
Abstract
Poor sleep and psychological symptoms (depressive or anxiety symptoms) are common following concussion. Saliva microRNAs (miRNAs) are promising concussion biomarkers based on associations with the presence of a concussion in prior work. The purpose of this study was to identify saliva miRNAs that differ between participants with and without concussion and show correlations with sleep and/or psychological symptoms. We analyzed salivary miRNAs and concussion symptom data from n = 272 concussion cases (median: 2.5 days postinjury, interquartile range: 2.41–10.65) and n = 153 uninjured controls. Concussion symptoms were quantified with the Post-Concussion Symptom Scale (PCSS). miRNAs were quantified with high-throughput small RNA sequencing. Spearman rank correlations were used to evaluate the association between miRNAs and PCSS sleep (“Drowsiness,” “Trouble falling asleep”) and PCSS psychological (“Feeling More Emotional,” “Irritability,” “Sadness,” and “Nervousness”) symptoms. Wilcoxon rank sum tests evaluated differences in miRNAs between the concussion versus the uninjured controls. All tests were evaluated at the Benjamini-Hochberg adjusted alpha level of 0.05. We identified 14 miRNAs that were correlated with sleep symptoms (n = 5 positively and n = 9 negatively correlated), 28 with psychological symptoms (n = 12 positively and n = 16 negatively correlated), and 4 miRNAs with both (miR-361-5p, miR-4510, miR-193a-3p, miR-3074-5p). Gene targets for miRNAs correlated with both sleep and psychological symptoms were significantly enriched within “spinocerebellar ataxia,” “dopaminergic synapse,” “axon guidance,” and “circadian entrainment” pathways. Of the four miRNAs correlated with both psychological and sleep symptoms, miR-4510 and miR-193a-3p were also differentially expressed between the concussion cases and the uninjured controls. miR-4510 and miR-193a-3p were differentially expressed in individuals with concussion and were correlated with both sleep and psychological symptoms. Additional research is needed to confirm the temporal relationship between alterations in these miRNAs, concussion occurrence, and onset of sleep and/or psychological symptoms.
Introduction
Concussion represents a complex neurological injury that can disrupt multiple domains of functioning. 1 Symptoms, while nonspecific to concussion alone, are grouped into various domains including cognitive, physical, emotional, or sleep-related symptoms2,3 that may present acutely postinjury, or develop in the subsequent days to weeks postinjury.2,3 Although most individuals return to usual activities within 2–4 weeks,4,5 up to 35% of individuals will develop persisting symptoms (i.e., symptoms lasting >28 days) after concussion (PSaC).6,7 The development of secondary complications including PSaC is a major concern as these complications can lead to additional time loss from athletics, decreased academic engagement, decreased peer interactions, and increased health care utilization.8,9
Sleep and psychological symptoms are common and interrelated secondary symptoms following concussion.10,11 The overlap between sleep disturbances and psychological symptoms extends beyond concussion, as the pathophysiological links between sleep and psychiatric disorders such as anxiety and major depressive disorder (MDD) have also been explored.12,13 While the direct pathophysiology between sleep and psychiatric disorders remains unclear, several studies suggest there is a significant interplay between sleep disturbances and the exacerbations of psychiatric disorders.12–15 Using a Mendelian Randomization design, Sun et al. 14 confirmed a bidirectional and causal relationship between sleep disturbance and psychiatric disorders including MDD. A systematic review of experimental research on sleep and emotion also found strong evidence that periods of extended wakefulness, shortened sleep duration, and/or nighttime awakenings adversely affect human emotional experiences such as negative affect and depressive symptoms, reduction of positive affect, and increased anxiety symptoms. 15 Postconcussion sleep health may be bidirectionally linked with psychological health. Identification of biological mechanisms underlying these overlapping symptoms may help to motivate personalized treatment approaches to improve concussion recovery prognosis.
Accurate prognosis of concussion recovery remains a significant clinical challenge given the heterogeneity in concussion symptom presentation and recovery trajectories. In current practice, clinicians often rely on self-reported symptoms, which, although validated and clinically relevant, are inherently subjective and susceptible to under- or over-reporting.16–18 There is a need for objective biological measures that reflect underlying pathophysiological processes and can be integrated with existing clinical tools (e.g., Post-Concussion Symptom Scale [PCSS]). Objective biomarkers have the potential to address this gap by enabling risk stratification, refining prognosis, and identifying patients who may benefit from early, targeted interventions. For example, biomarker-informed guidance could augment clinical decision-making to identify the optimal treatment pathway for individual patients. More broadly, integrating biologically informed markers with clinical assessment may allow for more efficient allocation of postinjury recovery guidance by identifying optimal guidance. In addition, these biomarkers may help identify modifiable biological pathways, informing the development of targeted therapeutics and enabling objective monitoring of treatment response.
