
Abstract
Purpose:
The aim of the present investigation was to observe the effects of different energy densities of ultraviolet A photobiomodulation (UVA PBM) on human embryonic stem cell-derived mesenchymal stem cells (hESC-MSCs).
Methods:
The hESC-MSCs were cultured and irradiated with UVA PBM at energy densities ranging from 0.5 to 5.0 J/cm2 at a wavelength of 355 nm. Key biological parameters, including proliferation, viability, and migration, were evaluated at 72 h postirradiation.
Results:
Compared with the control, the UVA PBM significantly increased cell proliferation and viability at energy densities ranging from 0.5 to 2.0 J/cm2 and 0.5 to 3.0 J/cm2, respectively. Further, UVA PBM notably increased the migratory capacity of hESC-MSCs within the 0.5–2.0 J/cm2 range. Interestingly, higher energy densities beyond this threshold were found to be ineffective in promoting these cellular responses. In addition to examining proliferation, viability, and migration, this study explored the role of UVA PBM in osteogenic differentiation. Following irradiation, hESC-MSCs were induced toward osteogenesis, and differentiation was assessed through Alizarin Red S staining to evaluate calcium deposition. The results indicated that UVA PBM at optimal low-energy densities not only enhanced early proliferation but also supported osteogenic potential, as evidenced by increased mineralization in treated cells. To further understand the molecular mechanisms underlying these effects, quantitative PCR (qPCR) was performed to analyze the expression of key osteogenic markers, including RUNX2, Alkaline phosphatase, and Osteocalcin (OCN). Gene expression analysis revealed significant upregulation of these markers in the UVA PBM-treated groups, suggesting that UVA PBM promotes osteogenesis at the transcriptional level. UVA PBM at low energy densities significantly enhances the proliferation, viability, migration, and osteogenic differentiation of hESC-MSCs, potentially through the modulation of osteogenic gene expression.
Conclusion:
These findings highlight the therapeutic potential of UVA PBM in regenerative medicine and bone tissue engineering, particularly dentistry.
Keywords
Introduction
Currently, photobiomodulation (PBM) or low-level laser therapy has become a key tool in cell culture and has been used to encourage the proliferation, migration, and differentiation of many cell types, such as fibroblasts, epithelial cells,1–4 and osteoblasts, with calcification-related genes, such as osteoglycin. 5 Many studies are available regarding the effects of different visible and infrared lasers on stem cells, but only a few publications are available regarding the use of ultraviolet (UV) lasers on stem cells. UV light is mostly composed of ultraviolet A (UVA) and ultraviolet B (UVB). UVA (wavelengths ranging from 320 to 380 nm) is considered harmful or beneficial to cells and tissues.6–8 UVA irradiation is responsible for many alterations in cell biology. UVA light accounts for 90% of UV irradiation from solar light, which may significantly contribute to human life on Earth. UVB (wavelengths ranging from 275 to 320 nm) irradiation induces useful alterations in keratinocytes by causing intramembranous changes. 9 In addition, UVB irradiation at low energy levels increased the motility of melanoma cells via autocrine interleukin-8 secretion. 10
Mesenchymal stem cells (MSCs) are multipotent stromal cells capable of differentiating into various cell lineages, including osteoblasts, chondrocytes, and adipocytes, making them highly valuable in regenerative medicine and tissue engineering. 11 The modulation of MSC behavior, including proliferation, migration, and differentiation, is influenced by various physical and biochemical factors, among which UV light has gained attention for its potential biological effects.
Recent studies have highlighted the role of UV light in influencing the osteogenic differentiation of MSCs. For example, UVB irradiation has been shown to increase the expression of osteogenic markers such as RUNX2, Alkaline phosphatase (ALP), and Osteocalcin (OCN), potentially through the activation of reactive oxygen species (ROS)-mediated signaling pathways. 12 Moreover, UV exposure can stimulate the production of vitamin D, which plays a critical role in calcium metabolism and bone formation. 13 In addition, controlled UV irradiation has been found to regulate the Wnt/β-catenin signaling pathway, a key regulator of osteoblast differentiation and bone homeostasis. 14
In the context of human embryonic stem cell-derived mesenchymal stem cells (hESC-MSCs), the effects of ultraviolet A photobiomodulation (UVA PBM) on osteogenic differentiation remain underexplored. In this study, different energy densities of UVA PBM were used to irradiate hESC-MSCs to increase their proliferation and differentiation. Therefore, the aim of this investigation was to observe the effects of different energy densities of UVA PBM exposure on the biological responses of hESC-MSCs, with a focus on proliferation, viability, migration, and osteogenic potential. In addition, this study evaluated the expression of key osteogenic genes via quantitative PCR (qPCR) to elucidate the molecular mechanisms underlying UVA PBM-induced osteogenesis.
Materials and Methods
Culture of human embryonic stem cell-derived mesenchymal stem cells
For this investigation, hESC-MSCs were obtained from the Stem Cell Unit, College of Medicine, King Saud University, Riyadh. 15 Complete medium (Dulbecco’s modified Eagle’s medium [DMEM]/high glucose, 10% fetal calf serum, 1% essential amino acids, and 1% Anti-anti) was used to culture hESC-MSCs. hESC-MSCs were incubated in a humidified chamber containing 5% CO2 at 37°C. hESC-MSCs were harvested via trypsin and ethylenediamine tetraacetic acid solution (Gibco), Thermo Fisher Scientific, Waltham, MA, USA and incubated at 37°C for 3–5 min. Then, complete medium was added to collect the cells, which were subsequently centrifuged at 500g for 5 min to collect the cell pellet, which was subsequently transferred to 48-well plates. hESC-MSCs were incubated overnight before UVA PBM irradiation.
Ultraviolet A photobiomodulation
UVA PBM at 355 nm was used to irradiate hESC-MSCs with different energy densities, such as 0.5, 1.0, 1.5, 2.0, 2.5, 3.0, 3.5, 4.0, 4.5, and 5.0 J/cm2, as explained previously.16,17 A power density of 25 mW/cm2 in continuous wave mode was used for irradiation. The irradiation duration was 20-200 sec, along with that of the control group (nonirradiated). The control group of hESC-MSCs was not irradiated and was maintained under the same conditions as the other treatment groups. The irradiation process was performed only once. The medium of the hESC-MSCs was replaced with sterile phosphate-buffered saline (PBS) before irradiation to minimize the loss of laser energy. Sterile conditions for hESC-MSCs were maintained during laser irradiation. Fresh complete medium was added to each well after laser treatment, and the hESC-MSCs were incubated further. All experiments were performed in triplicate for statistical analysis during this study.
Cell proliferation determination
The proliferation of hESC-MSCs was determined by 3-[4,5-dimethylthiazol-2-yl]−2,5-diphenyltetrazolium bromide (MTT) assay, a highly water-soluble formazan dye, after UVA PBM irradiation. hESC-MSCs (4 × 104 cells per well) were seeded into 48-well plates and incubated at 37°C overnight. The next day, hESC-MSCs were irradiated with UVA PBM as described above and then further incubated for 72 h. At the end of the experiment, 100 µL of MTT solution (0.5 mg/ml in PBS) was added to each well, and the plate was incubated at 37°C for 2 h. After that, the MTT solution was replaced with an equal volume of isopropanol to dissolve the formazan crystals, and the plate was incubated on a shaker for 2 h. The absorbance was measured at 549 nm with a microplate reader (BioTek, USA).18,19
Cell viability rate
Trypan blue staining is useful for counting viable and nonviable cells, as explained in our previous publications. 17 Briefly, 4 × 104 cells were seeded in each well of a 48-well plate and incubated overnight. Then, hESC-MSCs were irradiated with UVA PBM irradiation as described above. At the end of the experiment, after 72 h, hESC-MSCs were harvested via trypsinization (0.25% trypsin–0.01% ethylenediamine tetraacetic acid (EDTA) suspension; Gibco). An equal volume of cells was stained with an equal volume of trypan blue, and the resulting mixture was incubated at room temperature for 2 min. Then, viable (unstained) and nonviable (stained) cells were counted via a Neubauer chamber under a phase contrast microscope (Olympus IX51, Tokyo, Japan). The number of viable cells in the UVA PBM-treated group was compared with that in the control group (nonirradiated), and graphs of cell viability were generated.
Cell migration assay
Scratch or wound healing assays were used to determine the number of migrated hESC-MSCs treated with UVA PBM irradiation. We used our previously published protocol to perform this assay. 17 Briefly, 4 × 104 cells were seeded in each well of a 48-well plate and incubated overnight to allow the cells to settle. The next day, a “scratch or wound” was created in the center of each well via a sterile 200 µL pipette tip, after which the hESC-MSCs were irradiated with UVA PBM as described above. The PBS was removed, and fresh medium was added to each well after UVA PBM irradiation. Then, the hESC-MSCs were incubated further. At the end of the experiment at 72 h, photographs were taken during migration from each group via a camera attached to a phase-contrast microscope (Olympus) at a power of 10×. For statistical analysis, all photographs were taken from three different “scratch or wound areas” at the same magnification. Then, the number of migrated hESC-MSCs in the “scratch area” from the control and irradiated groups was determined, and graphs were plotted for the cell migration assay.
