
Abstract
Hedgehog signaling was discovered more than 40 years ago in experiments demonstrating that it is a fundamental mediator of limb development. Since that time, it has been shown to be important in development, homeostasis, and disease. The hedgehog pathway proceeds through a pathway highly conserved throughout animals beginning with the extracellular diffusion of hedgehog ligands, proceeding through an intracellular signaling cascade, and ending with the activation of specific target genes. A vast amount of research has been done elucidating hedgehog signaling mechanisms and regulation. This research has found a complex system of genetics and signaling that helps determine how organisms develop and function. This review provides an overview of what is known about hedgehog genetics and signaling, followed by an in-depth discussion of the role of hedgehog signaling in craniofacial development and carcinogenesis.
Introduction
The Hedgehog (Hh) signaling pathway is a highly conserved pathway involved in a multitude of cellular functions including development and homeostasis. The Hedgehog gene (hh) was originally identified in Drosophila in the seminal screening experiments of Nusslein-Volhard and Weischaus that led to a Nobel Prize in 1995. 1 The gene is so named because a loss of function mutation in Drosophila led to the flies being covered with denticles, or little spikes, which gave them the appearance of hedgehogs. Further studies found that there were 3 mammalian counterparts to the Drosophila hh gene, Sonic hedgehog (Shh), Indian hedgehog (Ihh), and Desert hedgehog (Dhh).2-5 Subsequent research has shown that the Hh signaling pathway is of vital importance in human development and physiology. Furthermore, it has important implications in tissue engineering and cancer therapeutics.
Skeletal and craniofacial development is a complex process that is controlled by numerous pathways. The Hh pathway has emerged as one of the most important specifically related to craniofacial and limb development. The importance of vertebrate Hh proteins was clearly demonstrated when Sonic hedgehog expressing cells grafted on to developing chick wings produced the same effect as grafts of the “polarizing regions” from the posterior side of the wing- both of these procedures created limbs with mirrored digits. 2 It was shown that the diffusion and concentration of this secreted morphogen, hedgehog ligand, was responsible for the antero-posterior axial patterning of limbs. Additionally, it was also quickly realized that Hh signaling was vital in craniofacial development when Shh deficient mice were found to have cyclopia. 6
Recent research has revealed that hedgehog signaling is also involved in many other homeostatic and pathologic processes in humans, potentially making it a useful therapeutic target. For example, aberrant Hh pathway signaling has been linked to numerous cancers including medulloblastoma, basal cell carcinoma, glioblastoma, and breast cancer.7-9 Hh has also been implicated in the maintenance and differentiation of stem cells, and modulation of Hh signaling holds promise in regenerative medicine.10,11
An updated review of hedgehog is timely given that the understanding of hedgehog signaling is rapidly changing and growing. This warrants an up to date overview of the progression of research on the intricacies of the pathway and associated clinical applications. This is highly relevant for the field of craniofacial surgery, as the role of Hh in development is of increasing focus in both normal and aberrant processes such as craniosynostosis and head and neck cancers. Molecular considerations of Hh signaling will be reviewed and we will summarize the research from the past 40 years while highlighting important aspects of this complex pathway in development and disease.
Signaling Mechanism
Hedgehog signaling is a highly conserved pathway across animals. In broad terms, Hedgehog ligands are formed and released from a cell, drift to their target cell, bind to their receptor, and initiate an intracellular cascade leading to the activation of transcription factors that control gene expression. The intracellular signaling is highlighted in Figure 1.
