
Abstract
Background:
The escalating antibiotic resistance necessitates alternative strategies like bacteriophages, utilized in livestock for growth promotion and infectious disease treatment. To select promising phage candidates, this study aimed to characterize Escherichia phages with potential against swine-associated Escherichia coli.
Methods:
Four E. coli phages were characterized for morphology, efficiency of plating, temperature stability, tolerance of simulated gastric fluid, long-term storage in various solutions, and lytic activity of individual and combined phages against pathogenic E. coli in tryptic soy broth and simulated intestinal fluid.
Results:
All phages exhibited infectivity and stability across 4°C–45°C, with varying sensitivity to low pH. The phages maintained stability in culture medium and select buffers at 4°C for at least 52 weeks. All phages exhibited significant lytic activity, with the cocktail showing enhanced reduction in certain conditions.
Conclusion:
These four Escherichia phages demonstrate potential as biosanitation agents for swine housing and biocontrol strategies against swine colibacillosis, warranting further in vivo investigation.
Introduction
The increasing problem of antibiotic resistance has been projected to significantly raise annual deaths, thus increasing awareness of antimicrobial-resistant (AMR) pathogens. The misuse and overuse of antimicrobial agents in agriculture and human medicine is a major factor in the development and spread of AMR pathogens. Antibiotic residues in the environment and food animals also pose risks to human health, including effects on the immune system and gut microbiota.1–3 Nevertheless, the use of antimicrobials for both human and animal welfare remains essential, inevitably leading to the continued use of antibiotics. Thus, alternative methods are urgently required to combat pathogens, especially AMR strains.
Bacteriophages (phages) are viruses that replicate within bacterial hosts, leading to host cell lysis and the release of progeny virions. The ecological relationship between phages and their hosts indicates that phages can be isolated from the related locations. Initially applied therapeutically in Eastern Europe a century ago with limited Western use, phages are now experiencing a global resurgence for various applications due to increasing research and implementation worldwide. Phage therapy employs various strategies, including conventional use of natural phages, dual therapy combining phages with antimicrobials, the application of genetically modified phages for targeted effects, and the use of phage-derived proteins, such as endolysins. 4 Phages have been utilized in various other fields, for instance, environmental applications as bioindicators for water contamination detection, 5 industrial applications such as food biocontrol and biosanitation, 6 agricultural applications in both crop and livestock sectors, 7 and veterinary applications in the form of phage therapy for both farm and companion animals. 8
In the livestock sector, phages have been developed and employed for promoting growth through feed additives and treating infectious diseases with their antimicrobial properties. 9 A study evaluating a commercial phage product, probiotics, and their combination as pig feed additives demonstrated that phage supplementation significantly improved average daily gain, average daily feed intake, and feed-gain ratio. Furthermore, pigs receiving phages showed higher fecal levels of beneficial bacteria (Bifidobacterium spp. and Lactobacillus spp.) and lower levels of potentially harmful bacteria (Clostridium spp. and Coliforms) compared to the control and probiotic-only groups. 10 Recognizing this potential, the selection of robust and effective phages necessitates a thorough understanding of their fundamental characteristics and lytic capabilities.
Addressing the critical need for targeted biocontrol in swine production, this study analyzed the characteristics of four distinct Escherichia phages. Their infectivity, stability, storage conditions, and lytic activity were determined. Moreover, the lytic activity of individual phages and their synergistic potential as a cocktail were quantified against a combined pathogenic E. coli culture under conditions relevant to swine nurseries.
Materials and Methods
Bacterial strains and phages used in this study
Four phages, named biotsu, sutha, kaset, and nithesis, were isolated, with E. coli strains M484, M486, M491, and M493 serving as their respective hosts. These E. coli strains were originally isolated from the intestinal specimens of pigs with porcine diseases. 11 The characteristics of each phage were determined, and plaque assays were performed using their specific isolation host. Prior to each experiment, bacterial hosts were cultured overnight in tryptic soy broth (TSB) at 37°C. The plaque assay was performed to determine the phage titer by mixing phage suspension with an overnight bacterial culture and molten tryptic soy agar (TSA) containing 0.45% agar. The mixture was overlaid on a TSA plate. Plaques were counted after incubation at 37°C for 16 h and calculated as plaque-forming units per milliliter (PFU/mL).
Morphology investigation
Phage suspension (>109 PFU/mL) was applied as a droplet onto a parafilm sheet. A formvar-coated copper grid was then carefully placed onto the phage droplet for 10 min to allow phage adsorption. The grid was then negatively stained with a 2% uranyl acetate solution for 2 min. Phage morphology was subsequently examined using a Hitachi HT7700 transmission electron microscope (Hitachi, Japan). The size of 10 particles of each phage was measured. The size measurements are presented as mean ± standard deviation.
Efficiency of plating
The experiment was carried out using the plaque assay technique of the tested phages with their respective host strains. The plates were incubated at 10°C, 15°C, 20°C, 25°C, 30°C, 37°C, 42°C, and 45°C for 18 to 48 h. 12 The efficiency of plating (EOP) was calculated using the following formula: phage titer at each temperature divided by the highest phage titer observed across all tested temperatures. The EOP value reveals phage lysis efficiency at each temperature: EOP > 0.5 = high efficiency, 0.5 > EOP > 0.2 = medium efficiency, 0.2 > EOP > 0.001 = low efficiency, and EOP < 0.001 = inefficiency.
