54 Chromatographic separation has been performed using high-performance or ultra-high-performance liquid chromatography, typically employing sub-2 µm C18 columns with water and acetonitrile as mobile phases. Mobile-phase modifiers such as ammonium formate, ammonium acetate, formic acid, or acetic acid have been included to enhance ionization efficiency and improve detection of low endogenous endocannabinoid concentrations. 55
Collectively, these studies demonstrate substantial methodological heterogeneity in the quantification of salivary AEA and 2-AG. Given the lipophilicity and chemical instability of these analytes, degradation and isomerization during sample collection, processing, and storage are likely to contribute to inter-study variability.45,56,57 The development of standardized methodologies is therefore essential to improve analytical reliability and comparability across studies. Nevertheless, compared with blood, saliva contains relatively few cells, reducing the potential for ex vivo enzymatic synthesis of AEA and representing a potential analytical advantage of this matrix. 58
The Endocannabinoid System in the Oral Cavity
Endocannabinoid metabolism
AEA and 2-AG possess the ability to cross cellular membranes through passive diffusion or active transport. 59 Despite this capability, concentrations of AEA and 2-AG concentrations have not correlated between different biomatrices,60–62 including between blood and saliva.39,43 These results likely reflect the nature of endocannabinoid signaling, which is produced on demand and acts mainly in a paracrine or autocrine manner, so that concentrations in each biofluid largely reflect activity in nearby tissues. 63 This contrasts with steroid hormones such as cortisol, progesterone, and testosterone, which are produced by a small number of specialized tissues, circulate systemically, and act as classical endocrine signals with widespread effects across multiple tissues.64,65 Circulating steroid hormones enter saliva via intracellular diffusion; thus, salivary levels primarily reflect their free, biologically active concentrations present in the blood. 66
In mammals, salivary glands include the three major glands, namely, the submandibular, sublingual, and parotid glands, as well as numerous smaller accessory glands. 67 Matias and colleagues analyzed human submandibular and accessory gland homogenates from maxillofacial surgery patients and detected AEA, 2-AG, OEA, and PEA, as well as messenger ribonucleic acid encoding DAGL-α/β, NAPE-PLD, and FAAH. 39 Similarly, immunohistochemical staining of the mouse submandibular gland (SMG) has also revealed NAPE-PLD expression within myoepithelial cells, 68 while in rats, intercellular AEA levels increased in the SMG following ethanol administration. 69 Together, this evidence suggests that salivary glands serve as a site for endocannabinoid metabolism, which then subsequently determines salivary levels. The endocannabinoid tone in saliva therefore likely reflects the localized activity of the endocannabinoid system, showing its dynamic responses and interindividual differences.
Endocannabinoid function
Salivary glands are composed mainly of acinar, ductal, and myoepithelial cells, and saliva secretion begins with the transport of water and ions from the bloodstream into ducts through the acinar cells. The contraction of myoepithelial cells forces the fluid through the ducts, allowing ductal cells to modify its composition, resulting in final saliva secretion. 70 Each of these cellular types contains the receptors for both sympathetic and parasympathetic neurotransmitters of the autonomic nervous system (ANS), which allows for integrated control of saliva volume and composition. 71 Several early studies in animals have demonstrated that endocannabinoid binding to CB1 and/or CB2 receptors directly on acinar, ductal, or myoepithelial cells alters various intracellular signaling pathways, resulting in decreased saliva secretion and altered saliva composition.69,72–77 For example, Prestifilippo and colleagues discovered that AEA binding to CB1 and CB2 in the SMG of male rats reduced cyclic adenosine monophosphate via inhibition of adenylyl cyclase. 73 However, in a more recent study, CB1 receptor immunostaining was observed exclusively on cholinergic neurons innervating the mouse SMG. 68 In this study, pharmacological inhibition or genetic knockout of FAAH increased AEA levels within SMG tissue and reduced salivation, an effect attributed to decreased acetylcholine release through CB1 receptor activation. Similarly, CB1 receptor staining was identified on parasympathetic nerve axons in the mouse parotid gland, where CB1 agonist treatment inhibited both stimulated and unstimulated salivation. 78 In other tissues, endocannabinoids also interact closely with the ANS, where neuronal stimulation triggers endocannabinoid synthesis, which in turn suppresses further neurotransmitter release.79,80 These findings suggest that endocannabinoids modulate salivary secretion indirectly by regulating presynaptic neurotransmitter release, rather than acting solely through direct effects on glands. It is worth noting that endocannabinoids have also been demonstrated to influence salivary secretion centrally, acting on hypothalamic autonomic circuits that regulate parasympathetic and sympathetic outflow to the salivary glands. 76 Salivary endocannabinoid concentrations may therefore reflect the fine, real-time regulation of secretion by autonomic activity and hold promise as biomarkers of this activity in health and disease.