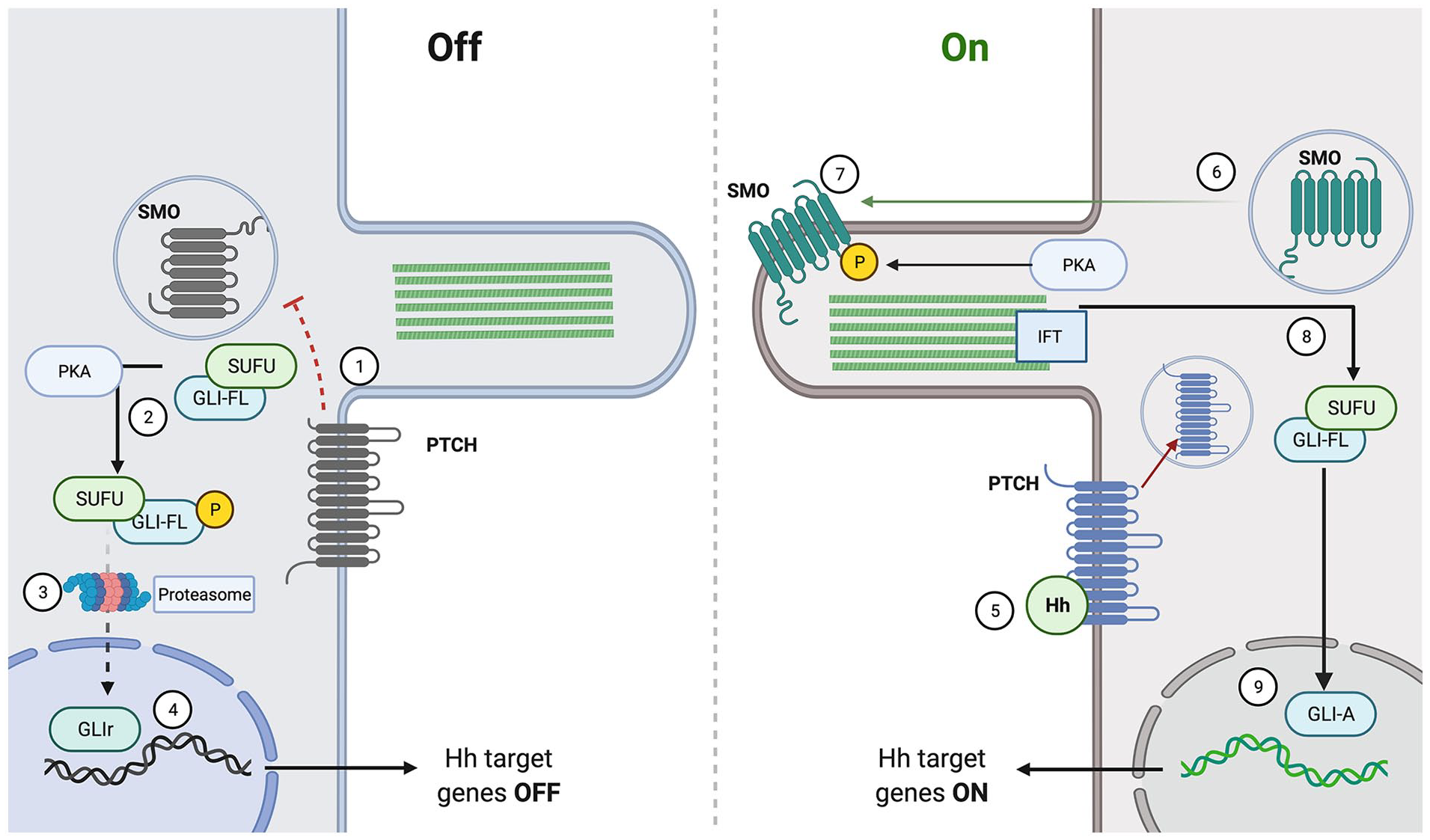
Canonical hedgehog signaling.