Temperature and acid tolerance
For the temperature stability test, each phage suspension was diluted in pre-warmed TSB to a final titer of approximately 107 PFU/mL. The diluted suspensions were then incubated at 4°C, 28°C, 39°C, and 45°C for time intervals of 2, 4, 6, 12, and 24 h. At each specified time point, samples were collected, and plaque survival was determined using the plaque assay technique.
For acid tolerance assessment, phage suspensions were subjected to simulated gastric fluid (SGF) 13 and incubated at 39°C for 2 h. Following this incubation period, samples were taken, and the plaque assay was performed immediately for phage enumeration.
Phage storage assessment
An aliquot (10 µL) of each phage stock (1010 PFU/mL) was added to 990 µL of the different test solutions. Phage long-term storage was assessed over a period of 52 weeks (for 0, 2, 4, 8, 12, 16, 20, 24, 36, and 52 weeks) with six different solutions (TSB, saline-magnesium [SM] buffer, 0.9% NaCl, 5% glucose solution, Dulbecco’s phosphate-buffered saline [DPBS] with Ca2+ and Mg2+, and DPBS without Ca2+ and Mg2+). The experiments were performed at 4°C. Phage titers were quantified at the indicated time intervals using the plaque assay technique.
Phage lytic activity investigation
Lytic activity was assessed through two experimental sets: individual phage assays and phage cocktail assays. For individual phage assays, each phage was tested against its specific host at multiplicities of infection (MOIs) of 1 and 10 in both simulated intestinal fluid (SIF) (10 mg/mL pancreatin and 20 mg/mL bile salts in 50 mM KH2PO4, pH 6.8) at 39°C and TSB at 30°C. For the phage cocktail assays, all phages and their respective hosts were combined in SIF and TSB to achieve MOIs of 10 and 1, respectively. Control groups, without phage addition, were included in both assays. All experiments were conducted for 3 h, with samples collected at hourly intervals for bacterial enumeration.
Statistical analysis
One-way analysis of variance with Tukey’s post-hoc test (IBM SPSS Statistics v.23) was run to assess statistical significance of the temperature, acid tolerance, and phage storage experiments. Linear mixed-effects models for repeated measures executed via R (version 4.4.2) 14 and RStudio (version 2024.12.0.467) 15 were employed to determine the differences in bacterial titer at each time and condition for the lytic activity investigation. For all analyses, statistical significance was defined as a p-value of <0.05.
Results
Phage morphology
The TEM micrograph revealed distinct morphologies of phage icosahedral heads and tails (Fig. 1). Plaque appearances of each phage were provided along with the phage morphology. Phages biotsu and nithesis exhibited symmetric icosahedral heads with average diameters of 53.28 ± 1.73 nm and 59.40 ± 1.54 nm, respectively, and short tails measuring 11.64 ± 1.69 nm and 14.78 ± 1.79 nm. In contrast, phages sutha and kaset possessed long, contractile tails with average lengths of 99.85 ± 2.05 nm and 100.00 ± 2.54 nm, respectively. The phage sutha displayed an asymmetrical icosahedral head with an average width of 85.45 ± 1.62 nm and an average length of 93.73 ± 2.52 nm, while the phage kaset had a symmetric icosahedral head with an average width of 64.48 ± 1.42 nm and an average length of 64.03 ± 1.48 nm. Based on these morphological characteristics, phages biotsu and nithesis were classified as podoviruses, whereas phages sutha and kaset were classified as myoviruses.

Transmission electron micrographs and plaque appearances of
Efficiency of plating
The EOP assay demonstrated that phages biotsu and kaset exhibited high lytic efficacy across all tested temperatures (Figs. 2a, c). Phage nithesis also displayed a strong lytic efficacy across the temperature range, although its EOP was moderate at 10°C (Fig. 2d). Phage sutha showed the highest EOP at 37°C (Fig. 2b) and maintained high infectivity at both 30°C and 42°C. Moderate plating efficiency was observed for phage sutha at 15°C, 20°C, and 45°C. Notably, phage sutha showed no detectable infectivity at 10°C.