Salivary microRNAs (miRNAs) are a promising concussion biomarker because they are unique to concussion and are effective at differentiating between concussion cases versus controls. Following concussion, select miRNAs appear to be preferentially associated with underlying concussion pathophysiology and show expression patterns that differ from miRNA changes observed after exercise, musculoskeletal injury, or ischemic brain injury.19–22 While overlap in differentially expressed miRNAs can occur across injury types, experimental models demonstrate traumatic brain injury-associated miRNAs often target biological pathways that differ from those implicated in ischemic injury, potentially reflecting distinct mechanisms of injury and recovery. 20 Antrobus et al. 23 provides a comprehensive review of research studies that have evaluated salivary miRNA biomarkers, among other epigenetic biomarkers, that differ between participants with mild concussion versus controls. Cases include those with sports and nonsports related concussion, samples are often collected within hours to up to 60 days of the initial injury, and diagnosis of concussion and quantification of symptoms are based on variety of clinical tools including World Rugby Head Injury Assessment, Sport Concussion Assessment Tool 3 (SCAT-3), SCAT-5, Glasgow Coma Scale (GCS), Zurich Concussion in Sport Group guidelines, and Standardized Assessment of Concussion (SAC). Control groups are often composed of participants with no recent history of concussion or participants with recent musculoskeletal or orthopedic injury, often matched based on age and/or sports participation. Although beyond the scope of the current paper, salivary miRNAs are also effective at identifying those with persistent or prolonged concussion symptoms24,25 as well as differentiating between patients with differing levels of concussion severity.19,26
Importantly, salivary miRNAs often outperform (higher AUC) concussion predictive models based on clinical, parent, and/or patient-reported symptoms.25,27 Most importantly, salivary miRNAs are correlated with changes in miRNAs in cerebrospinal fluid following traumatic brain injury and thus, provide a potential window into central nervous system dysregulation 28 during concussion recovery. However, associations between salivary miRNAs and specific concussion-related symptoms including sleep disturbance and psychological symptoms have not been comprehensively evaluated. The purpose of this study was to build on prior work by examining the association between miRNAs and both sleep and psychological symptoms that were quantified with the PCSS, in a large cohort of adolescents and young adults with concussion as well as uninjured peers.
Methods
Study population
We identified participants from a prior study 27 of adolescents and young adults. The parent study 27 was a prospective, multicenter study that enrolled participants (ages 5–66) with clinically diagnosed concussion and controls without recent concussion. The clinical diagnosis of concussion was defined according to the 2016 Concussion in Sport Group criteria (Zurich Concussion in Sport Group guidelines). 3 Participants with concussion were recruited within 14 days of injury from diverse clinical settings, outpatient primary care clinics, emergency departments, outpatient specialty care clinics, and sports medicine clinics, and included both sport- and non–sport-related mechanisms. Controls were recruited from similar clinical settings and required absence of concussion within the prior 12 weeks and resolution of any prior symptoms. Exclusion criteria for all participants included non-English primary language, current pregnancy, active periodontal disease, neurologic disorder, drug or alcohol dependency, existing upper respiratory infection, having legally appointed guardian, or inability to provide consent/assent due to intellectual disability. Concussion group-specific exclusionary criteria included the following: GCS ≤12 at the time of initial injury, presence of severe injury (hospitalization, skull fracture, penetrating head injury, and/or evidence of intracranial bleed on advanced imaging), or > 14 days between initial injury and assessment. Participants were excluded from the control group for any of the following: rheumatologic or neoplastic condition, concussion in the previous 90 days, or persistence of symptoms from a prior concussion. In this way, uninjured controls were matched at the group level on the basis of age and sex. Individuals with chronic mental health conditions (i.e., anxiety, depression), chronic headaches, postexercise fatigue, and attentional difficulties (attention-deficit hyperactivity disorder) were targeted for recruitment in order to isolate specific biologic signals related to brain injury, rather than secondary symptomology.
For the present study, our secondary analysis included a subset of participants and samples from the parent study. For this secondary analysis, we filtered the dataset to one saliva sample per individual, keeping the first available sample for analysis. Among participants with a saliva sample available, we excluded participants for the following: existing orthopedic injury at sample collection (n = 46), participant recruited as a post-exercise control (n = 77), incomplete PCSS data (n = 59), and low miRNA counts (n = 4). The final dataset for analysis included 425 participants, n = 272 participants in the concussion group and n = 153 participants in the uninjured control group.
Concussion symptom assessment
Consistent with Hickes et al., 27 PCSS data were collected from all participants (concussion cases and controls) at the time of sample collection. The PCSS quantifies 22 common concussion-related symptoms based on self-reported ordinal scales that range from 0 (no problem) to 6 (severe problem). 29 The tool was designed to monitor concussion recovery and to assess severity of individual concussion-related symptoms. We focused on PCSS scales reflecting psychological (“Feeling More Emotional,” “Irritability,” “Nervous,” and “Sadness”) and/or sleep (“Drowsiness” and “Trouble falling asleep”) symptoms. As described in Hicks et al., 27 saliva samples were collected using the Oragene Discover collection kits (DNA Genotek Inc, ON, Canada) swab procedure, consisting of 10 swab touches to predetermined saliva-rich regions where saliva pools in the sublingual and parotid areas. Samples were handled according to manufacturer specifications and shipped to a central processing lab within 14 days of collection. The timing of sample collection, sample volume, and proximity of sample collection to last meal was tracked for all samples.