Secreted Hh ligands fulfill the definition of morphogen, meaning they are able specify cell fate in a graded fashion. There are 3 Hh homologues in vertebrates: Dhh, Ihh, and Shh. These proteins are synthesized as ~45 kD precursor protein which then undergo auto-proteolysis by the C-terminal domain to form 19 kD lipidated proteins from the amino-terminal domain which contains the signaling mechanism.12,13 The basic structure of the Hedgehog ligand is shown in Figure 2. These smaller lipidated proteins have a 2 lipid modifications, a palmitoyl acid on the N-terminal and a cholesterol moiety on the C-terminal.12,14 These lipid modifications make the protein more closely associated with the cell membrane and are an important determinant of accurate Hh signaling. Studies have shown that Hh ligands without the lipid moieties diffuse farther but lead to a low threshold activation and do not enter or activate cells in the same way that the modified ligands do. 15 Cholesterol is frequently a modification added to proteins to localize them to the plasma membrane, and it has been shown that Hh with its lipid modifications is closely associated with plasma membranes. This has led to much research into how these lipid- associated structures diffuse across a large area. Two mechanisms have been proposed, one being that Hedgehog ligands are associated with exosomes, or other extracellular carriers, that help them move across an area and protect their hydrophobic tails,16,17 while another hypothesis proposes that the ligands can be released from their lipid modifications by matrix metalloproteases. 18 These lipid modifications and diffusion mechanisms are important in regulating the spread of the morphogen across its field of action. 19
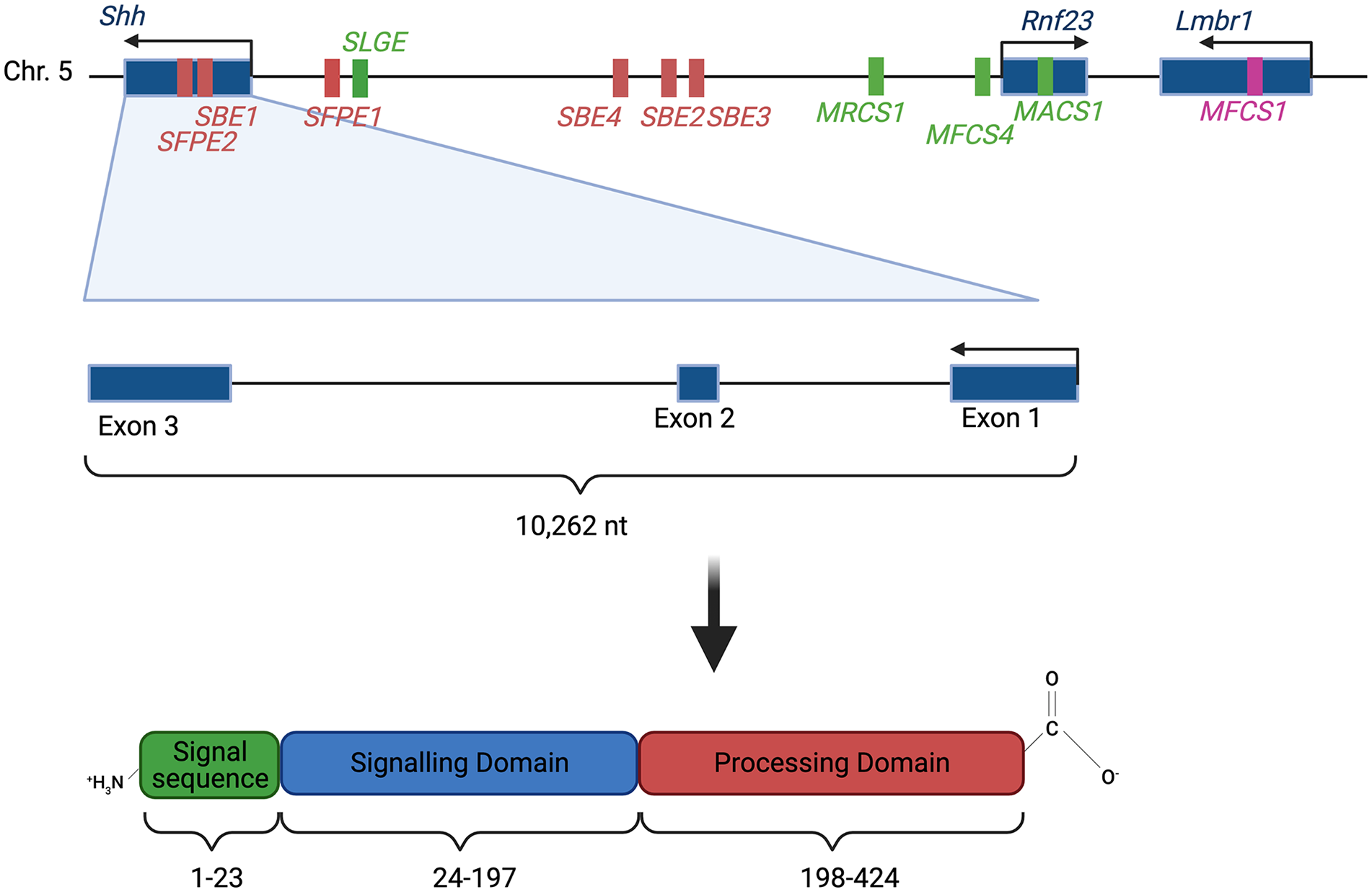
Sonic hedgehog (Shh) locus
In the canonical pathway, after formation and modification, the hedgehog ligands are released from the cell by the large transmembrane Dispatched (Disp) protein. 20 Recent research using fluorescence labeled proteins has shown that Disp is also involved in the re-uptake of released Hh ligands, which is important to the recycling of the ligands and long range Hh signaling using non-lipid modified ligands. 21 After being released from the cell and diffusing to its target cell using one of the mechanisms described above, the ligand then binds to the 12-span transmembrane receptor Patched (Ptch) which upon binding releases its inhibition of the 7-pass transmembrane protein Smoothened (Smo). 22 In the absence of a Hh ligand Ptch destabilizes Smo, but upon binding Ptch is internalized and degraded allowing Smo to translocate to the membrane. 23 Notably the PTCH gene has been identified as a tumor suppressor and then SMO gene has been identified as a oncogene in a variety of cancers including carcinomas and medullablastomas. 24