Osteoblast differentiation of hMSCs via an optimized induction protocol
For this study, human mesenchymal stem cells (hMSCs) derived from hESCs were used as a model for primary MSCs, which exhibit similar cellular and molecular phenotypes. These cells were cultured in DMEM supplemented with 0.25 mol/L
The cells were grown to 70–80% confluence at a density of 50,000 cells/mL. For osteoblast induction, the medium was replaced with osteogenic differentiation medium containing 10% FBS, penicillin–streptomycin, 0.003 mol/L ascorbic acid, 0.01 mol/L β-glycerophosphate, 10−8 mol/L calcitriol (vitamin D3), and 10−8 mol/L dexamethasone. Differentiation was conducted in 48-well plates, with the medium changed three times per week, and the plates were incubated for a total of 14 days for PCR and quantitative ALP activity assay and 21 days for ALP staining. 15
Quantitative ALP activity
To quantify ALP activity after osteogenic differentiation, we used a BioVision ALP activity colorimetric assay kit (BioVision, Inc., CA, USA) with some modifications. The cells were cultured in 48-well plates under the same conditions; then, on the day of analysis, the wells were rinsed once with PBS, and 50 µL of pNPP solution was added to each well and incubated for 2 min in the dark at room temperature. The Optical density (OD) was then measured at 405 nm.
Flow cytometry
Flow cytometry was performed on UVA PBM-treated and control single-cell suspensions. The cells were harvested with trypsin, neutralized in 10% serum, and resuspended in Fluorescence-Activated Cell Sorting (FACS) buffer (PBS with 1% FBS) at a concentration of 1 × 106 cells/mL. Approximately 1 × 105 cells were used per antibody stain. The MSC-specific antibodies used included CD14-APC, CD29-PE, CD31-FITC, CD34-FITC, CD44-PE, CD73-PE, and CD146-PE, with matched isotype controls (IgG1-FITC, IgG1-PE, and IgG-APC) from BD Biosciences.
The cells were stained with antibodies for 30 min on ice, washed twice with FACS buffer, and resuspended for acquisition. A minimum of 10,000 events per sample were acquired via a FACSCalibur (BD Biosciences) and analyzed with CellQuest Pro® software Kansas, USA.
Quantitative real-time PCR
Total RNA was extracted via an innuPREP RNA Mini Kit (Analytik Jena, Germany, REF No: 845-KS-2040250) as recommended by the manufacturer. Total RNA was quantified via a Nanodrop spectrophotometer (Nanodrop 2000, Thermo Fisher Scientific, USA). Complementary DNA (cDNA) was synthesized from 1 μg of RNA with a high-capacity cDNA reverse transcription kit (Applied Biosystems, USA) via a ProFlex PCR system with a multigene thermal cycler according to the manufacturer’s instructions. Relative levels of mRNA were determined from the cDNA via RT-PCR (Applied Biosystems Real-Time PCR Detection System) via a Fast SYBR Green PCR Kit (Applied Biosystems, UK) according to the manufacturer’s instructions. Following normalization to the reference gene, Glyceraldehyde 3-Phosphate Dehydrogenase (GAPDH), the quantification of gene expression was carried out via a comparative computed tomography (CT) method, where ΔCT is the difference between the CT values of the target and reference genes.
Statistical analysis
One-way analysis of variance (ANOVA) was used to compare the mean values of the quantitative variables across the categorical variables, followed by the post hoc Tukey HSD test. A p value of <0.05 was considered statistically significant. The data were analyzed via GraphPad Prism 4.0 (GraphPad Software, San Diego, CA, USA). Student’s t-test was performed for PCR data, and graphs were prepared using Microsoft Excel.
Results
Effects of ultraviolet A photobiomodulation on the proliferation of hESC-MSCs
The proliferation of hMSC-MSCs was assessed via the MTT assay, which is based on mitochondrial activity. This assay measures the optical density in wells, which indicates living cells present in the UVA PBM nonirradiated and irradiated groups. UVA PBM irradiation significantly increased the proliferation of hMSC-MSCs treated with different energy densities. Figure 1A shows that the proliferation of hMSC-MSCs increased by 1.52, 1.58, 1.45, 1.41, 1.31, 1.17, 1.16, 1.1, 1.0, and 0.98-fold at doses of 0.5, 1.0, 1.5, 2.0, 2.5, 3.0, 3.5, 4.0, 4.5, and 5.0 J/cm2 energy density, respectively, compared with that of the control, with a maximal and significant effect obtained following 1.0 J/cm2 (p < 0.001). In addition, UVA PBM increased proliferation from 2.5 to 4.0 J/cm2, but this increase was not significant. However, UVA PBM did not increase the proliferation of hMSC-MSCs when hMSC-MSCs were treated with energy densities ranging from 4.5 to 5.0 J/cm2.
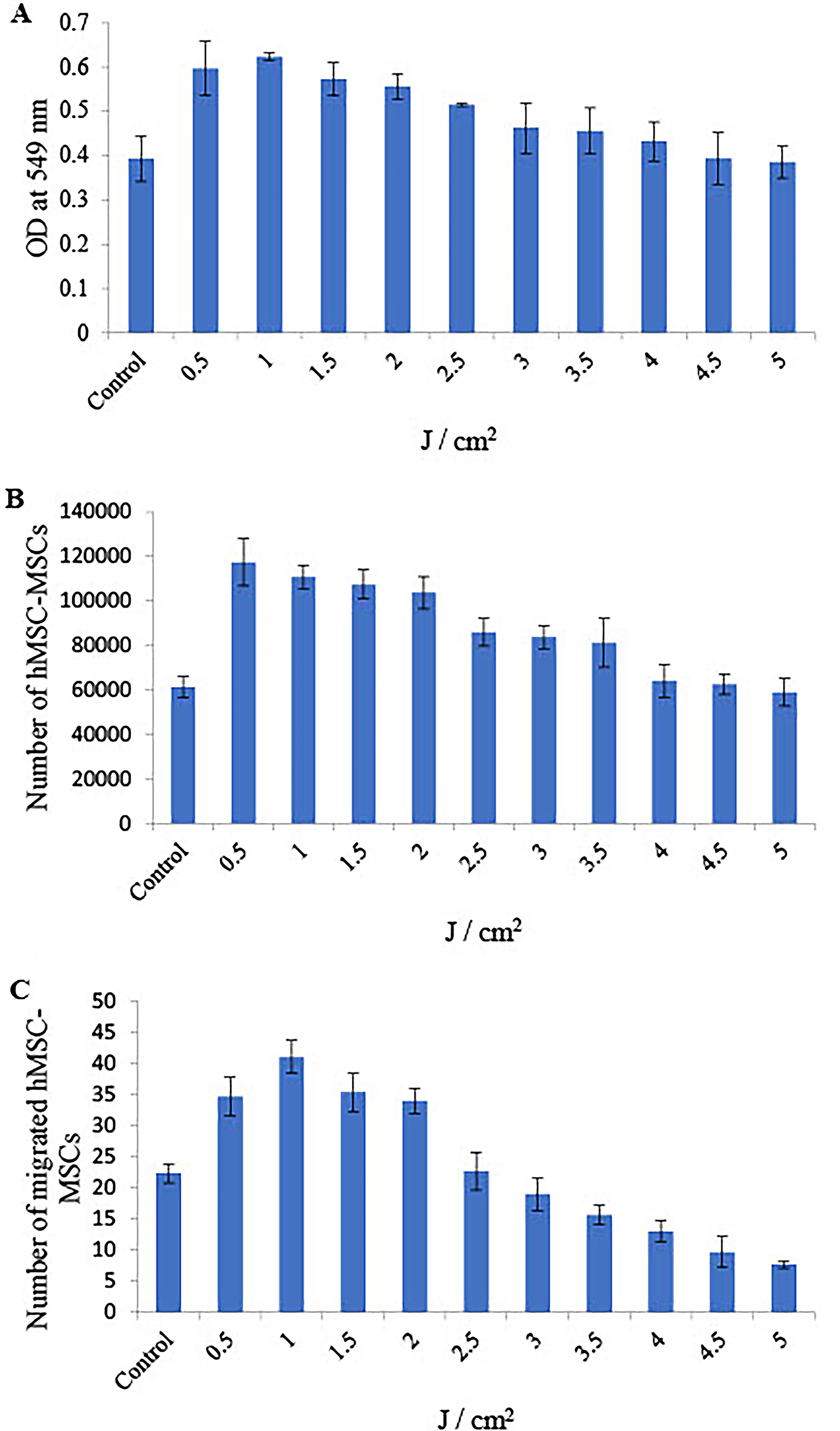
One-way ANOVA revealed that the energy density of UVA PBM from 0.5 to 2.0 J/cm2 was significantly different compared with control, and the lowest p value was <0.01 (Table 1).
