
Abstract
Introduction:
Burkholderia cepacia complex (BCC) infections pose a significant risk to people with cystic fibrosis and can result in high mortality rates. High levels of antimicrobial resistance within the BCC have increased interest in using bacteriophages (phages) as alternative therapeutics. However, most currently available BCC-targeting phages are temperate or lysogeny-capable, limiting their therapeutic use.
Methods:
To expand the pool of lytic Burkholderia phages, a phage targeting Burkholderia cenocepacia isolated from lake water was characterised by genome sequencing, electron microscopy, host range analysis, and bacterial killing assays performed at different temperatures.
Results:
The phage, designated as Burkholderia phage vB_BceS_Rocko (φRocko), possessed a 47 kb dsDNA circularly permuted genome and siphovirus morphology. At 37°C, antibacterial activity was modest and limited to actively growing bacterial cells. However, at higher phage densities and reduced temperatures (27°C and 30°C), pronounced bacterial killing was observed, particularly against stationary-phase cells. φRocko also exhibited activity against three additional BCC species, including both clinical and environmental isolates. Turbid plaques were produced by φRocko at cooler temperatures (23°C and 30°C), whereas plaques were largely absent at 37°C. Despite this, no lysogenic activity was detected at any temperature, suggesting that plaque morphology was not associated with lysogeny.
Conclusion:
The discovery of this novel phage adds another option to the limited arsenal of lytic phages active against BCC. Notably, φRocko’s ability to target stationary-phase bacteria may prove useful for therapeutic applications.
Introduction
The Burkholderia cepacia complex (BCC) is a group of >20 closely related Burkholderia species, that are ubiquitous in the environment and are causative agents of opportunistic infections, primarily in people with cystic fibrosis (CF; pwCF).1,2 In the United Kingdom, BCC is responsible for 1–4% of CF lung infections annually 3 and can cause severe and recurrent infections that may develop into the invasive “cepacia syndrome.” In particular, Burkholderia cenocepacia infections can result in disqualification from lung transplants due to increased mortality.4,5
Treatment of Burkholderia infections is complicated by high levels of intrinsic resistance to many antibiotics, including β-lactams, polymyxins, and aminoglycosides, 6 severely limiting the availability of therapeutic options. Bacteriophages (phages) and phage therapy represent an alternative approach for difficult-to-treat infections. 7 While these host-specific viruses have great potential, their application to Burkholderia infections is limited by the relatively small number of Burkholderia-targeting phages. Despite extensive environmental sampling efforts, just 10 lytic phages targeting the BCC have been reported,8,9 with the majority of phages being temperate phages, 9 capable of undergoing lysogeny through integration into the bacterial genome.8,9 Initial studies of lytic BCC-targeting phages noted that of the four lytic phages isolated, three had highly similar genomes, with one additional diverse phage, 10 suggesting a restricted but diverse phage population infecting BCC species. Similarly, a recently described lytic, flagellotropic jumbo phage exhibited a broad host range across multiple Burkholderia species, 11 reinforcing the notion that although the diversity of lytic Burkholderia-targeting phages is increasing, they are still rare. Temperate phages are considered less suitable for therapeutic use, as they may integrate into host chromosomes rather than entering lytic replication and can additionally transduce foreign DNA, that can increase bacterial resistance or virulence. 12 Advances in phage genetics have enabled the modification of temperate phages via the inactivation of integration-associated genes, 13 allowing their successful therapeutic use, including the treatment of Mycobacterium abscessus infections in pwCF.14,15 Because temperate phage-derived virions can also exhibit lytic activity against noncarrier strains without engineering, they are being investigated for therapeutic use, particularly against species like Burkholderia, for which lytic phages are scarce.9,16 Prophage-derived phages have demonstrated efficacy in preventing Burkholderia gladioli plant infections 17 and in controlling Clostridium difficile infections in vivo. 18 Additionally, studies propose inducing lysis in pathogenic strains through the direct activation of endogenous prophages as a therapeutic strategy.19,20 Nevertheless, obligately lytic phages currently remain the gold standard for therapeutics.
The successful use of phages to treat other CF lung pathogens has not yet translated into success against BCC lung infections, as the two reported attempts to treat Burkholderia infections in pwCF proved unsuccessful.21,22 In both instances, the patients had severe disease and had already undergone lung transplantation, and phage therapy was administered as a last resort. In each case, only a single lytic phage was available, and treatment was likely administered too late to enable patient recovery from advanced infection.21,22 Given the limited number of lytic Burkholderia phages and the challenges posed by slow-growing bacterial populations in chronic BCC infections, there remains a need to identify lytic phages capable of acting across all stages of infection. In this study, we isolated and characterized Burkholderia phage vB_BceS_φRocko, a novel lytic phage targeting B. cenocepacia K56-2, and examined its genomic features, host range, lifestyle, and behavior under different temperature conditions. In particular, we identified its unexpected activity against stationary-phase bacterial cells. Together, this study uncovers a rare example of a Burkholderia-targeting lytic phage with activity against metabolically restricted cells, which may prove useful for the treatment of chronic infections or biofilm control.
Materials and Methods
Microorganisms and culture conditions
All bacterial strains used in this study are listed in Supplementary Table S1. Burkholderia species were routinely cultured at 37°C, at 180 rpm in lysogeny broth (LB; Invitrogen, USA). For solid agar, 1% agar (VWR Chemicals, USA) was added to LB, and strains were grown at 37°C for 48 h. For phage propagation, experiments were conducted at 30°C, as is routine for many Burkholderia phages,10,23–25 and plates were incubated for 36 h. Plaque assays used 100 µL of stationary-phase bacterial cultures (∼5.6 × 109 colony forming units [CFU]/mL) and 100 µL of phage solution, with a 15-min adsorption period before 3 mL of 0.5% LB agar + 2 mM MgSO4 (Sigma, Japan) + 2 mM CaCl2 (Sigma, Japan) was added and poured over a solid agar base.
