
Abstract
Working memory (WM) is a capacity-limited control system that maintains and updates task-relevant representations for goal-directed cognition. Because WM declines with aging and is disrupted across neuropsychiatric and neurodegenerative disorders, neuromodulation has emerged as both a causal probe of circuit function and a potential therapeutic adjunct. This mechanistic narrative review evaluates evidence across transcranial direct current stimulation, transcranial magnetic stimulation/theta-burst stimulation, transcranial alternating current stimulation, transcranial random noise stimulation, and deep brain stimulation. Rather than asking whether neuromodulation uniformly enhances WM, we examine when different modalities alter distinct WM subprocesses. Across modalities, effects are generally modest and heterogeneous, but the literature increasingly suggests that stimulation acts less by adding “capacity” than by biasing gain, timing, and plasticity-related control variables that shape WM trajectories. More precise dosing, targeting, and computation-aligned outcomes are needed to establish reproducible and mechanistically interpretable effects.
Introduction
Working memory (WM) comprises a capacity-limited set of control operations that temporarily maintain, manipulate, and prioritize task-relevant information in the service of goal-directed cognition, reasoning, learning, and decision-making. 1 Contemporary accounts no longer treat WM as a passive storage buffer; rather, WM depends on dynamically stabilized representations that must be encoded, protected from interference, refreshed, and updated as task demands evolve.1,2 Crucially, WM is not a unitary endpoint. Common paradigms assay partially dissociable subprocesses, including maintenance, updating, interference control, and representational precision or feature binding, whose neural constraints are overlapping but not identical.1,3 In the present review, cross-study comparisons are therefore interpreted primarily at the level of the computation under examination rather than under a single umbrella WM score.
Individual differences in WM are functionally consequential across development, aging, and disease. WM capacity is closely linked to educational attainment across childhood and adolescence, 4 shows characteristic age-related vulnerability, 5 and is disrupted across major disorders of cognitive control, including schizophrenia and attention-deficit/hyperactivity disorder.6,7 These observations make WM both a mechanistically informative construct and a clinically relevant target for causal intervention.
Neuromodulation has accordingly become an important experimental and translational approach for studying WM. Noninvasive methods, particularly transcranial electrical stimulation (tES; including transcranial direct current stimulation, transcranial alternating current stimulation, and transcranial random noise stimulation) and transcranial magnetic stimulation (TMS), together with invasive approaches such as deep brain stimulation (DBS), provide forms of causal leverage that correlational neuroimaging alone cannot offer.8-11 By perturbing defined cortical or subcortical nodes and quantifying changes in specific WM subprocesses, these methods can test circuit-level hypotheses about maintenance, updating, gating, and interference control. At the same time, the current literature does not support a uniform enhancement account. Reported effects are typically heterogeneous and depend on stimulation modality, target, brain-level dose, baseline state, and task structure.12-16
This article is a mechanistic narrative review of neuromodulation effects on WM across stimulation modalities, targets, and populations. Its primary aim is not to determine whether neuromodulation uniformly improves WM, but to evaluate under what conditions distinct stimulation approaches alter distinct WM subprocesses, and how confidently such effects can be interpreted at circuit and systems levels. We therefore prioritize convergent evidence from recent meta-analyses, representative controlled trials, and studies bearing on target engagement and physiological interpretation. The evidential maturity of the major modalities is not symmetrical: confidence is strongest where the literature is broadest and methodologically most developed, particularly for tDCS and TMS as causal perturbation tools; it is more conditional for tACS and more preliminary or clinically narrower for tRNS and DBS when WM is the primary endpoint.12-16 Table 1 provides a compact comparative overview of representative findings by technique, target population, putative mechanism, and WM outcome domain.
Neuromodulation Modalities for Working Memory (WM): Canonical Targets, Interpretable Populations, Putative Mechanisms, and the Most Consistent Outcome Patterns. This Table Summarizes Where Current Evidence is Most Interpretable Across Modalities, How Each Technique is Most Plausibly Framed Mechanistically Within a Gain-and-Timing Account, and Which WM Domains are Most Commonly Assayed. It is Intended as a Comparative Synthesis of the Present Literature, Not as a Claim of Equivalent Evidentiary Maturity Across Techniques. Accordingly, Cross-Study Comparability Should be Interpreted at the Level of the Specific WM Computation Assayed, such as Maintenance, Updating, Interference Control, or Load-Sensitive Precision Rather Than at the Level of a Generic WM Label.
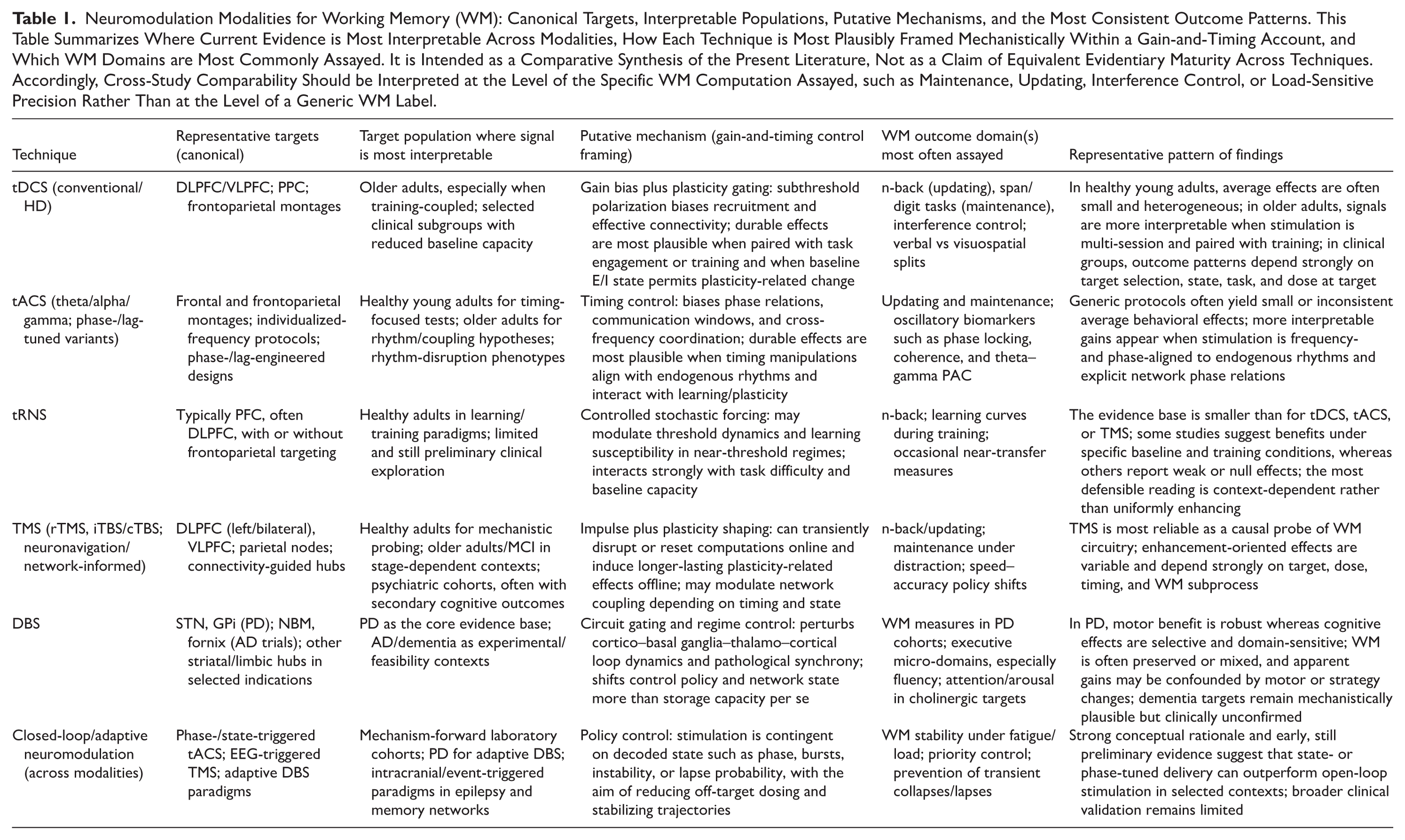
Transcranial Direct Current Stimulation (tDCS)
Biophysical Rationale: Biasing Rather Than Driving
Transcranial direct current stimulation (tDCS) applies a weak constant current at the scalp (classically 1-2 mA) to induce low-amplitude intracranial electric fields that bias neuronal polarization rather than directly elicit action potentials. The traditional anodal-“facilitatory” and cathodal-“inhibitory” heuristic remains serviceable as a first approximation, yet contemporary biophysics emphasizes that net effects in vivo depend on cortical folding, axonal–dendritic orientation relative to the induced field, tissue conductivities, montage geometry, and, critically, the ongoing network state. Polarity rules should therefore be treated as probabilistic tendencies rather than deterministic laws. Mechanistically, tDCS is best framed as a subthreshold gain-biasing intervention that shifts the likelihood that task-relevant ensembles recruit, synchronize, and update under demand, without imposing activity “from the outside.”8,12,17-19
A direct methodological implication follows: “1–2 mA” is a device setting rather than a brain dose. The biologically relevant dose is the intracranial field strength and its spatial distribution at functionally engaged tissue, which can vary substantially across individuals as a function of anatomy and montage. This dose uncertainty contributes importantly to heterogeneous outcomes and motivates current-flow modeling.8,12,17,19
Cellular and Plasticity-Linked Mechanisms
In the context of WM, the most defensible mechanistic claim is not that tDCS reliably boosts momentary performance in isolation, but that it can modulate learning and plasticity when stimulation interacts with ongoing task engagement. Behavioral and physiological after-effects that outlast stimulation are commonly interpreted within plasticity-linked accounts (including NMDA-dependent and calcium-mediated processes) that are strongly state-dependent: identical parameters can yield different outcomes depending on baseline state and task context. This supports a pragmatic interpretation of tDCS as a plasticity gate or learning-rate modulator, rather than a standalone cognitive enhancer.8,19,20
Neurochemical and Systems-Level Effects: Shifting the Plasticity Landscape