Significance Values Obtained from One-Way ANOVA of the Means of Three Cell Culture Experiments Examining the Viability, Proliferation, and Migration of Human Embryonic Stem Cell-Derived Mesenchymal Stem Cells Treated with Different Energy Densities of Ultraviolet A Photobiomodulation
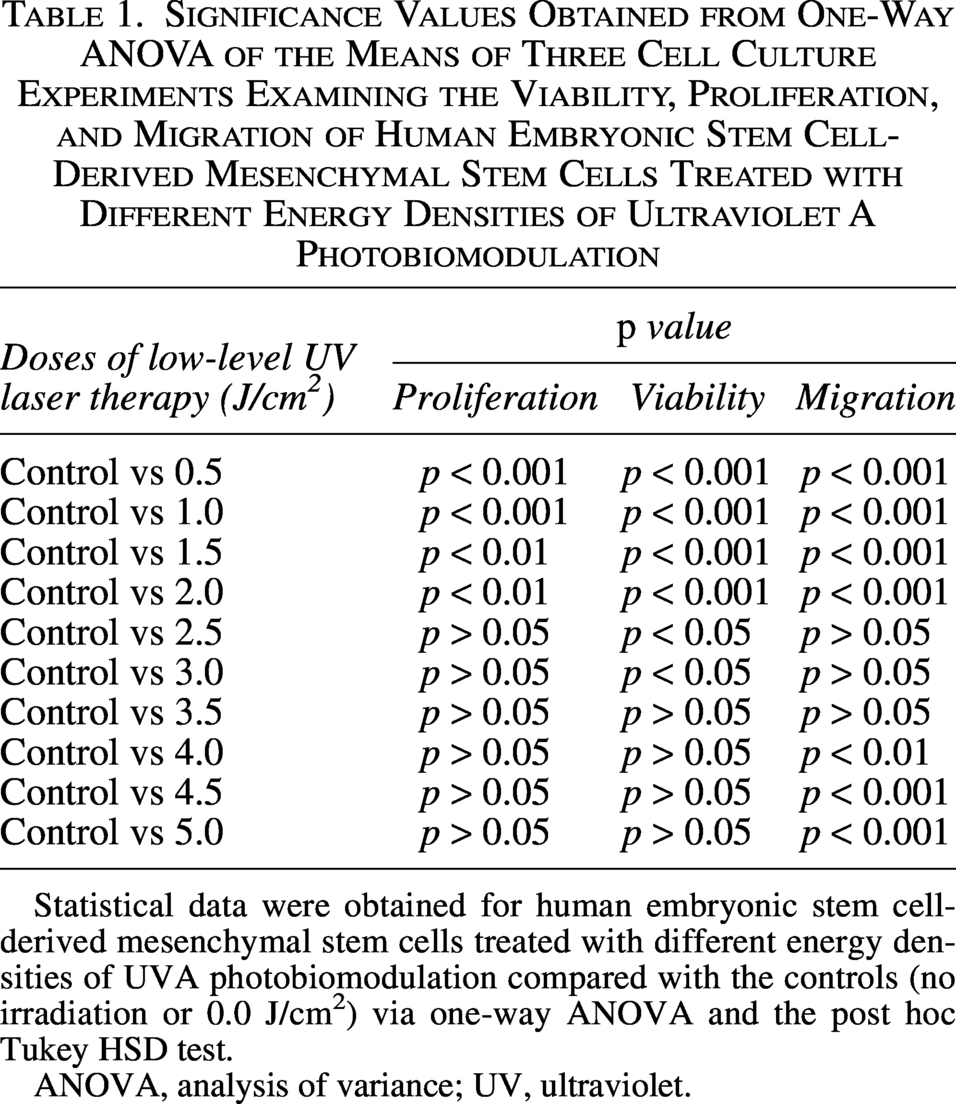
Statistical data were obtained for human embryonic stem cell-derived mesenchymal stem cells treated with different energy densities of UVA photobiomodulation compared with the controls (no irradiation or 0.0 J/cm2) via one-way ANOVA and the post hoc Tukey HSD test.
ANOVA, analysis of variance; UV, ultraviolet.
Effects of ultraviolet A photobiomodulation on the viability of hESC-MSCs
The experiments were performed in triplicate, and the average value was used to summarize the results obtained. UVA PBM irradiation was given to hMSC-MSCs once to observe the viability of hMSC-MSCs compared with that of nonirradiated hMSC-MSCs (control). In this experiment, the number of viable cells, stained by trypan blue dye, was determined at 72 h following irradiation, as illustrated in Figure 1B. UVA PBM significantly increased the viability of hMSC-MSCs by 1.92, 1.81, 1.75, 1.69, 1.4, 1.36, 1.32, 1.04, 1.02, and 0.96-fold at doses of 0.5, 1.0, 1.5, 2.0, 2.5, 3.0, 3.5, 4.0, 4.5, and 5.0 J/cm2, respectively, compared with the control, with a maximal and significant effect obtained following 0.5 J/cm2 (p < 0.001).
One-way ANOVA revealed that the energy density of UVA PBM from 0.5 to 3.0 J/cm2 was significantly different from that of the control, and the lowest p value was <0.05 (Table 1). In addition, UVA PBM was able to increase the number of viable hMSC-MSCs from 3.5 to 4.0 J/cm2 compared with the control, but this increase was not statistically significant.
Effects of ultraviolet A photobiomodulation on the migration of hMSC-MSCs
Compared with the control, UVA PBM significantly increased the number of migrated hMSC-MSCs from 0.5 to 2.0 J/cm2. As shown in Figure 1C, the number of migrated hMSC-MSCs treated with UVA PBM increased by 1.55-, 1.83-, 1.58-, 1.52-, 1.01-, 0.85-, 0.7-, 0.58-, 0.43-, and 0.34-fold at doses of 0.5, 1.0, 1.5, 2.0, 2.5, 3.0, 3.5, 4.0, 4.5, and 5.0 J/cm2 energy density, respectively, compared with that of the control, with a maximal and significant effect obtained following 1.0 J/cm2 (p < 0.001).
One-way ANOVA revealed that, compared with the control, UVA PBM significantly increased the number of migrated hMSC-MSCs from 0.5 to 2.0 J/cm2, and the p value was <0.001 (Table 1). On the other hand, UVA PBM at 2.5–5.0 J/cm2 failed to increase the migration of hMSC-MSCs compared with that of the controls.
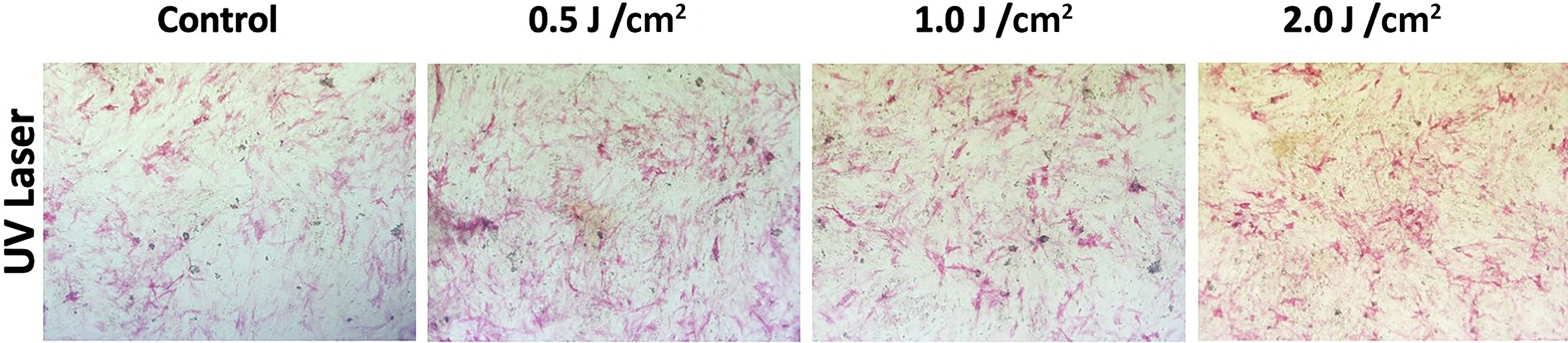
Photomicrographs of hMSC-MSCs treated with UVA PBM revealed that migration was most effective at an energy density of 1.0 J/cm2 compared with that of the control (Fig. 2).

Photographs of human embryonic stem cell-derived mesenchymal stem cells at 72 h after treatment with ultraviolet A photobiomodulation (UVA PBM) at energy densities of 0.0 J/cm2 or control
Ultraviolet A photobiomodulation enhanced the OB differentiation of hESC-MSCs
ALP activity was measured to observe the differentiation of hESC-MSCs from the UVA PBM treatment into osteoblasts. Three different energy densities of UVA PBM irradiation, 0.5, 1.0, and 2.0 J/cm2, were applied to hESC-MSCs, and control and osteoblast-induced medium were used to observe the differentiation of hESC-MSCs toward the osteoblast lineage. After UVA PBM irradiation, hESC-MSCs were incubated for a total of 21 days, after which differentiation was measured by ALP activity. These results revealed that UVA PBM increased differentiation under the two highest energy-density conditions.
Measurement of ALP activity via ultraviolet A photobiomodulation
These data showed that high-dose UVA PBM increased differentiation (Fig. 3). Our data revealed that UVA PBM had the biggest effect on the differentiation of osteoblasts at the two highest energy densities (Fig. 4); the effect was dose dependent. These findings indicate that UV lasers have a positive effect on differentiation.
Alkaline phosphatase (ALP) staining of human embryonic stem cell-derived mesenchymal stem cells (hESC-MSCs) stimulated with varying doses of ultraviolet A photobiomodulation (UVA; 0.5 J/cm², 1.0 J/cm², and 2.0 J/cm²) and subsequently differentiated into osteoblasts. The control group was not exposed to UVA PBM. Images were captured at 20x magnification, which revealed dose-dependent increases in ALP activity, as evidenced by the intensity of staining, indicative of osteoblast differentiation.