Phage isolation
For the isolation of a novel phage, 50 mL of water was taken from the lake bordering the Tocil Wood and Nature Reserve on the University of Warwick campus (GPS coordinates: 52.376148, −1.559658), and bacteria were removed using a 0.45 µM filter (Cytiva, USA). The water was combined with an equal volume of mid-exponential (OD600 nm 0.2) B. cenocepacia K56-2 wildtype or the aflagellated deletion mutant (ΔfliF) in 2× LB supplemented with 2 mM CaCl2 and 2 mM MgSO4, and the samples were incubated for 20 h at 30°C, at 170 rpm. Following incubation, bacterial cells were pelleted, and the supernatant was filtered through a 0.22 µM filter (Merck, Germany), before being plated in soft agar overlay plates containing B. cenocepacia K56-2 wildtype or ΔfliF. Following the initial isolation, all phages were routinely cultured on the wildtype strain. Plaques were cored using a P1000 tip, diluted in SM buffer (100 mM NaCl, 8 mM MgSO4, 10 mM Tris pH 7.5), and plated in soft agar overlay plates. Phages were isolated sequentially five times from the initial lysate. For the generation of phage stocks, completely lysed phage overlay plates (containing <1 × 105 plaque forming units [PFU/mL]) were flooded with 5 mL SM buffer and were incubated at room temperature for 30 min. The soft agar and SM buffer were collected and agitated by vortexing to release any additional phage, and solids were pelleted at 9,000 g before the supernatant was filtered through a 0.22 µM filter (Cytiva, USA). Phages were routinely stored at 4°C.
Genome assembly and analysis
Phage DNA was purified from >1 × 109 PFU/mL by phenol:chloroform extraction as outlined in the PCI/SDS DNA Extraction protocol at PhagesDB. 26 The DNA was submitted to MicrobesNG (United Kingdom) for Illumina Short Read paired-end sequencing. Reads were trimmed with Cutadapt v0.6.10, 27 down-sampled to 150× coverage with rasusa v2.0.0 28 and assembled with SPAdes genome assembler v3.15.5. 29 Genomes were annotated with Pharokka v1.7.1, 30 encompassing tools PHANOTATE for coding sequence (CDS) prediction, 31 tRNAscan-SE 2.0 for tRNA prediction, 32 and CRT for CRISPR prediction. 33 Functional annotation, including the detection of toxin, virulence, or antimicrobial resistance genes, was achieved through matching each CDS to the VFDB, 34 PHROG, 35 and CARD 36 databases using MMseqs2 37 and PyHMMER. 38 Contigs were matched to their closest relatives in the INPHARED database 39 using mash. 40 Genome plots were generated with pyCirclize. 41 Taxonomy was assigned using TaxMyPhage. 42 Genomes were scrutinized for lysogeny genes using PhageLeads. 43 Sequencing reads were indexed and mapped to the reference genome using SAMtools, 44 and the nucleic acid read mapping was visualized using Integrative Genome Viewer (IGV). 45 Single-nucleotide polymorphisms (SNPs) were identified using Snippy. 46 Genome comparisons of the amino acid sequences from annotated genes were made to the following phage genomes: vB_BceS_KL1, 23 vB_PaeS_SCH_Ab26, 47 vB_Pae-Kakheti25, 48 and vB_Pae_PA73 49 using Clinker.23,47–50
Plaque morphology
Images of plaques following incubation at 30°C for 36 h were acquired using a CanoScan LiDE 400 scanner (Canon, Japan). Plaque size was determined using ImageJ. 51 For comparison of activity across different temperatures, 5 µL of phage lysate at 1 × 107, 1 × 105 and 1 × 103 PFU/mL was plated onto a wildtype B. cenocepacia K56-2 lawn, and the plates were incubated at 23°C, 30°C, and 37°C for 48 h.
Transmission electron microscopy
Phages were purified from bacterial lysate using Vivaspin 20 centrifugal concentrator 300 kDa MWCO columns (Cytiva, USA), and the buffer was exchanged into SM buffer. A 5 µL drop of purified phage was applied to a glow-discharged carbon/formvar-coated 300 mesh copper TEM grid (EM Resolutions, United Kingdom). The grids were stained with 2% uranyl acetate for 1 min and were imaged on the JEOL 2100 Plus electron microscope (JEOL, Japan) with a Gatan ultrascan camera (Gatan, USA).
Lysogeny assay
Lysogeny was screened following the protocol outlined in Gordillo Altamirano and Barr 52 and using the Lysogeny Experiments protocol at PhagesDB. 26 Briefly, φRocko at concentrations of 1010, 109, 108, and 107 PFU/mL was spotted onto a soft agar overlay with wildtype B. cenocepacia K56-2 and was incubated for 48 h at 23°C, 30°C, and 37°C, respectively. These temperatures were maintained during subsequent culturing, except that all plaque soft agar overlay plates were all cultured at 30°C. Following incubation, bacteria were collected from the mesa and streaked onto a new plate. Sixteen single colonies per temperature were collected and were streaked onto a fresh LB agar plate and onto a soft agar overlay with wildtype B. cenocepacia K56-2. To check for dormant phages, all colonies, regardless of activity on the overlay plate, were streaked onto a fresh LB agar plate, and a single colony subsequently was tested for phage activity on a new overlay plate, and lysate was spotted onto another overlay plate. Bacterial cells from the culture that were used to make the lysate were tested for the presence of φRocko by polymerase chain reaction (PCR) with the 2× PCRBIO Taq Mix Red (PCR Biosystems, England) as per manufacturer’s instructions and with primers that bound to the endolysin gene (Fwd GAACCAATCAAAACCCCGACGCCG; Rev GATTGCCCACATTATGCCGCCCC). The products were analyzed on a 1% agarose gel. The test for lysogeny was repeated three times at 30°C and once at 23°C and 37°C.
Phage virion thermostability assay
Phage virion thermostability was tested by storing 8 × 109 PFU/mL of phage in 100 µL SM buffer in triplicate at 4°C, 23°C, 30°C, and 37°C for 48 h. Following the incubation period, the phages were diluted in SM buffer, and 10 µL was plated in technical triplicate onto soft agar overlay plate containing stationary phase culture, and was incubated at 30°C for 36 h. Recovered phages were quantified and analyzed for differences relative to the 4°C control sample.
Phage killing kinetics assay
Overnight cultures of wildtype B. cenocepacia K56-2 (prepared in biological triplicate) were diluted 1/100 into prewarmed media and were grown to mid-exponential (OD600 nm = 0.3–0.4) phase. Cultures were then diluted to an OD600 nm of 0.001 (4 × 106 CFU/mL) in LB + 2 mM MgSO4 + 2 mM CaCl2 and were combined with ɸRocko at an initial multiplicity of infection (MOI) of 250 and subsequent 10-fold dilutions, each in technical triplicate, to a final volume of 200 µL in a 96-well plate covered with a Breathe-Easy® membrane (Sigma, Japan). Plates were incubated in a Spark Multimode Microplate reader (Tecan, Switzerland) for 48 h at 27°C, 30°C, and 37°C with constant aeration (180 rpm). Measurements were taken every 20 min at OD600 nm. The area under the curve (AUC) was calculated in R
53
using gcplyr.