At the neurochemical level, converging evidence supports the plausibility that prefrontal tDCS can acutely reshape metabolite profiles measured by magnetic resonance spectroscopy (MRS). A concurrent tDCS–MRS study reported stimulation-associated increases in glutamatergic markers (Glx) in the left DLPFC persisting shortly after stimulation, consistent with a transient shift of the excitation–inhibition balance toward plasticity-permissive regimes. At the same time, such findings underscore that field magnitude and biochemical changes do not map one-to-one across individuals, reinforcing the need for mechanistic readouts rather than relying solely on nominal current. 20
At the systems level, WM computations are implemented by distributed frontoparietal control networks supporting maintenance, updating/gating, and manipulation. Accordingly, tDCS effects are often more interpretable when framed as perturbations of effective connectivity and gain control within executive circuits, rather than as isolated modulation of “DLPFC excitability.” Under this view, beneficial versus deleterious outcomes can emerge depending on whether stimulation stabilizes the network regime required by a given WM demand, or instead destabilizes it under a mismatch between dose, target, and state.8,12,17,19
Healthy Adults: Small Mean Effects, Large Moderation, and Dose–Target Specificity
In healthy young adults, the field has largely moved beyond the broad claim that “tDCS improves WM” toward a more conditional statement: mean effects are often small, while moderators can be large. Meta-modeling work focusing on anodal left prefrontal tDCS has emphasized that behavioral outcomes depend on where induced electric fields are maximized across cortical territories, implying that null results can arise from insufficient dose at functionally relevant tissue, even when scalp current is nominally matched.8,12,17-19,21
Reliability constraints also matter. Even when group means suggest modest effects, within-person effects across sessions may remain unstable unless state and dose are tightly controlled. A large preregistered registered report testing intra-individual reliability underscores the methodological burden of demonstrating repeatable behavioral modulation from single-session, low-dose tDCS. 21
Older Adults: A Clearer Signal When Paired with Training
Evidence is more encouraging in healthy older adults, particularly when tDCS is paired with multi-session cognitive training; a recent meta-analysis reported significant WM improvements, with stronger effects in higher-intensity, extended protocols. 22
Major Depressive Disorder (MDD): Plausible Adjunctive Benefits, Not Yet Definitive for WM
In MDD, executive and WM dysfunction can persist beyond mood symptoms, making prefrontal tDCS an attractive adjunct aimed at restoring control-network dynamics. Recent quantitative syntheses suggest that while tDCS shows antidepressant efficacy and can improve certain cognitive domains, robust and selective enhancement of WM is not consistently demonstrated, in part due to heterogeneous endpoints and protocol variability. 23 A plausible interpretation is that MDD comprises multiple neurocognitive biotypes; without stratification by baseline cognition, medication status, and physiological markers, group-level WM effects may remain diluted. 23
ADHD: Safety is Encouraging; Cognitive Effects Remain Protocol- and Development-Dependent
For ADHD, tDCS is appealing as a scalable and generally well-tolerated option, yet WM outcomes appear strongly contingent on montage, dosing schedule, and developmental neurophysiology. Meta-analytic evidence supports overall safety and suggests that symptomatic improvements can occur under some protocols, while also emphasizing substantial variability and the need for stronger sham-controlled designs with clinically meaningful cognitive endpoints. 24 Conceptually, “more excitability” is not automatically “better computation” in neurodevelopmental populations; maladaptive gain increases could plausibly worsen distractibility or destabilize control policies in subsets of individuals. 24
Schizophrenia: Network-Aware Targeting Beyond DLPFC
In schizophrenia, WM deficits implicate both prefrontal and parietal nodes, and target selection may be as consequential as intensity. A recent randomized double-blind trial comparing DLPFC-tDCS, PPC-tDCS, and sham during concurrent WM engagement reported more favorable WM-related outcomes under PPC targeting in that context, alongside physiological signatures consistent with circuit engagement. 25 These findings support network-aware targeting and suggest that effects may be more interpretable in specific subdomains and subgroups.
Interpretation of tDCS effects remains constrained by uncertainty in delivered brain dose, strong state dependence, and variability in WM outcome definitions. As a result, tDCS findings are best interpreted conditionally rather than as evidence of uniform WM enhancement.
Transcranial Alternating Current Stimulation (tACS)
Conceptual Rationale: Write to the Brain in Its Native Clock
Transcranial alternating current stimulation (tACS) delivers a sinusoidal (or otherwise periodic) weak electrical current through the scalp to generate intracranial electric fields that rhythmically bias membrane polarization. The core hypothesis is frequency- and phase-specific control: if WM computations depend on oscillatory timing, classically frontoparietal theta coordination, nested theta–gamma phase–amplitude coupling, and phase-dependent gating, then externally sculpting phase relations should shift WM performance by stabilizing (or destabilizing) those timing regimes.10,26,27 Two mechanistic routes are typically invoked, and they are not mutually exclusive: (i) entrainment/resonance, where stimulation nudges ongoing oscillators toward a targeted frequency/phase; and (ii) plasticity-by-timing, where repeated phase-biased spiking and synaptic integration induce longer-lasting changes consistent with spike-timing–dependent plasticity (STDP).
Mechanistic Granularity: Phase is the Control Variable, Not Excitability
Unlike tDCS, which is often framed as tonic gain-biasing, tACS is best understood as temporal control: it perturbs when neurons are most excitable, thereby biasing spike timing, effective connectivity, and cross-area synchrony. The strongest causal story for WM is therefore not “tACS increases activity,” but “tACS changes coordination,” especially frontoparietal theta phase alignment and theta–gamma coupling that can function as a multiplexing scheme for maintenance/manipulation.26-29 A crucial refinement is to treat frequency and phase as task-conditional parameters. Even under controlled laboratory conditions, the same nominal stimulation (eg, ~6 Hz frontal theta) can be pro- or anti-performance depending on whether it aligns with (a) the individual’s endogenous peak frequency, (b) the task epoch (encoding vs maintenance vs updating), and (c) the intended inter-regional relationship (synchronization vs desynchronization).10,26,27
Evidence in Healthy Adults: Early Optimism, Then Effect-Size Shrinkage Under Stricter Designs
The empirical literature in healthy adults has been volatile, partly because early datasets were small and protocols heterogeneous, and partly because more recent work has emphasized preregistration, stricter blinding controls, and more conservative outcome modeling. A systematic review/meta-analysis reported a moderate overall effect of active tACS versus sham on WM in healthy adults, while also highlighting strong moderators (task, load, region, and session structure). 14 In contrast, a later meta-analysis incorporating newer trials concluded that current protocols yield only small improvements (and in some contexts “barely” enhance WM), consistent with a trend toward diminished pooled effects as additional null/near-null studies accumulate. 30 Overall, current evidence supports small average effects with potentially meaningful condition-specific gains.