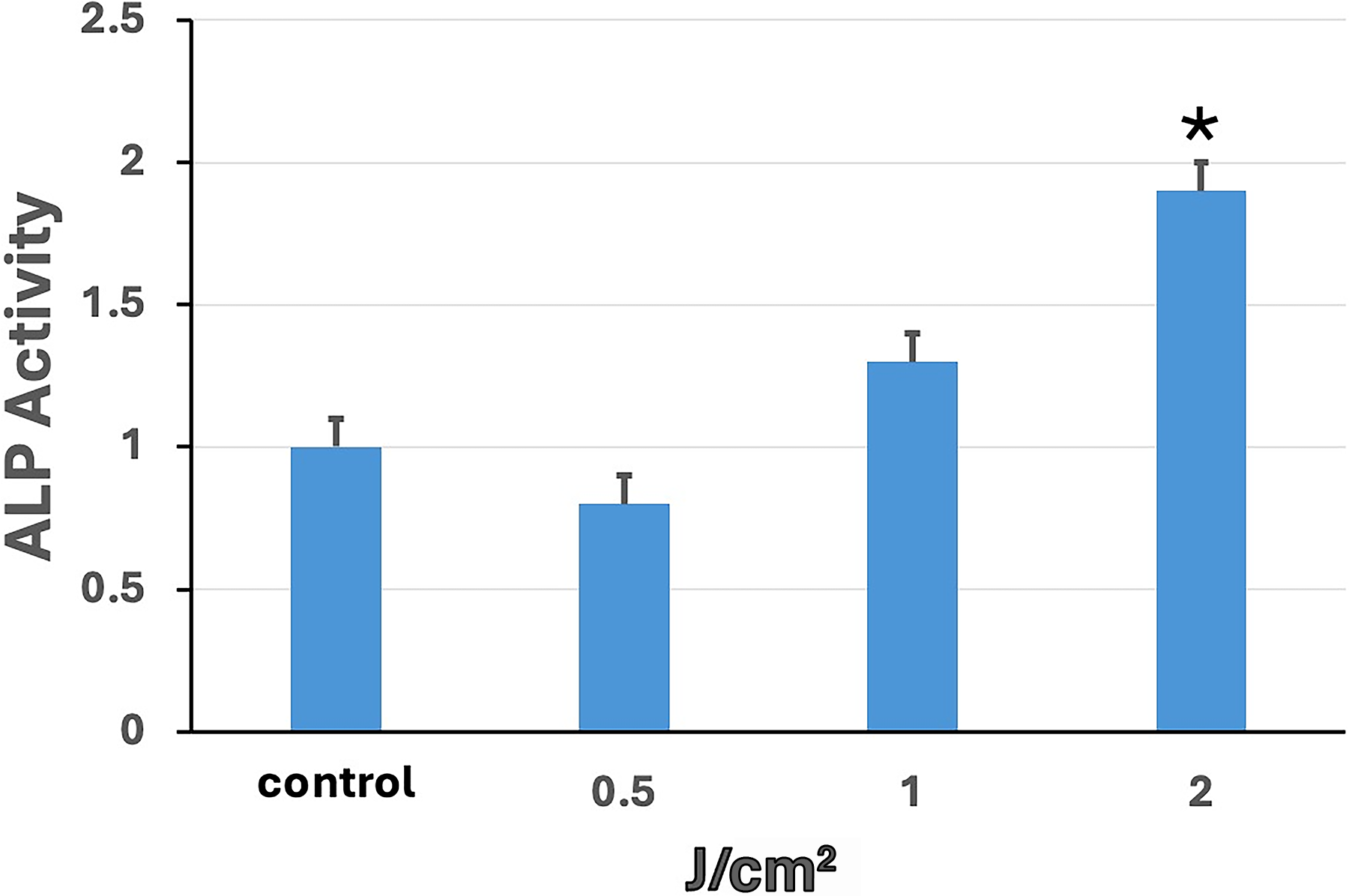
Alkaline phosphatase (ALP) activity of human embryonic stem cell-derived mesenchymal stem cells (hESC-MSCs) treated with ultraviolet A photobiomodulation (UVA PBM) at energy densities of 0.5, 1.0, and 2.0 J/cm² compared with that of the control group (Cnt). ALP activity was quantified, revealing a dose-dependent increase in activity with increasing UVA PBM energy density. Significance of differences between irradiated and controls: *p < 0.01.
Flow cytometry results analysis
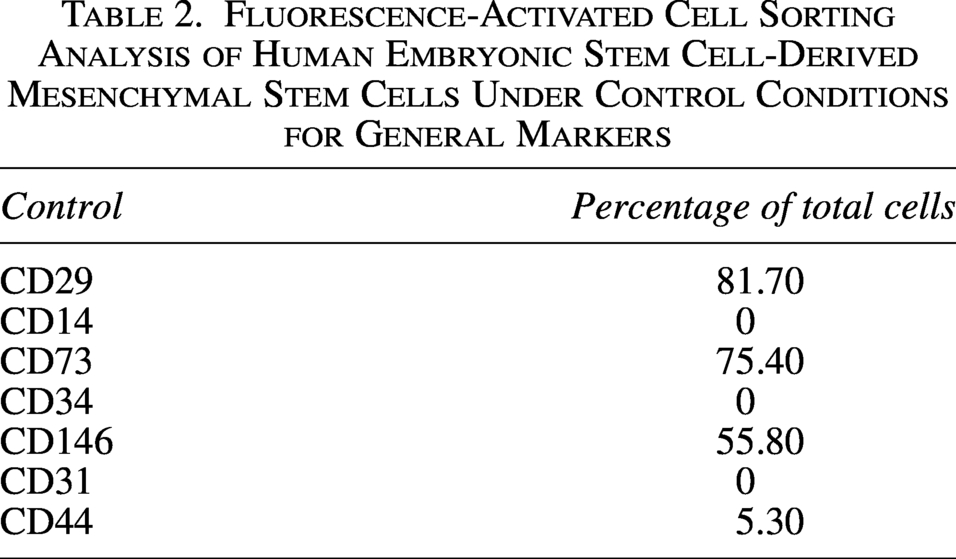
We performed FACS on human Bone Marrow Mesenchymal Stem Cells (BMSC) cells without any treatment and found that key MSC markers were present and highly expressed in our cells, whereas only CD44 was expressed at low levels (Table 2). We treated hESC-MSCs with UVA PBM at different energy densities, such as 0.5, 1.0, and 2.0 J/cm2. We discovered that 0.5 J/cm2 UVA PBM treatment increased the expression of all the key markers CD29, CD73, and CD146; however, the expression of CD44 slightly decreased. However, higher doses of UVA PBM reduced marker expression but maintained higher expression than the control conditions did, indicating that UVA PBM treatment is beneficial for hESC-MSCs (Table 3).
Fluorescence-Activated Cell Sorting Analysis of Human Embryonic Stem Cell-Derived Mesenchymal Stem Cells Under Control Conditions for General Markers
The hESC-Derived Mesenchymal Stem Cells Were Treated with Different Energy Densities of UVA Photobiomodulation at 0.5, 1.0, and 2.0 J/cm2 and Then Subjected to FACS Analysis
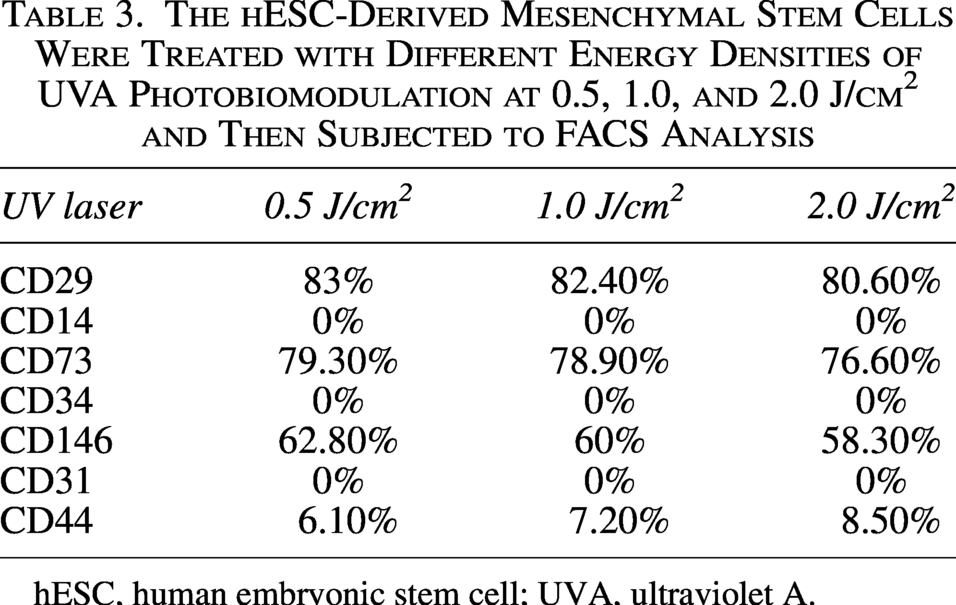
hESC, human embryonic stem cell; UVA, ultraviolet A.
Figure 5 shows human stem cells in the control condition without any treatment and with the ISO-type control. We observed that there was no background staining with any fluorescence. We observed that, when the cells were treated with UVA PBM at 0.5 and 1.0 J/cm2, the expression of CD29 and CD73 was highest and increased compared with the control and other treatments. These findings indicate that UVA PBM treatment promotes MSC marker expression (Fig. 6). We noticed that the UVA PBM had the greatest effect on the MSC expression of key cell surface markers such as CD29 and CD73; however, no treatment had any effect on the negative markers CD14 and CD31.