54
The virulence index (Vi) and growth reduction coefficient (GRC) were determined using the equation 1–
Host range analysis and efficiency of plating
Concentrated phage lysates (>1 × 1010 PFU/mL) were spotted (5 µL) onto soft agar overlay plates containing each of the Burkholderia species outlined in Supplementary Table S1 and activity was observed after 48 h at 30°C. Phages were quantified by spotting 10 µL of phage lysate diluted in SM buffer in technical triplicate onto a soft agar overlay plate containing stationary phase bacterial culture and incubated at 30°C for 48 h. Plaques were quantified and the EOP relative to the B. cenocepacia K56-2 wildtype strain was determined using the following equation:
Visualization and statistical analysis
Plots were generated in R 53 using ggplot2. 55 For comparison between two samples in the virion thermostability and host range EOP assays, a Mann–Whitney U test was performed using R to determine any statistically significant differences. For comparisons of changes in the phage kill curve, a Kruskal–Wallis test was performed using R.
Results
Phage isolation and characterisation
Six phages were isolated from the lake bordering the Tocil Wood and Nature Reserve on the University of Warwick campus against two B. cenocepacia K56-2 host strains (wildtype and ΔfliF): four using wildtype B. cenocepacia K56-2 (phages 1–4) and two using B. cenocepacia ΔfliF (phages 7 and 8). Both strains recovered similar numbers of plaques that had a shared morphology.
Short read sequencing of the dsDNA genomes confirmed that the phages isolated on the same hosts were 100% identical at the nucleotide level, while phages isolated on different hosts shared 99.18% nucleic acid identity.
A representative phage was selected from each of the identical sets for further analysis (phages 1 and 8 were isolated against wildtype and ΔfliF, respectively). Transmission electron microscopy revealed that both phages displayed siphovirus morphology with long flexible tails (184 ± 4 nm) and heads of 49 ± 4 nm (Fig. 1).
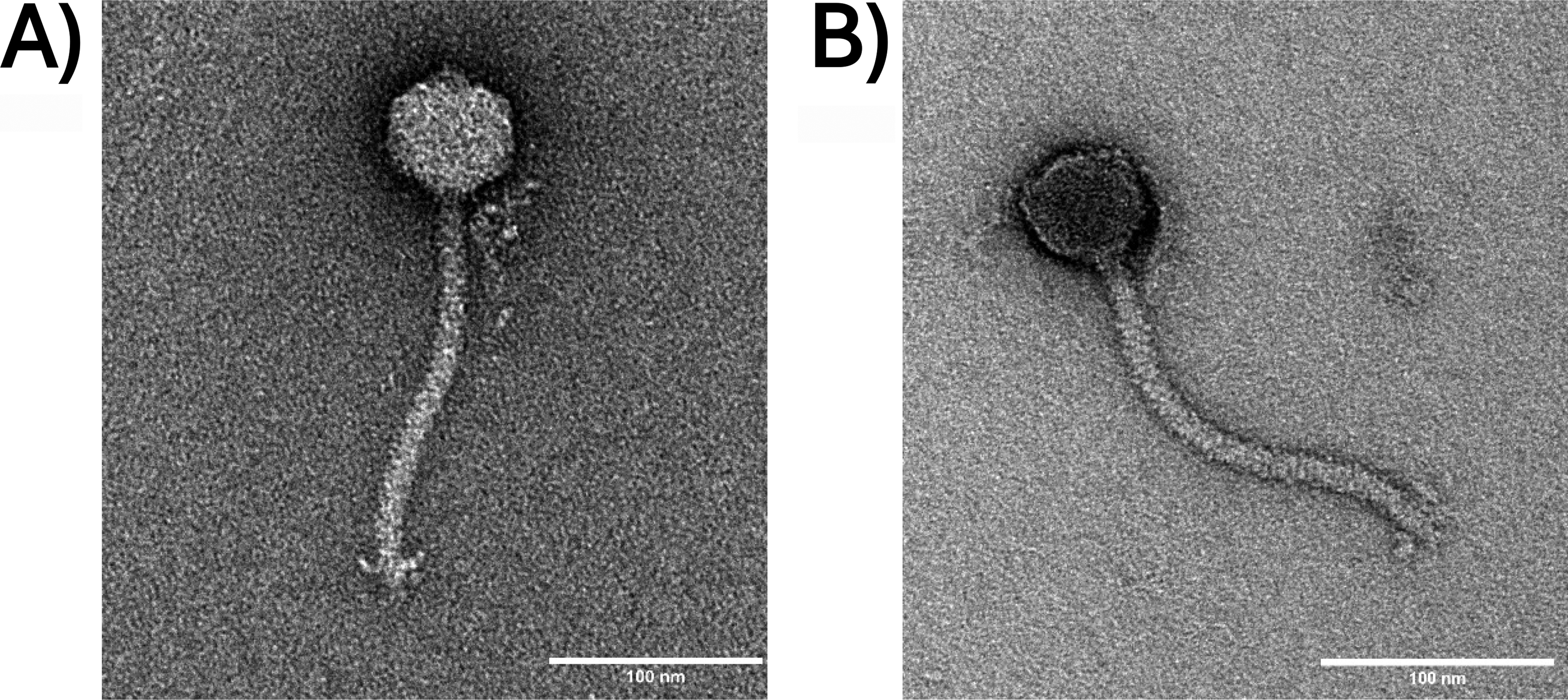
Transmission electron micrographs of φRocko
Phage genome analysis, annotation, and organisation
Genome analysis showed that phage 1 and phage 8 had genome sizes of 43,738 bp and 43,737 bp respectively and shared an average GC content of 55%. A total of 355 SNPs were detected between the genomes (Supplementary Table S2). These SNPs were likely present during the initial isolation, as it is unlikely that they arose independently during the five rounds of purification. Overall, the genomic similarity confirms that these isolates represent two variants of the same phage: Burkholderia phage vB_BceS_Rocko (φRocko; accession ERS25458009) represented by phage 1 and the closely related variant Burkholderia phage vB_BceS_Rocko.8 (φRocko.8; accession ERS25458010) represented by phage 8.
The SNPs were distributed across 41 of the predicted 58 ORFs in φRocko (Fig. 2), resulting in one less ORF in φRocko than in φRocko.8. Five ORFs did not align between the two genomes (Supplementary Table S3) and φRocko contained three fewer structural genes than φRocko.8. Notably, the major capsid protein was annotated in φRocko.8 (RCKO8_CDS_10 Supplementary Fig. S1), but not in the φRocko sequence. Given the presence of seven missense mutations and one stop mutation in the φRocko.8 region relative to φRocko, the absence of the major capsid protein in φRocko is likely due to an annotation error, rather than a true biological difference. Both genomes encoded a MazG protein, a pyrophosphohydrolase, which has been implicated in viral metabolism 56 and neither genome encoded any antimicrobial resistance (AMR) or toxin genes as determined using Pharokka. Furthermore, both genomes lacked any lysogeny markers, including transposases, excisionases and recombinases and PhageLeads 43 analysis did not identify any temperate markers or virulence-associated genes. As φRocko and φRocko.8 showed no detectable differences in activity, all subsequent analyses focussed exclusively on φRocko.