Phase- and Network-Engineering Approaches: Synchrony as a Causal Lever
A particularly informative class of studies treats tACS as a network phase controller rather than a local modulator. Classic causal work demonstrated that externally induced frontoparietal synchronization can enhance WM performance, supporting the claim that coupling is not merely correlative but behaviorally consequential.28,29 More direct phase-geometry tests now exist: for example, an experiment comparing in-phase versus anti-phase theta tACS across frontal and parietal sites reported phase-dependent WM differences consistent with a synchrony-based causal mechanism. 31 Separately, multi-node, network-level protocols that impose non-zero phase lags between large-scale networks and assess WM during concurrent neuroimaging further reinforce a key point for narrative reviews: the same carrier frequency can help or harm depending on phase geometry and baseline subgroup (“fast” vs “slow” performers), even when stimulation intensity is held constant. 32
Older Adults and Aging: Cross-Frequency Coupling as a Plausible “Best Bet,” with Load-Dependent Gains
Aging shifts WM bottlenecks toward noisier coordination, weaker neuromodulatory gain control, and less reliable cross-frequency coupling, which makes theta–gamma coupling protocols conceptually attractive: they attempt to restore a multiplexed timing code rather than simply elevate drive.26,27 A randomized, sham-controlled, triple-blinded multi-session study in older adults applied repeated peak-coupled theta–gamma tACS over DLPFC during n-back training (16 sessions) and reported improvements that were selective to higher load (eg, 2-back sensitivity) alongside decision-bias changes and supporting a “coupling restoration + training signal” synergy model, rather than a uniform enhancement claim.33,34 Even when behavioral effects are modest or endpoint-specific, this line of work is mechanistically valuable because it operationalizes “WM enhancement” as target engagement of coupling and load-conditional benefit, not just post-hoc performance shifts.
Clinical and Translational Status: Promising Niches, But Still Thin Ice
In psychiatric and neurocognitive disorders, tACS is appealing because many syndromes show oscillatory abnormalities (theta control deficits, altered gamma, dyscoupling). However, current clinical syntheses emphasize feasibility/tolerability more than decisive cognitive efficacy, largely because trials remain underpowered and parameter dispersion is high (frequency, montage, dose, timing, and endpoints vary widely).35,36 In parallel, guidance-oriented papers on clinical tACS primarily function as conservative boundary setters, helpful for safety, governance, and responsible protocol design, rather than as evidence that WM enhancement is robust across disorders.37,38
Interpretation of tACS–WM effects is constrained by variability in effective cortical dose and sensory confounds that can compromise blinding; task selection strongly affects interpretability. Within these limits, tACS appears most promising when stimulation is aligned to endogenous rhythms and explicit phase relationships.
Transcranial Random Noise Stimulation (tRNS)
Conceptual Rationale: Why “Noise” Can be Computationally Useful
Transcranial random noise stimulation (tRNS) is a noninvasive electrical neuromodulation method in which a zero-mean, randomly fluctuating current is applied through scalp electrodes, typically spanning a broadband frequency spectrum (often 0.1-640 Hz) or a high-frequency band (commonly 100-640 Hz).15,39-44 Unlike tDCS (constant bias) or tACS (narrowband rhythmic forcing), tRNS is best conceptualized as a stochastic perturbation: it injects controlled electrical variability into cortical tissue to modulate gain, threshold dynamics, and plasticity susceptibility in task-relevant networks. The mechanistic intuition is not that noise “adds information,” but that, under specific nonlinear regimes, noise can improve the reliability of signal transmission by pushing near-threshold neuronal populations across effective activation boundaries, a principle formalized in stochastic resonance and related equivalent-noise frameworks. 45 Because WM depends on regime properties such as the stability of recurrent activity, gating thresholds, and frontoparietal coordination, it is a plausible target for noise-based modulation, with outcomes shaped by state dependence and individual heterogeneity.46,47
Biophysical and Cellular Hypotheses: Stochastic Resonance, Sodium-Channel Dynamics, and Intensity Nonlinearity
At the tissue level, tRNS induces fluctuating intracranial electric fields that bias transmembrane potentials subthresholdly, altering recruitment probability rather than directly triggering action potentials. Two mechanistic hypothesis families recur. First, stochastic-resonance and equivalent-noise accounts: in systems operating near a threshold, an optimal noise level can enhance sensitivity to weak inputs by increasing the probability that informative fluctuations cross decision or activation boundaries. Although this account is most strongly supported in perceptual paradigms, it provides a principled lens for WM: if WM failures reflect intermittent drops below a stability threshold (eg, lapses in maintenance or gating), injected noise could, in the proper regime, reduce lapse probability by assisting effective threshold crossing during encoding or updating. Second, fast channel and excitability dynamics: high-frequency tRNS has repeatedly been associated with increased cortical excitability and after-effects persisting beyond stimulation in motor-cortex assays, suggesting a plausible physiological substrate for plasticity-relevant modulation.15,39-45,48,49 Importantly, effects are not expected to be linear in “dose.” As with other tES modalities, intensity and baseline state can produce direction-like reversals because inhibitory and excitatory elements may be differentially recruited as stimulation approaches functional thresholds. 50 This matters for WM because “more excitability” is not synonymous with “better control”: excessive gain can destabilize selective maintenance, increase distractibility, or bias speed–accuracy trade-offs. Device current (1-2 mA) is not the brain dose; the effective variable is the induced cortical field, which varies with anatomy and montage.
Parameter Space: Why Spectrum (Not Just Amplitude) Matters
tRNS is unusually parameter-sensitive because its defining property is spectral structure. Mechanistic evidence suggests that high-frequency bands are often more effective for increasing excitability than lower-frequency noise, and that excitability enhancement may require a sufficiently wide bandwidth rather than a narrow sub-band. 44 Two implications follow for WM studies. First, trials labeled “tRNS” but implemented with meaningfully different spectra may not be comparable. Second, null findings can reflect spectral mismatch (or insufficient effective bandwidth) rather than a true absence of effect. For a mechanistic narrative review, spectrum should therefore be reported with the same seriousness typically reserved for tACS frequency: band limits, bandwidth, sampling characteristics, and whether the noise is full-spectrum versus high-frequency-limited should be treated as first-order protocol descriptors.15,39,48,49,51
Working-Memory Outcomes in Healthy Adults: Small Means, Strong Moderation
In healthy adults, the tRNS–WM literature remains relatively limited, but comparative studies suggest that DLPFC-targeted tRNS can, in some settings, outperform tDCS and sham, with the clearest signal observed in individuals with lower baseline WM and findings broadly consistent with threshold-sensitive and ceiling-related interpretations.46,47 Other well-controlled studies, however, report weak or null effects under alternative conditions.15,39,41-44,46-49 Overall, the evidence supports a context-dependent effect rather than a uniform enhancement claim.
Clinical and Translational Evidence: Promising Niches, Insufficient Generalization
tRNS is clinically attractive because it is typically well tolerated and can improve blinding relative to modalities that produce stronger cutaneous sensations.15,39,49 However, evidence for WM remediation remains preliminary and disorder-dependent. In ADHD, controlled work combining tRNS with cognitive training reports symptomatic improvements and physiological signatures, while executive-function outcomes including WM remain mixed or endpoint-dependent and consistent with the possibility that shifting arousal/gain control can move symptom scales without guaranteeing task-specific WM transfer. 52 In neurodegenerative and basal-ganglia contexts, pilot work in Parkinson’s disease with mild cognitive impairment suggests feasibility and modulation of attention/executive outcomes, motivating larger mechanistic trials; WM-specific claims should nevertheless remain conservative until adequately powered replication clarifies which WM subcomponents (updating, interference control, load-dependent capacity) are affected. 53 More broadly, recent systematic reviews surveying tRNS across psychiatric indications emphasize safety but also substantial protocol dispersion and small samples, limiting inference for cognition-first targets. 49
Interpretation remains limited by dose variability, regime dependence, and endpoint heterogeneity. Consistent benefits appear most likely in lower-baseline individuals and training-coupled protocols.
