
Abstract
Background:
Alzheimer’s disease (AD) is a leading cause of dementia in older adults, and effective and widely applicable treatment options remain limited. Photobiomodulation (PBM) has shown promise for AD. However, reported estimates of the delivered dose after transcranial propagation vary widely, limiting translation from animal models to clinical settings.
Objective:
Building on our team’s prior clinical findings, this study evaluated whether an 810 nm/10 Hz pulsed PBM regimen improves cognitive performance and reduces Aβ42 burden in APP/PS1 mice.
Methods:
APP/PS1 mice received PBM using an 810 nm LED pulsed at 10 Hz. Irradiation was delivered for 540 sec/day, 6 days/week, for 7 weeks, with a scalp-surface power density of 0.025 W/cm2 and an energy density of 13.5 J/cm2. Cognitive function was evaluated using the Morris water maze, and Aβ42 burden was quantified by immunofluorescence.
Results:
Cortical and hippocampal Aβ42 plaque burden was reduced, p < 0.01. Exploratory correlation analyses suggested an association between hippocampal Aβ42 plaque number and reversal-learning performance in the histological subset, p = 0.02. The microglia-Aβ42 colocalization ratio increased by 7.53%, p = 0.03, indicating enhanced spatial association between microglia and Aβ42 after PBM.
Conclusions:
These findings support further evaluation of this 810 nm/10 Hz pulsed PBM regimen in AD mouse models and highlight the value of standardized PBM parameter reporting in preclinical studies.
Introduction
Alzheimer’s disease (AD) affects ∼36 million people worldwide, and its prevalence is increasing rapidly. 1 It is the fifth leading cause of death among adults aged 65 years and older. 2 By 2050, the global societal burden is projected to reach $9.1 trillion, largely driven by population aging. 3 At present, no curative therapy is available, and existing pharmacological interventions are primarily symptomatic and do not halt disease progression. 4 Accordingly, there is an urgent need to develop effective therapeutic and preventive strategies.
In recent years, photobiomodulation (PBM) has been widely investigated as a potential intervention for neurodegenerative disorders because it is noninvasive, can be delivered at low irradiance, is minimally associated with thermal injury, and has readily tunable dosimetric parameters. 5 The mechanisms underlying PBM are complex and involve multiple targets and pathways. A commonly cited target is mitochondrial cytochrome c oxidase, whose redox state may be modulated by PBM, thereby enhancing ATP production and modulating reactive oxygen species levels.4,6 Water molecules have also been proposed as photosensitive receptors that may contribute to signal transduction through vibrational energy transfer.7,8 The amyloid cascade hypothesis posits that amyloid beta (Aβ) burden in the brains of patients with AD may drive neuronal loss and cognitive decline. 9 PBM has been reported to confer neuroprotective effects via multiple routes, including the mitigation of mitochondrial dysfunction, modulation of microglial responses, and reduced Aβ burden.10–12 Prior studies suggest that PBM-associated reductions in Aβ burden and improvements in cognition may be associated with microglial involvement in Aβ-related pathology.13–16 However, no consensus standards exist for PBM transcranial penetration or target-tissue dose. 17 Accordingly, direct extrapolation of animal findings to clinical settings remains highly uncertain.
Our team previously reported that whole-brain near-infrared light therapy delivered at 810 nm and 10 Hz, with a power density of 0.09–0.10 W/cm2, produced a significant 3.2-point improvement in Mini-Mental State Examination (MMSE) scores relative to baseline (p = 0.02), and this improvement persisted for at least 2 months after treatment. 18 Building on these findings, the present study evaluated an 810 nm/10 Hz PBM regimen in APP/PS1 mice while adhering to safety protocols. This design provided a controlled preclinical evaluation of the parameter set by assessing post-treatment cognitive performance and Aβ markers in mice.
In this study, mice received whole-brain LED irradiation at a scalp-surface power density of 0.025 W/cm2 for 540 sec/day, corresponding to an energy density of 13.5 J/cm2. Treatment was delivered six times per week for 7 weeks using 10 Hz pulsed light. We hypothesized that PBM would improve performance in the reversal phase of the Morris water maze (MWM) while reducing Aβ42 burden. PBM was associated with microglial morphological changes and increased Aβ42-microglia colocalization in the cerebral cortex and hippocampus, consistent with microglial involvement in the tissue response to Aβ42.
Materials and Methods
Research design
Male APP/PS1 double-transgenic mice aged 5 months and weighing ∼23 g (n = 40; PrP-hAPP K595N/M596L [APPswe] × PrP-hPS1 dE9) were obtained from Beijing HFK Bioscience Co., Ltd. The mice were black, and their scalp hair was shaved before irradiation. In the APP/PS1 line, substantial Aβ burden typically begins to accumulate around 4–5 months of age, 19 while more significant behavioral deficits are more commonly observed at the later stage, around 7 months of age.20,21 Mice were maintained under specific pathogen-free conditions and acclimatized for 7 days before the experiment. Mice were eligible for inclusion based on genotype, sex, age, and health status at enrollment. Sample size was determined according to Festing and Altman’s guidelines and supported by an a priori power analysis in G*Power v3.1.9.7 (α = 0.05, power = 0.80, effect size d = 0.92), which indicated 20 mice per group. 22 At enrollment, each mouse was assigned a unique code, the scalp was shaved, and mice were randomized at the individual level to the AD_Nir or AD_Sham group using a permuted coding sequence (n = 20/group). The individual mouse was regarded as the experimental unit. No animals were excluded from treatment or behavioral analyses after group allocation, and the exact n for each analysis is reported in the corresponding results section or figure legend. To minimize potential confounding, irradiation was performed at a fixed time each day, mice were handled in a random order during treatment, and cage positions were rotated weekly within the rack. Allocation concealment was not used. Because the PBM device emitted visible red light, treatment operators were aware of group allocation during irradiation. However, husbandry staff were blinded to group assignment, and behavioral assessments were performed without reference to animal ID codes or allocation lists.