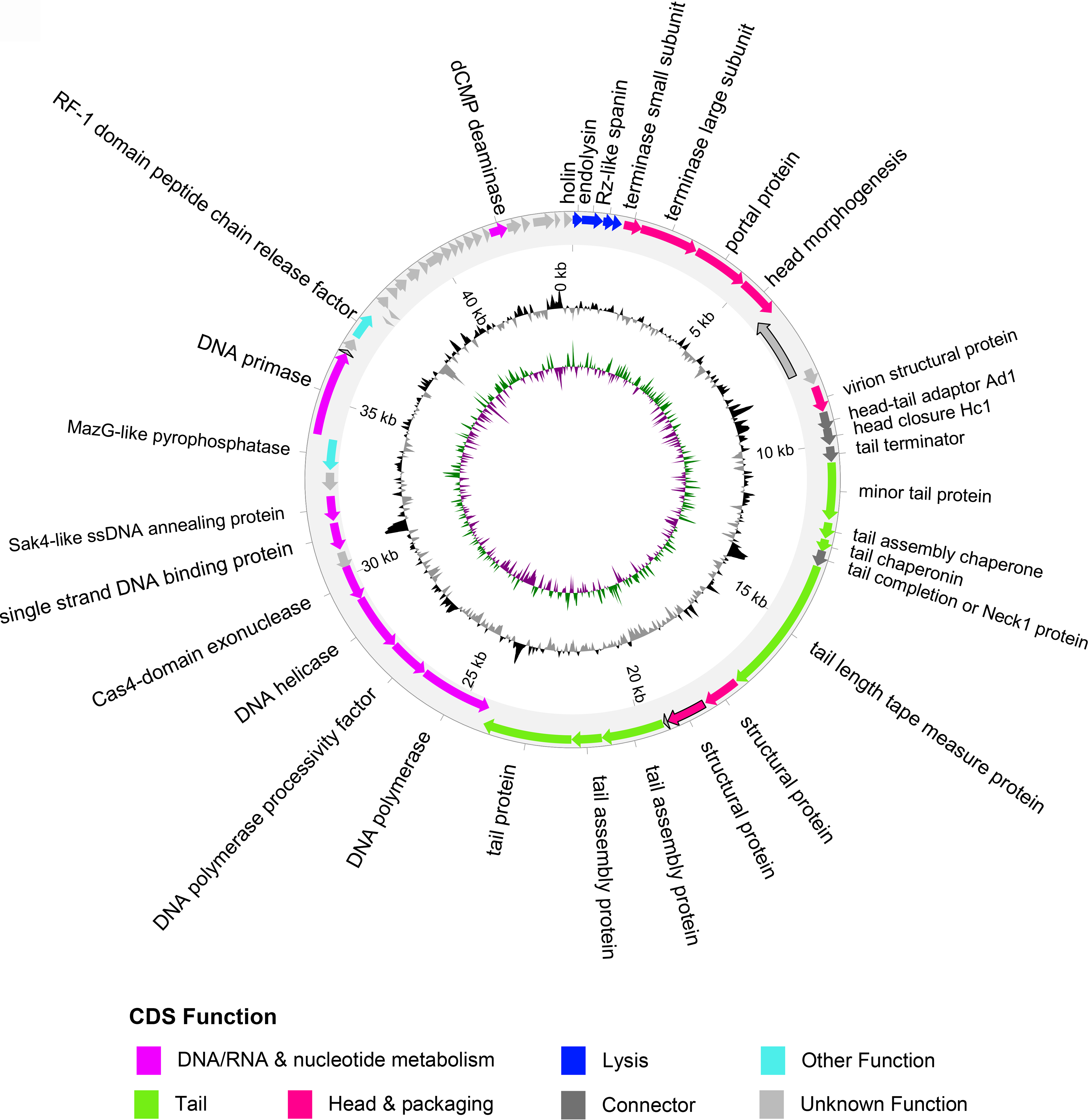
Graphical representation of the φRocko genome. The outer ring shows the CDSs, with colors corresponding to their predicted functions (legend underneath). Genes outlined in black highlight the CDSs that differ between the two variants of φRocko. The middle ring shows the GC content: black represents positive GC content and gray represents negative GC content. The inner ring denotes the GC skew: where purple represents positive GC skew and green represents negative GC skew. CDSs, coding sequences.
To determine the DNA packing strategy employed by the phages, the read alignment patterns of all six phages were scrutinized in IGV. For all phages, coverage was consistent, with no sudden drops or peaks. Furthermore, each phage displayed a different origin and wrap-around ends were identified in all sequences, suggesting that the φRocko genome is circularly permuted 57 with random termini. As such, all φRocko sequences were reordered to align with the holin gene. This provided a consistent start position and facilitated comparison with related phages identified through TaxMyPhage, 42 which classified φRocko as a new phage species within the Kilunavirus genus, based on the ICTV taxonomy and the average nucleotide identity (ANI) threshold matching this genus. φRocko shared 70.6% similarity with the other Burkholderia Kilunavirus phage vB_BceS_KL1 (KL1) 23 and similarity with three Pseudomonas aeruginosa targeting phages (Fig. 3): Ab26 (55.5%), 73 (56.2%) and Kakheti25 (57%). Although PhageLeads did not identify any temperate lifestyle genes in the φRocko genome, the software detected in the KL1 phage genome predicted a cro gene, (Fig. 3: red gene), which regulates phage genome integration.58,59 φRocko and the Pseudomonas phages each contain homologs of this gene, but PhageLeads did not identify these homologs as cro genes in any of the other phage genomes.
