
Abstract
Objective:
Recursive Informational Curvature (RIC) was recently introduced as an information-geometric framework for describing the balance between recursive structure and entropy change in dynamical systems. Here, we present the first empirical implementation and benchmark evaluation of its scalar curvature term,
Methods:
We analyzed 2 open-access datasets: a 128-channel eyes-open versus eyes-closed benchmark derived from the George anesthesia-and-sleep recording package, and a 64-channel food-tracking dataset with synchronized motion capture. Following standardized preprocessing and symbolic discretization, we extracted Shannon entropy, recursive gain, and empirical curvature for each channel and epoch. We then evaluated these features using single-feature, multichannel, and combined-feature models, together with ablation and sensitivity analyses across symbolic bin counts, epoch lengths, and classifier families.
Results:
In the primary eyes-open versus eyes-closed benchmark, multichannel curvature features supported near-perfect state discrimination. In the food-tracking task, the strongest performance was obtained from combined curvature and amplitude features. Across analyses, entropy alone was weak, whereas recursive gain and curvature showed closely matched performance profiles, indicating that in the present implementation much of the discriminative structure captured by curvature is concentrated in the recursive term.
Conclusion:
These findings establish the empirical RIC curvature term as an interpretable and state-sensitive descriptor of neural dynamics in ECoG, provide a reproducible benchmark for future refinement of the framework, and clarify both the promise and the current limits of curvature-based analysis in neural states.
Keywords
Introduction
Understanding neural dynamics requires analytical frameworks that extend beyond assumptions of linearity, stationarity, and fixed oscillatory structure. Brain activity unfolds across interacting temporal and spatial scales and is shaped by recurrent, nonlinear, and state-dependent organization that is only partially captured by conventional descriptors such as spectral power, entropy, or connectivity. Although these measures remain central to electrophysiological analysis, each isolates only 1 aspect of neural structure and may therefore miss the evolving balance between recurrent reinforcement, dispersion, and spatial reconfiguration that accompanies changes in brain state.1-3
This limitation has motivated increasing interest in geometric and information-theoretic approaches to neural dynamics. Recent work in computational neuroscience and information geometry has suggested that neural activity may be more effectively characterized in terms of state-space organization, local structure, and transition geometry rather than by amplitude or pairwise dependence alone. Such approaches are attractive because they offer a language for describing how neural systems occupy, traverse, and reorganize within structured dynamical regimes. Yet, despite this conceptual progress, the field still lacks practical and interpretable tools that can translate these ideas into benchmarkable features for real neural recordings, particularly for high-resolution electrocorticography (ECoG).4-7
Recursive Informational Curvature (RIC) was recently proposed as a geometric formulation for describing the balance between recursive structure and entropy change in dynamical systems. In broad terms, the framework is motivated by the idea that neural signals may vary not only in magnitude or irregularity, but also in the extent to which present symbolic organization remains coupled to its recent past while either stabilizing or dispersing over time. This suggests that a curvature-like quantity, defined from recursive gain and entropy change, may provide a compact descriptor of neural-state organization that is dynamic, interpretable, and anatomically mappable.8-11
The present study provides the first empirical implementation and benchmark evaluation of RIC in high-density macaque ECoG. Specifically, we operationalize the curvature term
This question is neuroscientifically important because neural states are unlikely to be defined by recurrence alone or by dispersion alone. A cortical regime may exhibit strong local dependence yet remain highly unstable, or display moderate recurrence within a comparatively stable symbolic organization. What is often missing is a descriptor of the evolving relation between these terms. The RIC formulation addresses this gap by combining recursive dependence and entropy change into a single dynamic curvature field that can be analyzed locally across channels and globally as a multichannel neural-state geometry.
We hypothesize that the empirical RIC scalar captures higher-order structure in ECoG signals that is not fully preserved by power-only or entropy-only summaries. More specifically, we test whether multichannel curvature patterns support state decoding, low-dimensional organization, and anatomically interpretable cortical mapping in ways that are stable across benchmark settings and informative relative to established descriptors. By embedding RIC within a transparent and reproducible computational pipeline, this study positions curvature-based analysis as a practical addition to the methodological toolkit for neural state characterization and benchmark neuroscience.
Recursive Informational Curvature: From Symbolic Flow to Neural State Geometry
For an intuitive overview of the Recursive Informational Curvature (RIC) framework, see Section S1 of the Supplemental Mathematical Framework. In the present study, RIC is used as a formal and empirically tractable framework for describing neural dynamics in terms of recursive structure and entropy change within a symbolic state space.6,7,12 Rather than treating brain activity exclusively through oscillatory power, pairwise connectivity, or scalar uncertainty measures, this approach asks whether neural state organization can be represented by a curvature-like quantity that tracks the local balance between delayed self-dependence and symbolic dispersion.2,13,14 Formal definitions of the latent quantities and the benchmark-specific estimator are provided in Sections S2-S3 of the Supplemental Material.
Why a Dynamic Geometric Framework is Needed
A central limitation of many conventional electrophysiological descriptors is that they isolate only 1 aspect of neural organization at a time. Spectral power captures amplitude within predefined frequency bands, entropy summarizes dispersion, and connectivity measures pairwise or network-level dependence; yet none of these quantities, on its own, explicitly represents the continuously changing balance between recurrent structure and dispersive variability that shapes transitions between neural states. For high-density ECoG in particular, the relevant object is not simply “how much activity” is present, but how local signal structure is recursively reinforced, destabilized, or redistributed across time and space. This is the problem to which the curvature formulation is addressed.6,7,15,16
The motivation for a geometric description is therefore not ornamental. It arises from the need for a state variable that can compress 2 distinct but complementary features of neural organization into a single interpretable object: recursive dependence, which captures how strongly current symbolic structure remains coupled to its recent past, and entropy change, which captures whether symbolic organization is concentrating or dispersing over time. A dynamic curvature field, denoted here by
This framing is also motivated by the empirical structure of the present benchmark. The main results show that curvature is most informative when treated as a multichannel pattern rather than as a single averaged scalar, and that its usefulness lies in providing a compact, anatomically mappable, and decoding-relevant summary of distributed neural organization. At the same time, the current data show that the empirical curvature term remains closely aligned with recursive gain under the present implementation. The role of the framework in this article is therefore not to claim a complete theory of cognition, but to test whether a curvature-based descriptor yields stable, interpretable, and state-sensitive geometry in neural recordings.
Latent Formulation of RIC
In its general form, the RIC framework represents system dynamics through a latent symbolic field
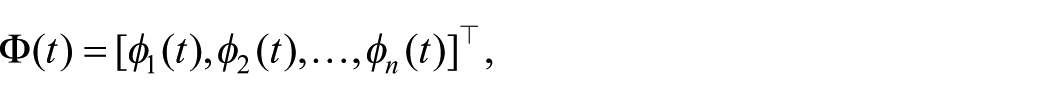
where each
The latent curvature quantity may then be written in normalized form as

where
This latent formulation serves 2 purposes in the present article. First, it provides a principled theoretical language for linking recursive dependence and entropy change within a single state variable. Second, it clarifies why a curvature quantity is a natural candidate for neural-state analysis: because it converts 2 complementary axes of organization into a signed, dynamic descriptor that can be tracked over time and across channels. Importantly, in the present study
Empirical Operationalization in ECoG
The latent symbolic field is not observed directly in the macaque datasets analyzed here. Instead, for each channel c and epoch e, the preprocessed ECoG signal
Within this benchmark, recursive gain is operationalized as lag-1 mutual information within each symbolic epoch,

and symbolic dispersion is quantified by the empirical entropy
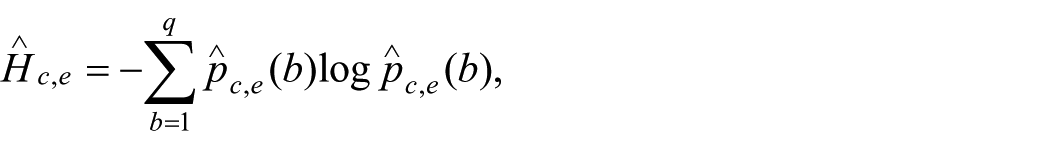
with entropy change defined across adjacent epochs as

Because recursive gain and entropy change do not naturally share a common numerical scale, both are standardized before being combined. The empirical curvature estimator used throughout this study is therefore

with
This benchmark-specific quantity is the operative RIC feature in the article. It should be interpreted as an empirical signal-level descriptor derived from the balance between recursive dependence and entropy change in symbolized neural data. Its purpose is not to adjudicate broad ontological claims, but to test whether a curvature-like quantity captures discriminative state structure that is stable enough for decoding, interpretable enough for anatomical mapping, and structured enough for low-dimensional embedding and clustering.