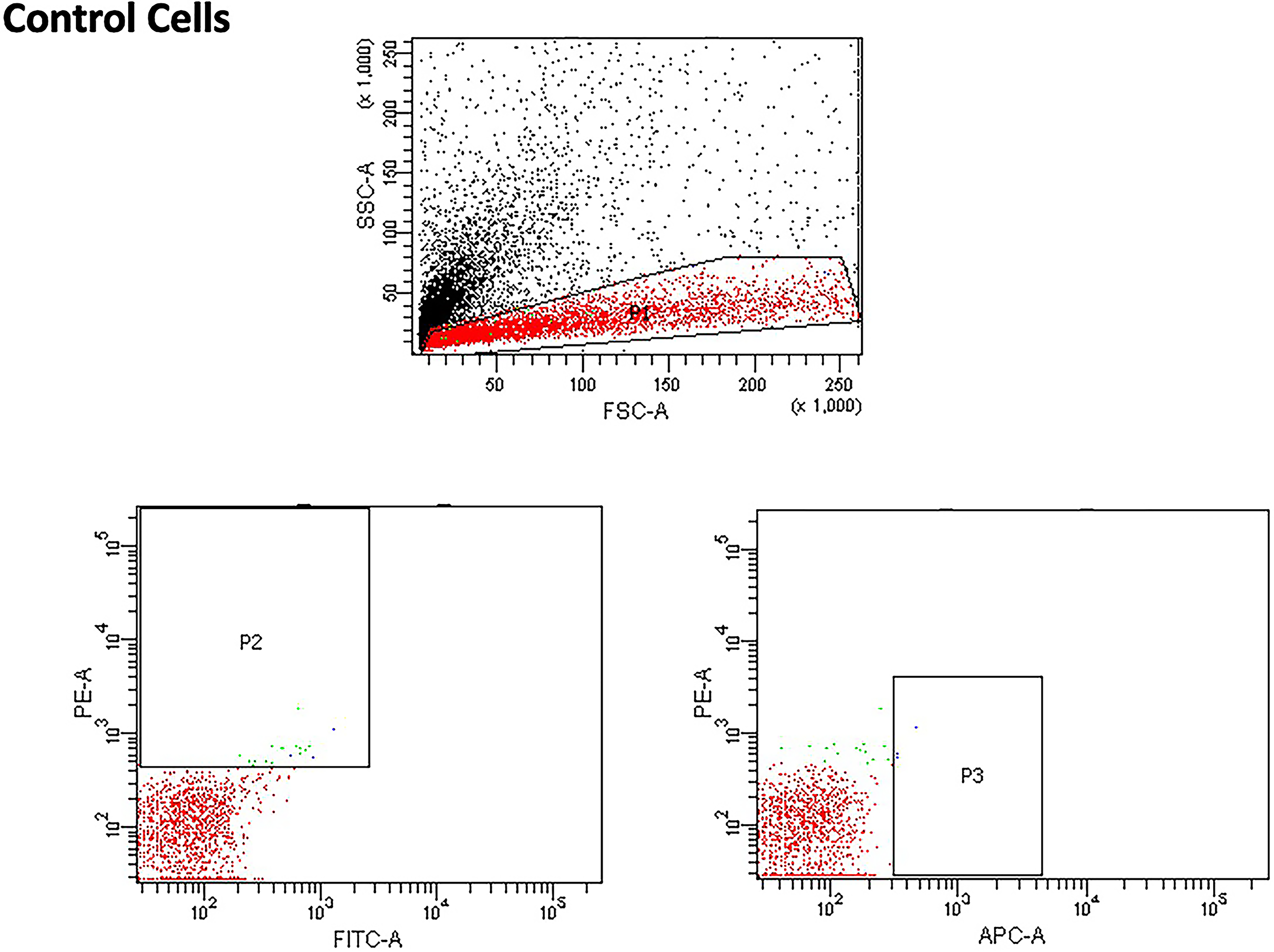
Forward scatter and side scatter and ISO-type controls, which were the same for the ultraviolet A photobiomodulation (UVA PBM) and all energy densities.
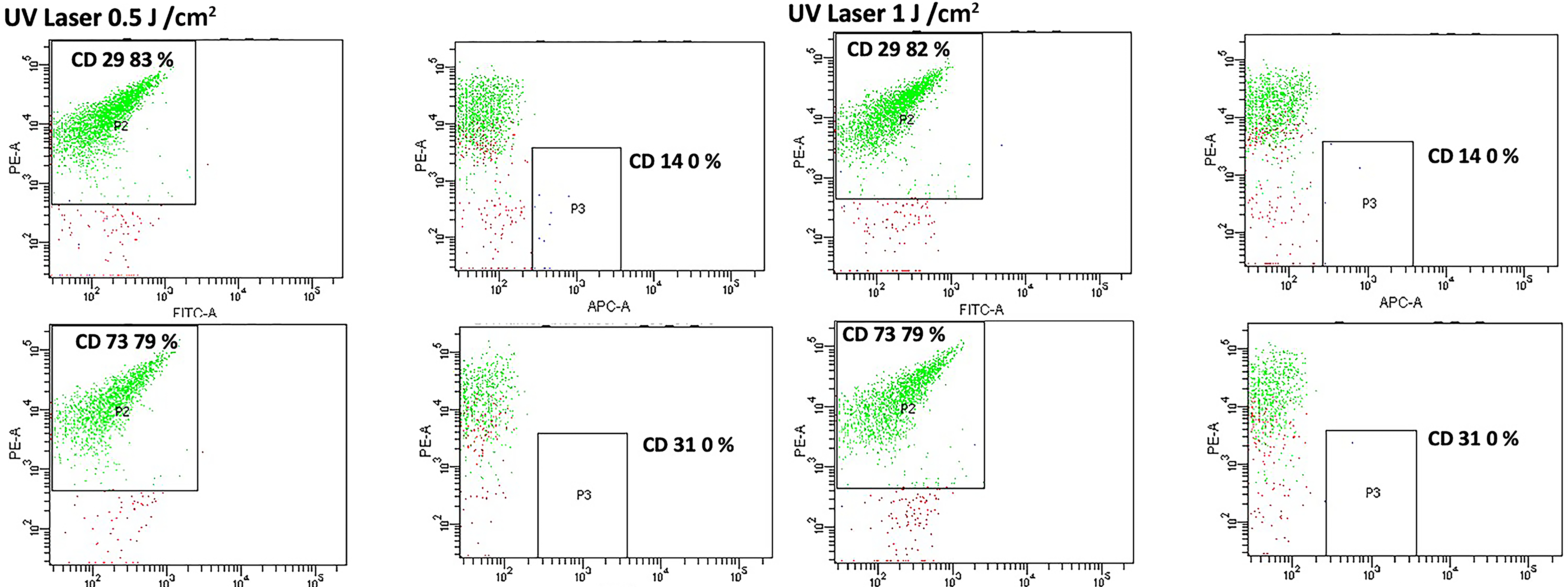
Flow cytometry (FACS) analysis of human embryonic stem cell-derived mesenchymal stem cells (hESC-MSCs) treated with ultraviolet A photobiomodulation (UVA PBM) at energy densities of 0.5 J/cm² and 1.0 J/cm² to assess the expression of the surface markers CD29, CD73, CD14, and CD31. UVA PBM treatment maintained the characteristic mesenchymal stem cell (MSC) marker profile (CD29 and CD73 positivity) and minimal expression of hematopoietic (CD14) and endothelial (CD31) markers, indicating preservation of the mesenchymal phenotype following treatment.
Gene expression analysis for key osteoblastic genes
ALP was the first gene to be investigated. We observed ALP gene expression with UVA PBM and its three different energy densities. UVA PBM presented the highest gene expression at all three energy densities (Fig. 7). The next gene we investigated was RUNX2, which is another key early osteoblast marker gene for osteoblast differentiation, and UVA PBM significantly upregulated RUNX2 up to threefold at 1.0 J/cm2 (Fig. 8). The next gene we investigated was BMP4, another key gene but a mid-differentiation gene that is expressed at the progenitor stage. As shown in Figure 9, UVA PBM did not increase BMP4 expression substantially at 0.5 J/cm2, and UVA PBM increased the expression of BMP4 at 1.0 and 2.0 J/cm2.