RNA processing and quantitation
As described in Hicks et al., 27 salivary RNA was extracted according to the Oragene RNA purification protocol. The yield and quality of the RNA samples were assessed using the Agilent Bioanalyzer. The Illumina TruSeq Small RNA Sample Prep protocol was used for RNA library preparation. Samples were sequenced with the Illumina HiSeq 2500 platform at a targeted depth of 10 million reads per sample. FastQ files were then filtered to remove poor-quality reads and trimming adapter sequences. Reads were then aligned to the build hg38 of the human genome using Partek Flow software with the Bowtie2 aligner. Total reads were quantified with miRBase precursor and mature-microRNA v21.
Quality control procedures were implemented at both the sample and transcript level as described in prior work. 24 Samples with raw miRNA read counts <10,000 were excluded (n = 4). Among the 2500 known human miRNAs, transcripts were retained if the raw read count was >10 in at least 10% of samples. Transcripts that did not meet this minimum abundance criteria were filtered out as low-abundance transcripts. All transcripts meeting filtering criteria, 265 miRNAs, were log2 transformed, quantile normalized, median centered, and standardized (mean = 0 and standard deviation = 1) for downstream analysis.
Statistical methods
Descriptive statistics were used to describe demographics and clinical characteristics in the concussion and uninjured control groups. Spearman’s rank correlation coefficients were used to test the correlation between miRNAs and individual PCSS symptoms representing sleep or psychological health issues. Wilcoxon rank sum tests were then used to test for differences in miRNA expression between the concussion group and the uninjured control group. The Benjamini–Hochberg (BH) method 30 was used to adjust for multiple comparisons. Relationships between miRNAs significantly (BH adjusted p value <0.05) correlated with sleep or psychological symptoms were confirmed using ordinal logistic regression models. Age, sex, BMI, timing of sample collection (i.e., hour of sample collection within a 24-h period due to the influence of time of day on sleep and psychological symptom severity as well as miRNA expression), and self-reported race were adjusted for in all ordinal logistic regression models. All p values from the multivariable ordinal logistic regression models were Bonferroni adjusted based on the number of miRNAs tested. Among candidate miRNAs, multivariable logistic regression models adjusted for age, sex, self-reported race, and timing of sample collection were used to confirm that candidate miRNAs were differentially expressed between concussion cases versus controls. Candidate miRNAs were defined as miRNAs that that were significantly (BH adjusted p value <0.05) correlated with all three outcomes: (i) presence of concussion (case vs. control), (ii) sleep symptom severity (“Drowsiness,” “Trouble Falling Asleep”), and (iii) psychological symptom severity (“Feeling More Emotional,” “Irritability,” “Nervous,” and “Sadness”). All p values from the multivariable logistic regression models (concussion case vs control) were Bonferroni adjusted based on the number of miRNAs tested. For the sake of consistency throughout the remainder of the paper, when reporting the relationship between miRNA levels and quantitative PCSS symptoms, effects are described as “correlations,” miRNAs are either positively or negatively correlated with specific symptoms. When evaluating the relationship between miRNA levels and the presence of a concussion, miRNAs that differ between individuals in the concussion vs the control group are described as differentially expressed, either up- or down-regulated in the concussion group relative to the control group.
Enrichment analysis
We evaluated the pathophysiologic functions of miRNAs by testing whether their downstream gene targets were enriched within known biological pathways and/or processes. We focused on miRNAs correlated with one or more of the sleep symptoms and one or more of the psychological symptoms. Enrichment analysis was performed using miRPath V4.0 using the microclip (brain) and microT-CDS algorithms (threshold >0.80) with miRbase V22.1 and Kyoto Encyclopedia of Genes and Genomes (KEGG) annotations. Enriched terms were assessed at a BH-adjusted p < 0.05.
Standard protocol approvals, registrations, and patient consents
This secondary study was reviewed and approved as exempt by the Colorado Multiple Institutional Review Board (COMIRB#: 24-1386), and the requirement for informed consent was waived. Informed consent and/or assent were obtained from all participants, as appropriate, in the parent study.
Results
The average age among the 425 participants was 18.0 (±5.0) years. Differences in demographics and clinical characteristics in the concussion group vs the uninjured control group are described in Table 1. In the concussion group (n = 272), saliva samples were collected at a median of 2.5 days post-injury (interquartile range: 2.41–10.65 days). Concussions occurred most frequently during participation in sports (188/272, 69.1%). The median overall PCSS symptom severity score across all concussion cases was 16 (IQR: 4–39). Loss of consciousness and memory loss occurred in 15.1% (41/272) and 32% (87/272) of participants in the concussion group, respectively.