The release of inhibition on Smo leads to an intracellular signaling cascade as seen in Figure 1. Smo is a GPCR- (G-Protein Coupled Receptor) like protein and its activation leads to a synergy with G protein-coupled receptor kinase 2 (GRK2) which starts a phosphorylation cascade. Smo activation also requires localization to the primary cilium (PC). Fluorescence microscopic analysis localized Smo to the PC and showed that mutations in the hydrophobic domain necessary for this PC localization led to a lack of activation. 25 The activation of the intracellular cascade by Smo is passed through the Fused (Fu) and Suppresor of Fused (Sufu) proteins. In the inactivated state Sufu constrains the glioma-associated oncogene transcription factors (Gli) in the cytoplasm. Upon activation the Sufu-Gli complex moves to the PC and Fu antagonizes the repressive effects Sufu leading to the nuclear localization of the Gli transcription factors.26,27 In short, Hh ligand binding causes dissociation of Gli from Sufu and it then moves to the nucleus. A knockout Sufu−/− mouse model showed constitutive, Hh independent, activation of the pathway, demonstrating the function of the Sufu protein in repressing Hh signaling. 28
The Gli family consists of 3 zinc finger transcription factors Gli1/Gli2/Gli3 that were among the first zing finger proteins to have their 3D structure resolved. 29 As their name suggests they were first discovered in relation to cancer, but they have numerous and varied functions in vivo.30,31 The Gli transcription factors exist in 2 forms, their transcription activating form known as Gli-A and their transcription repressing form known as Gli-R. The balance between these 2 forms determines the Gli code, which is the gene expression profile that Hh signaling produces. 32 Studies have shown that proteolytic processing of the Gli2 and Gli3 proteins leads to their repressive forms. This proteolysis is performed by proteins such as the SCFβ-TRCP complex. 33 When Hh signaling is in the “off” formation, the Gli proteins are phosphorylated by PKA which activates the proteolytic processing. These repressive proteins translocate to the nucleus and switch “off” target genes.32,34 When the Hh signal is switched “on”, the Gli transcription factors localize to the nucleus in their active forms where they target a host of genes, many of which have been identified in genome wide expression analysis studies.35-37 In the active form, Smo inhibits proteasomic processing of Gli, allowing for the full-length Gli proteins to activate transcription of their target genes. 27 Notably it has been found that tissue specific signals are necessary for the Gli proteins to regulate certain genes, meaning that Hh gene activation is position dependent and can have pleiotropic effects. 38