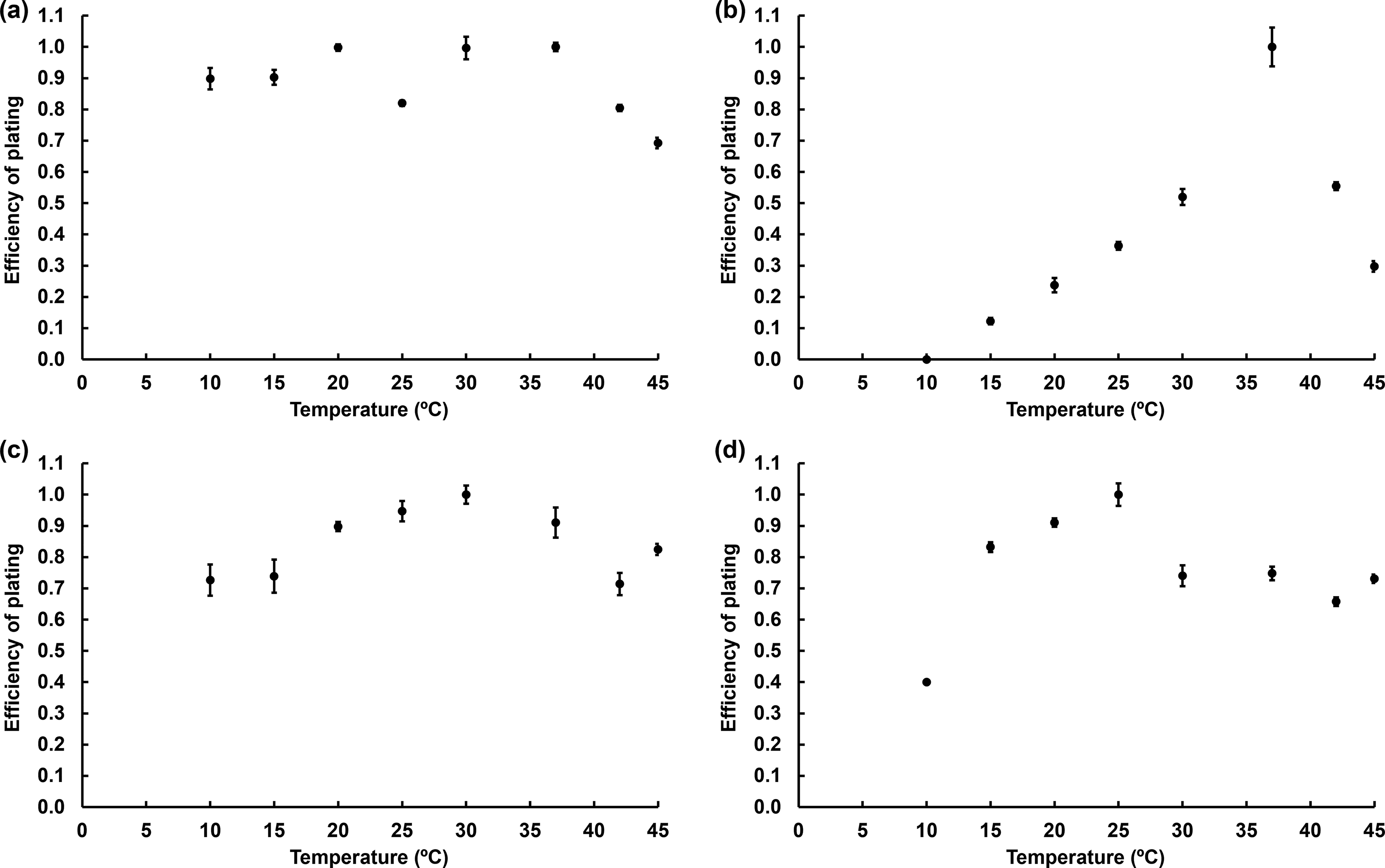
Efficiency of plating of
Temperature and acid tolerance
Phage biotsu demonstrated stability across all tested temperatures for at least 24 h (Fig. 3a), whereas the other phages exhibited minor reductions in titer at 39°C and 45°C. Phage sutha and nithesis showed a significant decrease in phage titer at 39°C (at 24 h) and at 45°C (starting from 12 h) (Fig. 3b, d), with a maximum reduction of <0.4 log PFU/mL observed at 45°C at 24 h. Phage kaset showed a detectable and significant titer reduction starting at 2 h, reaching a decrease of 0.11 and 0.14 log PFU/mL when tested at 39°C and 45°C, respectively (Fig. 3c).
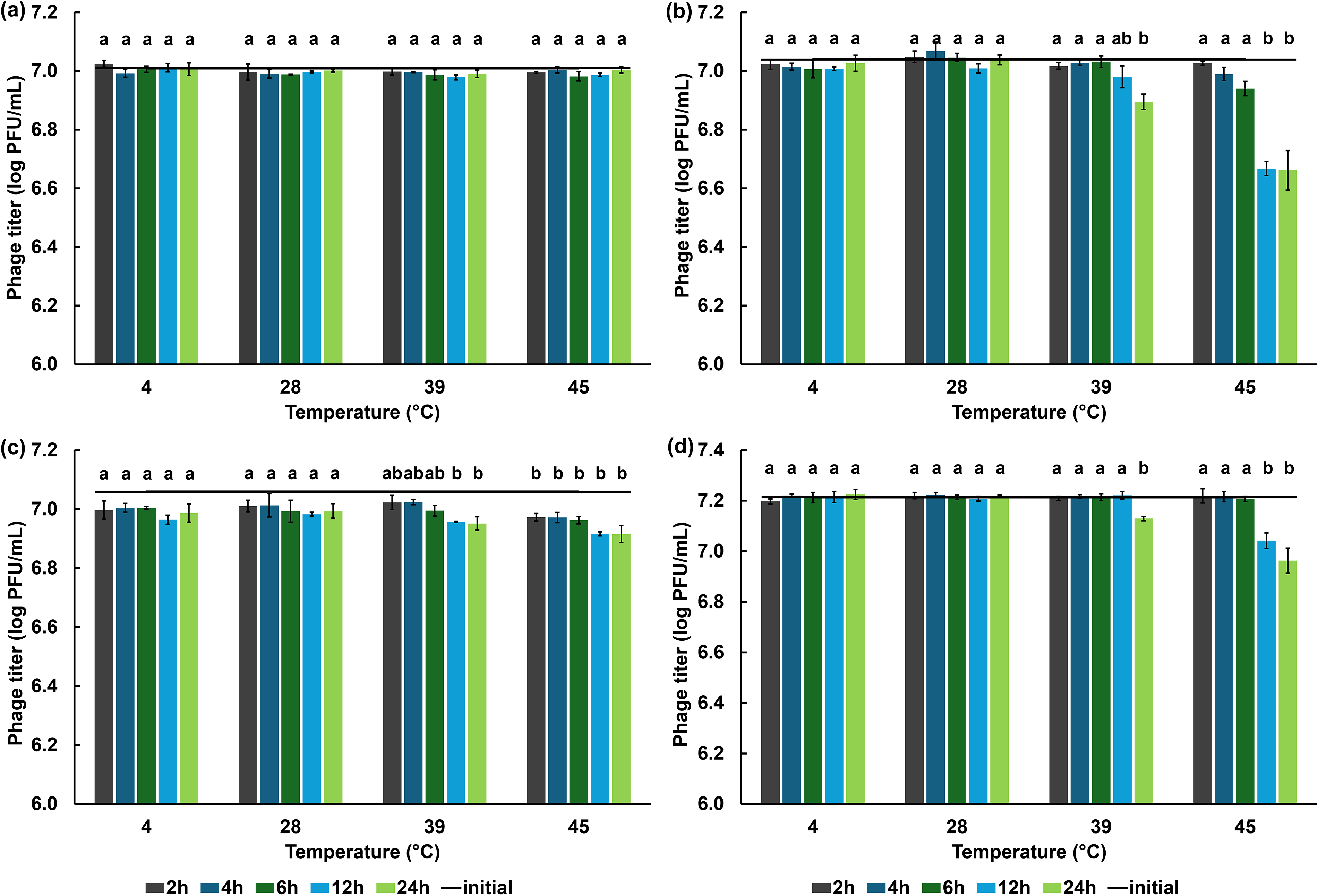
Temperature stability of