Transcranial Magnetic Stimulation (TMS)
Why TMS is Conceptually Different from tES
Transcranial magnetic stimulation (TMS) perturbs cortical tissue by inducing brief, high-amplitude electric fields via time-varying magnetic pulses. Unlike tES, where induced fields are weak and predominantly subthreshold, TMS can directly evoke neuronal firing, with primary effects thought to arise from stimulation of axonal elements and transsynaptic recruitment within superficial cortical gyri. This distinction matters for WM because TMS supports 2 partially distinct scientific uses: it can function as an online, “lesion-like” causal probe (single-pulse or time-locked stimulation designed to disrupt a computation), and it can operate as a plasticity-shaping intervention (repetitive protocols intended to induce durable changes in excitability, coupling, and network regime). Interpretation of cognition-facing protocols, particularly repeated-session stimulation over the prefrontal cortex, should be anchored in contemporary international safety and dosing guidance, which remains foundational for both experimental design and clinical translation.9,54
Protocol Families and What They Are Expected to Modulate
In practice, commonly used rTMS protocols can be grouped into families that carry different mechanistic expectations, although none should be treated as deterministic. Conventional frequency-coded rTMS uses low-frequency stimulation (around 1 Hz) that is often associated with suppressive network effects, and higher frequencies (approximately 5-20 Hz) that are often associated with facilitatory effects; these labels, however, function best as statistical tendencies moderated by baseline state, intensity, target, coil orientation, and concurrent task context. Pattern-coded theta burst stimulation (TBS) compresses stimulation into short trains that are typically interpreted as plasticity-biased perturbations, with intermittent TBS (iTBS) often framed as plasticity-enhancing and continuous TBS (cTBS) often framed as suppressive, again with strong dependence on dose, state, and cortical physiology. Accelerated, high-dose, and individualized stimulation paradigms are increasingly replacing one-size-fits-all approaches, enabling controllable interventions with explicit dosing logic and target verification.13,55
Mechanistic Bridge to Working Memory: Plasticity and Control-State Stabilization
WM depends on distributed frontoparietal control circuits that coordinate maintenance, updating/gating, manipulation, and interference control. 1 The most defensible mechanistic framing for rTMS/TBS effects on WM is therefore not “local excitation of DLPFC,” but state-dependent modulation of network gain and effective connectivity. Under this view, repetitive stimulation can reshape prefrontal–parietal coupling, bias oscillatory regimes linked to cognitive control (including theta-range coordination and cross-frequency organization, depending on protocol and measurement), and alter the stability of task-relevant, attractor-like control states that support WM under load. This also predicts bidirectional outcomes when timing, target, and task demands are mismatched.55-58
Healthy Adults: Small Mean Effects, Large Conditional Effects
Across healthy cohorts, meta-analytic evidence indicates small average cognitive gains (Hedges’ g ≈ 0.1) alongside substantial heterogeneity across tasks and protocols.13,55 Online stimulation effects further depend on timing and frequency, with both facilitatory and disruptive outcomes observed. Consequently, TMS is best positioned as a causal probe of working memory circuitry, while enhancement effects remain variable and context-dependent. Contemporary designs therefore emphasize target engagement and mechanistic validation (eg, TMS–EEG, fMRI connectivity) rather than relying solely on mean behavioral improvement.59,60
Older Adults and Neurodegenerative Contexts: Promise, But Endpoint- and Stage-Dependent
In aging and neurodegenerative contexts, reduced baseline network efficiency creates a larger window for modulation. Quantitative syntheses indicate that rTMS can yield cognitive benefits, but effects are endpoint- and stage-dependent, with heterogeneity driven by target selection, dose, and baseline severity. 61 For working memory, the most defensible synthesis is that benefits are subset-specific and emerge when stimulation is adequately dosed, aligned to the dominant frontoparietal bottleneck, and integrated with task engagement rather than delivered as a stand-alone intervention.
Major Depressive Disorder (MDD): Cognition is Clinically Central But Under-Standardized
TMS is now a mature clinical tool in depression, and consensus guidance emphasizes protocol optimization, safety, and individualized targeting strategies in routine care.62,63 However, WM improvement is not guaranteed. Apparent cognitive gains may arise indirectly via improvements in mood, sleep, psychomotor slowing, or motivation, or may reflect more direct reconfiguration of prefrontal control circuitry, depending on protocol and patient phenotype. A systematic review evaluating neurocognitive effects of TBS in treatment-resistant depression suggests that TBS may improve domains including attention, executive function, and WM, but the evidence remains constrained by modest sample sizes, heterogeneous cognitive batteries, and limited mechanistic mediation testing. 64 The most defensible near-term model is synergy: TMS/TBS modulates control states and plasticity, with cognitive benefits emerging when paired with structured training and evaluated using harmonized WM measures.
Schizophrenia: Early Signals, Inconsistent RCT-Level Efficacy, and a Targeting Problem
WM deficits in schizophrenia are profound and functionally central, making neuromodulation attractive, yet evidence remains mixed when WM is treated as a primary outcome. A meta-analysis focused on rTMS for WM deficits in schizophrenia reported that active and sham groups often show comparable post-treatment WM changes in available randomized trials, underscoring that the clinical evidence base remains preliminary for WM-specific efficacy claims. 65 Higher-level syntheses (including umbrella reviews) suggest that some meta-analytic signals for WM improvement may exist, but overall evidence quality is limited and protocol heterogeneity remains a major constraint. 66 Mechanistically, neuroimaging RCT work highlights a critical nuance: rTMS can show measurable target engagement at the neural level without a clean behavioral WM gain. For example, a randomized trial of prefrontal rTMS in schizophrenia reported reductions in prefrontal BOLD signal variability and an interpretable physiological change, yet these neural shifts did not map straightforwardly onto improvements in WM performance.67,68 This type of dissociation strengthens the case for biomarker-guided designs and mediation logic, and it also points to a likely target-selection problem: WM outcomes may depend on computationally appropriate hub choice (prefrontal vs parietal vs network-level targets), adequate dose, concurrent cognitive engagement, and stratification by baseline circuit integrity and symptom–cognition biotypes.
Interpretation of TMS effects on working memory is constrained by uncertainty in effective prefrontal dosing, strong state dependence, and heterogeneity in WM outcome constructs.
Deep Brain Stimulation (DBS)
Clinical Context and Why DBS Matters for Working Memory
Deep brain stimulation (DBS) is an invasive neuromodulation modality in which chronically implanted electrodes deliver patterned electrical pulses to deep gray-matter targets and/or adjacent white-matter pathways, thereby modulating distributed cortico–subcortical circuits.11,69,70 Clinically, DBS is established for movement disorders, most prominently Parkinson’s disease (PD), where stimulation of the subthalamic nucleus (STN) or globus pallidus interna (GPi) can provide durable motor benefit in appropriately selected patients.71-73 From a cognitive-systems perspective, DBS offers powerful causal leverage over basal ganglia–thalamo–cortical “gating” architecture: deep targets sit at control bottlenecks that regulate when cortical representations are stabilized, updated, suppressed, or released. For WM, the central value of DBS is therefore not a generic “enhancement” narrative, but mechanistic inference: it reveals how WM depends on subcortical gatekeeping, oscillatory regime control, effective connectivity, and neuromodulatory context and why deep stimulation often yields selective cognitive trade-offs rather than global cognitive shifts.11,69,70,74,75
Mechanisms: From Local Stimulation to Network Reconfiguration
At conventional clinical parameters (eg, high-frequency stimulation commonly used in PD), DBS should not be conceptualized as a simple on/off switch for a nucleus. Contemporary biophysics and systems neuroscience support a composite mechanism in which stimulation (i) disrupts pathological local firing patterns, (ii) preferentially recruits axonal elements (including fibers of passage) that propagate effects downstream, and (iii) reshapes large-scale oscillatory regimes and effective connectivity across cortico–basal ganglia–thalamo–cortical loops. This network-level framing matters for WM interpretation: cognitive effects are rarely the result of “stimulating cognition,” but instead of shifting the operating point of control circuitry.11,69,70 Consequently, outcomes are strongly contingent on stimulation field geometry, electrode location relative to functional subterritories, laterality, and pre-existing circuit reserve.16,76-79 These considerations support a predictive view: cognitive effects are probabilistic functions of anatomy, connectivity, baseline cognition, and stimulation policy.
Canonical Targets and Cognitive Trade-Offs
STN (Subthalamic Nucleus): Control Gating with a Robust Verbal-Fluency Vulnerability
STN is embedded in frontal–basal ganglia loops implicated in inhibitory control, switching, and conflict regulation. It is therefore unsurprising that STN-DBS can improve motor dynamics while perturbing cognitive micro-domains that rely on the same gating architecture. Across syntheses, the most consistently observed cognitive signal is a selective vulnerability in verbal fluency (phonemic and/or semantic), whereas WM outcomes are more variable and task-dependent. Importantly, this variability is mechanistically interpretable: small differences in electrode placement, field spread, and individual network integrity can shift whether stimulation stabilizes an adaptive control regime or destabilizes it, especially for tasks that require flexible updating under interference.16,76-79
GPi (Globus Pallidus Interna): Often “Cognitively Gentler,” But Not Categorically
GPi-DBS is frequently described as having fewer cognitive side effects than STN-DBS, plausibly because programming can be more motor-focused depending on anatomy and clinical strategy. However, the STN–GPi cognitive separation is not absolute. Fluency vulnerability and executive-control shifts can appear across targets, and WM differences are not uniformly reproducible across cohorts. The clinically useful inference is therefore conditional: GPi may be preferred when cognitive preservation is prioritized, but target choice should be embedded in individualized risk estimation rather than relied upon as a categorical rule.16,72,77,78,80-82
NBM (Nucleus Basalis of Meynert): Cholinergic Leverage in Dementia Contexts
The nucleus basalis of Meynert (NBM) provides major cholinergic innervation to the cortex, making it a mechanistically attractive target given acetylcholine’s role in attention, cortical gain control, and plasticity.83,84 Current clinical work in Alzheimer’s disease (AD) and related dementias remains largely feasibility- and target-engagement–oriented. Reported signals, when present, tend to cluster around attention/arousal, neuropsychiatric domains, and sleep-related outcomes more than consistent, domain-specific WM enhancement. A nontrivial complexity is state interaction: increasing cholinergic drive may be beneficial for attentional stability in some regimes yet counterproductive if it disrupts sleep architecture or consolidation-relevant dynamics.85-87
Fornix DBS: Hippocampal Network Engagement with Heterogeneous Clinical Payoff
Fornix DBS aims to modulate hippocampal–diencephalic memory circuitry. Across studies, cognitive outcomes appear heterogeneous, plausibly reflecting disease stage, residual connectivity, and variance in which networks are actually engaged by a given electrode placement and stimulation field. In this context, “target choice” alone is unlikely to rescue outcomes without stratification by network reserve and stimulation policies that respect state dependence (when to stimulate, not only where). For WM specifically, the conceptual relevance of fornix DBS is indirect but important: it illustrates that stimulation of memory circuitry can yield mixed cognitive effects unless the intervention is computationally aligned with the deficit and the remaining network can still express adaptive dynamics.86,88-90
Cognitive Outcomes in Parkinson’s Disease: What is Reproducible Versus Confounded?