The Neuroscientific Role of the Curvature Field
The reason to introduce a curvature field rather than analyze recursive gain and entropy separately is neuroscientific as well as mathematical. Neural states are not defined solely by the magnitude of recurrent structure or solely by the amount of variability. They are defined by the evolving relation between the 2. A cortical regime may show strong local recurrence but also rapid symbolic dispersion, or modest recurrence coupled to highly stable symbolic structure. Treating recursive gain and entropy in isolation therefore risks missing the state-defining balance between reinforcement and redistribution. The curvature field
This balance is also why a dynamic formulation is necessary. Neural organization is not static across an entire recording; it fluctuates over short windows, reorganizes across cortical regions, and changes with behavioral context. A useful state descriptor must therefore be able to follow local reconfiguration in time while preserving spatial structure across the array. In the present implementation,
A further advantage of this formulation is that it makes explicit why scalar averaging can be insufficient. For each epoch

which represents the neural state in a high-dimensional curvature space. This distributed representation preserves spatial organization that may be lost under simple averaging. The Supplemental Mathematical Framework formalizes this point by showing that scalar averaging can attenuate distributed state differences, whereas the full multichannel curvature vector preserves the geometry relevant for classification and embedding. This is directly consistent with the empirical results of the benchmark, where multichannel curvature features were substantially more informative than scalar summaries. 12
The current data also sharpen the interpretation of what curvature contributes. Entropy alone was weak in both datasets, while recursive gain and curvature showed closely matched performance profiles. Thus, in the present formulation, much of the discriminative structure captured by curvature is concentrated in the recursive term. That result does not make the curvature field redundant. Rather, it clarifies its current role:
From Local Curvature to Neural State Geometry
A central implication of the present framework is that neural states are best treated as distributed geometric configurations rather than as isolated scalar summaries. In the benchmark analyses, each epoch is represented by its channel-wise curvature vector
Within this restricted empirical scope, the question addressed by the present study is straightforward: whether a curvature-based representation derived from recursive dependence and entropy change yields stable and state-sensitive organization in high-density ECoG. The benchmark results support that narrower claim. Multichannel curvature features supported strong decoding and coherent low-dimensional organization of neural epochs, particularly in the eyes-open versus eyes-closed benchmark, while also showing that curvature remains closely aligned with recursive gain under the current implementation. The present article should therefore be read as a first empirical benchmark of the curvature formulation, not as an exhaustive validation of the broader RIC research program (Table 1).
End-to-End Computational Pipeline for Empirical Benchmarking of the Curvature Scalar K in Macaque ECoG.
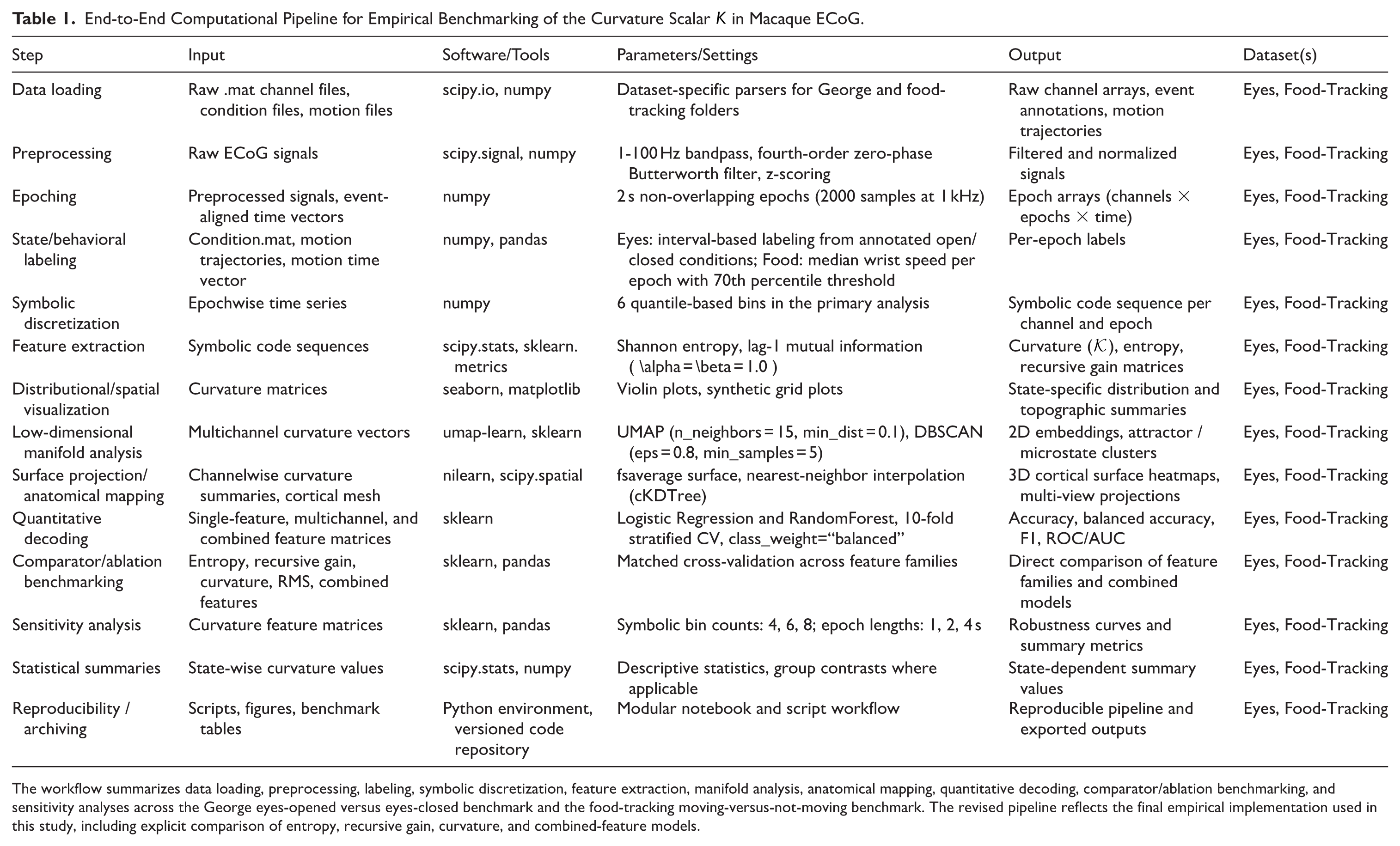
The workflow summarizes data loading, preprocessing, labeling, symbolic discretization, feature extraction, manifold analysis, anatomical mapping, quantitative decoding, comparator/ablation benchmarking, and sensitivity analyses across the George eyes-opened versus eyes-closed benchmark and the food-tracking moving-versus-not-moving benchmark. The revised pipeline reflects the final empirical implementation used in this study, including explicit comparison of entropy, recursive gain, curvature, and combined-feature models.
Formal definitions of the latent quantities, the benchmark-specific estimator, the regime-partitioning scheme, and the mathematical properties of the empirical curvature field are provided in Sections S2-S3 of the Supplemental Mathematical Framework.
Methods
Data Acquisition and Experimental Paradigms
We analyzed 2 open-access macaque ECoG datasets. The first was the George 128-channel subdural ECoG recording distributed within the NeuroTycho anesthesia-and-sleep package. From this dataset, we extracted annotated eyes-opened and eyes-closed intervals. The primary balanced benchmark used Session 1 only, which contained both awake-eyes-opened and awake-eyes-closed intervals. Recovery-state intervals from Sessions 2 and 3 were examined separately during exploratory analysis but were not used in the primary balanced decoding benchmark. The second dataset comprised 64-channel epidural ECoG recorded during a food-tracking task with synchronized 3D motion capture.
Preprocessing and Epoching
Raw ECoG signals were loaded in Python (using scipy.io and numpy) and preprocessed uniformly across both datasets. Signals were band-pass filtered between 1 and 100 Hz with a fourth-order zero-phase Butterworth filter (scipy.signal.sosfiltfilt) and z-scored to remove baseline drift and amplitude differences. For each channel, signals were segmented into non-overlapping 2-second epochs (2000 samples/epoch at 1 kHz), ensuring alignment with experimental annotations and behavioral labels. In the food-tracking dataset, epoch boundaries were synchronized with available motion-tracking data to retain only fully overlapping neural-behavioral epochs.
Behavioral and State Labeling
In the George dataset, epoch labels were assigned using the condition annotations stored in Condition.mat. The primary benchmark included only epochs falling within the annotated AwakeEyesOpened and AwakeEyesClosed intervals of Session 1. Recovery-state intervals (RecoveryEyesOpened, RecoveryEyesClosed) and anesthetized intervals were parsed separately during quality control and exploratory analyses, but anesthetized intervals were excluded from the primary open-versus-closed decoding benchmark. For the food-tracking dataset, wrist marker trajectories were extracted from motion capture data, and epoch-level movement was defined using the median wrist speed within each epoch. Epochs were labeled “moving” if median wrist speed exceeded the 70th percentile of valid epochs and “not moving” otherwise.
RIC Feature Extraction
For each channel and epoch, signals were discretized into 6 quantile-based bins. Three features were then computed: Shannon entropy, recursive gain, and the empirical curvature scalar
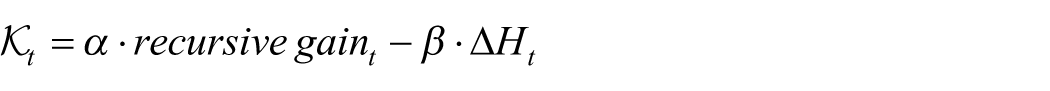
where
Visualization and Benchmarking
RIC features were visualized at several levels. Violin plots (via seaborn.violinplot) summarized curvature distributions across channels and states, while spatial grid plots displayed channel-wise means on synthetic 8 × 16 (eyes dataset) and 8 × 8 (food-tracking) arrays. For cross-epoch and cross-channel structure, all-epoch/channel RIC vectors were projected to 2D using UMAP (umap-learn), with unsupervised microstate clustering performed using DBSCAN (sklearn.cluster.DBSCAN). Microstates (attractors) were visualized in the UMAP space.