Upon exposure to SGF, phage biotsu exhibited complete inactivation under all tested conditions, while the other phages had different levels of acidic tolerance (Fig. 4). Phage sutha was undetectable at pH 2.5; however, survival was observed at pH 3, 3.5, and 4, albeit with significant titer reduction. In contrast, phage kaset demonstrated survival in SGF across all tested pH levels with a 1.5 log PFU/mL reduction at the lowest pH (pH 2). Phage nithesis did not tolerate low pH. At pH 4, the titer decreased by 5.6 log PFU/mL, becoming undetectable at more acidic conditions.
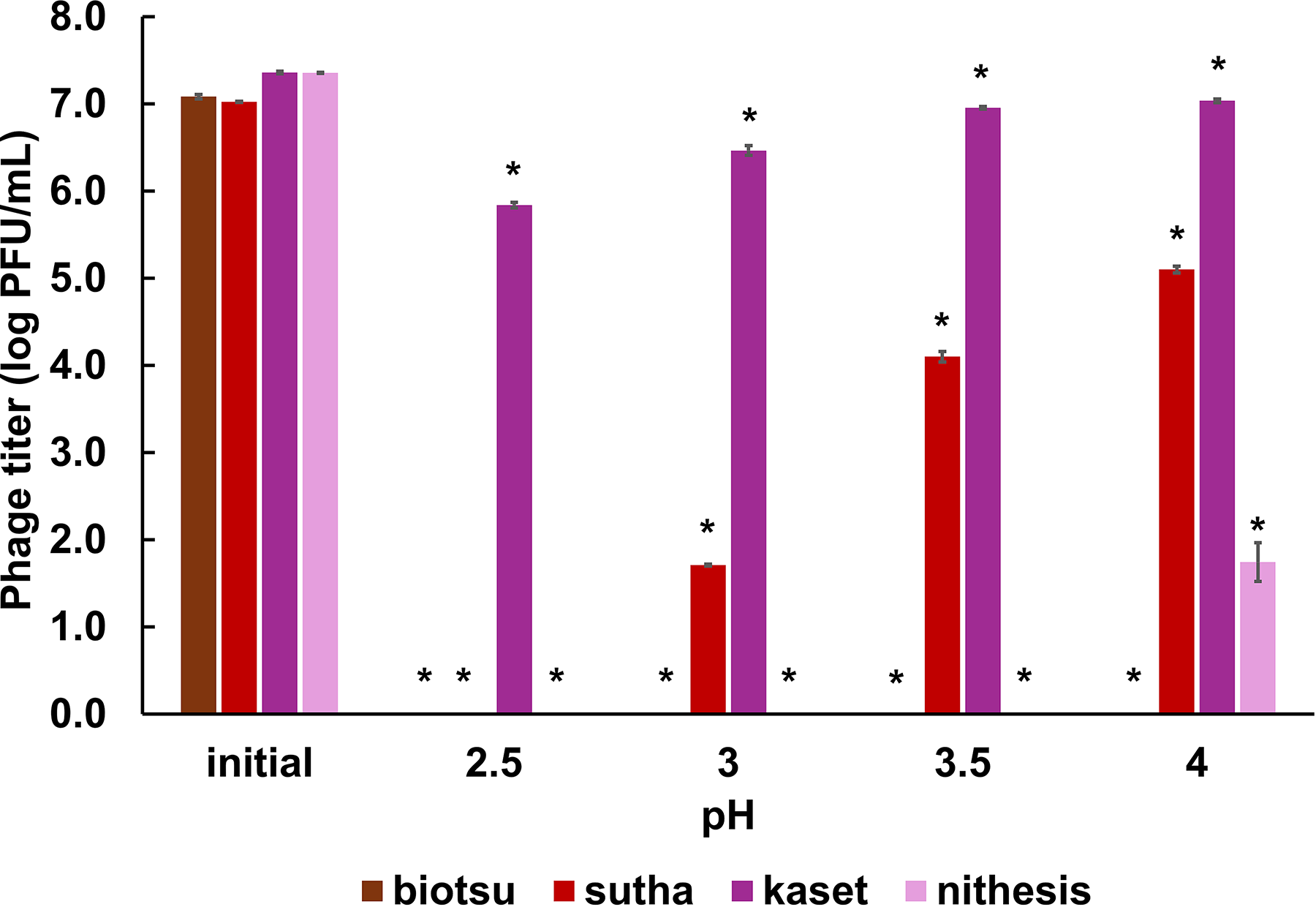
Simulated gastric fluid stability of phages biotsu, sutha, kaset, and nithesis at four different pH values after 2 h of incubation. Results are represented as mean ± SE. Asterisks (*) indicate a statistically significant difference (p < 0.05) in phage titer compared to their respective initial titer for each phage.