In PD, DBS produces robust motor benefit, while cognitive outcomes are best summarized as selective and domain-sensitive rather than globally deleterious or enhancing. When preoperative screening excludes significant baseline cognitive impairment, group-level global cognition measures are often broadly stable. By contrast, verbal fluency, particularly phonemic fluency, emerges as one of the more reproducible vulnerability signals across comparative analyses. WM outcomes are mixed: performance may appear preserved, reduced, or occasionally improved in ON–OFF comparisons, but apparent gains can reflect motor, strategy, fatigue, or speed–accuracy effects rather than true WM capacity changes. Interpretation should therefore focus on specific subcomponents (maintenance, updating, gating, interference control) and control for motor confounds.16,71-73,77,78,81,82
Dementia/Alzheimer’s Disease: Plausible Targets, Insufficient Confirmation
DBS for Alzheimer’s disease remains experimental. Although targets such as the fornix and NBM are mechanistically plausible, current evidence for reproducible cognitive benefit is limited and heterogeneous. In the near term, DBS is best regarded as a circuit-probing approach rather than a reliable means of WM enhancement.84,85,88-90
For DBS, adaptive delivery may be more mechanistically appropriate than continuous stimulation when cognitive failures are intermittent and state-dependent. At the same time, DBS remains invasive and ethically constrained for cognitive indications, particularly in dementia, where consent capacity and risk–benefit balance are critical. Clinical experience in PD further cautions against simplistic enhancement claims, as cognitive effects can trade off across domains depending on parameters and vulnerability. Accordingly, DBS is best regarded as established for motor symptoms, with uncertain and context-dependent cognitive effects.
Closed-Loop/Adaptive Neuromodulation for Working Memory
Why “Closing the Loop” is the Real Intervention Variable?
Most WM neuromodulation remains open-loop: stimulation is delivered on a fixed schedule, implicitly treating the brain as a time-invariant target. WM, however, is a metastable control process whose success depends on transient regimes, arousal, oscillatory phase, frontoparietal coupling, and neuromodulatory tone, that fluctuate over sub-second to minute scales. Under these conditions, fixed protocols average across helpful, neutral, and disruptive states, producing the familiar pattern of small mean effects and high heterogeneity. Closed-loop neuromodulation addresses this by treating efficacy as state-contingent: stimulation is useful only when delivered at the right time, in the right state, and, for rhythmic interventions, at the appropriate phase relative to endogenous activity. On this view, the critical dose variable is not stimulation duration, but the control policy linking stimulation to brain state.
Operational Definition: From Schedule to Control Policy
A minimal closed-loop architecture can be expressed as a control pipeline:
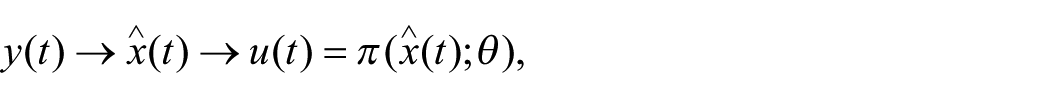
where
If slower-timescale adaptation is included, the controller parameters may be updated across trials or sessions according to:

where
Illustrative Closed-Loop/Adaptive Neuromodulation Architectures for Working Memory. This Table Summarizes Candidate Control Architectures Discussed in the Current Literature. It is Intended as a Conceptual Framework for Linking WM-Relevant Computational Objectives to Measurable State Features and Controller Policies, Rather Than as a Summary of Uniformly Validated Clinical Protocols.

What States Matter for WM?
Closed-loop success depends on choosing state variables that are (i) measurable in real time, (ii) causally coupled to computation, and (iii) sufficiently stable within-person to support policy learning. For WM, converging work motivates 4 biomarker families. First, oscillatory phase (especially frontal/frontoparietal theta phase) is attractive for phase-locked stimulation because phase can index windows of excitability and routing. Second, amplitude/power variables (theta/alpha power; gamma power; theta–gamma coupling) can serve as threshold triggers for “incipient failure” states. Third, connectivity/synchrony metrics (frontoparietal phase coherence; cross-frequency coupling; network-level integration measures) reflect the distributed nature of maintenance and updating. Fourth, computational/behavioral proxies (decision uncertainty, conflict, surprise, lapse probability, speed–accuracy regime shifts) can serve as triggers when physiology is artifact-contaminated or when the relevant state is better captured computationally than spectrally. A closed-loop protocol must therefore declare which WM subprocess it aims to stabilize, rather than treating WM as a single endpoint.3,94
Noninvasive Closed-Loop Paradigms: Early Signals, Strong Constraints
The cleanest noninvasive closed-loop demonstrations exploit phase as an explicit control variable rather than treating stimulation as a tonic intervention. Phase-tuned tACS paradigms illustrate this directly: behavioral benefit becomes contingent on the online alignment between stimulation phase and endogenous rhythms, making the policy, not the waveform alone, the experimentally meaningful object. EEG-informed TMS work converges on the same logic: the physiological and behavioral impact of identical pulses can shift substantially depending on instantaneous oscillatory state, supporting the mechanistic rationale for brain-state–gated stimulation.91,94,95 Hybrid strategies also exist: one can impose an oscillatory reference (eg, theta tACS) and then lock brief stimulation bursts to a defined phase of that reference, approximating a controllable phase geometry when real-time phase tracking is unreliable. 96
Three bottlenecks dominate noninvasive translation. The first is the artifact problem, but it differs across stimulation classes: concurrent EEG is typically most strongly contaminated during tACS because the stimulation waveform overlaps directly with oscillatory frequencies of interest; tDCS more often introduces slow drifts and offset-related distortions; and tRNS produces broadband contamination that complicates real-time spectral estimation. The second is latency: detection plus hardware/software delays can miss the targeted state entirely. The third is nonstationarity: biomarkers can change meaning with fatigue, learning stage, medication, or phenotype.97,98
Lessons from Invasive Closed-Loop Stimulation: Cognition is Stateful, and Timing Can Dominate Dose
Implantable systems provide the strongest proof-of-principle that state-dependent stimulation can shift cognitive outcomes by intercepting pathological excursions rather than “raising cognition” globally. Proof-of-principle work in other domains further supports this logic: in rodent seizure models, closed-loop stimulation triggered by pathological dynamics can limit network progression and reduce subsequent memory impairment, while human intracranial studies show that state-contingent stimulation can rescue functional networks, improve memory performance, and strengthen hippocampal network connectivity.98-101 Human intracranial studies further suggest that decoded brain-state stimulation can improve memory outcomes and that individual connectivity profiles help predict responders.99-101 The WM-relevant inference is sharp: if WM failures often manifest as intermittent regime collapses (lapses, intrusion states, runaway updating, maladaptive gating), then closed-loop policies become the coherent control strategy, whereas continuous stimulation risks either inefficacy (wrong states) or destabilization (disrupting already-optimal regimes).
Clinical Translation: Closed-Loop as Precision Stabilization, Not Amplification
Closed-loop WM neuromodulation is best framed as precision stabilization: preventing maladaptive state transitions rather than adding capacity. A credible translational program, therefore, proceeds by (i) specifying the failure mode (eg, lapse states under high load in aging; distractor susceptibility in ADHD; integration failures in schizophrenia), (ii) selecting within-subject biomarkers that track that failure mode, (iii) defining an explicit objective
Closed-loop and adaptive neuromodulation may provide the most mechanistically coherent route to more reproducible WM effects when failures are intermittent and state-dependent. The aim is not to “boost” WM in the abstract, but to deliver state-contingent control that stabilizes the appropriate network regime at the appropriate epoch, ideally with preregistered mediation pathways linking physiology, computation, and behavior.