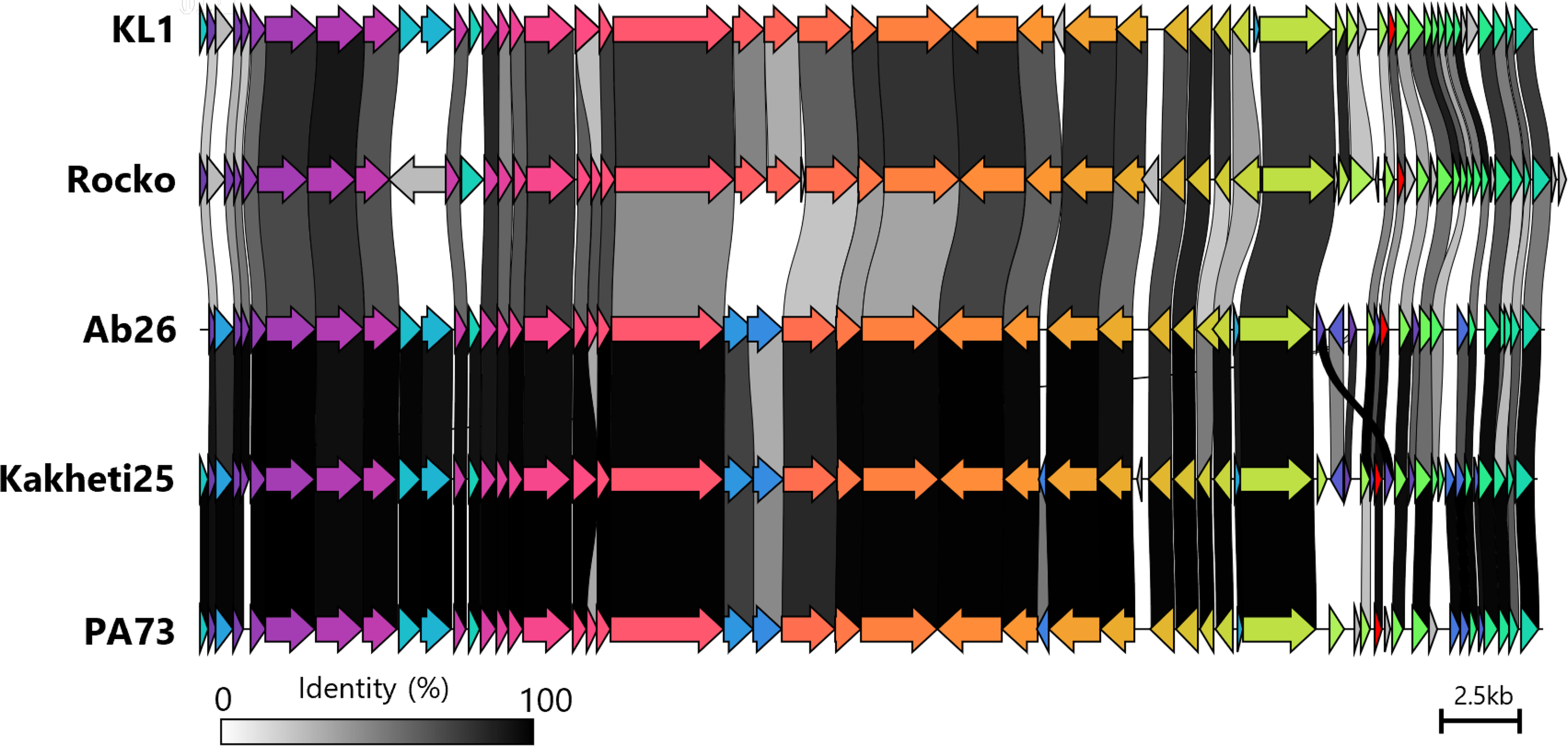
Genomic comparison of the novel φRocko and its closest relatives (Burkholderia phage KL1 and Pseudomonas phages Ab26, Kakheti25 and PA73). Coding sequences are color coded based on shared homology, and links between phages are colored to reflect their shared identity (legend on bottom left). The cro gene identified in KL1 and its homologs in the other phages are colored in red. Scale bar represents 2.5 kb.
Phage plaque morphology, host receptor analysis, and host range
φRocko formed plaques ranging from 0.3 mm to 3 mm in diameter with the larger plaques having an uneven shape and jagged edges relative to the smaller plaques that lacked this uneven shape (Fig. 4A). Plaques began to appear after 24 h and were fully formed after 36 h. As φRocko produces turbid plaques at 30°C, we looked at the formation of lytic zones at 23°C and 37°C (Fig. 4B). The plaques at 23°C were virtually indistinguishable from those at 30°C, except for a slightly larger hazy ring at the periphery of the zone. At 37°C, the lytic zones were substantially reduced with activity only visible at very high phage concentrations (>108 PFU/mL spots) and individual plaques could not be distinguished (data not shown). This reduction was independent of the phage virion thermostability at 37°C, as the concentration of φRocko did not significantly reduce over the same time period compared to the 4°C control (Fig. 4C).
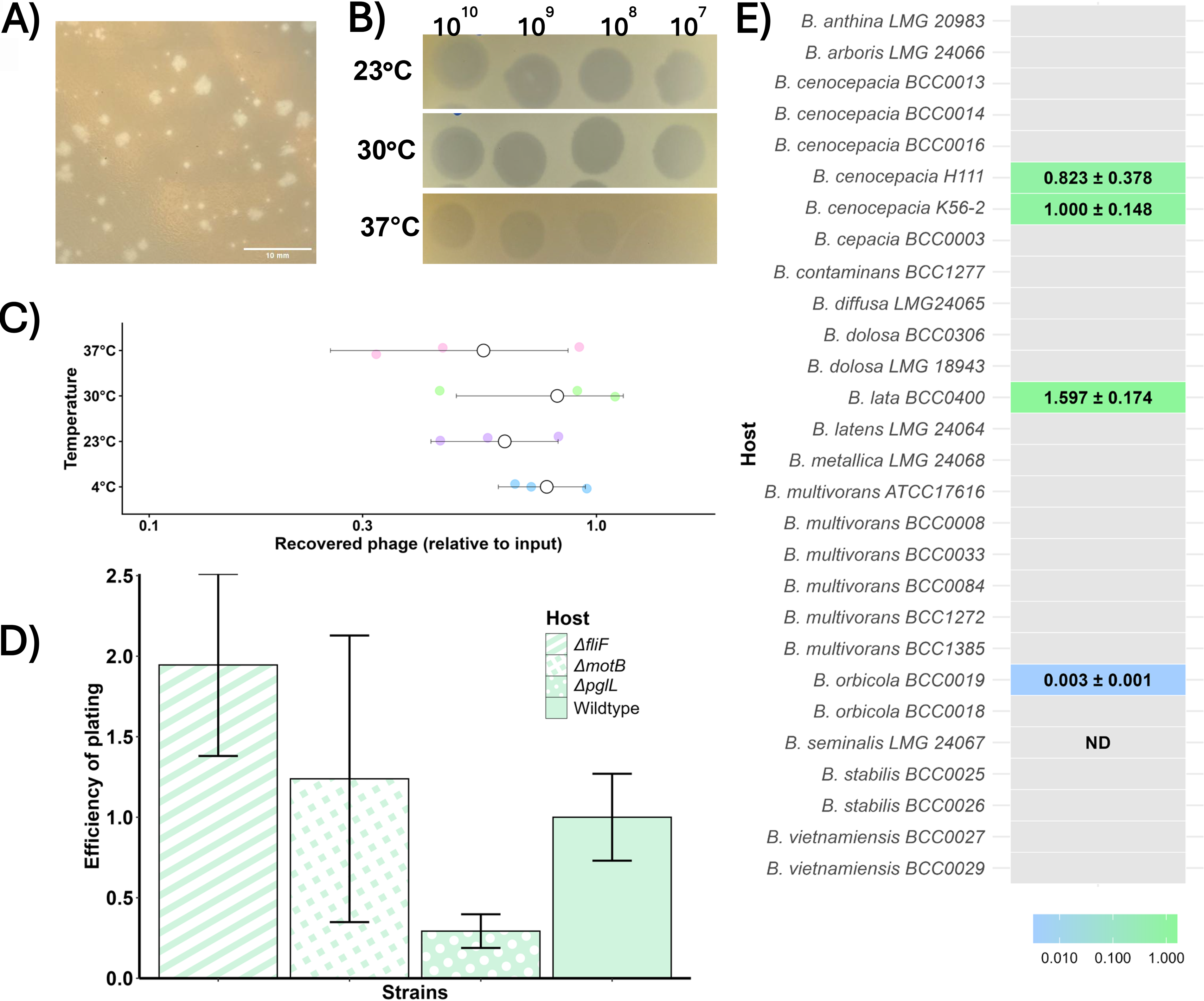
Plaque morphology and host-range analysis of φRocko.