As shown in Fig. 1a, the experiment lasted 7 weeks, and procedures were conducted 6 days/week. From weeks 1 to 7, PBM was administered daily at 9:00 AM for 9 min. The PBM parameters were 810 nm wavelength, 25 mW/cm2 power density, 10 Hz pulse frequency, and 50% duty cycle (Table 1; Fig. 1b). The AD_Nir group received active PBM, whereas the AD_Sham group underwent identical handling with the device inactive. During each session, awake mice were gently placed in a custom restrainer that limited gross body movement while leaving the shaved dorsal scalp exposed to the LED array; sham-treated mice were restrained for the same duration with the LED array turned off. Thus, restraint and handling procedures were matched between groups, differing only in whether the LED array was active. MWM testing was performed during weeks 6 and 7 (MWM_Week1 and MWM_Week2). After behavioral testing, brain tissue was collected for subsequent analyses. All procedures followed the 3Rs principles and were approved by the Animal Ethics Committee of Beihang University (BM20210156).
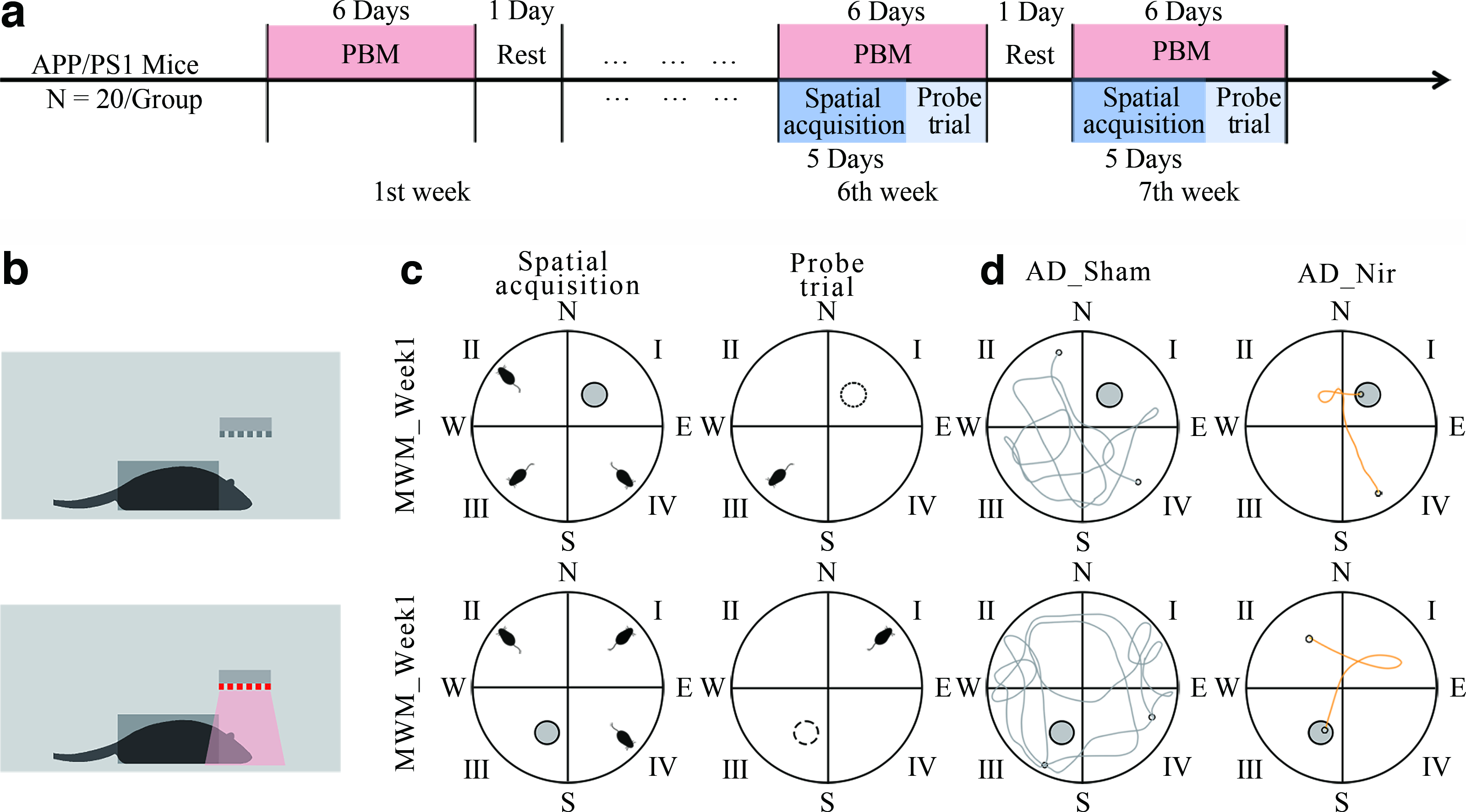
Experimental procedure.