Mechanistic Integration: Gain-and-Timing Control Over Working-Memory Trajectories
WM is not a unitary process that can be uniformly amplified by stimulation. Rather, it is a controlled dynamical trajectory whose stability, updating, and routing must be regulated across time: a time-evolving representational state that must (i) resist interference long enough to be useful, (ii) update when task demands change, and (iii) coordinate routing across distributed circuitry. Across tDCS, TMS, tACS, tRNS, and DBS, a mechanistically coherent synthesis is that neuromodulation primarily perturbs control parameters of WM dynamics, effective gain and timing, rather than directly injecting “more memory” (Figure 1). This framing explains the field’s characteristic signature: small mean behavioral effects, but potentially meaningful conditional effects (specific tasks, states, and individuals), because the sign of the perturbation depends on where a subject sits in a stability–flexibility landscape. By the stability–flexibility landscape, we mean the trade-off space in which stronger recurrent stabilization improves maintenance but can impair rapid updating, whereas weaker stabilization facilitates updating at the cost of representational fragility. Figure 2 provides an integrative overview of the framework developed here, linking major neuromodulation modalities to gain, timing, and adaptive-stability control axes, their effects on core working-memory subprocesses, and the broader roadmap toward biomarker-guided and closed-loop precision neuromodulation.

Gain–timing–stability control axes for working-memory neuromodulation. Three complementary control dimensions: gain (excitability/plasticity bias), timing (phase/entrainment windows), and stability (state-triggered control), converge on core working-memory subprocesses (maintenance, updating, gating, and interference control), providing a compact control-theoretic map linking stimulation design to cognitive function.
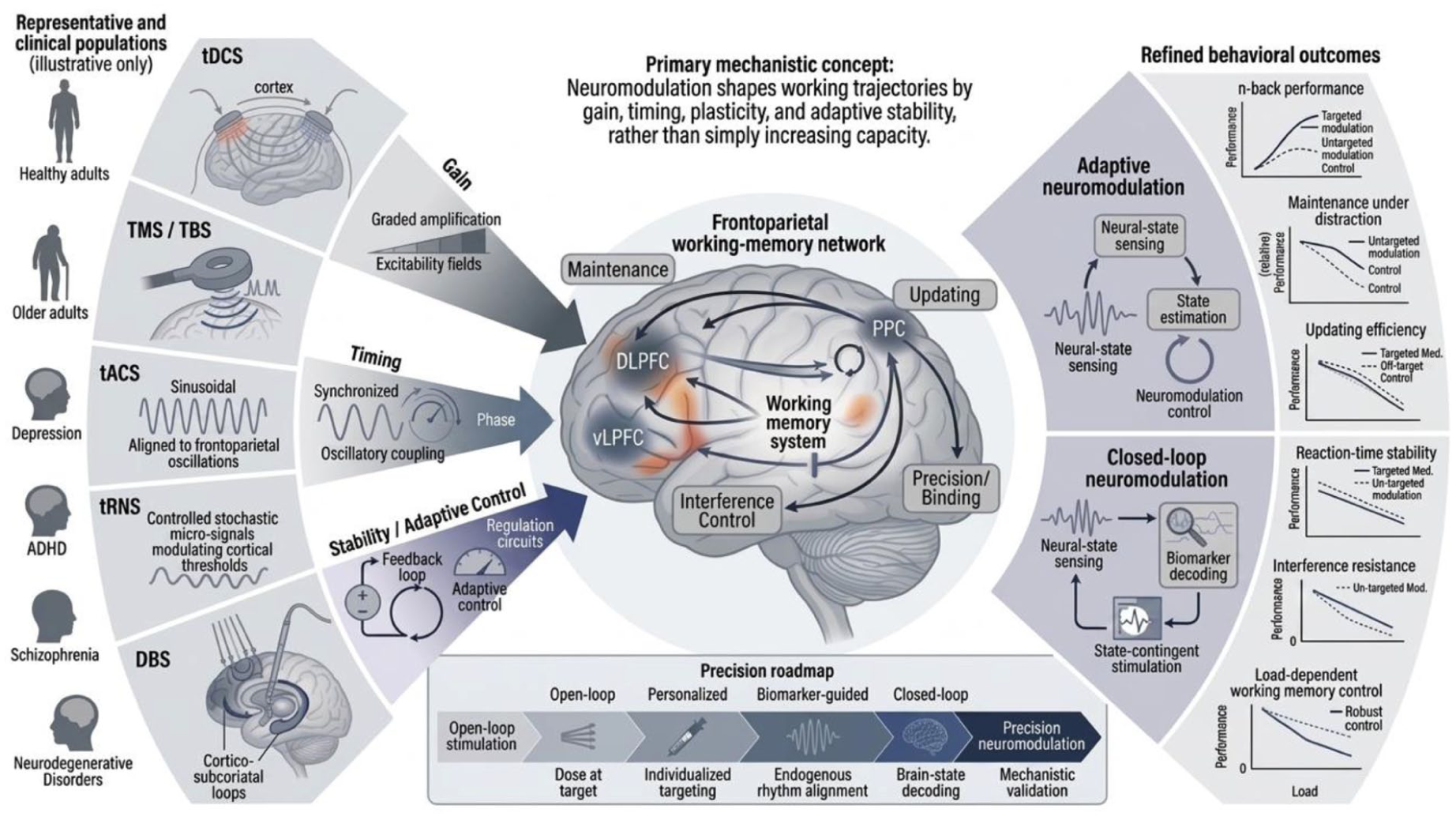
Mechanistic control architecture of neuromodulation for working memory. Across stimulation modalities, effects on working memory are best interpreted as perturbations of 3 partially dissociable control axes: gain, timing, and adaptive stability, rather than as uniform increases in mnemonic capacity. These control dimensions converge on core frontoparietal working-memory subprocesses, including maintenance, updating, interference control, and precision binding, and map onto progressively more precise intervention strategies ranging from open-loop stimulation to biomarker-guided and closed-loop neuromodulation. The figure summarizes the central claim of this review: reproducible working-memory effects depend on aligning modality, target, dose, and control policy with the specific computation being stabilized.
Plasticity Substrate: NMDA/STDP as Write Rules (Metaplastic Control)
Durable stimulation effects are best interpreted as modulation of synaptic update rules rather than instantaneous performance boosts. For tDCS, pharmacological evidence shows that after-effects can be gated by NMDA-receptor–dependent processes, consistent with a plasticity-permissive shift rather than a direct “drive” mechanism. 103 For patterned TMS, theta-burst stimulation (TBS) provides a canonical example: brief patterned trains can induce LTP/LTD-like changes in human cortex that outlast stimulation, again aligning with NMDA/Ca2+-linked consolidation logic. 55
A second refinement is temporal specificity: spike-timing–dependent plasticity (STDP) formalizes that synaptic change depends on millisecond-scale pre/post timing.104,105 When neuromodulation biases spike timing relative to endogenous rhythms, it can, at least in principle, bias plasticity phase-dependently, creating a bridge from oscillatory timing control (theta/alpha scaffolds; burst events) to longer-term circuit reweighting (Figure 1).
E/I Balance as Gain Control: The Stability–Flexibility Trade-Off
At the circuit scale, WM is constrained by a canonical compromise: stability (robust maintenance) versus flexibility (rapid updating). A sound reduction is to treat this as gain control implemented by excitation–inhibition (E/I) balance in recurrent frontoparietal circuitry. In attractor-style models of persistent activity, increasing effective recurrent gain deepens attractor basins (stronger maintenance) until the system risks rigidity/perseveration; decreasing gain shallows basins (easier updating) until representations become fragile and interference-prone.106,107
Human neurochemical measurements make the plausibility of “gain shifts” concrete: tDCS has been shown to modulate cortical inhibitory/excitatory metabolite markers (notably GABA; sometimes with complementary glutamatergic changes) in a polarity- and state-sensitive manner, implying that identical device settings can land differently across individuals and contexts. 108 Mechanistically, this makes baseline E/I state a first-order moderator: the same perturbation can move 2 people in opposite behavioral directions if they start on different sides of the stability–flexibility curve.
Oscillatory Coordination: Timing Control via Phase, Bursts, and PAC
If E/I balance sets how strongly representations persist, oscillatory dynamics shape when information is routed, bound, or updated. The contemporary picture is timing-based control organized around 3 primitives.