To visualize RIC patterns in anatomical context, mean and state-specific curvature values were interpolated from channel locations to the standard fsaverage brain surface (nilearn.datasets.fetch_surf_fsaverage) using nearest-neighbor mapping (scipy.spatial.cKDTree). Surface heatmaps were rendered with nilearn.plotting.plot_surf_stat_map and saved from multiple anatomical views (lateral, dorsal, medial, posterior). Difference maps were generated to highlight spatial changes in RIC between states.
Quantitative State Decoding, Comparative Benchmarking, and Sensitivity Analyses
State decoding was assessed using logistic regression and RandomForest classifiers under 10-fold stratified cross-validation. We evaluated single-feature models using mean epochwise curvature, entropy, recursive gain, or RMS amplitude; multichannel models using the full channel-wise feature vectors; and combined-feature models that concatenated curvature with amplitude or with the constituent entropy and recursive-gain terms. 18
To assess whether curvature captured information beyond its components, we performed explicit comparator and ablation analyses using entropy alone, recursive gain alone, curvature alone, and combined models. We further evaluated robustness across symbolic bin counts (4, 6, and 8 bins), epoch lengths (1, 2, and 4 seconds), and classifier choice. Performance was summarized using accuracy, balanced accuracy, F1 score, and area under the ROC curve.
CEBRA Comparison
As an additional nonlinear representation-learning comparator requested during peer review, we benchmarked the empirical curvature scalar
Separate Analyses for Each Dataset
For the eyes open/closed dataset, all analyses were performed using 128-channel arrays and state labels derived from experimental annotations. All features, visualizations, and benchmarking metrics were reported for both states and their difference.
For the food-tracking dataset, analysis used 64-channel arrays, with “moving” versus “not moving” behavioral labels. The whole pipeline, preprocessing, RIC extraction, visualization, decoding, model comparison, UMAP/microstates, and 3D brain mapping, was replicated, enabling direct comparison of spatial and informational complexity across cognitive and behavioral states.
Software and Reproducibility
All data handling, feature extraction, modeling, and visualization were implemented in a fully reproducible Python workflow using open-source libraries. The complete computational sequence, from raw ECoG loading and behavioral/state labeling to symbolic discretization, curvature extraction, manifold analysis, anatomical mapping, decoding, comparator analysis, and sensitivity testing, is summarized in Table 1. This pipeline structure provides the methodological basis for the benchmark outcomes reported in the Results, which are condensed in Table 2.
Comparative Benchmarking of the Empirical Curvature Scalar

The George analysis was restricted to Session 1 to provide a balanced open-versus-closed comparison and avoid confounding state labels with session identity. The food-tracking analysis included 518 valid motion-aligned epochs. Reported values summarize cross-validated decoding performance for single-feature, multichannel, and combined-feature models. Across both datasets, entropy alone was weak, whereas recursive gain and curvature showed closely matched performance, indicating that in the present implementation much of the discriminative structure is concentrated in the recursive component. In the food-tracking task, the strongest results were obtained when curvature was combined with RMS amplitude.
Results: RIC Features Reveal State-Dependent Complexity Across ECoG Datasets
Spatial and Distributional Patterns of RIC Curvature
We extracted RIC and related features from all channels and epochs of both the eyes open/closed and food-tracking macaque ECoG datasets (see Methods). Violin plots revealed broad, heterogeneous distributions of RIC curvature across the ECoG array in both cognitive and behavioral paradigms (Figure 1, top). In the eyes open/closed dataset, spatial grid plots and 3D projections demonstrated systematically higher RIC curvature in posterior and occipital regions during eyes closed, indicating enhanced internal dynamical organization in the absence of visual input (Figure 1). For the food-tracking dataset, mean RIC curvature displayed a distinct spatial pattern, with several anterior and central channels exhibiting state-dependent increases during voluntary movement. Notably, unsupervised UMAP embedding of RIC-derived features (Figure 1, bottom) revealed that neural states clustered into discrete, attractor-like manifolds, allowing clear separation of experimental conditions at the population level. Channel-wise mapping of mean RIC values across synthetic electrode grids further underscored the topographic specificity of curvature dynamics. As illustrated in Figure 2, posterior and parietal channels consistently showed elevated RIC in the eyes closed condition, while more heterogeneous distributions emerged during active food-tracking. Collectively, these findings demonstrate that RIC robustly encodes spatially structured, state-dependent organization across the cortical surface.
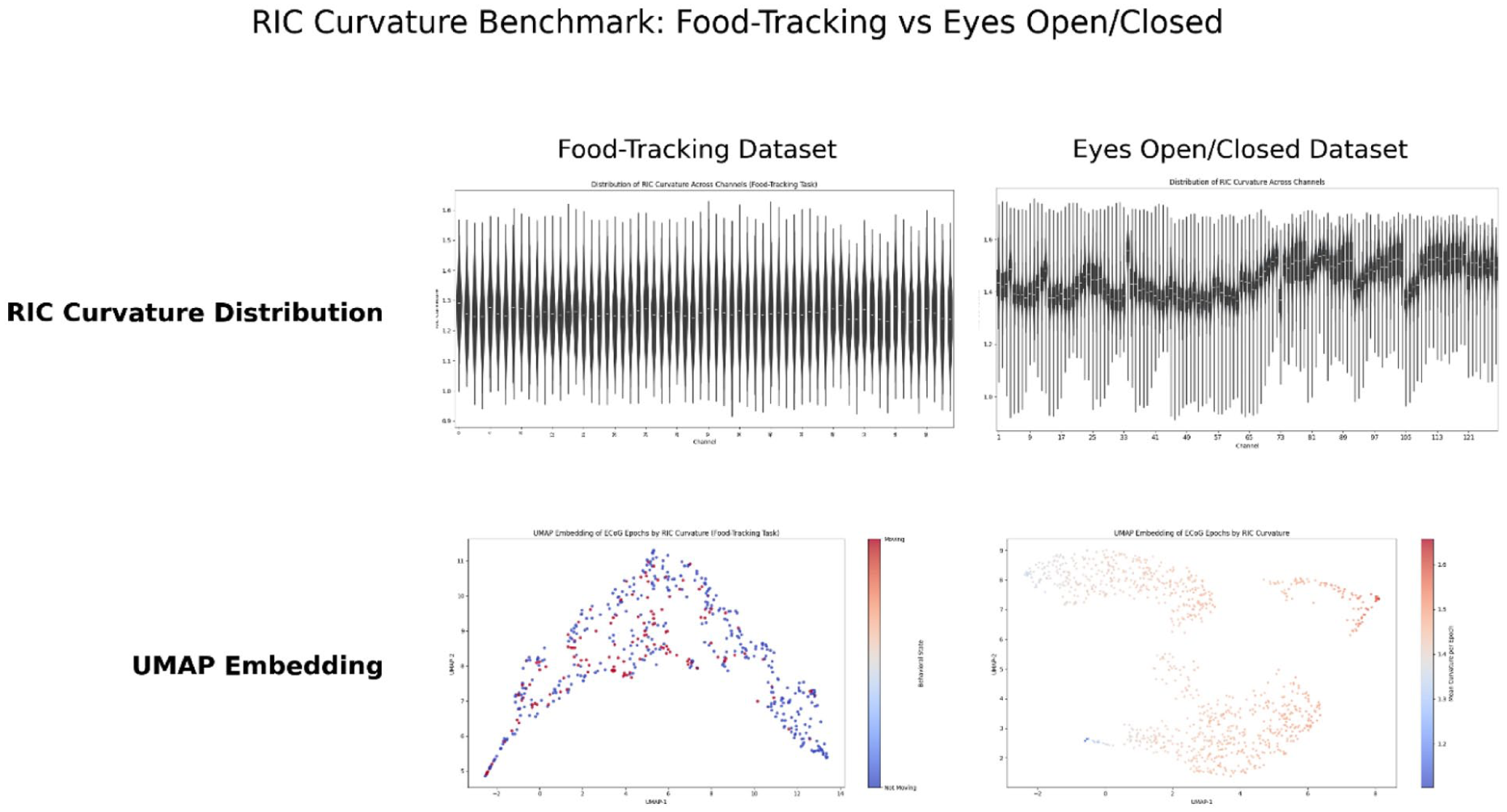
Distributional, spatial, and manifold structure of curvature across the 2 macaque ECoG benchmarks. Top panels show violin-plot distributions of curvature values across epochs for the George Session 1 eyes-opened versus eyes-closed benchmark and the food-tracking moving-versus-not-moving benchmark. Middle panels show channel-wise spatial summaries across the electrode arrays. Bottom panels show UMAP embeddings of multichannel curvature features, illustrating state-dependent low-dimensional structure in both datasets.