Irradiation Conditions and Energy
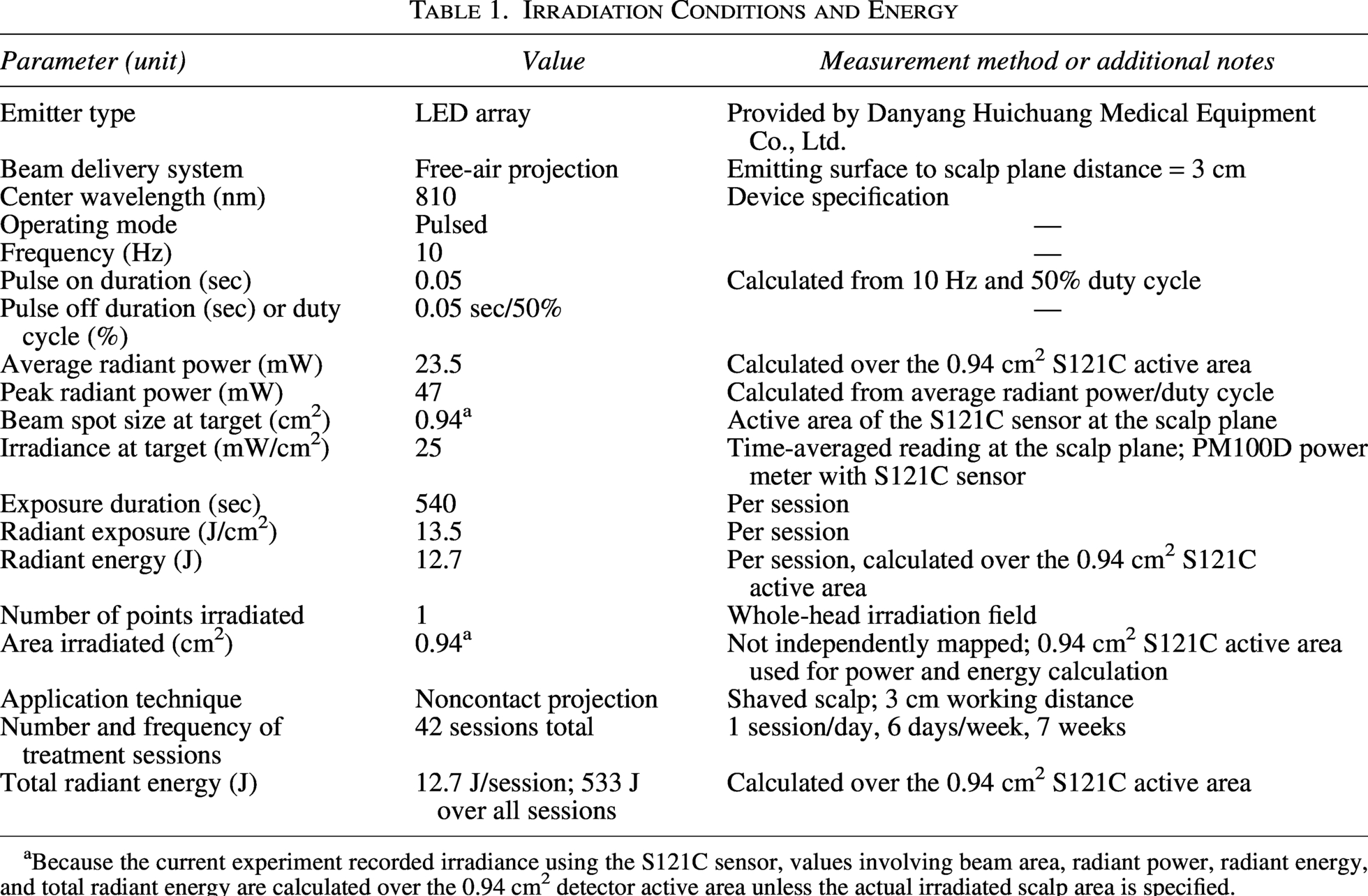
Because the current experiment recorded irradiance using the S121C sensor, values involving beam area, radiant power, radiant energy, and total radiant energy are calculated over the 0.94 cm2 detector active area unless the actual irradiated scalp area is specified.
Measurement of light transmittance in the mouse scalp-skull preparation
Near-infrared light was delivered using an LED panel, and optical power was measured with a digital power meter (PM100D; Thorlabs). Four 8-week-old male C57BL/6J mice were euthanized under deep isoflurane anesthesia. The APP/PS1 transgenic mouse line had been backcrossed onto a C57BL/6J background. 23 In addition, male C57BL/6J mice reach adult craniofacial shape by approximately postnatal day 60, supporting the use of 8-week-old mice as a young-adult cranial reference. 24
Brain tissue was removed while preserving the integrity of the scalp and parietal bone. Scalp-skull transmittance was recorded at randomly selected locations on the LED panel. Four mice, designated M-1 to M-4, were assessed, with 20–22 repeated measurements per mouse, yielding 82 datasets in total.
Treatment parameters
Based on our team’s prior clinical research, 18 this study used an LED array at 810 nm, pulsed at 10 Hz, with a time-averaged irradiance at the scalp plane of 25 mW/cm2. At 810 nm, tissue heating is minimal, 25 and modeling indicates that brain temperature increases by only 0.14°C under 0.1 W/cm2 illumination,26,27 suggesting that thermal contributions are negligible. To facilitate parameter comparability, we used radiant exposure and transmittance-based estimated cortical exposure as contextual metrics. 28 In the present mouse study, the scalp-surface single-session radiant exposure was 13.50 J/cm2. With a measured scalp-skull transmittance of 11–13%, the estimated cortical single-session radiant exposure was 1.49–1.76 J/cm2. For humans, we adopted a transmittance of 0.5–3% for 810 nm light through the scalp and skull.29–32 Accordingly, the surface single-session radiant exposure in the clinical study was 162–180 J/cm2, corresponding to an estimated cortical radiant exposure of 0.81–5.40 J/cm2. 18 These ranges partly overlapped with the estimated cortical radiant exposure in mice and fell within the reported effective radiant exposure window for cytochrome c oxidase of 0.3–3 J/cm2.33,34 These estimates are consistent with prior parameter settings and support the use of 0.025 W/cm2 for the animal experiments. 14 This comparison was used only to provide a parameter-selection context for the present animal experiment.
Morris water maze task
The MWM was used to assess spatial learning and memory (Fig. 1c) in accordance with the guidelines of Vorhees et al. 35 In MWM_Week1, mice underwent 5 days of spatial acquisition training. Each day, mice were released from the quadrant opposite the platform location while facing the pool wall and were given 60 sec to find the hidden platform. Mice that located the platform remained there for 15 sec; those that did not were guided to the platform and also remained there for 15 sec. Mean escape latency was recorded. A probe trial was performed 24 h after training by removing the platform and releasing mice from the quadrant opposite the original platform location, and swimming trajectories were recorded for 120 sec. In MWM_Week2, reversal learning was assessed by relocating the platform to the opposite quadrant. Mice completed three trials per day for 5 consecutive days, and the escape latency was recorded. A second probe trial was conducted 24 h later, with swimming trajectories recorded for 120 sec. Figure 1d shows typical swimming paths of the two groups of mice.
Paraffin section preparation
Mouse brains were fixed and processed for paraffin sectioning as previously described. 36 Immunofluorescence staining was performed on samples from each group (initially n = 6/group). 14 To support technical reproducibility and averaging, multiple nonoverlapping fields of view were acquired per animal. Samples with failed or inadequate staining were excluded from analysis. Following quality control, the final sample sizes were n = 4 in the AD_Sham group and n = 6 in the AD_Nir group.
Fixed specimens were removed from the fixative, rinsed, dehydrated, cleared, and embedded in paraffin to generate wax blocks. Blocks were chilled on ice for ∼30 sec, and two 5 μm sections were cut. Sections were precleared in 30% ethanol, flattened in a drying oven preheated to 42°C, air-dried at room temperature, and then placed on a 57°C drying rack for 30 min for preservation.