Briefly, the Hh pathway can also be activated in a non-canonical matter. In general non-canonical hedgehog signaling refers to when pathway activation is “Gli- independent”, that is it does not proceed through Gli mediated transcription, or does not proceed through the classic Ptch and Smo interactions.39,40 Numerous studies have shown that in cancer hedgehog signaling proceeds through Ptch-independent or Smo-independent routes, and that this non-canonical signaling can have crosstalk with a variety of other pathways.41,42 This non-canonical signaling is particularly important in explaining why certain cancers may be resistant to therapeutics targeting hedgehog. 43
Overall, hedgehog signaling follows a remarkably conserved pathway among animals from Drosophila to humans. Its unique aspects, such as the adjustment of ligand diffusion and tissue specific response, make it a fascinating and important area of study.
Genetics
The Hedgehog (Hh) gene is highly conserved from fruit flies all the way to humans. 44 In humans, the Sonic Hedgehog (SHH) gene is located on the long arm of the seventh chromosome and the Indian Hedgehog gene (IHH) is located on the second chromosome. 45 Desert Hedgehog (DHH) gene is located on the long arm of the 12th chromosome. 46 These genes code for Hh family proteins which play essential roles in development. 47 They are primarily active during development and are mostly inactive in adulthood, with the exception of stem cells and epithelial cells of some internal organs. 44 Given their role in development, it is no surprise that mutations of the Hh genes can result in a variety of congenital disorders.
Hh gene expression is highly variable depending on the tissue type and location. It is also highly specific spatiotemporally as the expression pattern of Hh genes has a large effect on how an organism develops. Thus, the expression of these genes is highly regulated. Of the mammalian Hh genes, only the regulation of Shh gene has been studied in great detail and more specifically mouse Shh has been studied most extensively as a model of human SHH. 47 Mouse Shh transcription is controlled by a number of independent enhancers, both long-range and local. Shh expression in the central nervous system is controlled partially by Shh floor plate enhancer 1 (SFPE1) and SFPE2. These local enhancers direct expression at the floor plate of the hindbrain and spinal cord. Shh brain enhancer 1 (SBE1) directs expression at the ventral midbrain and caudal diencephalon. The enhancers SBE2, SBE3, and SBE4 are located more distally, about 400 kb upstream of the Shh transcription site, and regulate expression in the ventral forebrain.47,48 The mouse Shh gene locus with the local enhancers mentioned here are highlighted in Figure 2.