Comparison of Participants in the Concussion vs the Uninjured Group

Correlations with sleep and/or psychological symptoms
We identified 14 miRNAs that were significantly (BH adjusted p < 0.05) correlated with sleep symptoms and 28 that were correlated with psychological symptoms (Fig. 1). Four miRNAs, miR-361-5p, miR-4510, miR-193a-3p, and miR-3074-5p, were correlated with both sleep and psychological symptoms (Fig. 1 and Table 2). Multivariable ordinal logistic regression analyses confirmed correlations between three of these miRNAs and individual PCSS concussion symptoms (Table 3). miR-4510 was negatively correlated (Odds Ratio [OR] < 1 and Bonferroni adjusted p value <0.05, Table 3) with the ‘Drowsiness’ and ‘Feeling More Emotional’ PCSS symptom scores. miR-193a-3p was positively (OR > 1 and Bonferroni adjusted p value < 0.05, Table 3) associated with the “Trouble Falling Asleep,” “Feeling More Emotional,” “Irritability,” “Sadness,” and “Nervousness” PCSS symptom scores. miR-3074-5p was inversely correlated (OR < 1 and Bonferroni adjusted p value < 0.05, Table 3) with the “Trouble Falling Asleep” and “Feeling More Emotional” PCSS symptom scores. miR-361-5p was inversely correlated (OR < 1 and Bonferroni adjusted p value < 0.05, Table 3) with “Feeling More Emotional” PCSS symptom scores but was not significantly associated with any of the sleep-related symptoms (Bonferroni adjusted p value > 0.05, see Table 3).

Overlap in miRNAs differentially expressed in concussion vs uninjured control group, correlated with PCSS psychological symptoms, and/or correlated with PCSS sleep symptoms. Description/Legend: Venn diagram representing overlap in miRNA discovery analyses testing
Candidate miRNAs Based on Significant Relationship with Multiple Outcomes, Differentially Expressed in Concussion vs Control Group, Correlated PCSS Sleep Symptoms and/or Correlated with PCSS Psychological Symptoms

↓ Downregulated in concussion group relative to uninjured control group, ↑ Upregulated in concussion group relative to uninjured control group.
(+) positively correlated with PCSS symptom subscale, (−) negatively correlated PCSS symptom subscale.
Ordinal Logistic Regression Results to Confirm Correlations between miRNA Levels and PCSS Symptoms
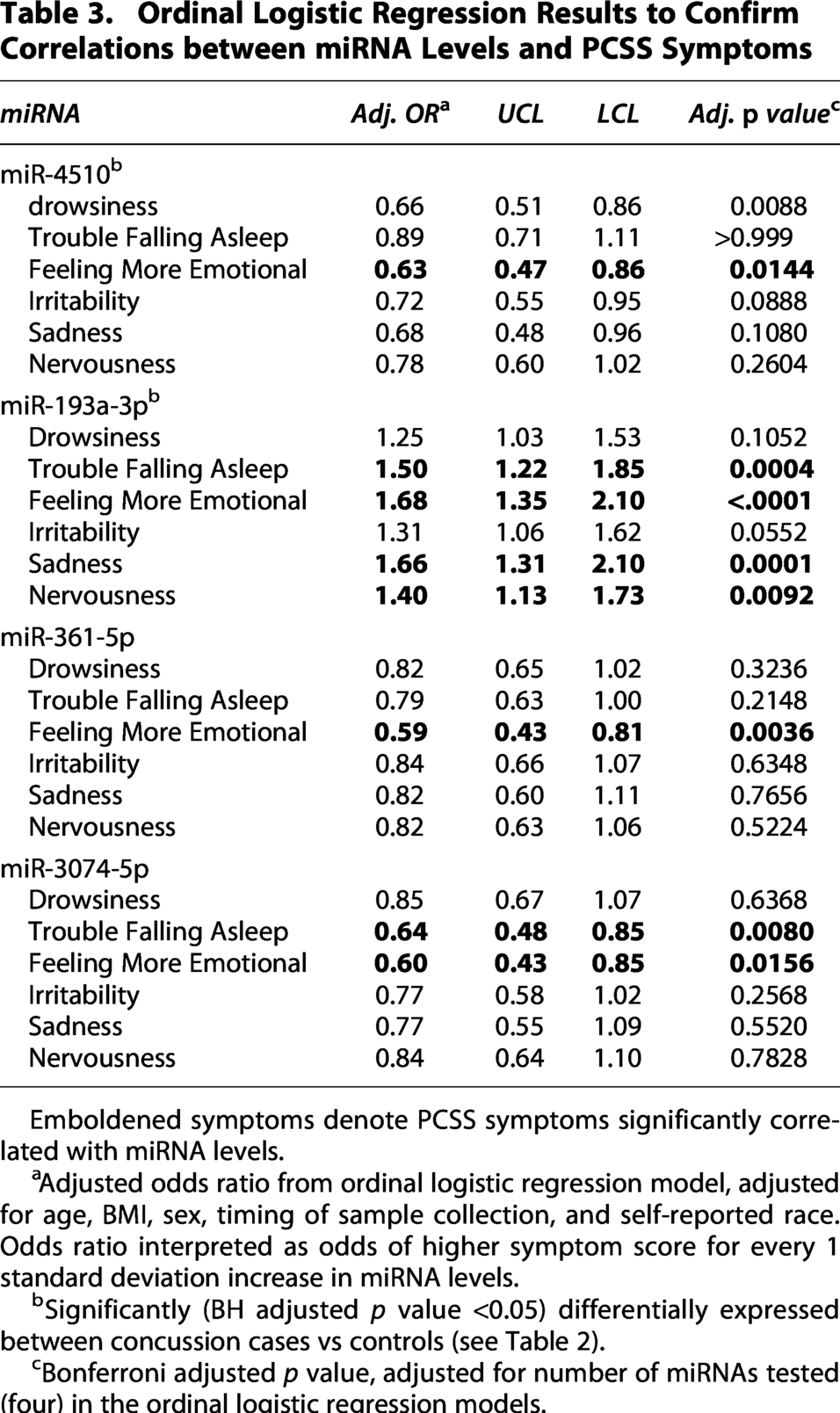
Emboldened symptoms denote PCSS symptoms significantly correlated with miRNA levels.