Tyramide signal amplification double staining
Aβ42 and Iba1 colocalization was assessed using tyramide signal amplification (TSA)-based double immunofluorescence.37,38 Paraffin sections were dewaxed and rehydrated, followed by antigen retrieval in citrate buffer at pH 6.0 at high temperature for 30 min. Sections were then incubated with 3% hydrogen peroxide in the dark for 10 min. The Iba1 primary antibody (Abcam ab178846) was applied overnight at 4°C, followed by the HRP-conjugated polymer secondary antibody MaxVision for 30 min at room temperature. TSA-FITC was then applied for 30 min at room temperature in the dark for signal development.
To remove residual antibody, sections underwent high-temperature treatment in antigen retrieval solution for 30 min. After cooling, sections were blocked with 3% BSA for 30 min and incubated overnight at 4°C with the Aβ42 primary antibody (Abcam ab201061). Sections were then incubated for 1 h at room temperature with a Cy3-labeled donkey anti-rabbit secondary antibody. Autofluorescence was quenched for 30 min, followed by 4′,6-diamidino-2-phenylindole (DAPI) counterstaining for 10 min in the dark. Sections were mounted with an anti-fade medium and scanned at 40× magnification.
Histological analysis
Immunofluorescence images were quantified in ImageJ. For each mouse, eight nonoverlapping fields of view measuring 300 × 300 μm were analyzed, including six cortical fields and two hippocampal fields, to evaluate microglial morphology, Aβ42 plaque area, and Aβ42 plaque number. 14 Aβ42 plaque area was defined as the total Aβ42-positive area (μm2) within the analyzed fields, and Aβ42 plaque number was defined as the number of discrete Aβ42-positive plaques within the same fields.
Statistical analysis
The primary outcome was MWM performance, which guided sample size calculation and hypothesis testing. Measures included escape latency during MWM_Week1 and MWM_Week2, platform crossings in the probe trial, target-quadrant swim distance, average swimming velocity, and total swim distance. Secondary outcomes included Aβ42 burden and microglial morphology. Analyses were performed using GraphPad Prism 9.0 and IBM SPSS Statistics. Normality was assessed with the Lilliefors test. For two-group comparisons, unpaired Student’s t tests or Welch’s t tests were used for normally distributed data, as appropriate; otherwise, Mann–Whitney U tests were applied. Effect sizes were reported as Cohen’s d for t tests, rank-biserial correlation for Mann–Whitney U tests, and partial η2 for repeated-measures analysis of variance (ANOVA). Pearson’s r and its 95% confidence interval (CI) were reported for correlation analyses. Repeated-measures ANOVA was used for day-wise MWM outcomes, with time as the within-subject factor, group as the between-subject factor, and time × group interaction included in the model. The Greenhouse–Geisser correction was applied when sphericity was violated. For daily post hoc between-group comparisons, p values were corrected using the false discovery rate method. Statistical significance was defined as p < 0.05, and data are presented as mean ± standard error of the mean. Data analysis was performed blinded to group allocation.
Results
Transmittance of the mouse scalp-skull preparation
Transmittance is summarized in Table 2. Mean transmittance was 0.12 ± 0.01, with a 95% CI of 0.11–0.12 and a standard error close to zero. Under the present experimental conditions, transmittance was stable with minimal inter-individual variability. Accordingly, values of 0.11–0.13 were used as representative inputs for simplified transmittance-based dose estimation.
Transmittance
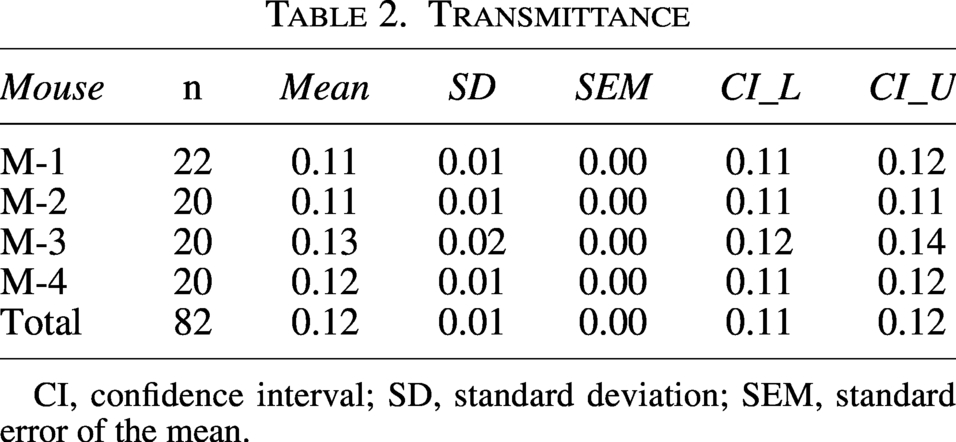
CI, confidence interval; SD, standard deviation; SEM, standard error of the mean.
For MWM_Week1, the day-by-group interaction for average swimming velocity was not significant, whereas a significant time effect was observed, F = 5.21, p < 0.01, partial η2 = 0.12 (Fig. 2a). In MWM_Week2, average swimming velocity likewise showed a significant time effect, F = 15.67, p < 0.01, partial η2 = 0.29 (Fig. 2f), while neither the day-by-group interaction nor the between-group effect indicated a pattern suggestive of altered training-related motor performance, despite a significant group effect, F = 6.51, p = 0.02, partial η2 = 0.15. Swimming distance in MWM_Week1 also showed a significant time effect, F = 13.79, p < 0.01, partial η2 = 0.27 (Fig. 2b), whereas the day-by-group interaction was not significant. In MWM_Week2, neither the time effect nor the day-by-group interaction for swimming distance reached significance, with F = 2.45, p = 0.06, partial η2 = 0.06 (Fig. 2g) and F = 0.96, p = 0.43, partial η2 = 0.03, respectively. Overall, these findings suggest that PBM did not produce a consistent effect on general swimming performance across training days.