Phage storage
All phages were sensitive to 5% glucose solution. Active phage titers significantly decreased over time depending on the storage solution (Fig. 5). For phage biotsu, the titer fluctuated continuously across all solutions, except for DPBS (Fig. 5a, Supplementary Table S1). In DPBS, a change in phage titer was observed at week 12, after which no significant change occurred up to week 54. Phages sutha and nithesis exhibited stable titers in all solutions except glucose solution for 36 weeks (Fig. 5b, Supplementary Table S2, Fig. 5d, Supplementary Table S4). In contrast, phage kaset exhibited a reduction in titer across all storage solutions throughout the 52-week period (Fig. 5c, Supplementary Table S3). Notably, phage kaset displayed significantly better stability in SM buffer and 0.9% NaCl solution at each weekly assessment. The maximal titer reduction for phage kaset was ∼1.5 log PFU/mL, whereas the other phages exhibited titer reductions of <1.0 log PFU/mL.
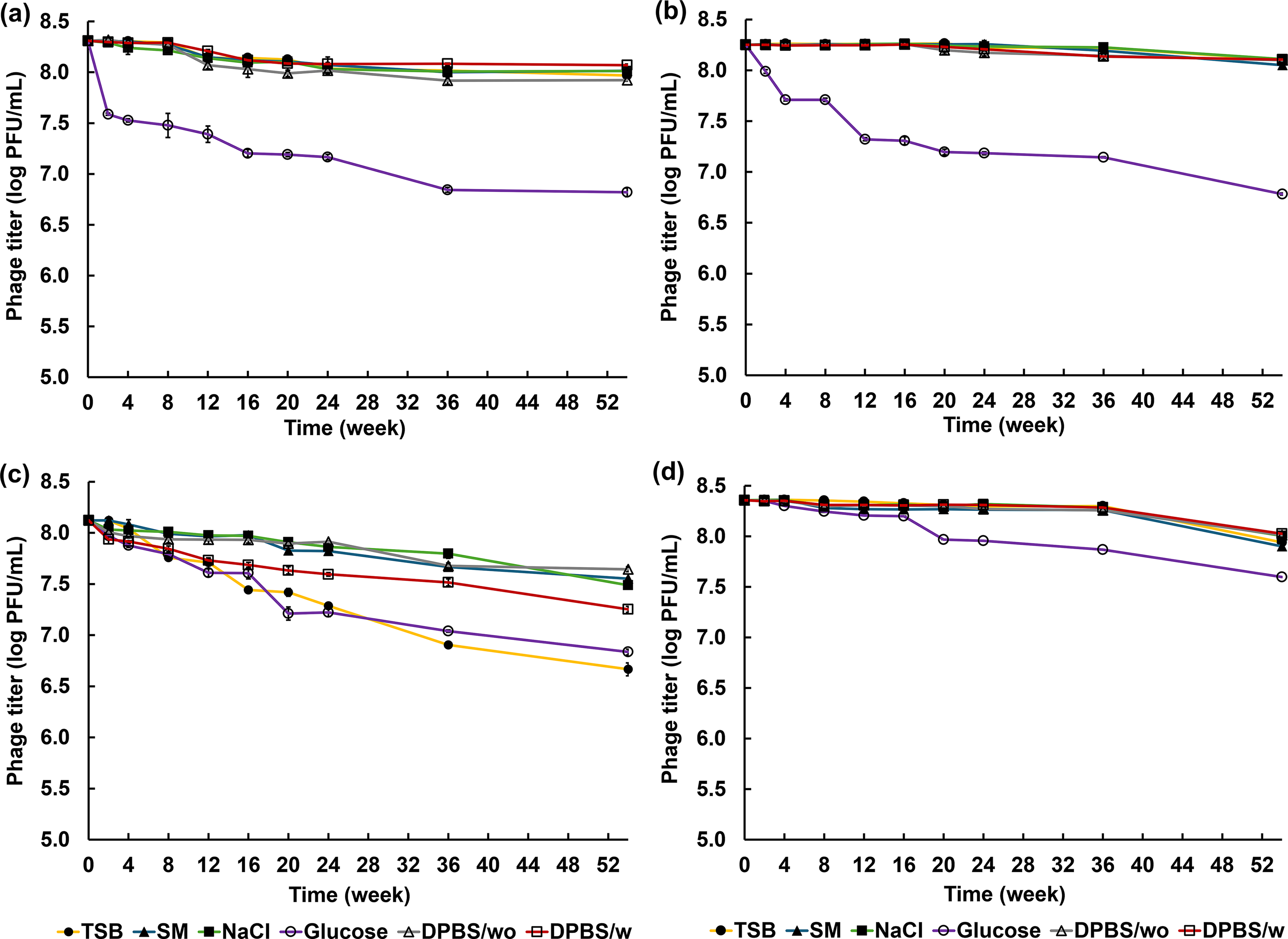
Long-term storage of
Phage lytic activity
As can be seen in Figure 6, bacterial titer was significantly influenced by the experimental duration. Furthermore, the phage activity varied depending on the MOI and the experimental condition. Phage biotsu and nithesis exhibited similar lysis patterns (Figs. 6a, d). Both