Interestingly, Thoungseabyoun and colleagues demonstrated that the subcellular distribution of cannabinoid receptors varies according to the type of autonomic input. 81 Specifically, sympathetic stimulation caused rapid redistribution of CB1 receptors to the apical membrane of acinar and ductal cells, whereas parasympathetic cholinergic stimulation left CB1 receptors confined to the basolateral membranes of ductal cells. Such context-dependent receptor redistribution may explain discrepancies in reported CB1 localization and function within the salivary glands. Endocannabinoids thus appear to influence salivary secretion through multiple mechanisms, through direct effects on glandular cells as well as indirect modulation of autonomic neurotransmission depending on cell type and neural input.
Although endocannabinoid metabolism has not yet been demonstrated in other oral tissues, cannabinoid receptor expression and functional activity have been documented. 82 For example, CB1 and CB2 receptors are present on epithelial cells of the tongue, 83 and administration of AEA or 2-AG in mice enhanced gustatory nerve responses and behavioral preference for sweet tastes. 84 Receptor expression has also been identified in the dental pulp and periodontal tissues, where endocannabinoids are thought to support tissue repair, cell growth, differentiation, and inflammation control. 85
Together, these findings suggest that the endocannabinoid system modulates oral functions in a complex manner through both gland-level and brain-level mechanisms. Salivary endocannabinoids may therefore provide insights into endocannabinoid system activity that are distinct from blood-based measures, underscoring the need for further research to clarify their physiological roles and potential as biomarkers.
Determinants of Salivary Endocannabinoid Concentrations
Acute effects
The endocannabinoid system is dynamically regulated by physiological challenges, with circulating endocannabinoid concentrations serving as indicators of system activity across diverse physiological states. 29 Recent findings indicate that these changes can also be detected in saliva, offering a noninvasive window into endocannabinoid system functioning.
Exercise, stress, and trauma
As part of a method validation study, Ney and colleagues demonstrated that concentrations of both AEA and 2-AG in saliva exhibit increases immediately following exercise, whereas corresponding cortisol responses were delayed. 38 This initial evidence demonstrated that physiological challenges could cause detectable changes in salivary endocannabinoid concentrations. Similarly, salivary 2-AG concentrations were significantly higher than those of a control group immediately following exposure to acute stress via the Maastricht Acute Stress Test (MAST). 43 In the same study, AEA concentrations did not differ from controls but positively correlated with both 2-AG and cortisol. In an alternative study, AEA showed significant time-dependent increases immediately after the MAST, whereas post-stress increases in 2-AG were not significant (Hargreaves et al., under review). These findings indicate that salivary AEA and 2-AG respond acutely to stress; however, further research is needed to confirm the robustness and consistency of these stress-induced changes.
The mechanisms underlying these acute changes in salivary endocannabinoid concentrations are not fully understood; however, their rapid responses suggest modulation by the SNS, which mediates the fast fight-or-flight response. 86 Supporting this link, salivary α-amylase (sAA), a surrogate marker of sympathetic activation, correlates with both AEA and 2-AG following a combined trauma film and cold pressor stress test. 87 Nevertheless, the validity of sAA as a direct marker of sympathetic activity remains debated,88,89 highlighting the need for studies employing pharmacological inhibition or examination of direct sympathetic markers to clarify the extent to which sympathetic activity has on endocannabinoid tone.