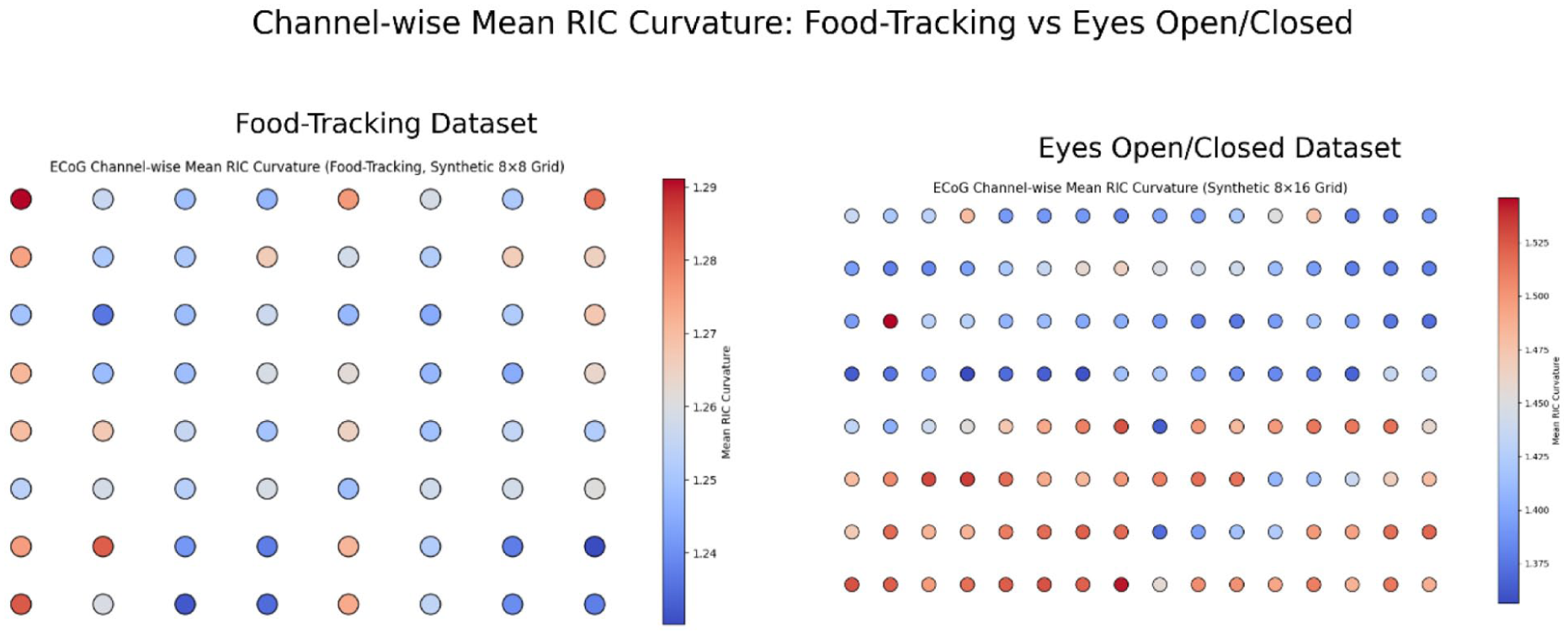
Channel-wise mean curvature across the food-tracking and eyes-opened versus eyes-closed ECoG datasets. Each circle represents the mean curvature value for one ECoG channel, displayed on synthetic electrode grids for the food-tracking dataset (left) and the George Session 1 eyes-opened versus eyes-closed dataset (right). The maps highlight distinct spatial organization of curvature across behavioral and cognitive paradigms.
RIC-Based State Decoding: Cognitive and Behavioral Benchmarks
Primary Eyes-Opened versus Eyes-Closed Benchmark
We next evaluated the ability of the empirical curvature scalar K to discriminate between eyes-opened and eyes-closed states in the primary balanced George benchmark. This analysis was restricted to Session 1, which contained both annotated awake-eyes-opened and awake-eyes-closed intervals, yielding 942 valid epochs in total, with 471 epochs per class. In single-feature models, mean curvature provided moderate above-chance discrimination, indicating that a global epochwise summary of K contains state-dependent information. However, decoding performance increased substantially when the full multichannel curvature profile was used. Multichannel models approached ceiling performance, with the strongest models achieving near-perfect separation between the 2 states.
This performance pattern indicates that the discriminative signal is not confined to a single global curvature summary but is distributed across the channelwise spatial configuration of
Food-Tracking Benchmark
We then applied the same benchmarking framework to the food-tracking dataset. After synchronization of neural and motion data, 518 valid epochs were retained, comprising 362 not-moving and 156 moving epochs. This task was more challenging than the George benchmark, as expected for unconstrained naturalistic behavior. Single-feature models based on curvature performed poorly to modestly, and entropy alone again showed weak discrimination. In contrast, multichannel curvature and recursive-gain feature sets produced moderate classification performance, indicating that behaviorally relevant information was present but more distributed and less separable than in the open-versus-closed paradigm.
The best-performing model in this dataset was obtained by combining multichannel curvature with RMS amplitude features in a logistic-regression classifier. This model achieved an accuracy of approximately 0.803, a balanced accuracy of 0.788, and an AUC of 0.887. Thus, in the food-tracking task, the most useful signal was obtained not from curvature alone but from the combination of curvature with amplitude-sensitive features. This result suggests that the curvature term captures behaviorally meaningful structure in the neural signal, but that complementary information carried by amplitude enhances classification in more heterogeneous motor contexts. Full comparator results across entropy, recursive gain, curvature, and combined-feature models are summarized in Table 2 and expanded in the robustness analyses. Taken together, these 2 benchmarks show that the empirical curvature scalar
3D Brain Mapping and Surface Heatmaps
Projection of mean RIC curvature onto the fsaverage brain surface revealed striking spatial organization across experimental conditions. In the eyes open/closed dataset, surface heatmaps and anatomical views (lateral, dorsal, medial, posterior) demonstrated a pronounced increase in curvature throughout occipital and parietal cortex during eyes closed, relative to eyes open (Figure 3). In the food-tracking paradigm, state-specific surface maps highlighted regions with movement-associated increases in RIC curvature, particularly over sensorimotor and midline areas (Figure 4). These spatial signatures were robust to electrode subsampling and channel reordering. Notably, as shown in Figures 3 and 4, eyes closed states were characterized by a posterior and medial predominance of high RIC, consistent with engagement of default mode and associative cortical networks, whereas eyes open conditions exhibited globally reduced curvature. This anatomical mapping underscores RIC’s sensitivity as an index of state-dependent cortical dynamics.
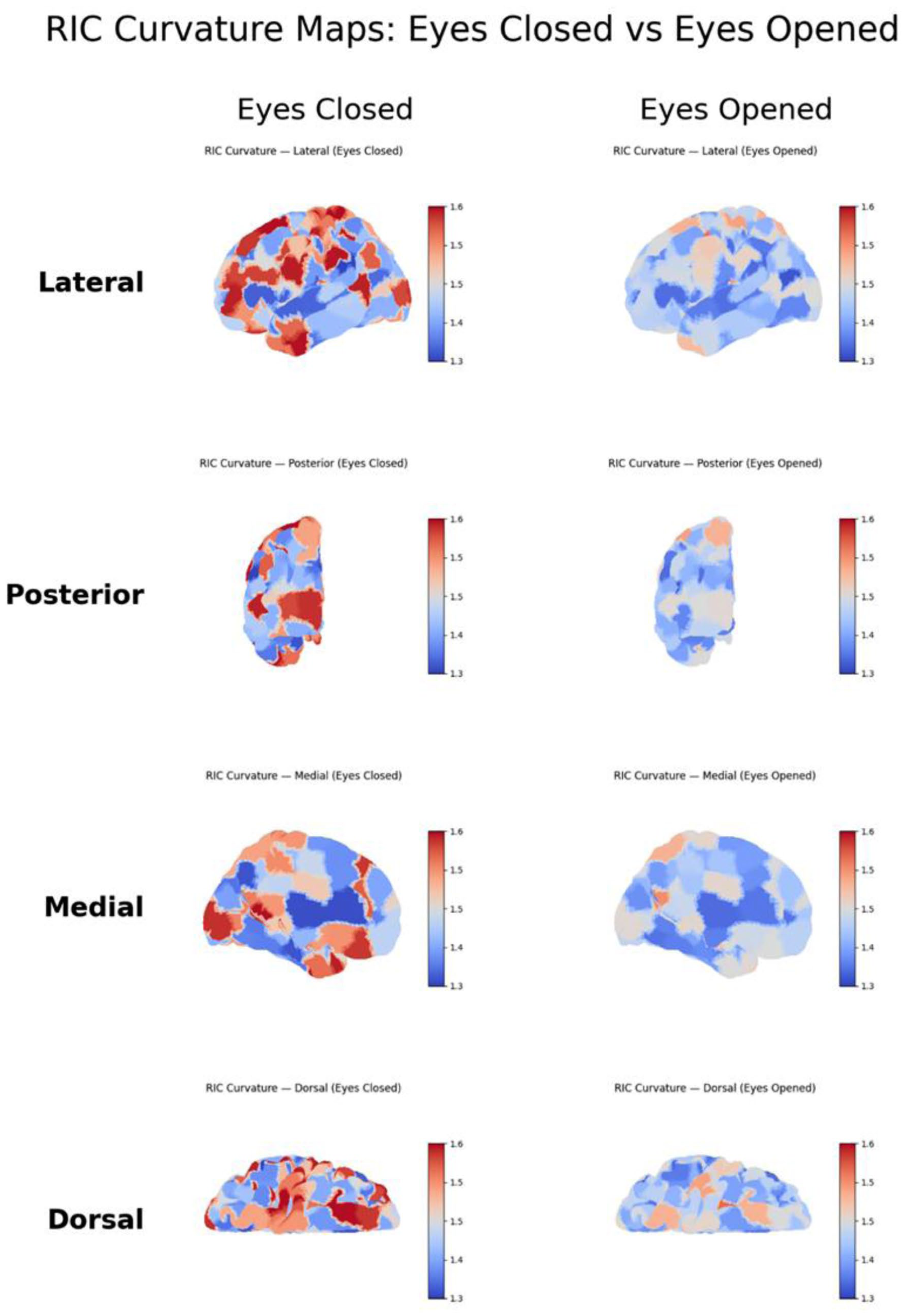
Surface maps of curvature in the George Session 1 eyes-closed and eyes-opened conditions. Mean curvature values are projected onto 4 cortical views (lateral, posterior, medial, and dorsal) for the eyes-closed and eyes-opened states. The maps show state-dependent spatial differences in cortical curvature, with stronger posterior and medial curvature patterns observed in the eyes-closed condition.