phages significantly reduced their hosts within the first hour, with host titers diminished by ∼4–5 log CFU/mL at the end point. No significant differences in bacterial reduction were observed between MOIs until the final hour for these phages. For phage sutha, the lytic patterns under each phage condition were significantly different (Fig. 6b). However, it reduced its host by ∼5 log CFU/mL in both experiments by the end. For phage kaset, a clear difference in phage activity was observed in the SIF experiment between MOI 1 and 10, but not in TSB (Fig. 6c). At the final point, the phage diminished its host by ∼4 log CFU/mL at both MOIs in TSB. In contrast, host reduction in SIF was 0.3 and 2.0 log CFU/mL at MOI 1 and 10, respectively.
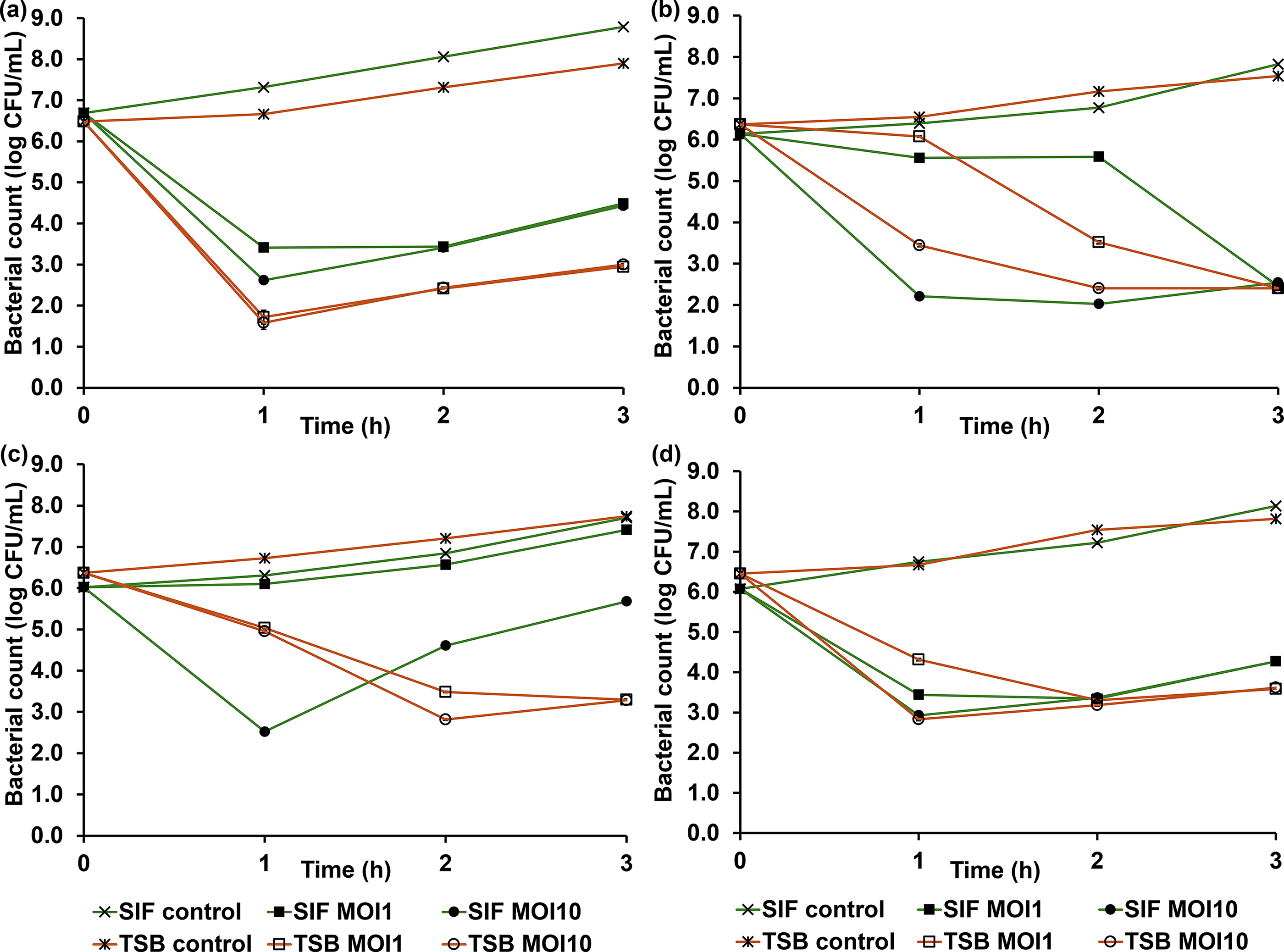
Lytic activity of individual phages:
In the phage cocktail experiment, control groups showed no significant difference during the first hour, whereas treatment groups exhibited significant variation in lytic activity (Fig. 7). A significant reduction in bacterial titters was observed in both SIF and TSB treatment groups from the first hour through the end of the experiment. At the end point, the phage cocktail demonstrated significantly higher lytic activity in TSB compared to SIF, resulting in bacterial reductions of 5.6 and 3.0 log CFU/mL, respectively.