In the same study, salivary 2-AG, OEA, and PEA declined significantly following combined trauma film and cold pressor exposure, and participants who did not exhibit a cortisol increase reported more intrusive memories—an effect moderated by AEA levels. 87 Moreover, salivary endocannabinoid concentrations have also been found to be associated with subjective stress, with higher distress correlating with lower levels of AEA and 2-AG after trauma film exposure. 62 As trauma film paradigms are commonly used to induce intrusive memories and model the etiology of PTSD, 90 salivary responses may therefore reflect CNS functionality involved in trauma memory formation.
Collectively, these findings highlight that salivary endocannabinoid levels are sensitive to not only physiological but also psychological states or activity. Their capability as accessible markers of both peripheral CNS functions and pathologies should therefore also be explored in further studies.
Food/alcohol intake
Despite the endocannabinoid system also playing a multifactorial role in metabolic regulation, 18 Matias and colleagues reported that AEA and 2-AG concentrations did not exhibit significant differences prior to and 1 h following a meal. 39 Moreover, postprandial increases in endocannabinoids and other NAEs discovered in separate studies were attributed to their release from food, as opposed to localized synthesis in the oral cavity.50,54 Tarragon and colleagues detected a significant link between highly processed food intake and elevated salivary AEA levels in women; however, no correlation was found between fatty acid intake and salivary concentrations of arachidonic acid. 91 Instead, arachidonic acid levels predicted waist circumference, suggesting that long-term, rather than immediate, dietary patterns influence metabolic status and consequently salivary endocannabinoid levels. Together, salivary endocannabinoid levels appear largely insensitive to acute food intake, a property that enhances their utility as biomarkers by reducing potential confounding from recent eating. Alternatively, ethanol administration has been demonstrated to increase AEA synthesis in the SMG of rats, 69 suggesting that alcohol consumption may acutely influence salivary endocannabinoid levels. However, this result remains to be confirmed in humans.
Testosterone administration
Wu and colleagues investigated acute salivary endocannabinoid changes following exogenous testosterone administration in humans. 52 While salivary AEA concentrations did not exhibit significant changes, they demonstrated that significant alterations were observed in the AEA/cortisol ratios, highlighting the disruption to the normal balance between the endocannabinoid system and HPA axis. These findings suggest that using analytical techniques that can examine the dynamic interactions between multiple biomolecules has the potential to offer additional insights into physiological regulation and endocannabinoid function.92,93
Pathological conditions
The endocannabinoid system becomes dysregulated across a variety of pathological conditions. 94 Emerging evidence suggests that such dysregulation is also reflected in salivary endocannabinoid concentrations, with several studies demonstrating altered levels in clinical populations so far.
Matias and colleagues discovered that obese, insulin-resistant individuals exhibited significantly elevated salivary concentrations of AEA, 2-AG, OEA, and PEA compared with normal-weight controls. 39 Interestingly, these elevations in saliva did not uniformly extend to plasma, as corresponding increases in 2-AG and OEA in obese individuals were not significant in this matrix. Furthermore, the difference between AEA and 2-AG concentrations in obese individuals versus controls was also significantly greater in saliva than in plasma, indicating that salivary endocannabinoid profiles are not only unique compared to those in plasma but also more sensitive to certain physiological differences.
Moreover, significantly lower levels of salivary 2-AG and OEA were observed in patients with orofacial pain compared with pain-free control subjects, 49 demonstrating that alterations in the endocannabinoid system associated with pain disorders may be observable in salivary endocannabinoid profiles. In a follow-up study, Heiliczer and colleagues focused on salivary endocannabinoids across distinct orofacial pain disorders. 95 While the sample sizes for these orofacial pain subgroups were relatively small, the findings revealed significantly decreased levels of 2-AG in patients with trigeminal neuralgia and tension-type headaches, alongside elevated AEA levels in individuals diagnosed with burning mouth syndrome. This research highlights the heterogeneity of salivary endocannabinoid profiles among patient subgroups, highlighting their potential utility in profiling specific disorders.