Adjusted odds ratio from ordinal logistic regression model, adjusted for age, BMI, sex, timing of sample collection, and self-reported race. Odds ratio interpreted as odds of higher symptom score for every 1 standard deviation increase in miRNA levels.
Significantly (BH adjusted p value <0.05) differentially expressed between concussion cases vs controls (see Table 2).
Bonferroni adjusted p value, adjusted for number of miRNAs tested (four) in the ordinal logistic regression models.
Concussion group versus uninjured control group
We identified 89 miRNAs that were differentially expressed (BH FDR adj. p < 0.05) between the concussion group (n = 272) compared to the uninjured control group (n = 153, Fig. 1).
miRNA prioritization—identification of candidate miRNAs
We reviewed the 21 miRNAs that were correlated with multiple outcomes (see overlapping regions in Fig. 1). Correlations (positive or negative) between these miRNAs and the three outcomes are described in Table 3 and Figure 2. We prioritized miRNAs that were correlated with all three outcomes, (i) differentially expressed between the concussion group vs the uninjured control group, (ii) correlated with one or more sleep symptom(s), (iii) correlated with one or more psychological symptom(s). miR-193a-3p and miR-4510 met these criteria and thus were considered candidates (Figs. 1 and 2). After adjusting for age, sex, BMI, timing of sample collection, and self-reported race, multivariable logistic regression models confirmed miR-4510 was downregulated in the concussion group (adjusted odds ratio [aOR] concussion vs. control: 0.34, 95% CI: 0.23–0.49, Bonferroni adjusted p value: <0.0001) and miR-193a-3p was upregulated in the concussion group (aOR concussion vs. control: 1.49, 95% CI: 1.13–1.97, Bonferroni adjusted p value: 0.0100).
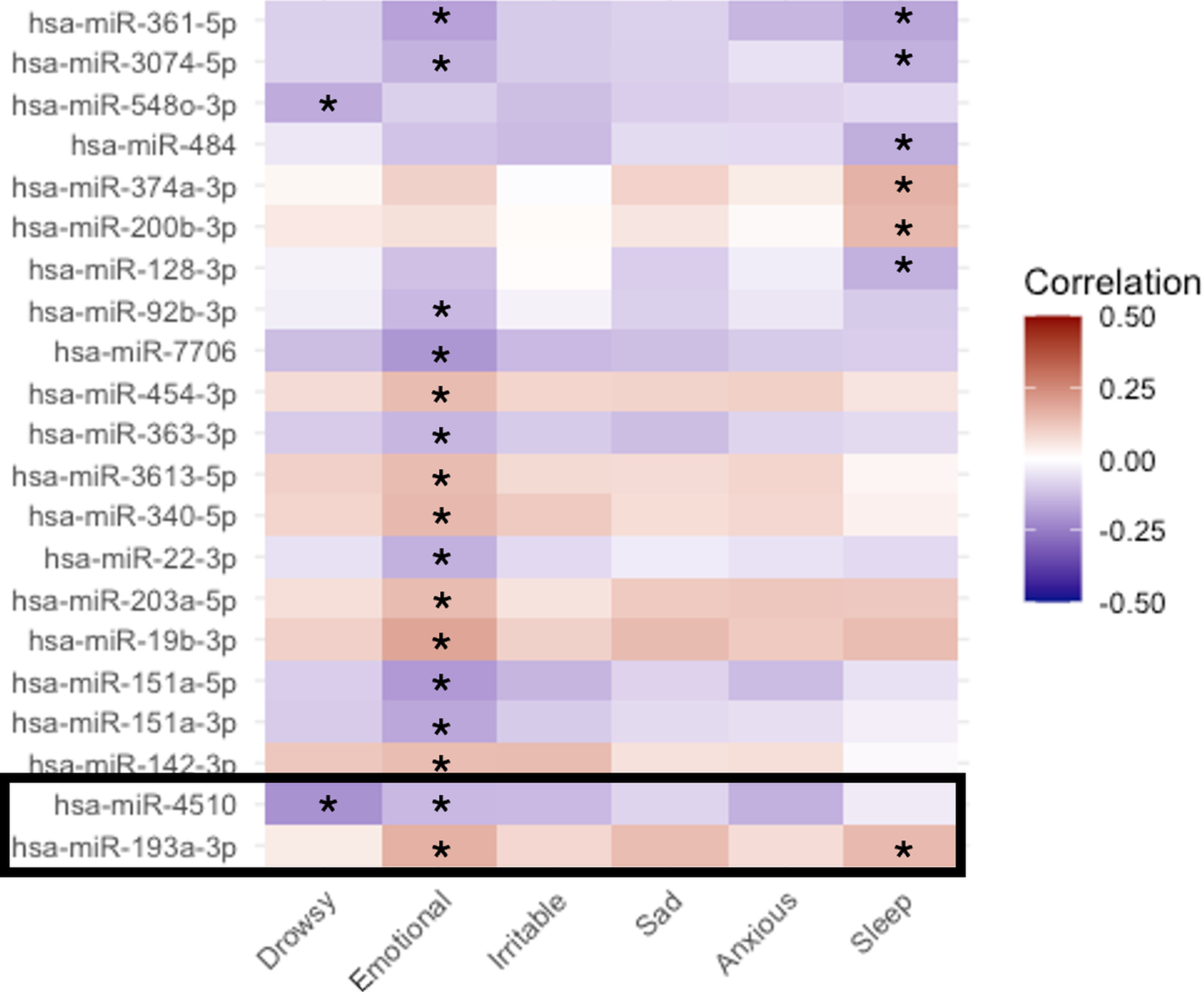
Heatmap of the strength of correlation between 21 candidate miRNA transcripts and PCSS psychological and sleep symptoms. Description/Legend: Heatmap representing strength of correlation (Spearman’s correlation coefficients) between candidate miRNA levels and PCSS psychological symptoms (“Feeling more emotional”, “Irritability”, “Sadness”, and “Nervousness”) and sleep symptoms (“Drowsiness”, and “Trouble falling asleep”). Candidate miRNAs include all transcripts significantly, BH adjusted p value < 0.05, correlated with multiple outcomes (concussion case vs uninjured control, psychological symptom(s), and/or sleep symptom(s)). The black box represents priority candidates, correlated with both psychological and sleep symptoms and differentially expressed between concussion cases and uninjured controls (see Fig. 1). On the x-axis, Drowsy = “Drowsiness”, Emotional = “Feeling more emotional”, Irritable = “Irritability”, Sad = “Sad”, Anxious = “Nervousness”, Sleep = “Trouble falling asleep”. *Indicates correlations significant at the BH adjusted p value < 0.05.
Enrichment results