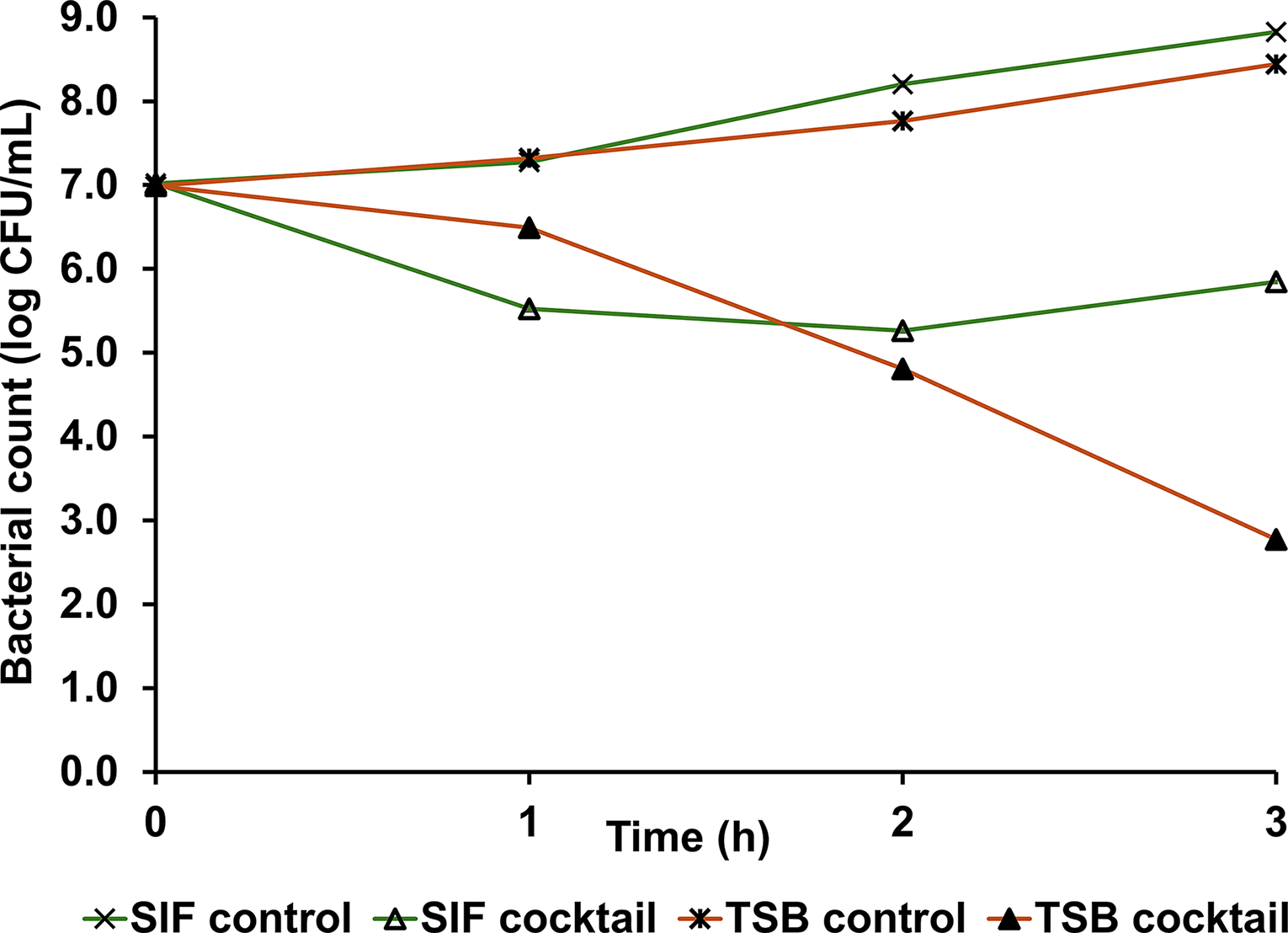
Lytic activity of phage cocktail in TSB at 30°C (MOI 1) and SIF at 39°C (MOI 10). Results are represented as mean ± SE.
Discussion
The escalating global health concern of antimicrobial resistance has increased interest in bacteriophages as therapeutic and biocontrol agents. As a result, sustained research and development efforts are focused on characterizing, optimizing, and implementing effective phage-based solutions across a wide range of applications. The extensive diversity of phages underscores the importance of selecting phages with appropriate efficacy for a specific use. This study characterized four potential Escherichia phages, focusing on their infectivity, stability, and lytic activity. Additionally, the lytic activity of these phages, individually and in combination, was evaluated under conditions simulating the porcine intestinal environment and nursery housing.