Outside the central nervous system, the mammals-fish conserved sequence 1 (MFCS1) enhancer controls Shh expression in the zone of polarizing activity (ZPA) of the limb buds. This enhancer is located even more upstream than SBE2, SBE3, and SBE4.47,48 Loss of function of MFCS1 has been shown to result in complete loss of Shh expression in the limb bud, resulting in truncation of the limb in mice. 49 Indeed, limb malformations are seen in humans with mutations in the MFCS1 enhancer. 50 Other notable regulators of Shh in the limb bud are the Alx4 gene and the erythroblast transformation specific (ETS) transcription factor genes Etv4 and Etv5. These function to decrease Shh expression in specific pattern in the limb bud, limiting its expression to the posterior margin of the limb. 48
Another set of enhancers located 600 to 900 kb upstream of the Shh gene regulate Shh expression in the epithelial lining of the pharyngeal cavity, lungs, and digestive tubes. MACS1 is the enhancer that drives expression in the posterior gut epithelium, the tracheal tube, the lungs, intestine, and urogenital tract. The enhancer MRCS1 controls expression in the tooth, hard palate, and tongue. A third enhancer, MFCS4 influences expression in areas overlapping the first 2 as well as in the epiglottis, arytenoids, and tympanic tube. Another enhancer, SLGE (Shh lung, gut enhancer) overlaps with the MACS1 enhancer and drive expression in similar regions, giving the regulatory elements some redundancy. 48
There are likely many other enhancers regulating Shh yet to be discovered.47,48 The specificity of these enhancers to certain areas of the organism demonstrate how the expression of Shh can vary greatly both spatially and temporally.
Role in Osteogenesis
Bone development can occur in 2 ways in vertebrates: via intramembranous ossification or via endochondral ossification. Intramembranous ossification, by which most craniofacial bones are formed, occurs when mesenchymal progenitor cells differentiate into osteoblasts and directly form bone. In contrast, the bones in the rest of the skeletal system are formed via endochondral ossification, a process in which mesenchymal progenitor cells first differentiate into chondrocytes before eventually being replaced by osteoblasts. Shh plays a role in both of these pathways. 51 Shh plays a key role in craniofacial development during intramembranous ossification and early patterning of the skeleton, while Ihh is recognized as a key regulator of endochondral ossification.51-53
Shh has also been noted to play a role in fracture healing and lower expression levels of Shh are thought to contribute to age-related delays in fracture healing. 54 A study of mice without fractures showed the overexpression of Shh resulted in increased osteoblasts, osteoclast, and osteoblast precursors which are important cells in the fracture healing process.55,56 Osteogenesis also involves a number of other signal molecules and pathways, including Wnt, BMP, and Notch. 53 Shh crosstalk with these other pathways lies at the root of numerous processes, including neuronal development and maintenance as well as carcinogenesis.57-59 Crosstalk between hedgehog and Notch signaling is observed during craniofacial development, and mice with Notch ligand disruption showed abnormal Shh signaling in the craniofacial skeleton. 60 Additionally, Shh was found to upregulate the transcription of osteogenic BMPs and synergistically increase alkaline phosphatase (ALP) activity, a measure of bone differentiation, in conjunction with BMP2.61-63 Lastly, inhibition of Wnt has been shown to decrease both Shh- and BMP2- induced ALP activity while conversely, Wnt induced ALP activity was not affected by Shh or BMP2 inhibitors, demonstrating a more complex relationship between Shh and Wnt. 64 It has been proposed that the pathways of all these molecules are linked through the transcription factor Runx2. 64
Ihh is important in regulating endochondral ossification. It is produced by chondrocytes adjacent to the proliferation zone in developing limbs. Ihh functions in conjunction with parathyroid hormone-related peptide (PTHrP) which is expressed by periarticular cells during endochondral ossification. Ihh stimulates PTHrP expression in periarticular chondrocytes, PTHrP then diffuses into the growth plate region to and promotes the proliferation of chondrocytes. Chondrocytesthen exit the cell cycle and undergo hypertrophy when PTHrP expression drops below a critical level as a result of dropping Ihh levels.65,66 These findings all show that both Ihh and Shh are part of a vital system leading to the differentiation and development of the skeletal system.
Function in Development
Hedgehog signaling is vital to the development of nearly every organ system in animals. When Christiane Nüsslein-Volhard and Eric Wieschaus first discovered Hh signaling in the 1980s they described its effect on embryonic development, specifically noting that hh controlled Drosophila cuticle pattern development. 1 Since that time Hh has been found to play a role in everything from the development of the brain to the limbs.