First, phase alignment for communication: selective routing can be implemented when communicating regions align excitability windows, “communication through coherence” as a mechanistic principle.109,110
Second, burst-based control: WM-relevant rhythmic signatures often appear as transient beta/gamma bursts rather than stationary oscillations, consistent with event-based stabilization, suppression, and readout.2,111
Third, cross-frequency coupling (CFC), especially phase–amplitude coupling (PAC), can regulate when fast local computation is expressed within slower control rhythms. Recent human evidence links hippocampal theta–gamma coupling to WM load/quality and to coordination demands that involve frontal timing, providing a concrete bridge from timing structure to prioritization and selection. 112
From a neuromodulation standpoint, tACS (and phase-tuned / closed-loop variants) is best interpreted as a timing intervention: it attempts to bias phase relationships and coupling structure toward regimes that implement better routing and prioritization. Empirically, frontoparietal/network-targeted tACS can modulate connectivity and behavior in ways consistent with this timing-control claim.28,29 More recent closed-loop approaches emphasize an even stricter point: behavioral impact depends on phase-contingent delivery, not stimulation “dose” alone.113,114
Network Control: Frontoparietal Policy + Cortico–Basal Ganglia Gating
At the system level, WM is a control policy: it decides what enters WM, what remains protected, what is suppressed, and what gets updated. Frontoparietal control networks implement top-down selection and maintenance; cortico–basal ganglia–thalamo–cortical loops implement gating and update control (computationally analogous to action selection). A mechanistic anchor here is the computational basal-ganglia gating framework, which formalizes how striatal/pallidal dynamics can regulate prefrontal updating versus maintenance, producing dissociable deficits depending on which gate component is perturbed. 74
This perspective predicts what the literature repeatedly shows: effects are computation-specific. The same stimulation can improve maintenance-under-distraction yet impair rapid updating, or shift speed–accuracy policies, depending on which node and which control subroutine is perturbed. It also motivates why individualized network targeting (rather than “DLPFC by default”) is the more principled path to reproducibility.
Neurotransmitter Systems: Dopamine, Noradrenaline, Acetylcholine as Hidden State Variables
Electrical stimulation not only perturbs membrane polarization; it can also shift neuromodulatory tone, which in turn reshapes gain, plasticity thresholds, and policy selection.
Dopamine (DA) is the classic latent variable: WM and cognitive control exhibit inverted-U sensitivity to DA, with excessive or insufficient DA degrading stability or flexibility depending on receptor/state context.115,116 Importantly, causal human evidence indicates that prefrontal TMS can influence dopaminergic release in connected striatal circuitry, providing a plausible pathway by which baseline DA state (and medication) can strongly moderate cognitive outcomes.117,118
Noradrenaline (NE) regulates arousal and “adaptive gain,” shaping exploration–exploitation balance and coding precision; this implies that NE state can amplify or blunt timing- and gain-based interventions even when stimulation parameters are unchanged. 119
Acetylcholine (ACh) supports attentional gating and plasticity, making the cholinergic state a natural “regime setter” for whether WM-relevant circuits operate in encoding-friendly versus interference-resistant modes.120,121
The methodological implication is straightforward: DA, NE, and ACh function as hidden state variables that can influence where an individual lies on the gain landscape and how plasticity gates respond to the same external perturbation.
Compact Unifying Model: Gain-and-Timing Control of WM Trajectories
A minimal scaffold clarifies the intended interpretation. Let x (t) denote a low-dimensional WM state,
If neuromodulation acts through gain- and timing-related control variables, then heterogeneity is expected rather than accidental. This implies that reproducibility depends less on averaging across nominally similar protocols and more on matching intervention parameters to baseline regime, target engagement, and task-defined computational demands.
Individual Variability & Personalized Dosing
Why Does the Same Protocol Rarely Mean the Same Intervention?
A recurring reason neuromodulation effects on WM look small and inconsistent at the group level is that “the same protocol” often fails to constitute the same biological exposure. In transcranial electrical stimulation (tES; including tDCS/tACS/tRNS), the device setting (eg, 2 mA) is not a neural dose; the relevant dose is the intracranial electric field at the tissue, its magnitude, orientation, and spatial distribution, which varies systematically with skull/CSF geometry, cortical folding, montage configuration, and individual anatomy. These sources of variance can shift not only “how much” tissue is engaged but which subregions are effectively targeted, and in some cases whether the net polarization aligns with the intended neural elements. In TMS, scaling intensity to motor threshold is a pragmatic calibration, yet it does not ensure equivalent field strength at prefrontal or parietal targets because scalp-to-cortex distance, coil orientation relative to gyral anatomy, and inter-individual head geometry strongly modulate induced fields outside M1.
As a result, nominally identical protocols can produce meaningfully different target engagement, large enough to obscure small mean cognitive effects and, in some cases, to reverse the direction of network-level consequences. Dose mismatch is therefore not a nuisance variable but a mechanistically central source of heterogeneity in WM outcomes.12,17,122,123 Key moderators and personalization levers are summarized in Table 3.
Individual Variability: Moderators of Response and Candidate Personalization Levers. This Table Organizes Heterogeneity in WM Neuromodulation into testable Moderator Classes Spanning Physical Dose Variance, State Variance, and Construct/Mechanism Variance. Its Purpose is Not to Imply That Personalization is Already Established, But to Indicate Where More Explicit Stratification and Modeling May Improve Interpretability.

Dose Variability is Only Part of the Problem
Electric-field (E-field) modeling and imaging-informed analyses increasingly support a dose–response logic for select physiological readouts, and in some contexts for behavioral variability: individuals receiving stronger (or more appropriately oriented) target fields may show larger shifts in neural markers, and occasionally larger WM changes.12,17 However, dose control is necessary but not sufficient. WM is a state-dependent control process, and responsiveness also depends on arousal, fatigue, strategy, neuromodulatory tone, medication, baseline excitation–inhibition regime, and disease-related circuit integrity. Even with comparable modeled fields, these factors can dominate variance; accordingly, the causal chain is better framed as dose × state × task rather than dose alone. Controlling physical dose reduces one major source of variance, but it does not eliminate regime dependence in a metastable cognitive system (see Table 4). 124
Protocol Parameter Space and Reporting Essentials for Working Memory (WM) Neuromodulation Studies. This Table Operationalizes Reproducibility Across Neuromodulation Modalities by Specifying the Minimum Reporting Set Needed to Interpret Heterogeneous WM Outcomes. It Emphasizes the Critical Distinction Between Device Settings and Brain-Level Dose-at-Target, Treats Timing/State Dependence as a First-Order Design Variable (Not a Nuisance), and Highlights Sham/Control and Safety Reporting Elements That Determine Whether Observed WM Effects Can be Causally Attributed to Target Engagement Rather Than Expectancy or Sensory Confounds.

The WM Signature: Responders, Non-responders, and Inverse Responders
Across modalities, WM responses often fractionate into responders, non-responders, and inverse responders, that is, individuals who reliably worsen under the same nominal protocol. This partition is expected under a gain-and-timing control view of WM trajectories. A gain-increasing perturbation may stabilize maintenance in underpowered systems (low baseline capacity, noisy control, reduced plasticity), yet destabilize selective control in systems near an optimal operating point, increasing distractibility, impulsive updating, or maladaptive persistence. Moderator-focused syntheses in prefrontal stimulation reinforce that timing (online vs offline), task demands (maintenance vs updating; interference control vs speeded performance), and baseline characteristics condition outcomes far more strongly than any uniform “enhancement” claim. In this framework, inverse responders are not anomalies to be averaged away; they are evidence that the intervention is pushing the system across a stability–flexibility boundary in the wrong direction for that individual and task. 125
What Does Personalized Dosing Concretely Entail?
Personalization is best understood as a layered strategy that separates physical targeting, dynamical alignment, and computational specificity. First, physical dose targeting replaces fixed device settings with model-informed dosing, such that montage or coil parameters and intensity are selected to achieve a predefined field range at the cortical target.122,126 Parallel logic applies to TMS, where field-based dosing and target-specific calibration are increasingly advocated, particularly outside the motor cortex.12,123 Second, for timing-sensitive interventions, especially tACS, personalization should incorporate endogenous peak frequency, phase geometry, and task-epoch alignment, because rhythm parameters and physical dose are likely to interact rather than operate independently. 12 Third, phenotype targeting aligns stimulation with the computation that is impaired, such as maintenance, interference control, or rapid updating, rather than treating all WM deficits as equivalent. 125
Personalization is Now Testable, Not Merely Explanatory
A methodological inflection point is that individualized dosing can be evaluated prospectively. Rather than invoking personalization post hoc to explain nulls, recent controlled designs have explicitly tested whether individualized, model-informed stimulation improves target engagement and whether engagement mediates behavioral change. This converts personalization from a narrative justification into intervention science: the hypothesis becomes preregisterable and falsifiable (individualized targeting → verified engagement → predicted WM shift), and failure points can be localized to modeling, state estimation, or endpoint definition. 127 Even so, in several modalities these personalization strategies remain testable hypotheses rather than validated solutions.
Reporting Discipline: Personalization Without Transparency Creates a New Reproducibility Crisis
As protocols become individualized, via model-informed montages, adaptive frequency selection, and biomarker-based stratification, reproducibility depends increasingly on transparent reporting. At minimum, studies should report modeled dose at target (including assumptions and uncertainty), targeting rationale, timing relative to task epochs and measured state variables, and preregistered primary endpoints and moderation plans. Existing reporting standards for concurrent tES–fMRI and consolidated guidance frameworks for tES study reporting provide a structural baseline; for WM neuromodulation, where effects are modest and moderator-sensitive and such standards determine whether results are interpretable, comparable, and replicable rather than merely “personalized” in an opaque way.128,129
Personal variability therefore supports the case for more explicit precision control in WM neuromodulation, including clearer definition of target computation, quantification of dose at target, alignment to endogenous state, and explicit testing of engagement–physiology–behavior pathways.