All four phages exhibited high efficiency of plating across a broad temperature range (10°C–45°C). However, phage sutha demonstrated a gradual loss of infectivity at temperatures deviating from its optimum of 37°C. Moreover, all phages maintained their stability over 24 h between 4°C and 45°C, with minor reductions only at 39°C and higher. Temperature is a critical factor in swine production, influencing growth, yielding, and critically posing a heat stress risk and meat yield, and importantly, heat stress poses a significant threat to pig health and can lead to mortality.16,17 Consequently, adjusting the temperature according to each growth stage is essential in swine production. Comfortable temperature ranges of 18°C–32°C are suggested for nursery and weaned pigs, with ideal mat temperatures between 30°C and 44.5°C. 18 Given that colibacillosis, caused by pathogenic E. coli, can affect pigs of all ages, 19 these bacteria can cause diseases under the wide range of temperatures encountered within swine housing and the pig body. The EOP and temperature stability results suggest that these phages are suitable for application throughout swine production, maintaining lytic activity across relevant temperature conditions. Furthermore, their stability at ambient temperatures suggests that refrigeration might not be required for short-term use.
Given that the bacteria in this study were isolated from the intestines of infected pigs, they were likely intestinal pathogens. While oral administration is a common approach for antimicrobial treatment of intestinal infections, the stability of biological agents like phages through the gastrointestinal tract is crucial for reaching their target site in the intestine. In this study, exposing the isolated phages to SGF at acidic pH levels revealed distinct patterns of gastric tolerance among the phage isolates. Colom et al. 20 reported up to ∼8 log PFU/mL reduction in phage titer after 1 hour in SGF, with encapsulated phages showing greater survival. In vivo studies in chicken caeca confirmed the better persistence of encapsulated phages compared to non-encapsulated phages. Both groups, however, effectively controlled Salmonella in vivo and persisted in the caeca until the end of the experiment. Similarly, other research has shown that while susceptible to SGF, free phages can still reach and remain active within the intestinal environment.21–23 However, encapsulation, high-dose administration, and repeated phage delivery have been proposed to maintain phage efficacy for oral phage therapy. 24
The phage storage solution is another critical factor for various administrative and production processes, as it can affect phage retention and infectivity. 25 Our finding that all phages were sensitive to the 5% glucose solution aligns with Duyvejonck et al. report. 25 Given that phage propagation in this work was performed in a medium broth, the direct use of TSB as a storage solution may offer several potential advantages, such as the preservation of phage infectivity and stability, and potentially lowering downstream costs associated with media replacement or specific buffers.
Lytic activity assessment revealed no significant difference in bacterial growth in control groups of both SIF and TSB, indicating that the pathogenic bacteria could remain metabolically active in both conditions. Overall, phage lytic activity was consistent across different MOI values within the same experimental condition. The result of SIF conditions demonstrated the reduction of tested E. coli within 3 h. Our study was based on the premise of mixing phages with feed, ensuring that piglets would receive phages with every meal. In general, piglets are fed either on demand via feeding machines or in 3–4 meals per day. For scheduled meals, farmers typically provide food every 2–3 h, depending on farm management. However, the phages demonstrated more effective bacterial reduction in TSB at 30°C compared to SIF at 39°C for both individual phages and the phage cocktail. Given the prevalence of colibacillosis throughout the swine life cycle, the presence of pathogenic E. coli in the barn environment is a critical factor, as suitable humidity and low temperatures support their survival. 26 Enhancing hygienic and biosecurity protocols to reduce environmental pathogens has been suggested as a key strategy for preventing and controlling colibacillosis. 27 For practical on-farm applications, a short contact time is preferred for effective bacterial reduction. The observed reduction in the E. coli population at 30°C revealed the suitability of these phages as rapid sanitation agents. As previously noted, a temperature of 30°C aligns with the suitable range for swine housing, 18 while normal pig body temperature is ∼39°C.28,29 These results indicated the potential of these four phages to control pathogenic E. coli in both swine environments and within the animals themselves.
Conclusion
All four Escherichia phages displayed distinct morphologies and characteristics. Their infectivity against their host bacteria was confirmed across a wide temperature range relevant to swine environments and physiology. Furthermore, all phages demonstrated stability in various suitable solutions and buffers for long-term storage, maintaining their infectivity for at least 1 year. Both individual phages and their cocktail demonstrated significant lytic activity, effectively reducing host bacterial counts in both SIF at 39°C and TSB at 30°C. These results indicate the potential of each phage, alone and in combination, as biocontrol and biosanitation agents for swine production.
Authors’ Contributions
The study conception, design and supervision were contributed by R.N., N.T., S.S., and N.I. Formal analysis, methodology, and investigation were performed by N.I. Resources and materials was provided by N.T. and S.S. Writing—original draft and visualization were done by N.I. Funding acquisition and project administration were contributed by N.I. and R.N. All authors read, comment, and approved the final article.
Footnotes
Acknowledgments
The authors acknowledge Pattaraporn Sriprasong, Peekarn Poomirut, Nittaya Kunsanit, and Nachakrit Lugsanagul for their experimental support.
Author Disclosure Statement
No completing financial interests exist.
Funding Information
This project is funded by National Research Council of Thailand (NRCT) (Grant number. N41A660177).
Supplemental Material
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.