Although the mechanisms underlying these changes remain unclear, both obesity and orofacial pain disorders are associated with heightened inflammation. This may, in turn, contribute to alterations in salivary endocannabinoid levels, given the established role of the endocannabinoid system in regulating inflammatory processes. 20 In rats, local and systemic administration of lipopolysaccharide, a bacterial endotoxin, increased AEA concentrations in SMG tissue, an effect mediated by elevated tumor necrosis factor-alpha.75,96 This study demonstrated that endocannabinoid concentrations in the salivary glands respond not only to local inflammatory conditions but also to systemic inflammatory signals. Salivary endocannabinoid levels may therefore not only mirror local pathological changes like orofacial pain but might also serve as accessible biomarkers reflecting broader inflammatory conditions throughout the body. Therefore, the sensitivity of salivary endocannabinoids to inflammation remains to be investigated further, especially since inflammation is increasingly implicated in the pathophysiology of various diseases. 97
Collectively, these findings suggest that, due to their sensitivity to multiple systemic, central, and local signals, salivary endocannabinoids may reflect unique pathophysiological profiles. Further investigation into the relationship between salivary endocannabinoids and various pathological disorders is therefore needed to determine their utility.
Interindividual differences
The endocannabinoid system interacts with multiple physiological pathways, contributing to substantial interindividual variability in basal endocannabinoid concentrations and their physiological responses. Such variability has been observed within studies quantifying salivary endocannabinoids so far.
Sexual dimorphism in the endocannabinoid system has been well-documented, 98 and numerous studies have reported higher levels of salivary AEA in men compared with women.43,62,87 Although the mechanisms underlying these differences remain unclear, higher FAAH expression has been observed in the SMG of female mice, 68 while circulating dehydroepiandrosterone sulfate has shown positive correlations with both AEA and 2-AG in human saliva. 99 Together, these findings suggest that sex-specific differences in salivary endocannabinoid tone may be influenced by both enzymatic activity and steroid hormone levels. However, not all studies have reported sex differences, with Tarragon and colleagues observing no sex-related effects 91 and Haviv et al. reporting differences in 2-AG only within specific neuropathic subgroups. 49
Age-related differences in salivary endocannabinoids have also been reported, although findings remain inconsistent. One study reported significantly higher 2-AG, but not AEA, in older age groups, 62 whereas another observed higher AEA, but not 2-AG, with increasing age. 87 Such differences may reflect age-related physiological changes that influence endocannabinoid metabolism.98,100,101
Collectively, these findings indicate that larger, well-characterized studies are required to clarify the influence of sex and age on salivary endocannabinoid concentrations. Importantly, interindividual differences may only become apparent within specific subgroups, contributing to variability across studies. Accounting for these factors will be essential for improving interpretability and reducing unexplained variability in future research.
Conclusions and Future Directions
Due to the invasive nature of tissue sampling, examining the endocannabinoid system in humans often requires the quantification of endocannabinoid ligands in biological matrices. Advancements in mass spectrometric techniques have enabled the quantification of endocannabinoids in saliva, a highly attractive biomatrix for diagnostic and research purposes.
Current evidence indicates that concentrations of salivary AEA and 2-AG are largely independent of circulating levels, providing novel, tissue-specific insights into endocannabinoid system activity under different physiological states. In contrast, more traditional salivary biomarkers, such as cortisol, estradiol, progesterone, and testosterone, are not produced locally but instead reflect unbound concentrations from the circulation.102–104 Consequently, their salivary levels can be influenced by local metabolism, complicating their interpretation as direct surrogates of physiological activity.66,102 Despite this feature, the potential utility of salivary endocannabinoids in research and clinical settings remains largely unexplored and remains to be determined.