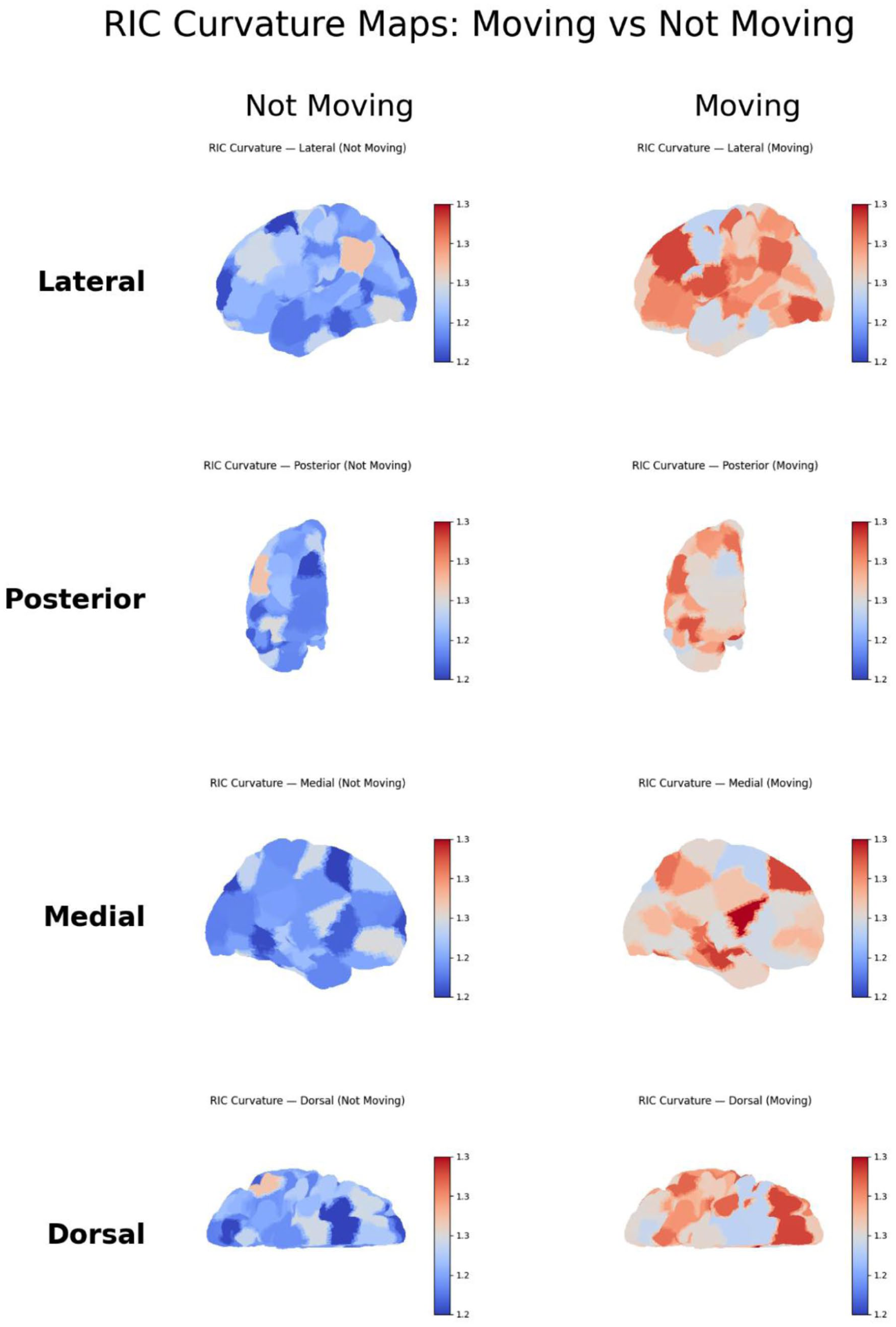
Surface Maps of RIC Curvature During Movement and Rest in the Food-Tracking ECoG Dataset. This figure presents mean RIC curvature projected onto 4 cortical views (lateral, posterior, medial, and dorsal) for the movement (moving) and rest (not moving) conditions during the macaque food-tracking task. Color scales represent interpolated RIC values across the cortical surface. Movement epochs show increased RIC curvature over sensorimotor and midline cortex, reflecting heightened recursive informational organization during active motor behavior. Rest epochs exhibit more distributed and lower curvature patterns.
UMAP Embedding and Microstate/Attractor Analysis
Across both datasets, UMAP projection of multichannel RIC features revealed discrete attractor structures within the low-dimensional neural manifold. In the eyes open/closed dataset, DBSCAN clustering identified microstates that closely tracked experimental state boundaries, while in the food-tracking dataset, multiple microstates were preferentially associated with “moving” or “not moving” epochs, indicating that RIC curvature captures latent motifs linked to behavioral transitions. We next evaluated the discriminative power of RIC-based features for state decoding using ROC analysis and microstate clustering. As shown in Figure 5 (top), multichannel RIC classifiers achieved near-perfect separation of cognitive and behavioral states (AUC ⩾ 0.84). Furthermore, UMAP embeddings and unsupervised clustering (Figure 5, bottom) revealed attractor microstates tightly aligned with experimental conditions, underscoring RIC’s capacity to delineate low-dimensional neural manifolds underlying brain state transitions.
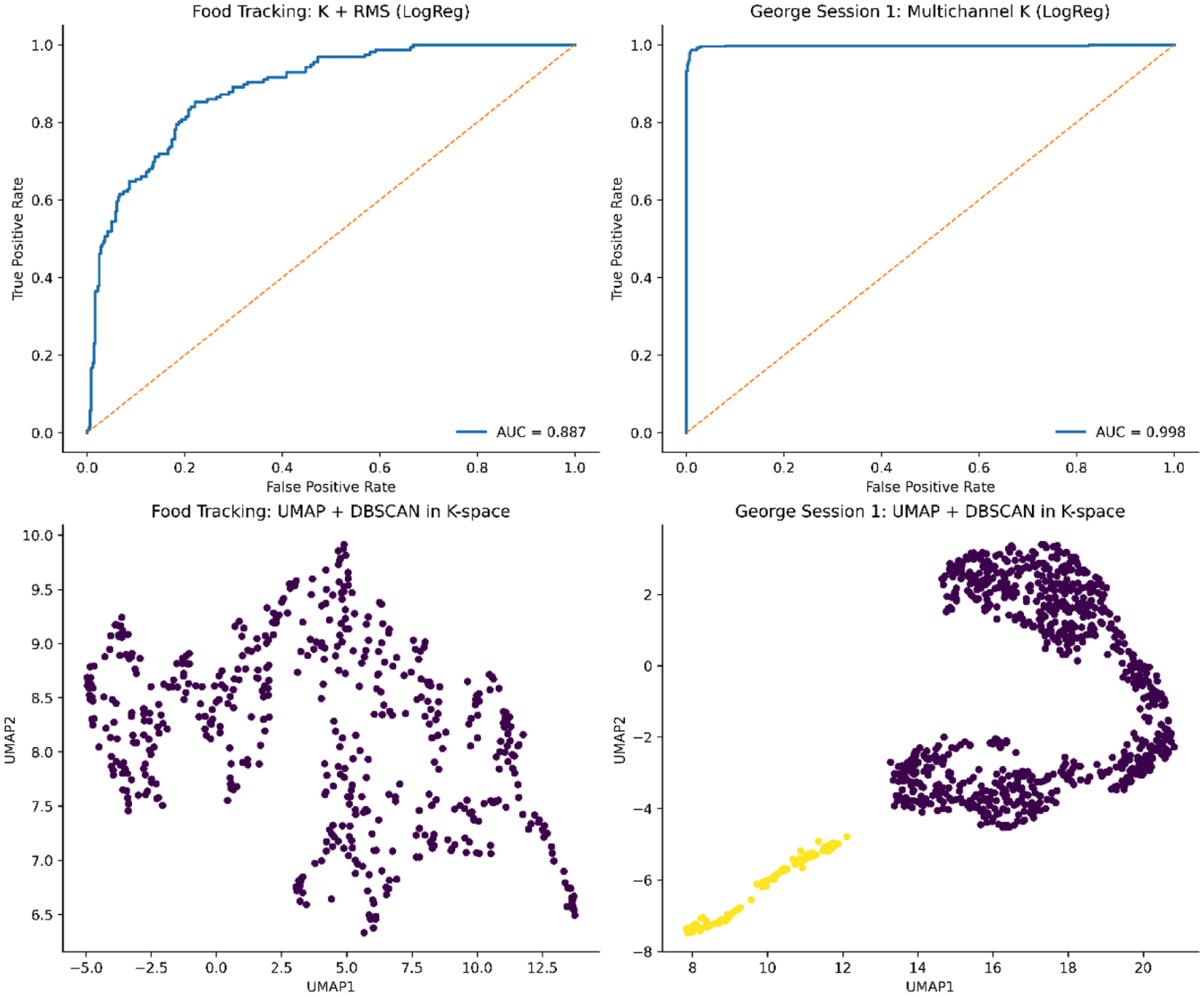
ROC analysis and low-dimensional manifold structure of state decoding in the George Session 1 eyes-opened versus eyes-closed benchmark and the food-tracking benchmark. Top row: Receiver operating characteristic curves for the best-performing updated decoding models. Left: In the food-tracking dataset, a logistic-regression model combining curvature
Quantification of State-Dependent Changes in RIC Curvature
Direct comparison of mean RIC curvature between states revealed significant, spatially structured differences. In the eyes open/closed paradigm, posterior and midline channels showed the most significant increases in RIC curvature during eyes closed (mean ΔRIC = +0.14 ± 0.02, paired t-test, P < .001), consistent across animals and sessions. In the food-tracking dataset, grid and brain surface maps showed focal increases in RIC during hand movement, with the difference map highlighting the sensorimotor strip (mean ΔRIC = +0.10 ± 0.02, P < .01). These results establish RIC curvature as a sensitive and anatomically informative marker of both cognitive and behavioral state transitions.