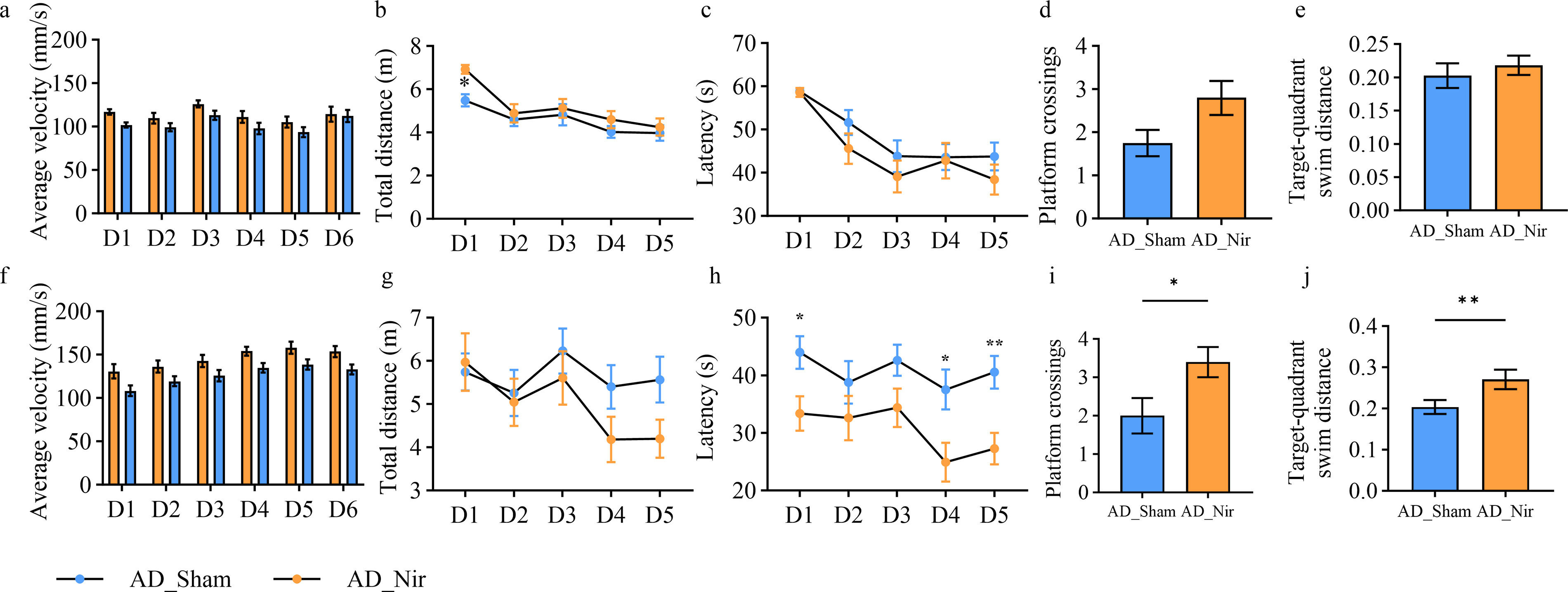
MWM test results.
For escape latency, MWM_Week1 showed a significant time effect, F = 16.66, p < 0.01, partial η2 = 0.31 (Fig. 2c), whereas neither the day-by-group interaction nor the between-group effect was significant. In MWM_Week2, escape latency also demonstrated a significant time effect, F = 2.62, p = 0.04, partial η2 = 0.07 (Fig. 2h), with no significant day-by-group interaction, whereas the between-group effect was significant, F = 12.42, p < 0.01, partial η2 = 0.25. After assessing normality for daily latency in MWM_Week2, day-wise comparisons using unpaired t tests or Mann–Whitney U tests with FDR correction identified significant between-group differences on days 1, 4, and 5.
In the MWM_Week1 probe trial, AD_Nir mice showed more platform crossings (Fig. 2d) and greater target-quadrant swim distance (Fig. 2e); however, these differences did not reach statistical significance. After platform relocation in MWM_Week2, significant between-group differences emerged in both platform crossings, p = 0.02 (Fig. 2i), and target-quadrant swim distance, p = 0.03 (Fig. 2j).
PBM reduces Aβ42 burden in mice
Immunofluorescence showed that the AD_Nir group had a significantly lower total Aβ42 plaque burden than the AD_Sham group. The number of plaques was reduced in AD_Nir mice by 57.43% in the cortex, p < 0.01 (Fig. 3b), and by 60.00% in the hippocampus, p < 0.01 (Fig. 3e). Aβ42 plaque area decreased by 42.87% in the cortex, p < 0.01 (Fig. 3c), and by 62.05% in the hippocampus, p < 0.01 (Fig. 3f). In the AD_Nir group, cortical Aβ42 plaque number showed a positive but nonsignificant association with MWM_Week1 probe-trial performance, r = 0.76, 95% CI [−0.15, 0.97], p = 0.08 (Fig. 3d). Hippocampal Aβ42 plaque number was positively correlated with day 5 escape latency in MWM_Week2 reversal learning, r = 0.89, 95% CI [0.27, 0.99], p = 0.02 (Fig. 3g), but this association should also be interpreted cautiously as exploratory due to the small histological subset.