Studies to date have shown that salivary endocannabinoids respond acutely to stress and exercise and display distinct profiles in individuals with conditions such as orofacial pain and obesity. These findings, if validated, suggest that salivary endocannabinoids could serve as real-time indicators of endocannabinoid system activity during various physiological processes and can help reveal disruptions associated with pathological conditions. Supporting this potential, salivary endocannabinoids have also been found to moderate instructive memory formation in cortisol nonresponders and correlate with subjective stress responses. Their responses may therefore not only offer unique insights to peripheral processes but also CNS functioning. Understanding how salivary endocannabinoids reflect CNS activity may offer a unique opportunity to identify neurobiological disruptions associated with psychological disorders or provide a profile between different disorders for diagnostic use. This is particularly valuable given that current biomarkers, such as cortisol and inflammatory markers, often lack disorder specificity and show inconsistent patterns across conditions.105,106
In addition to endocannabinoid ligands, salivary microRNAs implicated in the regulation of endocannabinoid signaling, 107 as well as CB1 receptor gene methylation patterns, 108 have been measured from saliva samples. However, as with endocannabinoids themselves, the degree to which these nucleic acid-based markers reflect local endocannabinoid system activity rather than that of other tissues remains uncertain. Nevertheless, it may be beneficial to investigate the significance of these markers in future, as both microRNAs and DNA can cross the blood–brain barrier and therefore could also give insight into central endocannabinoid activity.
Currently, the significance of salivary endocannabinoid research is limited by the small number of studies investigating this subject. Salivary cortisol is an extensively studied biomarker, supported by a robust body of research that includes analysis protocols, mechanistic investigations, and longitudinal studies. 109 This evidence base has enabled a detailed understanding of its reliability, biological relevance, and temporal dynamics as a biomarker of HPA axis activity. On the contrary, the mechanisms behind AEA or 2-AG regulation in the oral cavity are not fully understood. Their rapid responses to stress and exercise suggest that acute AEA and 2-AG concentrations within tissues of the oral cavity are regulated by the SNS; however, this has not been directly tested using mechanistic research in human tissue. This may have been especially difficult to address due to the lack of viable human salivary gland cell lines; however, methods to bioengineer a salivary gland model have been developed, which would be of use for this purpose. 110 Clarifying the signaling mechanisms underlying changes in salivary endocannabinoid concentrations will be crucial for interpreting future results and determining their relevance as biomarkers.
In addition to underlying physiological mechanisms, the influence of biological factors (e.g., sex, age, ethnicity, and allele variations) and lifestyle variables (e.g., smoking, diet, alcohol use, and sleep) have largely been explored for established salivary biomarkers.111,112 How these factors affect salivary endocannabinoid concentrations has not been established; however, as the endocannabinoid system is tightly integrated with other neuronal and hormonal systems, it is likely that there is variability in its ligand concentrations across different populations. So far, numerous studies have reported sex and age differences in salivary endocannabinoid levels, while other have not observed these differences. Larger and more comprehensive investigations are therefore needed to generate robust insights and to identify potential subgroup-specific patterns. Accounting for these biological and lifestyle factors is also critical in future studies, as uncontrolled variability can obscure true effects in endocannabinoid-related outcomes and limit the interpretability of results. Moreover, clarifying how these factors modulate endocannabinoid signaling could enhance our understanding of the mechanisms behind population-specific vulnerabilities to disease and could offer valuable insights to support the development of targeted therapeutic approaches.
Temporal dynamics, controlled by the circadian rhythm, represent another key source of endocannabinoid fluctuation. Previous research has demonstrated robust but distinct daily oscillations in circulating levels of AEA and 2-AG,113,114 raising the question of whether similar patterns occur in saliva. Recognizing and characterizing circadian influences is vital to prevent confounding variability in analysis and will also provide a window into disruptions in temporal regulation of the system in disease.
Finally, technical variability in endocannabinoid quantification represents a separate challenge for salivary endocannabinoid research. Differences in analytical methods used to process biological samples can significantly impact the reliability and comparability of findings across studies. 45 As such, developing and implementing standardized protocols for analyzing salivary endocannabinoids is essential to improve consistency and support broader integration of results across the field.
Overall, salivary endocannabinoids have significant potential as markers of endocannabinoid system function and its role in health and disease, for both research and diagnostic applications. However, continued investigation is essential to unlock their capabilities and to ensure robust, reproducible findings across diverse situations and populations.
Authors’ Contributions
J.H. was responsible for the literature search and writing the original draft. L.J.N. was responsible for conceptualization, supervision, and critical review and editing of the article. O.V.L. contributed to review and editing.
Footnotes
Author Disclosure Statement
The authors have no conflicts of interest to declare.
Funding Information
This research received no external funding.