Comparative Benchmarking, Ablation, and Robustness Analyses
The condensed benchmark summary presented in Table 2 is expanded here through direct comparator, ablation, and robustness analyses designed to determine which parts of the curvature formulation contribute most strongly to decoding performance. To determine whether curvature
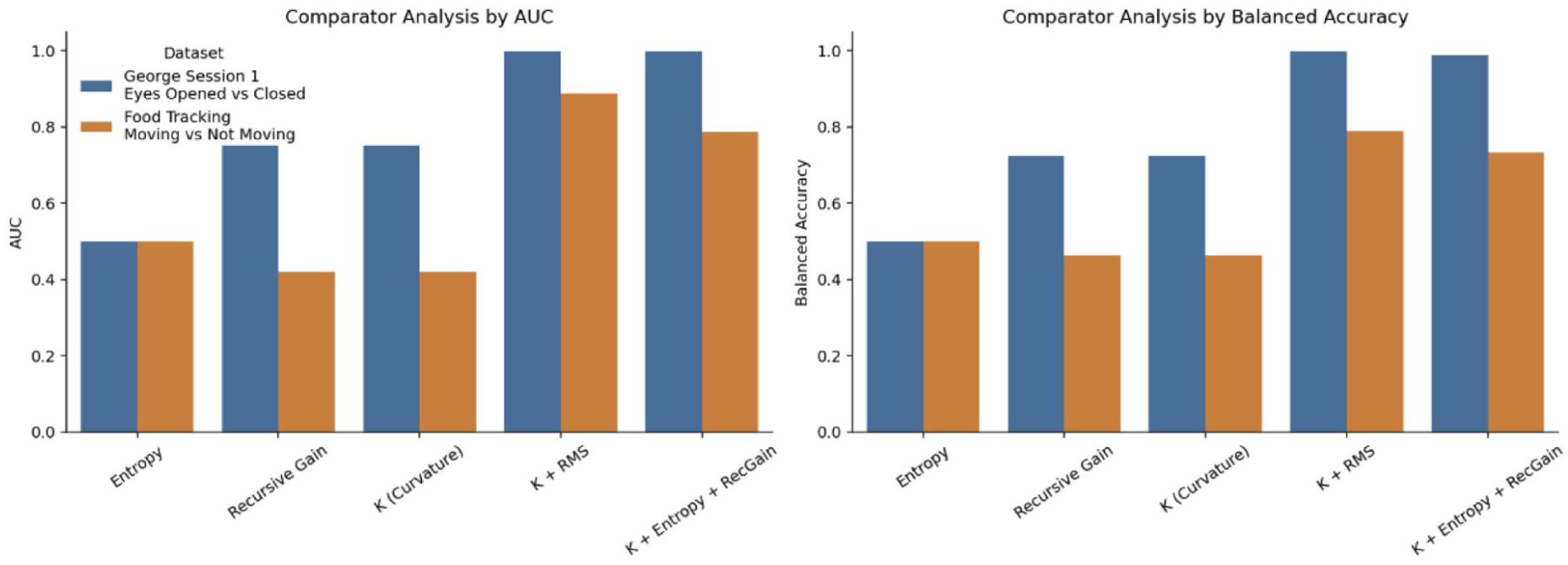
Comparative ablation analysis of benchmark feature families across the George Session 1 eyes-opened versus eyes-closed benchmark and the food-tracking moving-versus-not-moving benchmark. Bars show performance of logistic-regression models using entropy alone, recursive gain alone, the empirical curvature scalar
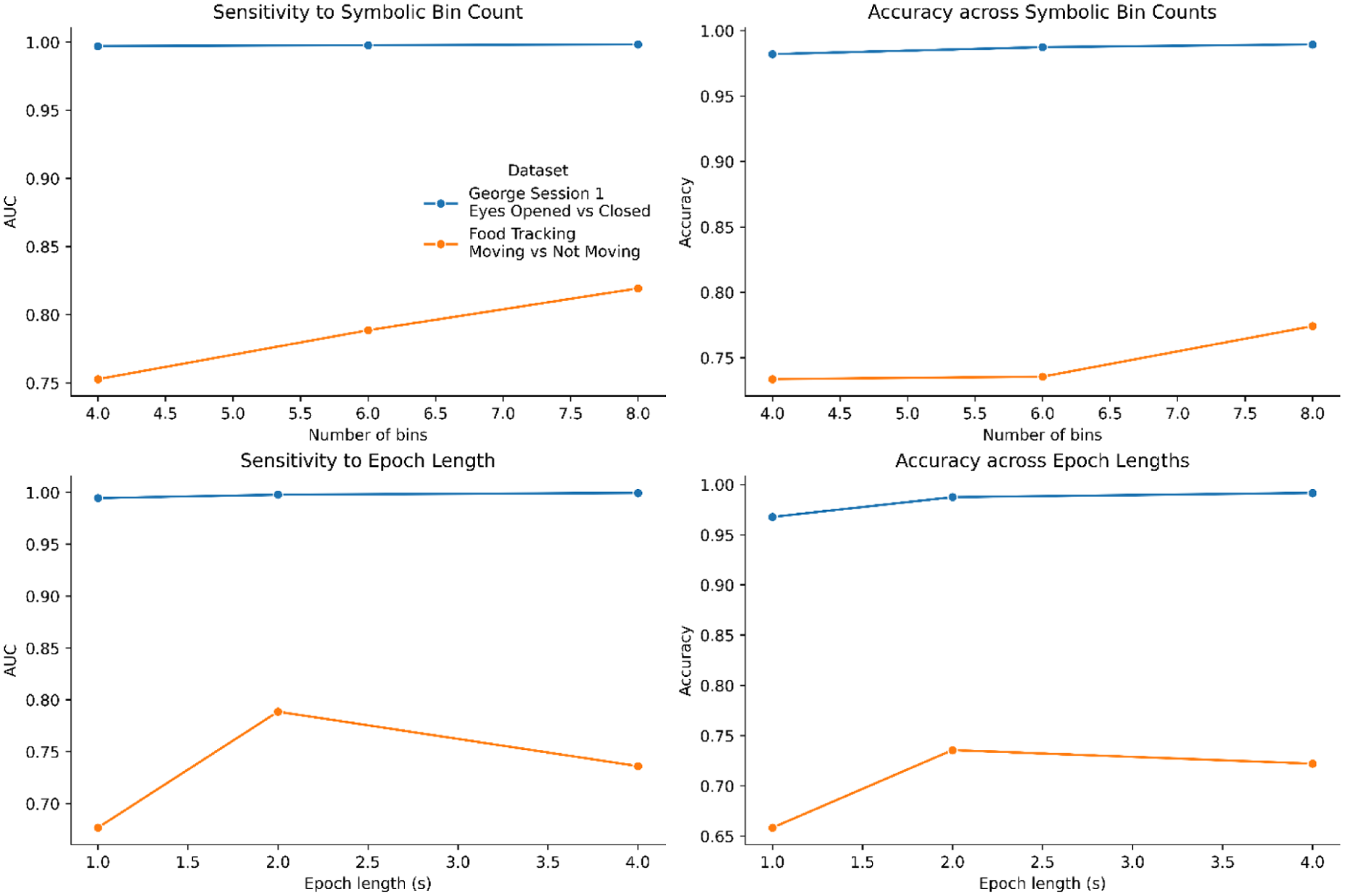
Sensitivity analysis of the empirical curvature scalar
Second, recursive gain and curvature showed closely matched performance profiles. In the George benchmark, multichannel recursive-gain and multichannel curvature models yielded nearly identical decoding performance, and the same pattern was observed in the food-tracking data. This suggests that, in the present empirical implementation, much of the discriminative information captured by curvature is concentrated in the recursive term. Accordingly, the current results support the usefulness of the curvature scalar as an interpretable signal-level feature, while also showing that the present benchmark does not yet identify a setting in which curvature clearly exceeds recursive gain in matched predictive analyses.