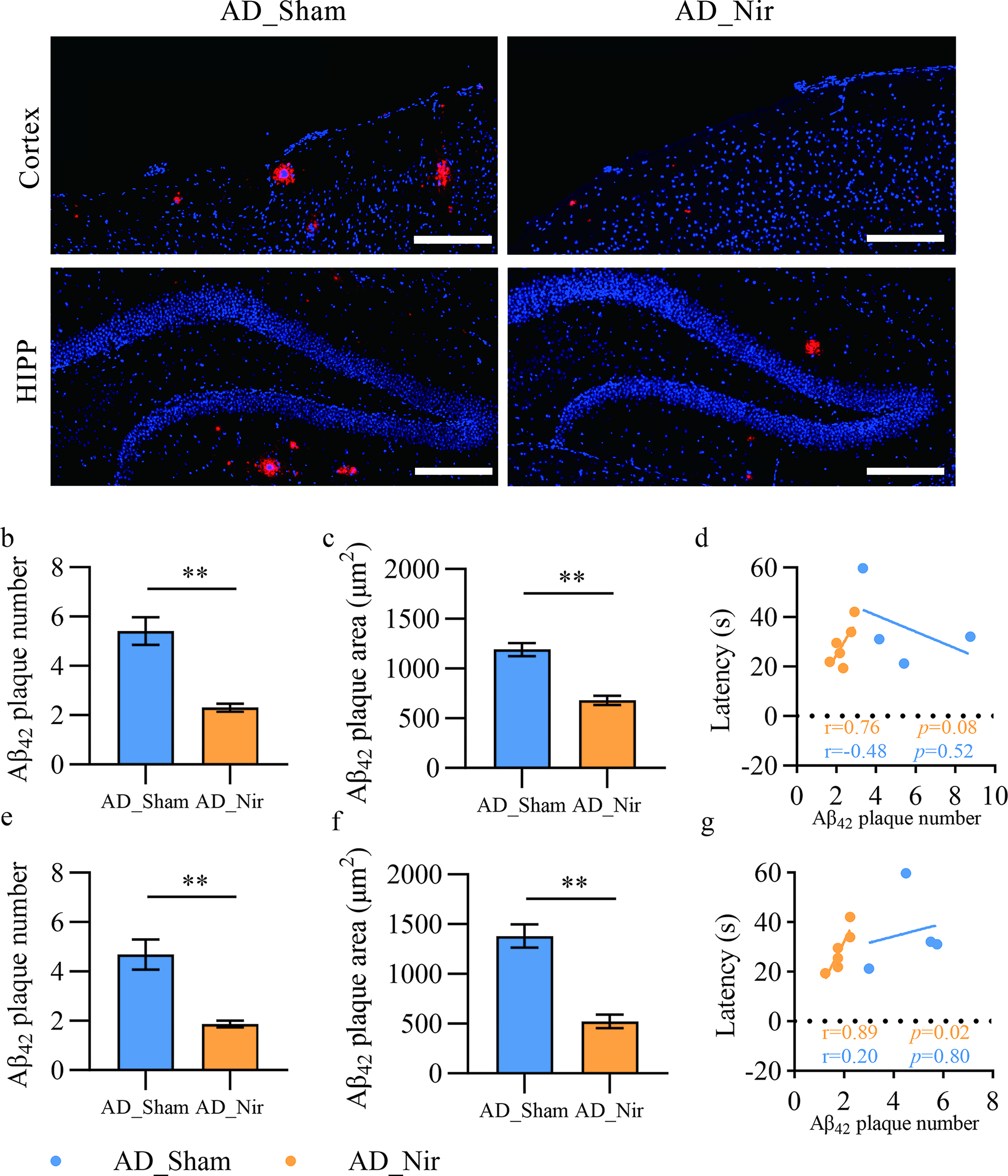
810 nm light reduces amyloid burden in APP/PS1 mice.
PBM is associated with microglial remodeling and reduced Aβ42 burden
Compared with AD_Sham mice, AD_Nir mice showed significant microglial morphological changes in the cortex: average branch length decreased by 12.18%, p < 0.01 (Fig. 4e), and maximum branch length decreased by 12.38%, p < 0.01 (Fig. 4f). The Aβ42-microglia colocalization ratio increased by 7.53%, p = 0.03 (Fig. 4g). In AD_Nir, the cortical Aβ42-microglia colocalization ratio was positively correlated with platform crossings in the MWM_Week2 probe trial, r = 0.97, 95% CI [0.74, 0.99], p < 0.01 (Fig. 4h). In the hippocampus, both the number of microglial branches and the number of junctions were reduced, p = 0.03 for each (Fig. 4i and j). Within AD_Nir, the number of hippocampal microglial branches was negatively correlated with the percentage of target-quadrant swim distance during MWM_Week1, r = −0.86, 95% CI [−0.98, −0.14], p = 0.03 (Fig. 4k). The number of hippocampal microglial junctions showed a similar negative correlation, r = −0.84, 95% CI [−0.98, −0.01], p = 0.04 (Fig. 4l).
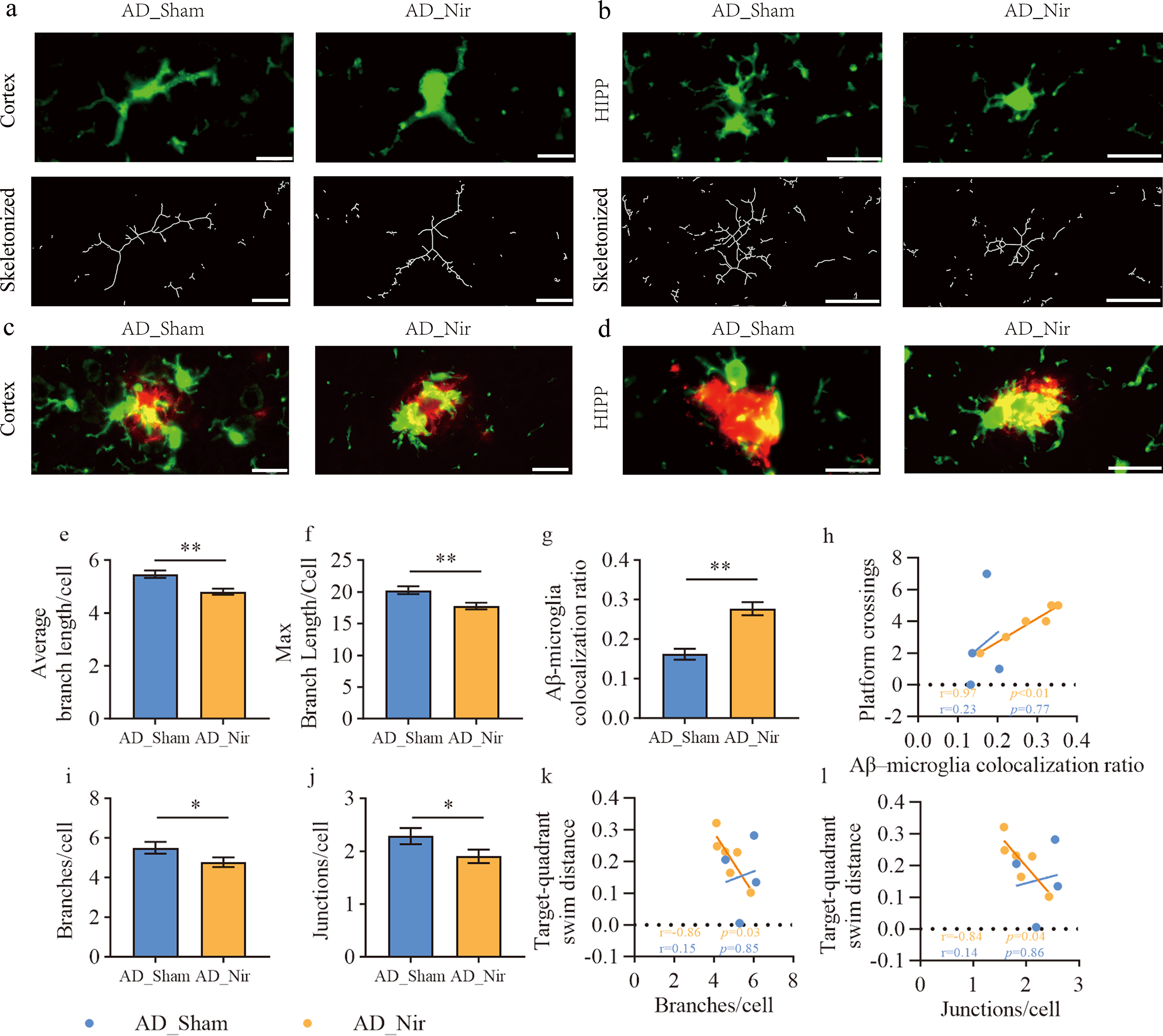
810 nm light is associated with microglial remodeling and increased Aβ-microglia colocalization in APP/PS1 mice.