We screened φRocko against a small selection of B. cenocepacia K56-2 mutant strains (ΔfliF—lacks a flagellum, ΔmotB—paralysed flagellum and ΔpglL—glycosylation deficient; Fig. 4D) in order to identify the phage receptor. No significant differences in infectivity were observed, suggesting that φRocko does not rely on either the flagellum or glycoproteins for infection.
To understand the host range, φRocko was screened against a panel of BCC strains and species (Supplementary Table S1; 5 B. cenocepacia strains and 14 different BCC species including type strains). φRocko showed quantifiable activity against two B. cenocepacia isolates, an environmental isolate of B. lata, the single representative of this species in the test panel and one of the two B. orbicola clinical CF isolates tested (Fig. 4E). Additional activity was observed against the B. seminalis isolate (a clinical CF isolate) at high phage titres (109–1010 PFU/mL), but individual plaques were not observed under the conditions tested. φRocko infected B. cenocepacia H111 at a rate comparable to the isolation strain B. cenocepacia K56-2 and the EOP of B. lata BCC400 was 1.5-fold higher, although this was not statistically significant (Fig. 4E). The EOP for B. orbicola BCC19 was very low compared to the host strain. For all species, the φRocko plaques were turbid and consistent in size and shape to the plaques formed on B. cenocepacia K56-2. Overall, the analysis from this panel suggests that the host range for φRocko may be limited.
Temperature-dependent phage killing dynamics and assessment of lysogeny
To understand how temperature influences φRocko activity, we analyzed the effect of the phage on bacterial growth in liquid culture across six MOIs at 27°C (Fig. 5A), 30°C (Fig. 5B), and 37°C (Fig. 5C). The strongest effect was observed at 27°C, particularly at the highest phage density (MOI 250), which caused a slight delay in the stationary phase. This corresponded to a ∼0.38 local virulence index score (Vi, Fig. 5D; purple). The most striking effect at this temperature occurred during the stationary phase, where high MOIs (250 and 25) resulted in a decrease in bacterial density after 20 h. Although the growth was partially recovered, these samples never reached the density of the uninfected control, with the overall differences in growth contributing to significant p values (1.8 × 10−44 for MOI 250 and 6.1 × 10−32 for MOI 25). This is reflected in the elevated growth reduction coefficient (GRC, Fig. 5E, purple) with scores of 0.52 for MOI 250 and 0.23 for MOI 25. At lower MOIs (2.5–0.0025), φRocko caused statistically significant reductions in growth, which are reflected in the Vi and GRC values of <0.1.
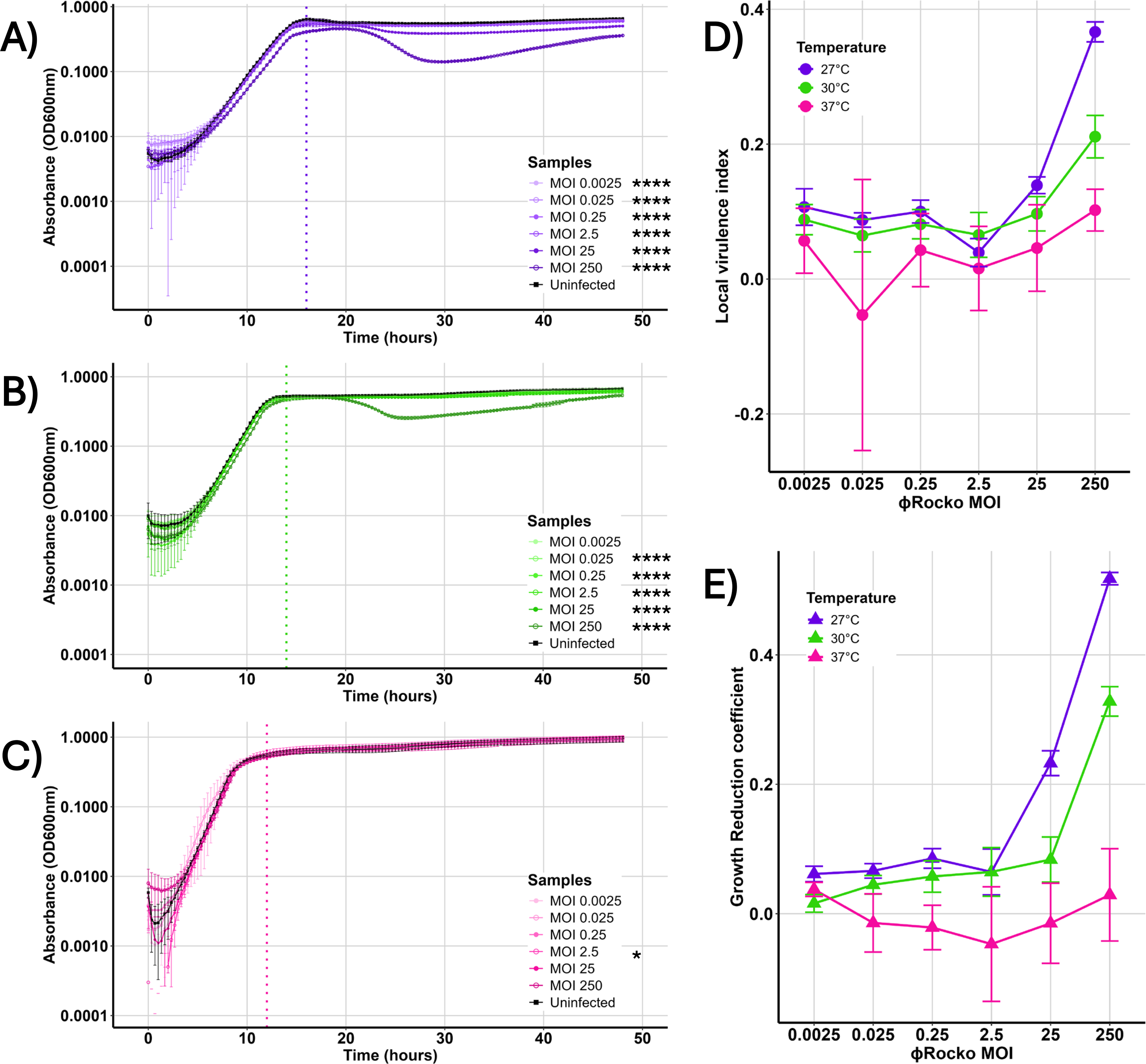
Effect of temperature on φRocko activity. Growth of B. cenocepacia K56-2 over 48 h at
Raising the temperature to 30°C attenuated the overall effect of φRocko on bacterial growth. At the highest phage density (MOI = 250), effect was observed on both the exponential and stationary phases (p value 6.5 × 10−47), but the magnitude was reduced relative to 27°C, with intermediate Vi and GRC scores (Vi = 0.21, GRC = 0.32; Fig. 5D–E, green). For lower density samples at 30°C, the Vi and GRC values were stabilized at ∼0.1, indicating a minimal biological impact despite statistically significant differences in growth. At 37°C, φRocko showed very little activity across all MOIs, with the growth curves closely aligning with the uninfected control. The only significant difference was identified at the MOI 2.5 sample (p value 0.01), which may reflect differences in the lag phase rather than phage-mediated activity. Consistent with this, the Vi and GRC scores were < 0.06, dropping to <0 for the GRC at MOIs = 25–0.025, reflecting lack of phage activity (Fig. 5D–E, pink). Notably, at MOI 250, the Vi was 0.1, despite the GRC being 0.03, showing that φRocko had a small effect on exponential phase bacteria at 37°C, but had no effect on stationary phase cells. Collectively, these data suggest that φRocko activity is strongly temperature dependent, with higher phage densities having a more profound effect on stationary phase cells.