Discussion: Limitations, Research Gaps, and a Roadmap Toward Precision Neuromodulation for Working Memory
Despite significant conceptual and technical advances, neuromodulation for WM remains in a transitional phase: we can perturb circuit dynamics in ways that are clearly causal, yet we have not consistently converted perturbability into reproducible, mechanistically interpretable, and clinically meaningful cognitive benefit. The core question is no longer simply whether stimulation can influence WM, but under what conditions such effects become reproducible, mechanistically interpretable, and clinically meaningful. Below, we compress the main limitations and research gaps into a smaller set of integrative themes aligned with the gain-and-timing framework developed earlier.
Variability is the Phenomenon, Not the Nuisance
Across modalities, WM outcomes show the same broad statistical pattern: small-to-modest mean effects accompanied by substantial between-study and between-person variance. Meta-analytic work in healthy young adults often finds pooled effects of prefrontal tDCS on WM that are small and sometimes compatible with zero once heterogeneity and bias are modeled,130,131 whereas older adults and selected clinical cohorts can show somewhat larger, though still modest, effects, especially with repeated sessions and training-coupled designs. 22 This pattern is consistent with the view that stimulation perturbs gain- and timing-related control variables whose behavioral impact depends on baseline regime, target engagement, and task demands. It also reinforces that pooled “WM” findings remain difficult to compare unless studies are aligned to the specific subprocess under examination, because maintenance, updating, interference control, and load-sensitive precision need not respond similarly to the same intervention.
Methodologically, heterogeneity is further amplified by parameter dispersion, underpowered samples, outcome multiplicity, selective reporting pressures, and incomplete specification of “brain dose” (field at target) relative to device settings. The immediate corrective is not simply more trials, but better epistemic infrastructure: preregistered primary endpoints and moderator hypotheses, adequately powered multisite designs, standardized reporting of dose-at-target and participant state, and routine publication of null results to reduce bias (see Table 4).128,129
From Plausible Stories to Causal Chains: Mechanism, Durability, and Transfer
The field now has several plausible mechanistic accounts, including plasticity-linked effects, E/I rebalancing, oscillatory coordination, network reconfiguration, and transmitter-system modulation, but mechanistic closure remains uncommon. Many studies assess behavior without physiology, whereas others measure physiology without testing whether it mediates behavioral change. A more mature standard is therefore explicit causal accounting: dose-at-target → physiological change → behavior, ideally with longitudinal follow-up to distinguish transient performance shifts from learning-related change.17,20,22,127,132 This is especially important because the unresolved problem is not only immediate efficacy, but also durability and transfer. Multi-session, training-coupled protocols can produce more persistent gains, yet replication remains uneven,22,132 and far transfer to everyday executive function is still limited. 133 Under a gain-and-timing framework, such limits are expected: transfer should occur primarily when real-world demands recruit the same control primitives engaged by the intervention. Future trials should therefore preregister transfer hypotheses, include ecologically meaningful endpoints, and extend follow-up duration rather than assuming laboratory gains will generalize by default.132,134
Clinical Translation: Stimulation is Rarely the Whole Intervention
Across several modalities, the most interpretable benefits appear when stimulation is embedded in a learning or rehabilitation context rather than delivered as a stand-alone enhancer.22,127 This reframes open questions: not just “what target and intensity,” but how to choose session number and spacing, how to integrate with pharmacotherapy, and what constitutes a clinically meaningful effect size for a given disorder and functional endpoint. For noninvasive modalities, safety profiles are generally reassuring under established guidelines, but systematic evidence on intensive, repeated, or home-based dosing, especially in pediatric contexts, remains comparatively thin and argues for prospective registries and harmonized adverse-event reporting.135,136 For invasive approaches like DBS, the risk–benefit and consent landscape is fundamentally different; cognitive indications demand conservative claims, robust consent frameworks, and unusually strong mechanistic justification.
Beyond Simplified Polarity and Localization Heuristics
Legacy heuristics (“anode excites/cathode inhibits,” “DLPFC stores WM”) are increasingly insufficient. Polarity effects can invert with field orientation, ongoing state, and network regime; and WM depends on distributed control policies rather than a single cortical locus. Paradoxical facilitation under inhibitory-leaning manipulations and mixed effects of parietal versus prefrontal targeting are not anomalies; they are signatures of control-parameter perturbation in a dynamical system. Progress therefore requires sharper theoretical commitments: specify which WM computation is being targeted (eg, maintenance, updating, interference control, or priority weighting), because conclusions drawn at the generic WM level may obscure subtype-specific effects. In this sense, dynamical-systems models, computational control accounts, and network/control theory are not decorative; they are the vocabulary needed to translate heterogeneous empirical effects into predictable engineering. 137
Roadmap: From Artisanal Protocols to Precision Neuromodulation
Any precision-oriented roadmap must be read against the current limitations of the evidence base: in several domains, the problem is not only insufficient personalization, but insufficient confidence in the robustness of the underlying effects.
A credible roadmap begins from a simple principle: neuromodulation should be treated as measurable, individualized control rather than recipe-driven perturbation. Personalized dosing and targeting should replace nominal device settings with brain-relevant exposure estimates, combined with baseline biomarkers and cognitive phenotyping to reduce responder/non-responder mixing.17,138 Closed-loop and state-triggered approaches address the equally important “when” problem: if WM failures are episodic and state-dependent, state-contingent delivery may improve efficacy while reducing unnecessary stimulation.100,139 In parallel, stimulation should be tested within synergistic designs that pair it with explicit learning signals and, where justified, pharmacological state-setting, but these combinations should be evaluated as mechanistic hypotheses rather than assumed advances.103,127 New technologies promising greater focality or deeper reach will matter only if they demonstrate target engagement, safety, and mediated mechanism-to-behavior links, rather than novelty alone.140,141
Ethics and Societal Impact: Enhancement is Not a Neutral Endpoint
As devices become cheaper, wearable, and home-deployable, ethics becomes inseparable from method. Equity of access, privacy of neurodata, informed consent (especially in children and cognitively impaired patients), and the risk of coercive or competitive “enhancement” environments require proactive governance. Neuroethics roadmaps emphasize transparency, risk communication, and safeguards against unsupervised or coercive use. 142 A precision paradigm also sharpens ethical duties: if we can predict subgroups likely to worsen under a protocol, deploying generic stimulation without screening becomes increasingly difficult to justify.
Taken together, the field will mature when WM neuromodulation becomes a controllable intervention rather than a probabilistic perturbation. The gain-and-timing framework offers both an explanation for heterogeneity and a blueprint for progress: quantify dose, model baseline regime, target the correct control node, intervene at the correct time, and evaluate outcomes that match the computation being stabilized. If pursued with open-science discipline and mechanistic restraint, neuromodulation can move from intriguing, selective effects toward reliable, personalized cognitive therapeutics, particularly for aging and neuropsychiatric populations where WM impairment is clinically consequential.
Conclusion
WM is a capacity-limited control system that stabilizes, protects, and updates task-relevant information. Because it declines with aging and is disrupted across neuropsychiatric and neurological disorders, neuromodulation has become an important tool for probing WM circuitry and, in some contexts, modifying performance.
Across modalities, the most defensible conclusion is not that stimulation simply “adds” working memory, but that it perturbs control variables governing WM dynamics, especially effective gain, timing, and plasticity-related state transitions. What can be concluded with greatest confidence is that neuromodulation can modulate WM-related circuitry and, under constrained conditions, alter performance in task-relevant ways. What remains less certain is the extent to which such effects generalize across heterogeneous protocols, populations, and WM endpoints, particularly for modalities with smaller or clinically narrower literatures. The practical implication is therefore methodological rather than promotional: future progress depends on explicit specification of dose at target, task-defined computation, endogenous state, and target-engagement pathways, ideally in designs that can distinguish physiological engagement from behavioral outcome.
As neuromodulation becomes more scalable, responsible translation will require clear ethical governance, including appropriate screening, informed consent, privacy protection, and safeguards against unsupervised use. Under those conditions, neuromodulation may develop into a more reliable approach for stabilizing WM-related control in vulnerable populations.
Footnotes
Acknowledgements
We acknowledge the foundational contributions of researchers whose work informed and inspired this review.
Author Contributions
SKSR: Conceptualization, Study design, Evidence synthesis, Formal analysis, Writing – original draft. MAA: Writing – review & editing, Supervision. FG, JR: Literature acquisition, Critical revision of the manuscript. AS, MR, SH, MS: Critical revision of the manuscript for important intellectual content.
Funding
The authors received no financial support for the research, authorship, and/or publication of this article.
Declaration of Conflicting Interests
The authors declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Generative AI
No generative artificial intelligence tools were used to generate scientific content or analyses in this manuscript.