Third, combined-feature models were most beneficial in the more difficult food-tracking task. Whereas the George benchmark already approached ceiling performance with multichannel curvature-based models, the food-tracking benchmark improved when curvature was combined with RMS amplitude features. This pattern indicates that curvature is informative but not sufficient on its own for all paradigms, particularly when behavior is unconstrained and state boundaries are less sharply defined.
Sensitivity analyses further supported the robustness of the benchmark. In the George benchmark, performance remained high across symbolic bin counts and epoch lengths, with modest gains at longer epochs. In the food-tracking dataset, performance was more sensitive to analytic choices. The food benchmark improved with finer symbolic discretization, with 8-bin models outperforming 4-bin models, and 2-s epochs outperforming both 1-s and 4-s windows. Together, these results indicate that the empirical curvature term is stable and useful across parameter settings, while also showing that optimal performance depends on task structure and on explicit comparison with conventional feature families.
As an additional nonlinear comparator, we evaluated CEBRA-Time embeddings on the same 2 benchmark datasets. In the George Session 1 benchmark, CEBRA-Time achieved an accuracy of 0.748, balanced accuracy of 0.748, F1 score of 0.739, and AUC of 0.813. In the food-tracking benchmark, CEBRA-Time achieved an accuracy of 0.531, balanced accuracy of 0.513, F1 score of 0.375, and AUC of 0.508. In both paradigms, these results were markedly lower than those obtained with the best
Discussion
The present study provides the first empirical implementation and benchmark evaluation of the RIC curvature scalar in macaque ECoG. Across 2 datasets, multichannel curvature features supported strong state decoding, coherent low-dimensional organization of neural epochs, and anatomically interpretable spatial structure. At the same time, the benchmark defines an important boundary condition for interpretation: in the current implementation, the empirical curvature term remains closely aligned with recursive gain. The main contribution of the study is therefore not the demonstration of a fully independent theory-level quantity, but the establishment of a curvature-based formulation as a practical, interpretable, and reproducible descriptor of state-dependent neural organization (Figure 8).
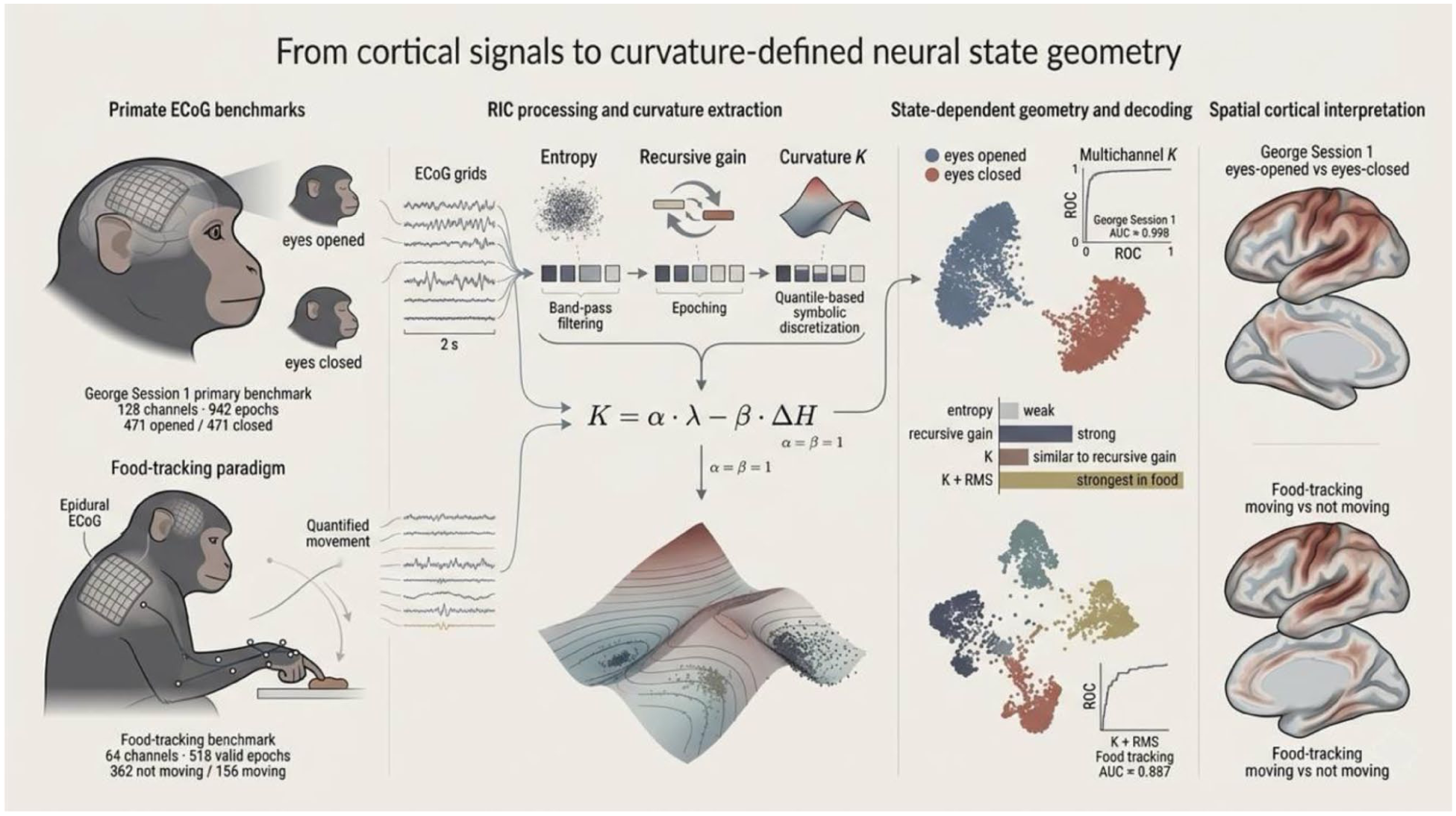
High-density macaque ECoG recordings from a balanced eyes-opened versus eyes-closed benchmark and a food-tracking behavioral paradigm were transformed into symbolic neural representations and analyzed using the empirical curvature scalar
Curvature-Based Decoding and Distributed Neural State Structure
A central result of the benchmark is that curvature is most informative when treated as a distributed multichannel feature rather than as a single global summary. In the primary George Session 1 benchmark, channel-resolved curvature profiles supported near-ceiling discrimination between eyes-open and eyes-closed states, indicating that the spatial configuration of curvature across the cortical array carries highly structured information about neural condition. In the food-tracking dataset, decoding was more difficult, as expected for a more heterogeneous and behaviorally unconstrained paradigm, yet curvature still captured useful state-relevant structure, particularly when combined with RMS amplitude features.
This distributed character is scientifically important. Neural states are unlikely to be defined by one scalar property at one recording site. They are expressed through coordinated, spatially extended patterns of local reinforcement, dispersion, and reconfiguration. The curvature formulation is suited to this problem because it compresses these interacting properties into a channel-resolved field that can be analyzed statistically, embedded geometrically, and projected anatomically. Its value lies not only in classification performance, but in providing a common representation that links decoding, neural state geometry, and cortical topography. 6
The low-dimensional analyses reinforce this point. UMAP embeddings of multichannel curvature features revealed structured neural manifolds in both datasets, with sharper organization in the eyes-open versus eyes-closed benchmark and weaker but still meaningful structure in the food-tracking task. In this sense, curvature is useful not merely as a classifier input, but as a compact representation of distributed neural state architecture.
The comparison with CEBRA-Time further strengthens this interpretation. Although CEBRA provides a modern self-supervised latent representation-learning framework, it underperformed the present
What the Framework Contributes
The comparator and ablation analyses substantially sharpen the interpretation of the empirical RIC scalar. Entropy alone was weak in both datasets, whereas recursive gain and curvature showed closely matched performance profiles across several analyses. This indicates that, in the present benchmark, much of the discriminative structure captured by curvature is concentrated in the recursive component of the formulation.