Discussion
Building on our prior 810 nm clinical findings, 18 we evaluated a clinically inspired PBM regimen in APP/PS1 mice, whose surface irradiance and exposure duration were selected with reference to transmittance-based dose estimates. The regimen was selected with reference to prior clinical use and simplified transmittance-based exposure estimates, which were used to provide dosimetric context for parameter selection rather than to establish a validated mouse-to-human dose conversion. Accordingly, APP/PS1 mice received PBM with an 810 nm pulsed LED at a scalp-surface irradiance of 0.025 W/cm2 and an energy density of 13.5 J/cm2, delivered at 10 Hz for 540 sec/day, 6 days/week for 7 weeks. In the MWM, groups differed in MWM_Week2 reversal learning, indicating improved performance in the reversal phase. 35 At the pathological level, intracerebral Aβ burden was significantly reduced and showed a positive but exploratory association with behavioral performance. Moreover, PBM was associated with microglial morphological changes, increased Aβ42-microglia colocalization, and reduced Aβ42 burden, consistent with a potential involvement of microglia in the tissue response to Aβ42.
In this study, we used a two-phase MWM paradigm to examine the effects of PBM on spatial learning and reversal learning. 33 MWM_Week1 comprised conventional acquisition training in which mice learned and retained the platform location. MWM_Week2 comprised a spatial reversal test in which the platform was moved to the opposite quadrant to assess reversal learning. 25 During MWM_Week1, groups did not differ in escape latency, swim speed, or swim distance, indicating no detectable effects on initial spatial learning or motor performance.39,40 Both groups showed a significant time effect, reflecting progressive acquisition across training days in both groups. In the MWM_Week1 probe trial on day 6, AD_Nir mice showed more platform crossings and greater target-quadrant swim distance, although these differences did not reach statistical significance. In MWM_Week2, AD_Nir mice exhibited shorter escape latencies than AD_Sham mice, indicating a significant between-group effect. During the reversal phase, these MWM_Week2 differences are most consistent with improved reversal-learning performance following PBM.41,42 Nevertheless, because no additional behavioral tasks were included, alternative explanations such as differences in search strategy, perseveration, or anxiety-related influences cannot be fully excluded. In addition, although restraint and handling procedures were matched between groups, and swim velocity and total swim distance did not suggest a consistent motor-performance confound, restraint-related stress was not directly assessed. Further behavioral studies will be needed to define the affected cognitive domain more precisely. 35
The reduction in Aβ burden observed here is consistent with most prior PBM reports.13,14,43,44 In APP/PS1 mice, 10 Hz irradiation at 1070 nm reduced Aβ burden, increased Aβ42-microglia colocalization ratios, and was accompanied by behavioral improvement. 14 Similarly, 808 nm continuous-wave light in AD transgenic rats reduced plaque burden and increased microglial recruitment, consistent with microglial involvement in the response to amyloid pathology. 43 Two additional APP/PS1 studies reported reduced Aβ, improved behavior and energy metabolism, and changes in microglial phenotype and inflammation-related markers after 6 weeks of 808 nm irradiation.13,44 Although PBM can exhibit a biphasic dose response, with benefit at lower doses and inhibition at higher doses, 45 low-dose PBM at 0.006 W/cm2 showed no measurable effect in 5xFAD mice, 46 indicating that efficacy depends on model, disease stage, and dosing regimen. Across studies, the absence of standardized target-tissue dose measurement, together with differences in devices and methods, introduces substantial variability and limits direct comparison of findings. Moreover, the present comparison is based on bulk transmittance scalars and does not capture the three-dimensional distribution of light within tissue, beam divergence, or the heterogeneous multilayer anatomy of the overlying tissues.28,47 These transmittance-based estimates should therefore be interpreted as approximate dosimetric context for parameter selection rather than as evidence of direct mouse-to-human dose transferability. The observed effects should also not be attributed solely to direct photon penetration to the cortex because extracerebral tissues, including scalp vasculature and skull bone marrow, may absorb or scatter part of the incident light and may also participate in neuroimmune responses. Skull bone marrow-meningeal border tissues represent biologically active neuroimmune interfaces that could modify PBM-related responses.48–50 More accurate cross-species light-dose mapping would require Monte Carlo photon-transport simulations in anatomically realistic models, which were not performed in the present study.