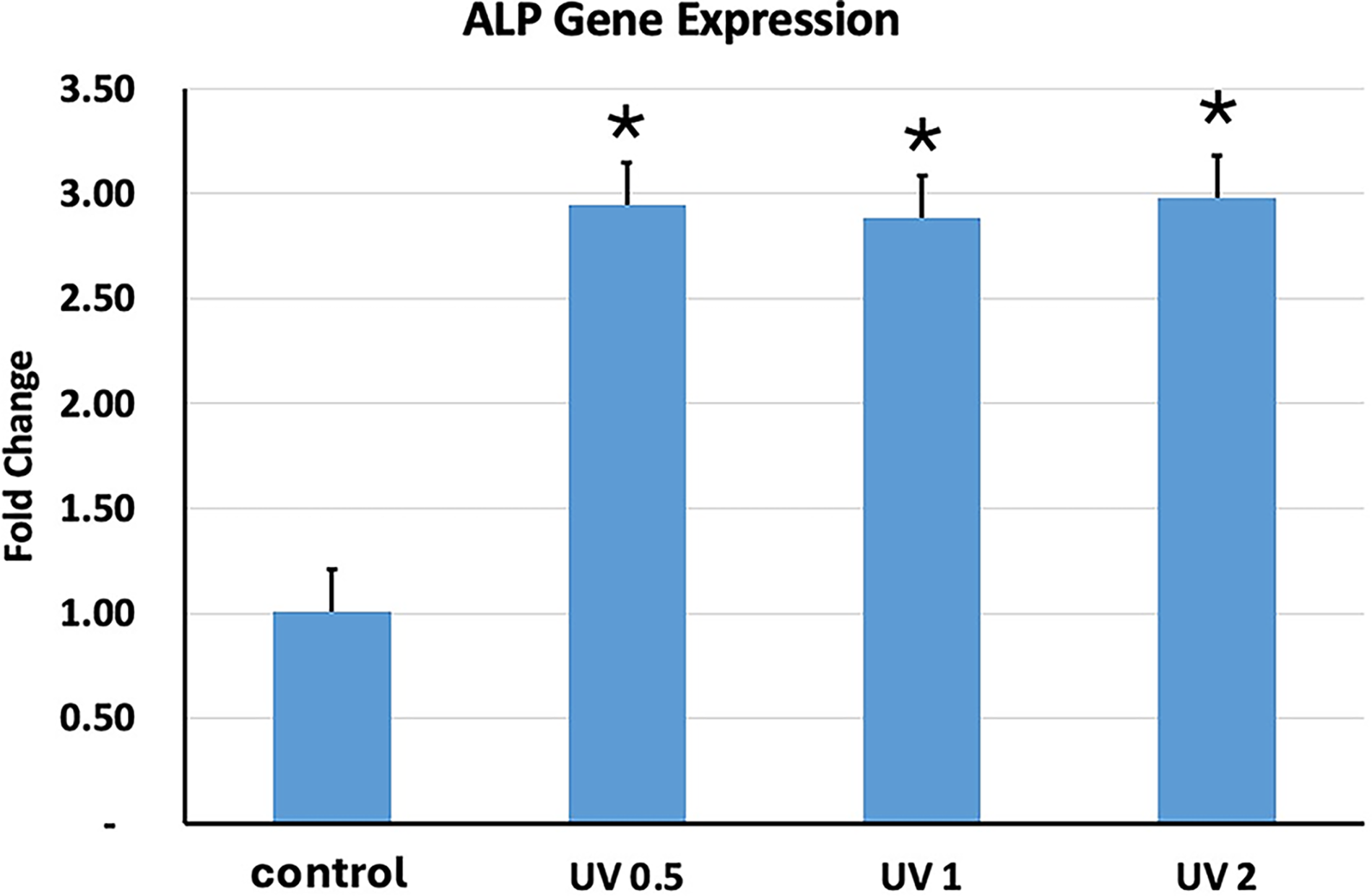
Quantitative PCR (qPCR) analysis of alkaline phosphatase (ALP) gene expression in human embryonic stem cell-derived mesenchymal stem cells (hESC-MSCs) treated with ultraviolet A photobiomodulation (UVA PBM) at energy densities of 0.5, 1.0, and 2.0 J/cm². The fold change in ALP expression was normalized to that of the untreated control (control), demonstrating a dose-dependent response to UVA PBM. The data indicate significant upregulation of ALP gene expression at 0.5 and 1.0 J/cm², with reduced expression observed at 2.0 J/cm². Significance of differences between irradiated and controls: *p < 0.01.
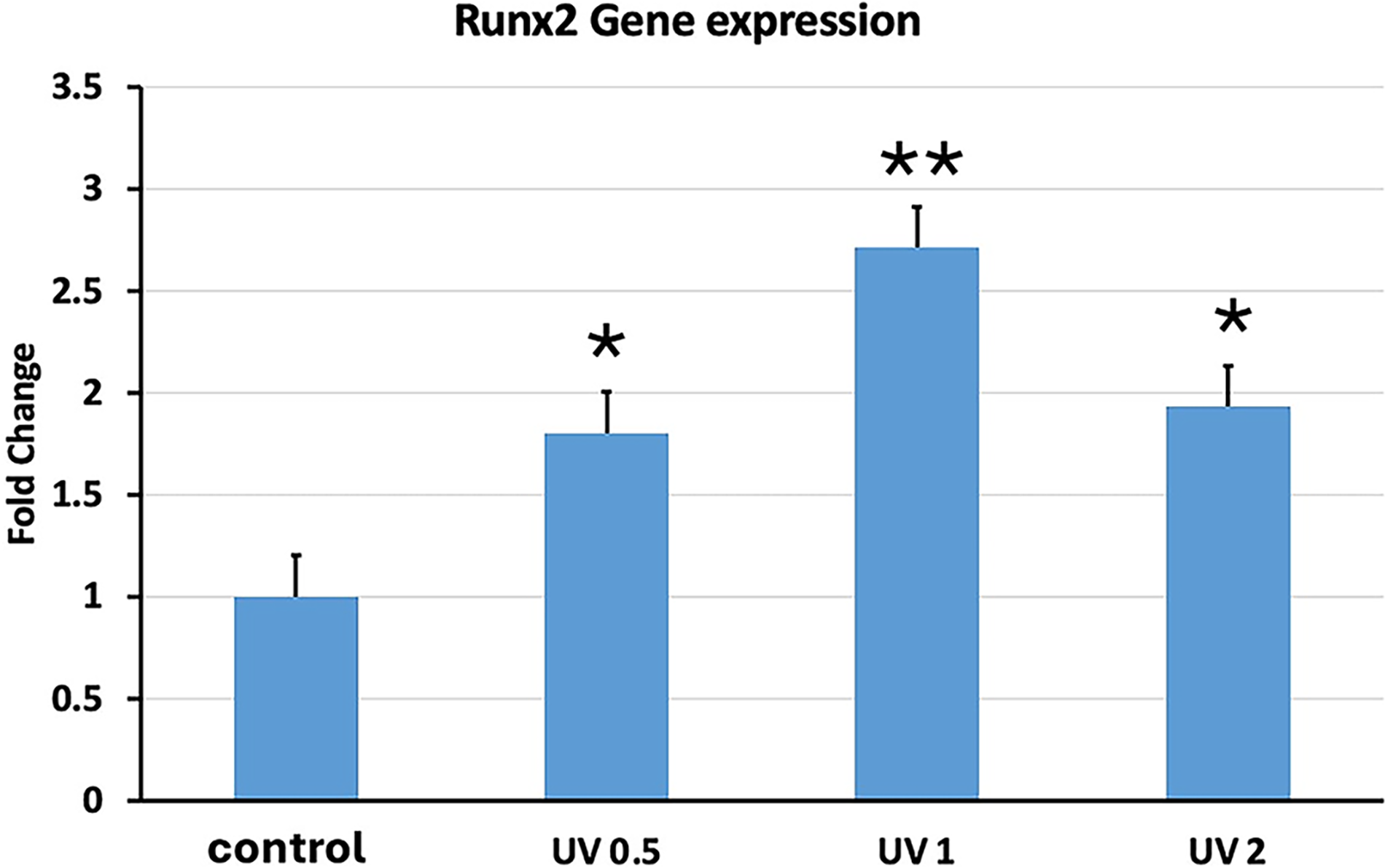
Quantitative PCR (qPCR) analysis of RUNX2 gene expression in human embryonic stem cell-derived mesenchymal stem cells (hESC-MSCs) treated with ultraviolet A photobiomodulation (UVA PBM) at energy densities of 0.5, 1.0, and 2.0 J/cm². The fold change in RUNX2 expression was normalized to that of the untreated control (control). The data revealed significant upregulation of RUNX2 expression at 0.5 and 1.0 J/cm², whereas a decrease in expression was observed at 2.0 J/cm², indicating a dose-dependent modulation of osteogenic gene activation by UVA PBM. Significance of differences between irradiated and controls: *p < 0.01; **p < 0.001.
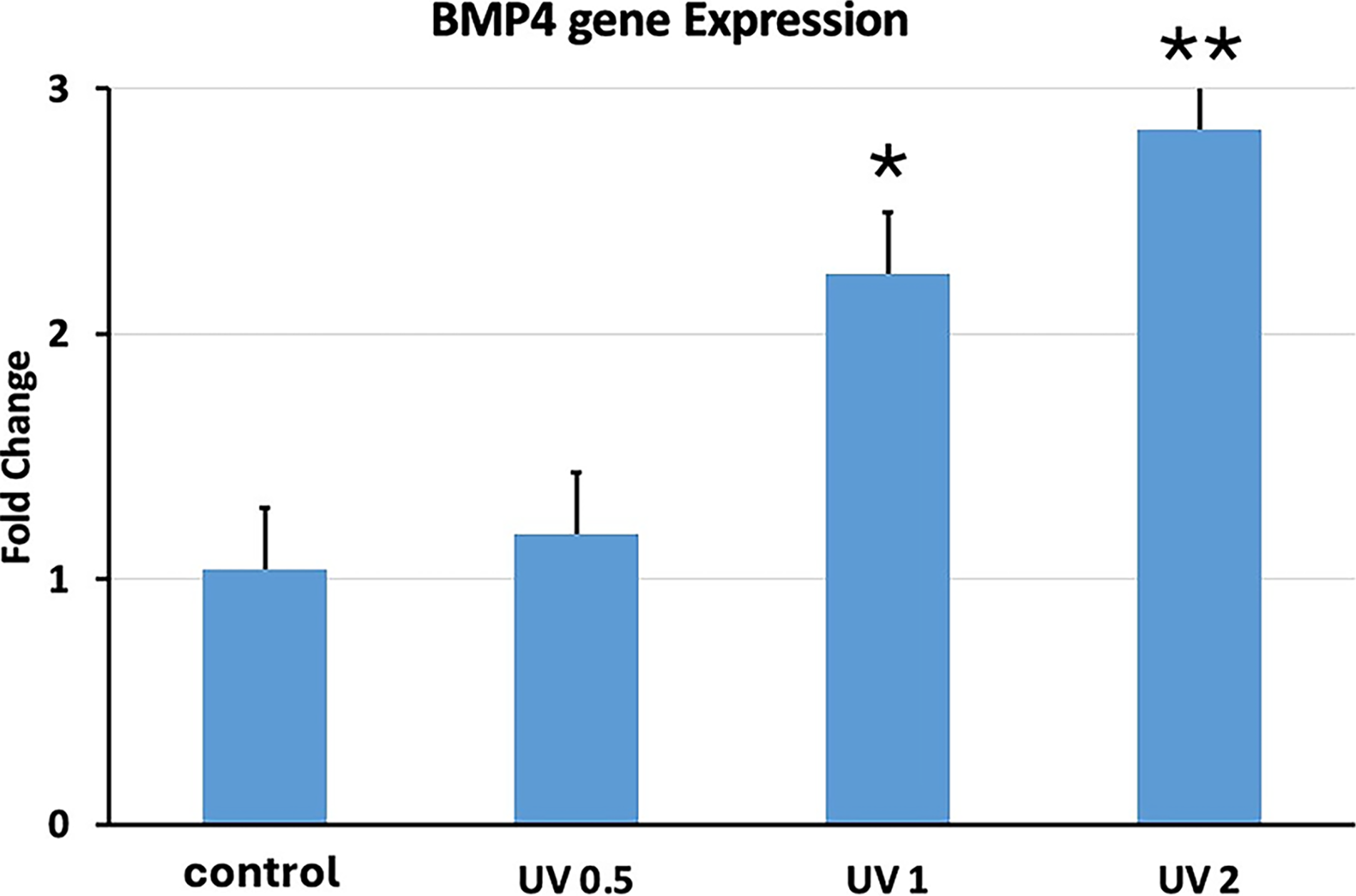
Quantitative PCR (qPCR) analysis of BMP4 gene expression in human embryonic stem cell-derived mesenchymal stem cells (hESC-MSCs) treated with ultraviolet A photobiomodulation (UVA PBM) at energy densities of 0.5, 1.0, and 2.0 J/cm². The fold change in BMP4 expression was normalized to that of the untreated control (control). The data indicate significant upregulation of BMP4 expression at all energy densities, with the highest expression observed at 2.0 J/cm², suggesting a dose-dependent enhancement of BMP4 activation by UVA PBM. Significance of differences between irradiated and controls: *p < 0.01; **p < 0.001.