Enrichment analysis was performed using genes targeted by miRNAs that were significantly (BH adj. p value < 0.05) correlated with both sleep and psychological health-related symptoms, miR-193a-3p, miR-3074-5p, miR-361-5p, miR-4510. We identified 85 pathways that were significantly (BH adjusted p < 0.05) enriched by genes targeted by miR-193a-3p, miR-361-5p, and miR-4510, see Figure 2. Among the brain-related KEGG pathways targeted by miR-4510, miR-361, and/or miR-193a were “Spinocerebellar ataxia” (BH adj. p = 1.06E-06), “Dopaminergic synapse” (BH adj. p = 1.06E-06), “Axon guidance” (BH adj. p = 1.45E-05), “Circadian entrainment” (BH adj. p = 3.79E-05), and “amphetamine addiction” (BH adj. p = 2.91E-04). A full list of all pathways identified in the enrichment analysis is described in Figure 2 and Supplementary Table S1.
Discussion
We identified several salivary miRNAs that were differentially expressed in participants in the concussion group compared to the uninjured control group, and furthermore, identified miRNAs were strongly correlated with sleep and/or psychological symptoms. Persistently altered levels of these miRNAs may serve as markers of biological processes accompanying concussion and may be associated with sleep disturbance and/or psychological symptoms.
We identified 7 miRNAs that were correlated with sleep symptoms and were differentially expressed between the concussion and uninjured control groups (see Table 3 and Fig. 1). Several of these miRNAs including miR-374a-3p, 31 miR-361-5p, 32 and miR-4510, 27 have been previously associated with concussion or concussion-related symptoms. Although the specific roles of these miRNAs in the development of sleep symptoms following concussion are not known, several studies have identified direct links between brain-specific miRNAs and sleep and/or circadian clock-related pathways.33,34 For example, miR-219-1 plays a role in regulating the length of the circadian day whereas miR-132, a light-activated transcript, modulates the impact of light on the circadian clock. 33 In-vivo and in-vitro work have also confirmed that miR-183, miR-96, and miR-182 can modulate circadian rhythms. 34 Inhibition of specific miRNAs including mir-138, directly resulted in changes in sleep duration, 35 demonstrating miRNAs as potential modulators of sleep behavior. Furthermore, increased expression of miR-193a-3p, which was positively associated with both ‘drowsiness’ and ‘trouble falling asleep’ in our study (see Table 3), has been observed in microglia following traumatic brain injury in mice. 36 Microglia, when chronically activated, contribute to the upregulation of nuclear factor-KB (NF-κB), a protein transcription factor that drives the production of pro-inflammatory cytokines such as IL-1β and TNF, both of which are considered sleep-regulating substances as they act to enhance or inhibit sleep. 37 Our findings indicate a complex relationship between miRNA levels and sleep-related symptoms.