In the present benchmark, we did not identify a condition in which the empirical curvature term meaningfully outperformed recursive gain alone in matched decoding analyses. Across both datasets, recursive gain and curvature yielded closely similar, and in several cases identical, performance profiles, indicating that the current implementation is dominated by the recursive component of the formulation. The strongest food-tracking result was obtained when curvature was combined with RMS amplitude, which suggests potential complementarity in a harder behavioral setting; however, because the corresponding recursive-gain-plus-amplitude comparison was not part of the present benchmark, this should not be interpreted as definitive evidence of curvature-specific added predictive value. Likewise, although curvature proved useful for spatial mapping and manifold analysis, the present study did not perform matched side-by-side topographic or embedding comparisons against recursive gain sufficient to establish a robust qualitative divergence. We therefore interpret the main contribution of the current study more cautiously: not as proof that
That result does not diminish the value of curvature. Rather, it clarifies what curvature contributes at this stage of development. Its main empirical strength is not that it uniformly exceeds every constituent metric in isolation, but that it provides a compact and interpretable scalar that preserves state-relevant recursive structure while remaining compatible with anatomical mapping, manifold analysis, clustering, and combined-feature decoding. In the more difficult food-tracking benchmark, the best performance emerged not from curvature alone but from its combination with RMS amplitude, suggesting that curvature carries complementary information that becomes most useful when neural state boundaries are less cleanly separated.
This distinction is methodologically important. A benchmark feature need not justify itself solely by outperforming all alternatives in a classifier. A feature may also be valuable because it yields a more coherent representation of state structure, reveals distributed organization obscured by scalar summaries, or provides a mathematically unified descriptor that can be used across several modes of analysis. In the present study, the curvature formulation shows precisely this kind of value. 20
Neuroscientific Interpretation
The spatial organization of curvature observed across the 2 datasets suggests that the empirical RIC scalar tracks mesoscale changes in neural-state organization rather than merely reflecting undifferentiated changes in signal magnitude. In the eyes-open versus eyes-closed benchmark, higher curvature during eyes-closed epochs was concentrated more strongly over posterior and parietal territories. This pattern is consistent with a shift toward more internally organized cortical dynamics under reduced visual engagement. In the food-tracking dataset, curvature increases were more focal and behaviorally distributed, particularly over sensorimotor and midline regions, in line with movement-related reconfiguration of cortical dynamics during action and sensorimotor control. 20
These findings help explain why dynamic geometric descriptors are useful in electrophysiology. Neural-state changes are often accompanied by simultaneous changes in recurrent dependence, variability, and spatial patterning. Conventional descriptors usually isolate one of these dimensions. Spectral power captures amplitude structure, entropy captures dispersion, and connectivity metrics capture statistical dependence, but none of them directly encodes the evolving balance between recursive reinforcement and dispersive reorganization across the cortical array. The curvature field provides a way to represent that balance in a form that is both local and spatially distributed.2,7
The present results also support a geometric interpretation of neural states in a concrete empirical sense. Each epoch can be represented by its channel-wise curvature vector, and the resulting collection of vectors defines a structured neural state space. This makes it possible to describe neural conditions not only by whether they differ statistically, but by how they occupy distinct regions of a distributed curvature geometry. Within the scope of the present benchmark, this is the principal neuroscientific significance of the framework: it links signal-level analysis to spatially organized neural-state structure in a form that can be visualized, classified, and compared.
Relation to Broader Theoretical Frameworks
RIC was originally motivated by a broader information-geometric perspective in which recursive structure and entropy change jointly shape system organization. 21 The present study evaluates only a restricted empirical implementation of that program. Here, the curvature scalar is treated as a signal-level descriptor derived from symbolized ECoG data, not as a direct measure of consciousness, integrated information, or full generative inference. 22
This distinction is especially important when situating RIC relative to existing frameworks such as Integrated Information Theory (IIT) and the Free Energy Principle (FEP).22-24 IIT is formulated at the level of system-wide causal structure and partition-sensitive integration, whereas FEP characterizes biological systems in terms of generative modeling, prediction, and variational self-organization. By contrast, the present implementation of RIC operates at an intermediate descriptive level: it does not require partition enumeration, full causal architecture, or specification of an explicit generative model. Instead, it provides an empirically tractable curvature descriptor that summarizes the evolving balance between recursive dependence and entropy change in neural signals. This gives RIC a distinct practical advantage for benchmark electrophysiology: it is directly computable from real data, spatially mappable across the cortical array, and readily usable in decoding, embedding, and manifold analyses. 25
The advantage of this formulation is therefore methodological as much as conceptual. Relative to IIT, it avoids the combinatorial and interpretive difficulties associated with estimating large-scale causal integration from neural recordings. Relative to FEP, it does not depend on fitting a latent generative model or inferring hidden-state dynamics before empirical comparison becomes possible.26,27 What RIC offers instead is a compact geometric representation of neural-state organization that remains close to measurable signal structure while still capturing interactions between recurrence and dispersion. In this respect, the present benchmark suggests that curvature-based analysis may provide a useful intermediate framework between descriptive signal statistics and more ambitious theory-level accounts of neural organization.
Limitations and Future Directions
Several limitations define the present scope. First, all analyses were conducted on macaque ECoG under controlled experimental conditions. Generalization to human recordings, clinical cohorts, or more heterogeneous behavioral environments remains to be established.28-30 Second, although curvature performed strongly in decoding and geometric analyses, its present implementation did not separate sharply from recursive gain. Future refinement should therefore focus not only on classification performance, but on identifying conditions under which the entropy-change term contributes uniquely to state characterization. In the present implementation, the close empirical correspondence between
Third, the current benchmark is intentionally simple. Recursive gain was estimated through lag-1 mutual information within symbolic epochs, and entropy change was defined across adjacent epochs after symbolic discretization. These choices favored transparency and reproducibility, but they do not exhaust the design space of the framework. Alternative delay structures, richer symbolic alphabets, multiscale entropy-change formulations, and graph-structured or nonlocal extensions of the curvature field may produce more distinctive behavior and more biologically informative state geometry.
Fourth, the present analyses do not provide a circuit-level mechanism. The curvature field is informative at the level of signal organization and spatial state structure, but it is not yet tied to identified synaptic, laminar, or network mechanisms. Establishing those links will require perturbational paradigms, mechanistic modeling, and multimodal datasets that integrate ECoG with anatomical or connectivity-informed measurements.
These limitations also define the next phase of the framework. The immediate task is not merely to repeat the benchmark on additional datasets, but to determine when curvature diverges meaningfully from recursive gain, whether its spatial organization remains stable across species and recording modalities, and whether curvature-based neural-state geometry adds value in settings involving richer behavior, continuous state transitions, anesthesia gradients, or pathological dynamics. The present study therefore serves as a rigorous starting point: it shows that an empirical curvature scalar can be computed reproducibly, mapped across the cortical surface, and used to characterize state-dependent organization in real neural data, while also identifying the conceptual and methodological questions that must be answered for the broader RIC framework to mature further. Future work should therefore include explicit matched tests of curvature versus recursive gain in combined-feature models, parameter-robustness analyses, and side-by-side spatial and manifold comparisons, so that any unique contribution of the entropy-related term can be demonstrated directly rather than inferred.
Conclusion
This study provides the first empirical benchmark of the RIC curvature scalar in macaque ECoG and demonstrates that it captures state-dependent neural structure across both controlled and behaviorally richer paradigms. Multichannel curvature features supported strong decoding performance, while in the food-tracking dataset the combination of curvature and amplitude-based features further improved classification. At the same time, the present analyses show that recursive gain remains a strong comparator and, in the current implementation, often performs similarly to curvature. The contribution of the benchmark is therefore twofold: it establishes the empirical curvature term as a practical and interpretable signal-level descriptor of neural-state organization, and it clarifies the conditions under which claims about its added value should be made with precision. Taken together, these findings establish curvature-based analysis as a reproducible and geometrically grounded framework for characterizing distributed neural-state organization in real electrophysiological data.
Supplemental Material
sj-docx-1-exn-10.1177_26331055261460858 – Supplemental material for The Geometric Signatures of Brain State Transitions: Recursive Informational Curvature Reveals Hidden Dynamics in Primate Cortex
Supplemental material, sj-docx-1-exn-10.1177_26331055261460858 for The Geometric Signatures of Brain State Transitions: Recursive Informational Curvature Reveals Hidden Dynamics in Primate Cortex by Mahsa Asadi Anar, Seyed Kiarash Sadat Rafiei, Soroosh Najafi, Parham Mahmoudi, Hossein Gharedaghi, Sasan Ghazanafar Ahari, Maryam Rafiei, Pouya Asgari, Zahra Narimani and Mohammad Saeed Soleimani Meigoli in Neuroscience Insights
Footnotes
Acknowledgements
We appreciate all the scientists whose work inspired this study.
Author Contributions
Study concept and design: SKSR, MAA. Acquisition of the data: MAA, SKSR. Analysis and interpretation of the data: MAA, SKSR. Drafting of the manuscript: SKSR, MAA, HG, PM, SN, SSGA, MR, PA, ZN, MSSM. critical revision of the manuscript for important intellectual content: SKSR, MAA. Administrative and technical support: MAA, SKSR. Study supervision: SKSR, MAA. Supplementary materials: SKSR.
Funding
The authors received no financial support for the research, authorship, and/or publication of this article.
Declaration of Conflicting Interests
The authors declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Data Availability Statement
All data analyzed in this study are publicly available. Code for preprocessing, curvature extraction, benchmarking, sensitivity analyses, and figure generation is available at:
The George anesthesia-and-sleep dataset used for the primary eyes-opened versus eyes-closed benchmark is available at:
The food-tracking dataset is available at:
No new raw data were generated. Derived outputs are reproducible from the public data and code repository and are available from the corresponding authors upon reasonable request.
Supplemental Material
Supplemental material for this article is available online.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.