As the phage killing kinetic assays suggest, φRocko was never able to clear cultures, yet was capable of producing high titres in liquid media (>1 × 1010 PFU/mL). This pattern of minimal bacterial killing aligns with the turbid plaques formed by the phage, yet contrasts with the observation of high phage productivity, suggesting that φRocko may use a passive phage release mechanism to largely avoid cell death. Such passive phage release mechanisms are characteristic of filamentous phages and have additionally been observed in at least one prophage infecting Lactobacillus lactis 60 but these have not been previously shown for any dsDNA phages infecting Burkholderia. Although the investigation of such a mechanism was beyond the scope of this study, the concept merits exploration in future work. To determine whether this behavior could reflect a temperate lifestyle, we performed lysogeny assays at 23°C, 30°C, and 37°C. No phage activity was detected in the subsequent bacterial isolates in the first or third generations at any temperature and PCR screening did not identify φRocko in the bacterial genome in any sample at the conclusion of the assay (data not shown). Together, these results suggest that φRocko is not temperate and cannot undergo lysogeny.
Discussion
Burkholderia infections remain amongst the most difficult to treat infections in pwCF due to their intrinsic drug resistance. While phage therapy offers a therapeutic alternative, a lack of lytic phages active against BCC hinders this advance. We have identified two variants of a novel lytic phage, φRocko and φRocko.8, adding to the 10 lytic phages previously characterized against this pathogen. 8 φRocko exhibited temperature-sensitive infection dynamics with activity favored at cooler temperatures (23–30°C) and could infect stationary phase cells at high phage densities. φRocko also displayed a limited host range, infecting <20% of the strains tested. This restricted range is consistent with the host ranges reported for other lytic phages targeting the BCC8,9 and may represent an intrinsic feature of phages infecting this genus. However, it is also likely influenced by the limited number of characterized lytic phages and the incomplete assessment of their host ranges. These features highlight the constraints and potential of φRocko as a therapeutic agent.
To assess the host-driven differences between the two φRocko variants, we compared the φRocko and φRocko.8 genomes and identified 355 SNPs that likely arose during the initial isolation on the B. cenocepacia wildtype and ΔfliF strains. Despite these genomic changes altering the predicted ORF content and many of the SNPs occurring in structural genes, φRocko.8 displayed indistinguishable morphology. These alterations may have contributed to functional differences between the variants and this warrants further investigation. Similarly, efforts to identify the phage host receptor were unsuccessful. A targeted screen of mutants was unable to identify any statistically significant differences between the mutants and the wildtype strain. Therefore, a more extensive mutant library is required to identify the receptor required for φRocko infection.
φRocko was most closely related to another B. cenocepacia-targeting phage KL1, a lysogen-capable phage isolated from sewage. 23 KL1 produces turbid plaques and exhibits delayed plaque formation, 23 characteristics shared with φRocko and also encodes a predicted cro gene. Although φRocko encodes a protein sharing 76% amino acid identity with KL1 Cro, this homolog was not flagged as cro in φRocko, nor in related Pseudomonas phages, including Kakheti25, a lytic phage that forms clear plaques. 48 Consistent with a strictly lytic lifestyle, φRocko lacks genes associated with lysogeny, including an integrase protein, which is typically >250 amino acids, 61 substantially larger than the 20+ hypothetical proteins encoded in the φRocko genome. KL1 has previously been shown to be retained in 22% of first-generation bacterial cells, following infection with the phage. 62 However, retention of KL1 was strain specific and while the presence of the phage genome was confirmed by PCR, its integration into the genome was not tested and could represent pseudolysogeny—a form of episomal replication—rather than true integration. While φRocko was screened for lysogeny in the third generation, it is possible that a small proportion of φRocko was transiently maintained in a limited number of cells and escaped detection. Such temporary maintenance of φRocko via pseudolysogeny could contribute to the delayed killing of stationary phase cells. Nevertheless, the absence of lysogeny-associated genes and the lack of detected lysogens, supports the conclusion that φRocko is functionally lytic.