Accumulating evidence indicates that microglia play an early protective role in mitigating Aβ42 toxicity. 51 In APP/PS1 mice, 808 nm PBM-associated behavioral improvement, reduced Aβ burden, and modulation of microglial responses may partly reflect mitochondrial targeting.52,53 PBM has been reported to modulate microglial responses in AD models, partly through mitochondrial metabolic regulation. In APP/PS1 mice, 808 nm PBM promoted oxidative phosphorylation and suppressed glycolysis in microglia; this metabolic shift was accompanied by anti-inflammatory and pro-phagocytic M2-like polarization rather than proinflammatory M1-like activation. 13 Aβ reduction associated with microglial remodeling has also been reported after 40 Hz light flicker and 40 Hz transcranial ultrasound stimulation, suggesting that distinct neuromodulatory approaches may converge on microglia-related amyloid clearance pathways.12,54 Beyond microglia, PBM may also support cognitive improvement through neuronal mitochondrial and plasticity-related mechanisms, including reported effects on neurogenesis and synaptogenesis.55,56 Microglia can form protective barriers around amyloid deposits, compacting fibrils into potentially less toxic structures and limiting further Aβ42 accrual on existing plaques. 57 Consistent with this, long-term near-infrared PBM in transgenic AD rats reduced plaque burden and enhanced microglial recruitment to plaques. 43 Cellular and in vivo data further indicate that near-infrared light reduces Aβ-induced microglial toxicity and improves neuronal survival, supporting parallel improvements in immune readouts and Aβ burden. 15 Activated microglia typically display enlarged somata, fewer branches, and shorter branch lengths. 58 In the present study, PBM was associated with microglial morphological changes in the cortex and hippocampus, increased the Aβ42-microglia colocalization ratio, and reduced Aβ42 burden. These findings are consistent with microglial involvement in the tissue response to Aβ42 and may suggest a potential contribution of microglia to the observed reduction in Aβ42 burden under the regimen used here. 59 However, because the present evidence is limited to microglial morphology and Aβ42-microglia colocalization, it does not establish the functional phenotype of microglia or demonstrate a specific microglia-mediated mechanism of Aβ42 clearance. 60 To verify the proposed microglia-related clearance pathway, further mechanistic studies will be required, including assessment of microglial metabolic markers, phagocytosis-related markers, cytokine profiles, and mitochondrial function. 61
Limitations
This study had a limited sample size; notably, the histological and microglial analyses were based on a smaller post-quality-control subset, so effect sizes and correlations may be unstable and require confirmation in larger cohorts. Because no age-matched wild-type group was included, behavioral effects should be interpreted as improvement relative to sham-treated APP/PS1 mice rather than restoration toward normal. The transmittance-based estimates do not fully account for potential contributions from scalp vasculature and skull bone marrow. Future studies should incorporate anatomically realistic Monte Carlo light-transport modeling together with tissue-level experimental validation. Because the device emitted visible red light, the treatment operator could not be blinded. Although handling and treatment procedures were standardized, subtle operator-related bias cannot be excluded; future studies should improve treatment-stage blinding using opaque shielding, visually matched sham devices, or separate personnel. Finally, PBM-related mechanisms, including mitochondrial oxidative phosphorylation, microglial polarization and phagocytosis, neurogenesis, synaptogenesis, and neuronal mitochondrial function, require direct validation in future studies.
Conclusions
This study evaluated an 810 nm/10 Hz pulsed PBM regimen in APP/PS1 mice and reported simplified transmittance-based exposure estimates to improve PBM parameter transparency. APP/PS1 mice received 810 nm pulsed LED irradiation at a scalp-surface irradiance of 0.025 W/cm2 and an energy density of 13.5 J/cm2, delivered at 10 Hz, 6 days/week for 7 weeks. Treated mice showed improved performance in the MWM reversal phase. Aβ burden decreased in the cortex and hippocampus and showed exploratory associations with behavioral improvement in the MWM. An increased Aβ42-microglia colocalization ratio suggested enhanced spatial association between microglia and Aβ42, consistent with microglial involvement in the tissue response to amyloid pathology.
Authors’ Contributions
Y.Z.: Investigation, data curation, writing—original draft, and visualization; H.Q.: methodology and data curation; Z.L.: Investigation; R.G.: Data curation; D.L.: Conceptualization, supervision, and writing—review and editing; W.S.: Formal analysis, visualization, and writing—review and editing; and D.W.: Conceptualization, supervision, and writing—review and editing.
Supplemental Material
sj-docx-1-pho-10.1177_25785478261465371 — Supplemental material for Transcranial 810 nm Pulsed Photobiomodulation Improves Learning and Reduces Aβ42 Burden in APP/PS1 Mouse Model of Alzheimer’s Disease
Supplemental material, sj-docx-1-pho-10.1177_25785478261465371 for Transcranial 810 nm Pulsed Photobiomodulation Improves Learning and Reduces Aβ42 Burden in APP/PS1 Mouse Model of Alzheimer’s Disease by Ye Zhang, Huiting Qiao, Zeping Lv, Rong Guo, Deyu Li, Wei Song, and Daifa Wang
Supplemental Material
sj-docx-2-pho-10.1177_25785478261465371 — Supplemental material for Transcranial 810 nm Pulsed Photobiomodulation Improves Learning and Reduces Aβ42 Burden in APP/PS1 Mouse Model of Alzheimer’s Disease
Supplemental material, sj-docx-2-pho-10.1177_25785478261465371 for Transcranial 810 nm Pulsed Photobiomodulation Improves Learning and Reduces Aβ42 Burden in APP/PS1 Mouse Model of Alzheimer’s Disease by Ye Zhang, Huiting Qiao, Zeping Lv, Rong Guo, Deyu Li, Wei Song, and Daifa Wang
Footnotes
Acknowledgments
The authors extend their thanks to Pony Testing International Group for providing the experimental environment. They gratefully acknowledge Danyang Huichuang Medical Equipment Co., Ltd. for providing the PBM equipment used in this study.
Ethical Considerations
Protocols for animal experiments were approved by the Animal Experimental Ethics Committee of Beihang University (approval no. BM20210156). During the experiment, we strictly adhered to all ethical standards to ensure the welfare of the animals.
Data Availability Statement
Data supporting the findings of this study are available upon request from the corresponding author.
Author Disclosure Statement
D.W. is an associate professor at Beihang University and serves as Chief Scientist at Danyang Huichuang Medical Equipment Co., Ltd., which provided the PBM equipment used in this study. To minimize potential bias, group allocation was randomized, and animal care staff and behavioral assessors were blinded to group allocation. The remaining authors declare no competing interests.
Funding Information
This work was supported by the Major Science and Technology Special Program of Jiangsu Province (BG2024025) and horizontal projects of the integration between industry and education (No. KH54429301).
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.