Another gene we investigated was Col1a, which is also a mid- to late-differentiation gene found in all osteoblasts and bone. Figure 10 shows that UVA PBM at all energy densities significantly increased the expression of Col1a. UVA PBM increased the expression of Col1a more than 2.5-fold at 1.0 J/cm2. The final gene we investigated was osteonectin (ON), which is a late-stage marker that is found only in mature osteoblasts. Figure 11 shows that UVA PBM could increase the expression of ON up to 2.5-fold at 1.0 J/cm2. Our results clearly show that UVA PBM can significantly increase the expression of osteoblastic genes and increase the differentiation of hESC-MSCs.
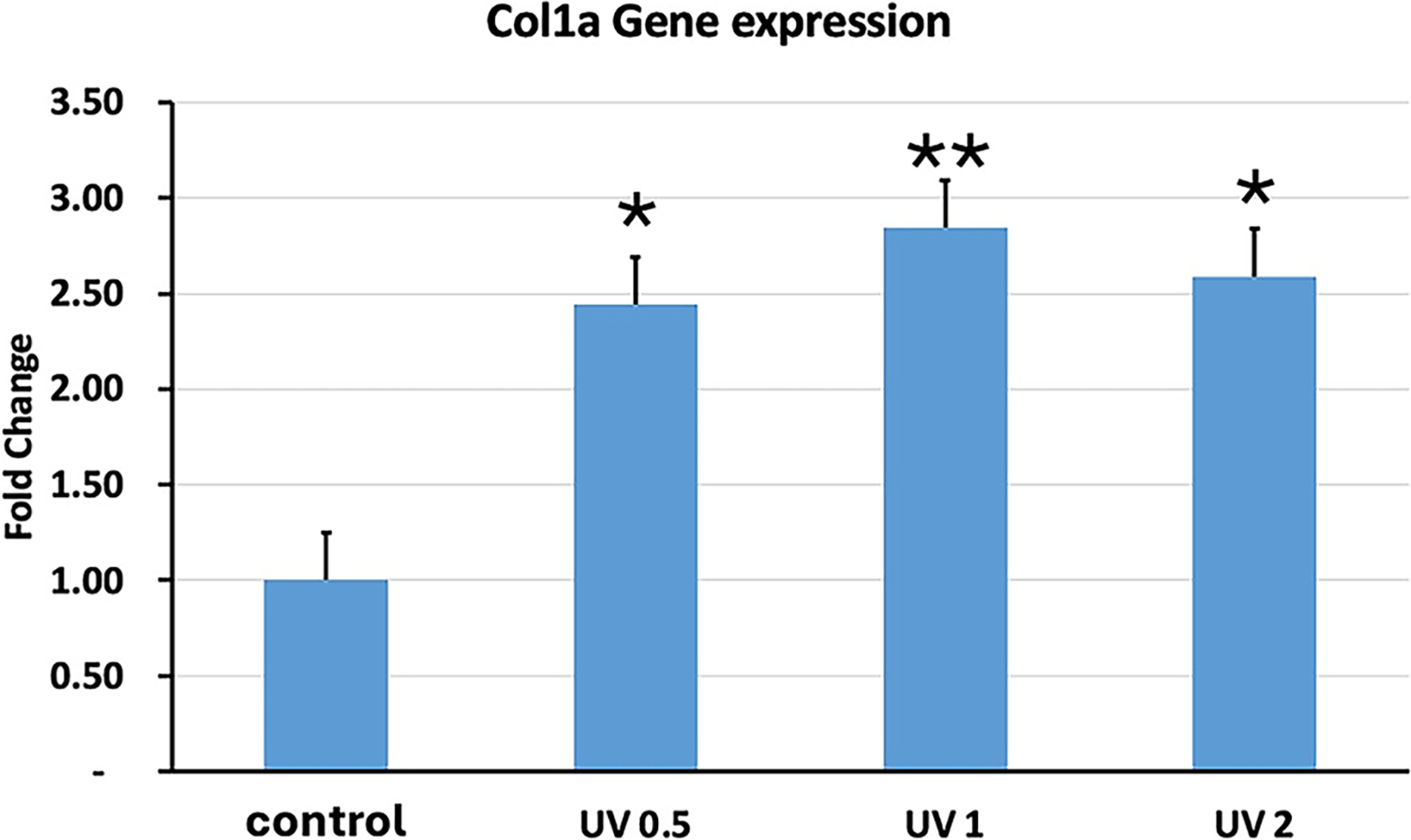
Quantitative PCR (qPCR) analysis of Col1a gene expression in human embryonic stem cell-derived mesenchymal stem cells (hESC-MSCs) treated with ultraviolet A photobiomodulation (UVA PBM) at energy densities of 0.5, 1.0, and 2.0 J/cm². The fold change in Col1a expression was normalized to that of the untreated control (control). The data revealed significant upregulation of Col1a expression at 0.5 J/cm² and 1.0 J/cm², with a slight reduction observed at 2.0 J/cm², indicating the dose-dependent regulation of collagen type I gene expression by UVA PBM. Significance of differences between irradiated and controls: *p < 0.01; **p < 0.001.
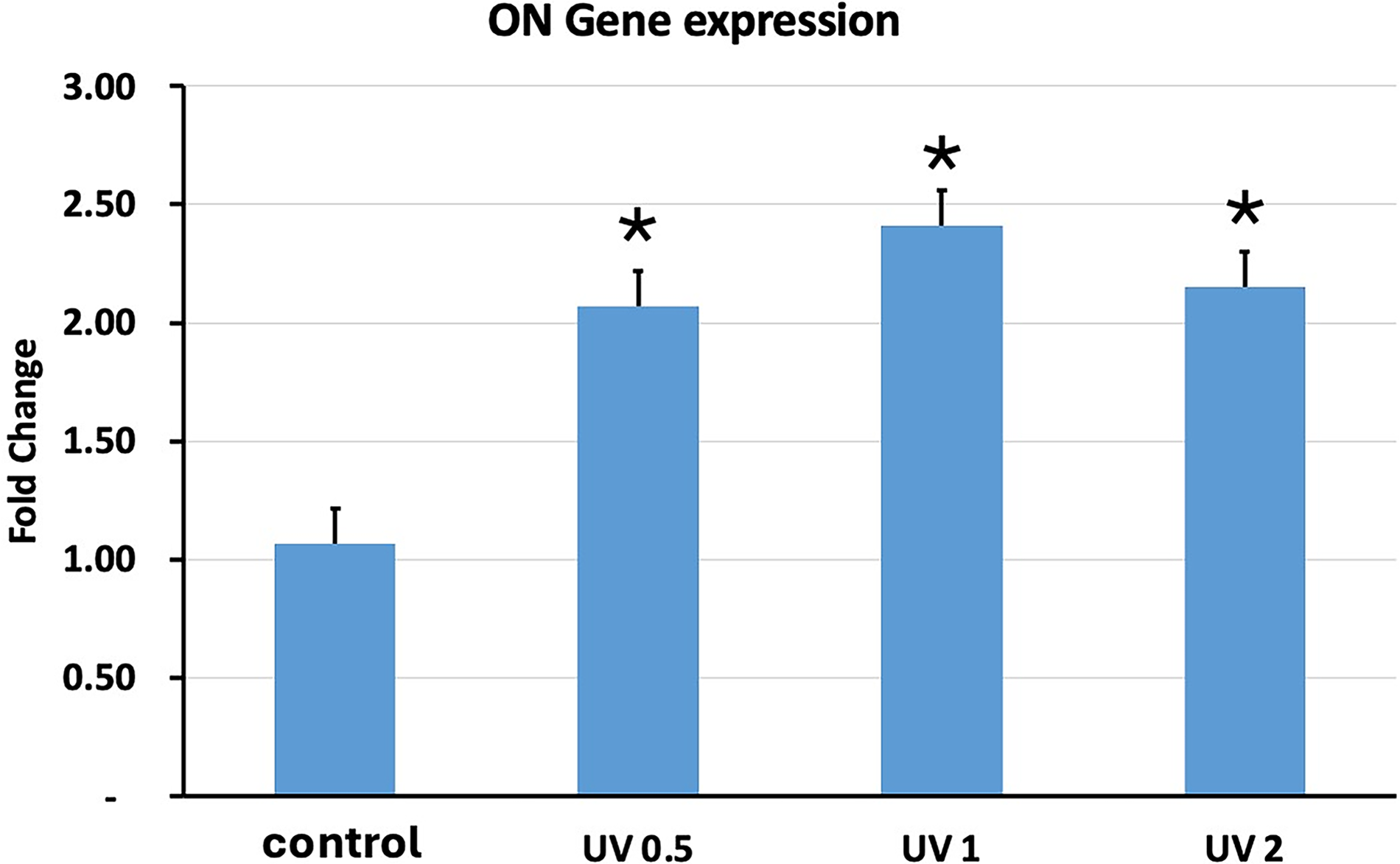
Quantitative PCR (qPCR) analysis of osteonectin (ON) gene expression in human embryonic stem cell-derived mesenchymal stem cells (hESC-MSCs) treated with ultraviolet A photobiomodulation (UVA PBM) at energy densities of 0.5, 1.0, and 2.0 J/cm². The fold change in ON expression was normalized to that of the untreated control (control). The data revealed significant upregulation of ON expression at 0.5 J/cm² and 1.0 J/cm², with a slight decrease at 2.0 J/cm², indicating that moderate UVA PBM enhances ON gene activation, whereas increased energy may reduce ON expression. Significance of differences between irradiated and controls: *p < 0.01.