The ability of φRocko to impact stationary phase bacterial cells was most pronounced at 27°C, where high phage densities (MOIs 250 and 25) resulted in a measurable reduction in cell density during the stationary phase. The GRC captures this impact best, integrating the effects across the full curve, whereas the Vi only captures the disruption to exponential growth. Although all φRocko-infected samples at 27°C temperature differed significantly from the uninfected control, the effects at MOIs ≤ 2.5 were biologically minimal, as reflected by the Vi and GRC values below 0.1. At 30°C, a similar but attenuated pattern of phage activity was observed, indicating that the modest temperature increase reduced the efficacy of φRocko infection. Although impact to both the exponential and stationary growth phase were observed at the highest phage density, the magnitude of this effect was reduced relative to the 27°C across both the Vi and GRC. Once the phage density dropped below MOI 25 at 30°C, the Vi and GRC values remained constant, sitting around 0.1, regardless of the reducing phage concentration. Given the small p-values across these samples, it is likely that this very slight impact on growth is phage mediated, which may lend the phage to being useful in low densities in the presence of additional phages. At 37°C, a slight dose-dependent response can be observed in the Vi values between MOIs 250 and 25. However, this was not shared in the GRC, where all samples returned values close to 0, showing the lack of impact of φRocko at 37°C outside a minor effect in the exponential phase. While the overall effect of φRocko is still modest compared to many phages that can decimate a population, the killing of stationary phase cells is an important and less frequently observed feature.63–65 Stationary phase cells are metabolically distinct from actively growing cells and often feature less receptors compared to their exponential counterparts, which for many phages makes them unsuitable hosts. However, the stationary phase may be more reflective of how the bacteria exist in the general environment, where the ideal growth conditions are not always met. 66 The metabolic restriction of stationary phase is also biologically similar in a number of ways to a biofilm67,68 and it might therefore be possible that these stationary phase active phages like φRocko, could have antibiofilm properties. Another factor that may influence the differences in infectivity across temperatures and growth phases, is φRocko’s ability to adsorb to host cells. Cell membrane composition changes between growth phases 69 and in different temperatures, 70 altering phage receptor availability and affecting a phage’s ability to bind to host cells. Investigation into φRocko’s ability to adsorb under these different conditions would allow a deeper understanding of its behavior across temperatures and growth phases and will provide key information for future studies.
The increased activity at lower temperatures observed in φRocko has been previously observed in numerous low-temperature active phages,71–73 including those isolated against Pseudomonas and Escherichia.74–77 These phages, which were isolated from a range of sources, demonstrate significantly reduced or completely ablated activity at higher temperatures and have been of interest for their applications in food manufacturing71,74,78 and biofilm control, 73 due to their activity in these environmental conditions.79,80 The molecular mechanisms that control for low temperature activity are unknown and thus, we cannot discriminate any genes in the φRocko genome that would corroborate the low-temperature preference we have observed, although the bacterial metabolism and overall phage virion thermostability is likely to be involved. 75 As some studies have shown that certain BCC clinical isolates are indistinguishable from environmental isolates,81,82 it is possible that φRocko is adapted for activity on environmental isolates and that there is enough similarity to the clinical isolates that we were able to observe some activity. Given its lack of activity at 37°C, φRocko in its current state would not be suitable for phage therapy of clinical BCC infections in the absence of other phages. However, in vitro evolution could be applied to increase φRocko’s temperature range83,84 and if achieved, the phage’s predisposition to target stationary phase bacteria could be a powerful tool in the treatment of biofilms85,86 or chronic infections.
Conclusion
This study describes the isolation and characterisation of φRocko, a novel phage with activity across four BCC species and a genomic profile consistent with therapeutic safety requirements. Its capacity to infect stationary phase cells at 27°C and 30°C is a particularly noteworthy finding, as this phenotype is relatively uncommon and may reflect adaptation to biofilm or environmental settings, where bacteria exist predominantly in a slow-growing state.
The principal limitation of φRocko is its failure to demonstrate significant activity at 37°C, which prevents its use as a standalone therapeutic for human infections. The mechanistic basis of this temperature sensitivity remains unclear and merits further investigation, along with broader testing against a wider panel of clinical BCC isolates to fully define its host range and resistance potential.
Despite this, φRocko’s stationary phase activity offers functional complementarity to phages that target actively dividing cells, making it a strong candidate for inclusion in phage cocktails targeting chronic BCC infections, such as those seen in cystic fibrosis patients. Future work focused on a phage cocktail design and the molecular basis of temperature-dependent phage infectivity, will be key to translating these findings towards clinical applications.
Data Availability
All genome sequencing data generated in this study are available under the project PRJEB95892 accessions ERS25458009 for Burkholderia phage vB_BceS_Rocko and ERS25458010 for Burkholderia phage vB_BceS_Rocko.8.
Author Disclosure Statement
No competing financial interests exist.
Supplemental Material
sj-docx-1-pta-10.1177_26416549261465080 — Supplemental material for Novel Burkholderia-Targeting Lytic Phage Rocko Preferentially Kills Stationary Phase Bacterial Cells
Supplemental material, sj-docx-1-pta-10.1177_26416549261465080 for Novel Burkholderia-Targeting Lytic Phage Rocko Preferentially Kills Stationary Phase Bacterial Cells by Jessica M. Lewis, Olivia Dutton, Tanaz Mistry, and Antonia P. Sagona
Supplemental Material
sj-csv-2-pta-10.1177_26416549261465080 — Supplemental material for Novel Burkholderia-Targeting Lytic Phage Rocko Preferentially Kills Stationary Phase Bacterial Cells
Supplemental material, sj-csv-2-pta-10.1177_26416549261465080 for Novel Burkholderia-Targeting Lytic Phage Rocko Preferentially Kills Stationary Phase Bacterial Cells by Jessica M. Lewis, Olivia Dutton, Tanaz Mistry, and Antonia P. Sagona
Footnotes
Funding Information
The authors acknowledge funding from the ![]() ). The authors would like to thank the University of Warwick, School of Life Sciences Media Preparation team for their assistance preparing the media used in this study and Dr. Saskia Bakker from the Advanced Bioimaging RTP at University of Warwick for her assistance with the transmission electron microscopy.
). The authors would like to thank the University of Warwick, School of Life Sciences Media Preparation team for their assistance preparing the media used in this study and Dr. Saskia Bakker from the Advanced Bioimaging RTP at University of Warwick for her assistance with the transmission electron microscopy.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.