We also identified 14 miRNAs that were strongly correlated with psychological health symptoms (irritability and/or anxiety) and were differentially expressed between the concussion and the uninjured control groups. Of these miRNAs, miR-4510, 27 miR-361-5p, 32 miR-151a-5p, 21 miR-142-3p, 38 miR-92b-3p, 24 miR-3613-5p, 39 miR-19b-3p, 40 miR-22-3p, 40 and miR-203a-5p 41 have been associated with the presence of concussion or concussion-related symptoms in prior work. More importantly, miRNAs are also related to the development of unique phenotypic subgroups after concussion, including a subgroup characterized by the presence of emotional symptoms. 42 In relation to psychiatric disorders, miRNAs have emerged as important biomarkers and even as potential treatment targets.43,44 miRNAs are dysregulated in MDD and are hypothesized to contribute to development of MDD based on their role in regulating pathways relevant to MDD etiology including neuroinflammation, neuroplasticity, neurotransmitters, hyperactivity of the hypothalamic-pituitary-adrenal axis, oxidative stress, and circadian rhythms.43,44 While there are no current miRNA-based therapies for depression and/or anxiety that are approved for human use, miRNA-based therapies are being studied in multiple animal models.45,46 For example, injection of miR-101 into the ventrolateral orbital cortex of rats was able to reverse depressive-like behavior in the chronic unpredictable mild stress depression rat model. 45 Overall, our work and others highlight correlations between miRNAs and psychological symptoms. Additional work is needed to understand how levels of psychological-related miRNAs change during concussion recovery and whether changes in these miRNAs modify clinical outcomes.
We identified miRNAs associated with both sleep and psychological health symptoms. We performed enrichment analysis to identify pathways targeted by these miRNAs. The “Circadian entrainment” and “Dopaminergic synapse” pathways were among the top significantly enriched pathways (see Supplementary Table S2 for a list of genes included in this pathway that are downstream gene targets of candidate miRNAs), supporting a direct biological link between miRNAs identified in our study and pathways relevant to both sleep and psychological health.47,48 “Spinocerebellar ataxia,” representing a group of neurodegenerative diseases impacting the cerebellum, was the top significantly enriched KEGG term (see Supplementary Table S2 for list of genes included in this pathway that are downstream gene targets of candidate miRNAs). This group of diseases is characterized by progressive loss of balance and coordination. 49 Although spinocerebellar ataxia represents a distinct pathophysiological disease process, these symptoms and cerebellar-related genes are potentially relevant to concussion due to known disruptions in balance and coordination deficits observed during concussion recovery 50 as well as the potential role of these disruptions in subsequent lower extremity musculoskeletal (MSK) injury risk, an adverse secondary outcome that is known to be increased following concussion. 51 However, MSK injury is complex, and additional work is needed to study whether miRNAs identified in the current study and their downstream gene targets contribute to increased risk of MSK injury.
We identified two miRNAs (miR-4510 and miR-193a-3p) that were associated with sleep disturbance, psychological symptoms, and were differentially expressed between the concussion and uninjured control groups. Both miRNAs target downstream genes that are relevant to the development of sleep issues and the onset of psychological symptoms. Based on the miRBD resource (mirdb.org), the PAK3 gene, involved in neurodevelopmental pathways, is one of the top predicted targets (Target score = 99) for miR-4510. Expression of PAK3 was observed to be significantly differentially expressed in postmortem brain tissue samples obtained from individuals with versus without clinical signs of depression. 52 miR-193a-3p targets the MAPK10 gene (Target score = 99, mirdb.org), a member of the MAP kinase (MAPK) family, which is expressed in neurons. Alterations in MAPK signaling pathway are hypothesized to contribute to the pathophysiology of MDD 53 and furthermore, based on the role of the MAPK pathway in circadian system entrainment, MAPK has been proposed as a treatment target for MDD. 54 Overall, both miR-4510 and miR-193a-3p represent ideal candidates for additional research to confirm their relevance to the broad spectrum of concussion pathophysiology, including postinjury recovery, sleep health, and psychological health.
Limitations
Due to our cross-sectional design, we cannot determine whether miRNA levels preceded sleep and/or psychological symptoms, or conversely, whether the symptoms caused changes in miRNAs. Additional work is needed to evaluate longitudinal changes in miRNAs as well as longitudinal changes in sleep and psychological symptoms during and following concussion symptom recovery. The sleep and psychological symptoms were collected with the PCSS. Although an effective and well-accepted instrument for quantifying concussion symptoms in the clinical setting, the individual symptoms quantified with this tool are broad. We selected individual symptoms that reflect sleep and psychological health-related symptoms. We acknowledge there is overlap in many of the symptoms included in the PCSS and thus, additional work needs to confirm correlations using instruments validated specifically for quantifying specific psychological health issues as well as specific sleep health issues. Furthermore, to align with our target patient population, inclusive of patients seeking care for concussion across outpatient primary care clinics, emergency departments, outpatient specialty care clinics, and sports medicine clinics, data were collected across numerous clinical environments, which potentially introduced heterogeneity related to the collection of pre-existing conditions. Although the prevalence of pre-existing conditions including history of depressive or anxiety disorders as well as ADHD were similar between the concussion and control groups, additional work is needed to evaluate the role of these and other pre-existing medical conditions in the development of specific symptoms during concussion recovery. Finally, point-of-care assessments for quantifying miRNAs do not currently exist. Technological advancements capable of more rapid quantification/measurement of miRNAs are necessary before miRNA biomarkers can be directly translated into the clinical setting.