Discussion
Photobiomodulation can enhance or inhibit biological parameters depending on the application of wavelengths and energy densities. PBM improves the therapeutic effectiveness of stem cell therapy through its ability to enhance the differentiation of stem cells. 20 The PBM has a nonionizing property, as it does not generate heat, sound, or vibration. 21 PBM enhances the proliferation of various cell types, such as keratinocytes, 22 fibroblasts, 23 osteoblasts, 24 and many other stem cells.25–27 The aim of the present investigation was to enhance biological parameters, including the proliferation and differentiation of hESC-MSCs, via UVA PBM irradiation. In this study, different energy densities of UVA PBM at a wavelength of 355 nm were applied to hESC-MSCs, and their proliferation, viability, migration, and differentiation were examined.
In this investigation, cell proliferation was evaluated via the MTT assay, which is based on the conversion of a substrate containing a tetrazolium ring into blue formazan by mitochondrial dehydrogenases. In this study, energy densities ranging from 0.5 to 5.0 J/cm2 were used to observe the effects of low doses of UVA PBM on the proliferation of hESC-MSCs and the inhibitory effects of high doses of UVA PBM. PBM has been reported to reduce the metabolism of cells and damage photoreceptors at high doses, which leads to cell death. 28 During the MTT assay, the optical densities of the treated and untreated (control) groups were measured after irradiation and compared with those of the control groups. These results showed that UVA PBM significantly increased proliferation at low energy densities (0.5–2.0 J/cm2). On the other hand, high energy densities inhibited the growth of hESC-MSCs. Our results are similar to those of a previously published study in which UV light enhanced proliferation at 3.0 J/cm2, 29 which strengthens our results. Moreover, it has been reported in previous studies that PBM increases the proliferation of fibroblasts and epithelial cells1,2 and increases the proliferation of osteoblast cells with calcification-related genes such as osteoglycin. 5 These data support our investigation.
Viability, a biological parameter, was measured via trypan blue staining. It is a blue dye. The unstained viable cells indicate living cells, and the stained nonviable cells indicate dead cells. 17 Our results showed that UVA PBM significantly increased the viability of hESC-MSCs from 0.5 to 3.0 J/cm2 at lower energy densities. Another biological parameter, migration, was measured via a scratch assay to observe the effects of UVA PBM. Migration is a simple, effortless, and low-cost method, 17 in which a scratch or gap area is created with the help of sterile micropipette tips in a 48-well plate containing hESC-MSCs. Our results revealed that UVA PBM at 0.5–2.0 J/cm2 significantly increased the number of migrated hESC-MSCs to close the gap. These results revealed that there was an increase in the number of UVA PBM-treated hESC-MSCs compared with the control, and the cells were moved to the center to close the gap after irradiation. In addition, photomicrographs also supported these results. Phase-contrast photographs of hESC-MSCs were taken after UVA PBM irradiation to observe the migration of hESC-MSCs (Fig. 2). It was observed that hESC-MSCs treated with low energy densities moved toward the center of the wells, which indicated that cell proliferation occurred. Our findings show that UVA PBM irradiation at lower energy densities (0.5–1.0 J/cm2) significantly enhanced hESC-MSC proliferation, viability, migration, and osteogenic differentiation. These results agree with the established concept of photobiomodulation, where low doses of light stimulate biological activity while higher doses may have inhibitory effects due to oxidative stress or photodamage.30,31
The increase in viability after UVA PBM, confirmed by Trypan blue exclusion, is consistent with earlier studies showing that low-level laser therapy improves mitochondrial activity and Adenosine Triphosphate (ATP) production in MSCs and fibroblasts.21,32 Enhanced migration observed in the scratch assay further supports reports that photobiomodulation promotes cytoskeletal rearrangements and motility, processes that are essential for wound healing.33,34 Together, these findings highlight UVA PBM as a potential adjunct to stem cell-based regenerative therapies.
Flow cytometry analysis confirmed preservation of the mesenchymal phenotype with high CD29 and CD73 expression and absence of hematopoietic (CD14, CD34) and endothelial (CD31) markers. These results comply with International Society for Cell & Gene Therapy (ISCT) guidelines for MSC characterization. 35 Interestingly, CD44 expression was lower than expected, which is consistent with other reports showing reduced CD44 levels in embryonic-derived MSCs compared with bone marrow MSCs. 36
Osteogenic differentiation was enhanced at low to moderate UVA PBM doses, as shown by increased ALP staining and upregulation of ALP, Runx2, Col1a, and Osteonectin. These findings align with previous studies demonstrating that photobiomodulation stimulates osteogenesis in MSCs by activating Mitogen-Activated Protein Kinase/Extracellular Signal-Regulated Kinase (MAPK/ERK) and Wnt/β-catenin pathways.37,38 However, a biphasic effect was observed, with reduced ALP and Runx2 expression at the highest dose (2.0 J/cm2). This pattern is consistent with previous PBM studies showing that higher doses can activate stress responses and reduce differentiation potential.39–41 In contrast, BMP4 expression continued to rise at higher doses, suggesting activation of compensatory or stress-related osteogenic signaling. 39
Most prior PBM research has focused on red and near-infrared wavelengths,32,37,40 whereas our study extends these findings by demonstrating that UV-A light, when carefully dosed, can also enhance stem cell function. Low-dose UV-A has been reported to induce beneficial ROS signaling without causing DNA damage, supporting our observations. 42 These results confirm that UVA PBM has a dose-dependent effect, with optimal stimulation at moderate doses and suppression at higher doses. From a translational perspective, UVA PBM may be particularly useful in dentistry and orthopedics, where bone exposure during procedures such as extractions, implants, and periodontal surgery requires rapid healing. By enhancing proliferation, migration, and osteogenic gene expression, UVA PBM could improve clinical outcomes in bone repair. This is in line with previous reports on the positive effects of low-level laser therapy in periodontal and bone regeneration.43,44
In summary, UVA PBM at low energy densities enhances MSC proliferation, migration, and osteogenic differentiation, consistent with previous PBM studies. These findings suggest that UVA PBM can be developed as a novel tool for regenerative medicine, though further mechanistic studies and long-term safety evaluations are needed before clinical translation.
Conclusions
This study demonstrates that UVA PBM irradiation exerts dose-dependent effects on hESC-MSCs. At low to moderate energy densities (0.5–1.0 J/cm2), UVA PBM significantly enhanced proliferation, viability, migration, and osteogenic differentiation, with increased expression of key markers including ALP, Runx2, BMP4, Col1a, and Osteonectin. In contrast, higher energy densities (2.0 J/cm2) led to a decline in some markers, suggesting a threshold beyond which photobiomodulation may become inhibitory. FACS analysis confirmed preservation of the mesenchymal phenotype across all treatment groups, with consistent expression of CD29, CD73, and CD146 and absence of hematopoietic and endothelial markers. The modest increase in CD44 expression with higher doses indicates subtle phenotypic changes but not lineage drift. Taken together, these findings highlight that UVA PBM, when carefully dosed, can promote osteogenic differentiation while maintaining MSC stability. This suggests potential applications in regenerative medicine and dental bone tissue engineering. However, further studies are required to clarify the underlying molecular mechanisms, confirm long-term safety, and optimize treatment parameters before translation into clinical practice.
Footnotes
Acknowledgement
This work was funded by the National Plan for Science, Technology and Innovation (MAARIFAH) King Abdul-Aziz City for Science and Technology, Kingdom of Saudi Arabia, grant number 13-MED-1390-02.
Author Disclosure Statement
The authors declare that they have no conflict of interest.