Conclusions
Our work supports the interrelationship between salivary miRNAs, psychological symptoms, and sleep symptoms after concussion. We evaluated correlations between salivary miRNAs and three outcomes: presence of concussion, correlation with sleep issues and correlation with psychological health issues. We identified two miRNAs, miR-4510 and miR-193a-3p, that were associated with all three outcomes. miR-4510 was lower in concussion cases and inversely associated with “drowsiness” and “feeling more emotional.” In contrast, miR-193a-3p was higher in concussion cases and positively associated with “trouble falling asleep” and “feeling more emotional.” These findings underscore the possibility that miRNA profiles may reflect the complex interaction between concussion, sleep symptoms and psychological health symptoms. However, given the cross-sectional design, the directionality and temporality of these relationships cannot be determined. It remains unclear whether miRNAs precede the development of sleep and psychological symptoms or whether they reflect downstream effects of these concussion features. Longitudinal studies are needed to validate these associations, establish temporal relationships, and determine whether these candidate miRNAs may serve as predictive and/or mechanistic biomarkers of concussion recovery.
Transparency, Rigor, and Reproducibility Summary
The authors affirm that this article is an honest, accurate, and transparent account of the data being reported. In the interests of transparency, rigor, and reproducibility, this study provides descriptions of all experimental methods, data analysis, and statistical tests used. The miRNA data used in the analysis are not publicly available. However, de-identified data including participant demographics and miRNA data are available from the corresponding author upon reasonable request. Data requests may require a signed data access agreement per institutional and/or ethics review board requirements.
Authors’ Contributions
P.M.C. contributed to study conception and design, statistical analysis and interpretation, and article drafting/revision. H.S. contributed to statistical analysis, data analysis and interpretation, and article drafting/revision. E.B.K. contributed to data analysis and interpretation, and article drafting/revision. J.C.W. contributed to data interpretation, and article drafting/revision. B.H. contributed to protocol and database development, participant enrollment, acquisition of data, and preparation of data for analysis. C.C.D. contributed to data interpretation and article drafting/revising. J.L. contributed to acquisition of data and article revision. R.P.O. contributed to acquisition of data, data analysis and interpretation, and article revision. C.O. contributed to acquisition of data and article revision. D.R.H. contributed to study conception and design, data interpretation, and article drafting/revision. S.D.H. contributed to study conception and design, acquisition of data, data analysis and interpretation, statistical analysis, and article drafting/revision.
Data Availability
Anonymized data not published within this article. De-identified data including participant demographics and miRNA data are available from the corresponding author upon reasonable request. Data requests may require a signed data access agreement per institutional and/or ethics review board requirements.
Footnotes
Acknowledgments
The study was partially supported by R42-NS119119 and R01-NS115942. SDH formerly served as a scientific advisory board member to Quadrant Biosciences and Spectrum Solutions. Neither party played a role in the analyses of this data or the creation of this article. The authors gratefully acknowledge the contributions of the following authors and collaborators who were involved in the collection and processing of data for the parent study: Gregory Fedorchak, Samantha DeVita, and Aakanksha Rangnekar, of Quadrant Biosciences; John Leddy and Mohammad N. Haider, of SUNY Buffalo; Zofia Gagnon, of Marist University; Aaron M. Yengo-Kahn and Scott Zuckerman, of Vanderbilt University; Chuck Monteith, of Colgate University; Thomas R. Campbell, of Bridgewater College; and Christopher Neville and Frank Middleton, of SUNY Upstate.
Author Disclosure Statement
J.C.W. is a member of American Medical Society for Sports Medicine (AMSSM). D.R.H. has received research support from the Eunice Kennedy Shriver National Institute of Child Health & Human Development (R03HD094560, R01HD108133), the National Institute of Neurological Disorders And Stroke (R01NS100952, R43NS108823), the National Institute of Arthritis and Musculoskeletal and Skin Diseases (1R13AR080451), 59th Medical Wing Department of the Air Force, MINDSOURCE Brain Injury Network, the Tai Foundation, the Colorado Clinical and Translational Sciences Institute (UL1 TR002535‐05), and the Denver Broncos Foundation. S.D.H. formerly served as the Chief Medical Officer and a Scientific Advisory Board Member for Quadrant Biosciences, Inc. and as a Scientific Advisory Board Member for Spectrum Solutions. S.D.H. is named as co-inventor on intellectual property involving the use of salivary microRNAs for the detection and management of mild traumatic brain injury by the Pennsylvania State University. All other authors declare no conflict of interest. All authors certify that the data presented here are original and are not under review at any other publication.
Funding Information
No funding was received for this study.
Supplemental Material
Supplemental Material
Abbreviations Used
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.