Hh Signaling in Embryonic Patterning
After its study in Drosophila, the study of Hh in vertebrates was keenly awaited by the scientific community given its implications in development. The 2 most well-studied organs in which hedgehog signaling plays a role are the developing neural tube and limb, but new functions continuously arise. The neural tube forms in the fourth to sixth week of development, when the dorsal ectoderm invaginates and pinches off into a tube that will become the central nervous system (CNS). 67 The ventral portion of the neural tube, the floor plate, is distinct from the dorsal portion. An early study found that when ectopically expressed in the CNS, Shh led to the expression of floor plate genes, and thereby determined ventral-dorsal polarity of the neural tube. 68 Other studies showed that Shh was sufficient to induce cells to differentiate into ventral cell types such as motor neurons. 69 The concentration gradient of Shh determines its differential effects, with motor neuron induction occurring at Shh concentrations 5-times lower than floor plate induction. 70 Shh is released by cells in the notochord, which lies ventral to the developing neural tube. This notochordal production induces a second Shh-producing center in the floor plate of the neural tube.71,72 Shh then diffuses across the developing neural tube, with the concentration being highest on the ventral aspect and lowest on the dorsal aspect. This concentration gradient is what creates the VD (ventral-dorsal) axis. Shh signaling determines the cell types by activation of various transcription factors across its gradient. Ultimately, this establishes the complex anatomy of the spinal cord, with different cell types in specific locations.
As discussed, the downstream effects of Hh signaling are mediated through the balance of Gli-A and Gli-R, and increased Shh binding to Ptch leads to an increased relative amount of GliA present in the cell. This was demonstrated when cells near the floor plate, which are exposed to more Shh, showed increased fluorescence when a GliA-GFP construct was introduced. 73 The mechanism of neural tube differentiation has been further elucidated with experiments showing that many of the Shh-responsive cis-regulatory elements in neural target genes are associated with binding sites for homeodomain proteins and SoxB proteins. 74 SoxB proteins are transcription factors active throughout the neural tube, and a fascinating experiment showed that ectopic Sox2 expression in the limb allowed for Shh-dependent activation of neural specific genes. 75 Thus, SoxB proteins are important tissue specific factors that help cells respond to Shh in a temporally and spatially specific manner.
Hedgehog is likely most well-known for its role in limb patterning. The developing limb begins as a small bud of proliferating mesenchymal cells covered by an ectodermal shell. 76 The zone of proliferating activity (ZPA) is a small group of posterior mesenchyme cells in the developing limb bud that is responsible for the correct pattern development. A series of landmark experiments by Saunders and Gasseling 77 heralding the role of the ZPA and morphogens demonstrated that transplanted posterior limb buds created duplicate digits. It was later found that Shh is expressed in the ZPA and mediates the development of the digits and the AP axis of the limb. Shh diffuses from the ZPA and creates a concentration gradient along the limb bud. Chiang et al 78 showed that Shh−/− mutant limb buds did not create duplicate digits when transplanted and showing the critical role of Shh in ZPA patterning. Further studies also showed that the dose and length of exposure to Shh led to specification of digits in the limb. 79 Hedgehog signaling is similarly activated in many other areas of the embryo as well, playing vital roles throughout the body.
Hedgehog in Craniofacial Development
The role of Hedgehog signaling in the development of calvarial and facial bones is a relatively new area of investigation. The craniofacial skeleton develops from a mixture of neural crest and paraxial mesoderm with species-specific differences. The majority of cranial and facial bones form through intramembranous ossification as previously discussed. The face forms from masses of mesenchymal stem cells that are grouped into a frontonasal mass as well as 2 maxillary and 2 mandibular prominences which fuse together. The skull forms from the cranial bones which start as bone plates that grow toward each other until they meet at fibrous junctions known as a sutures. The entire process is regulated by a complex network of interacting pathways, including hedgehog, which is highlighted in Figure 3.60,80
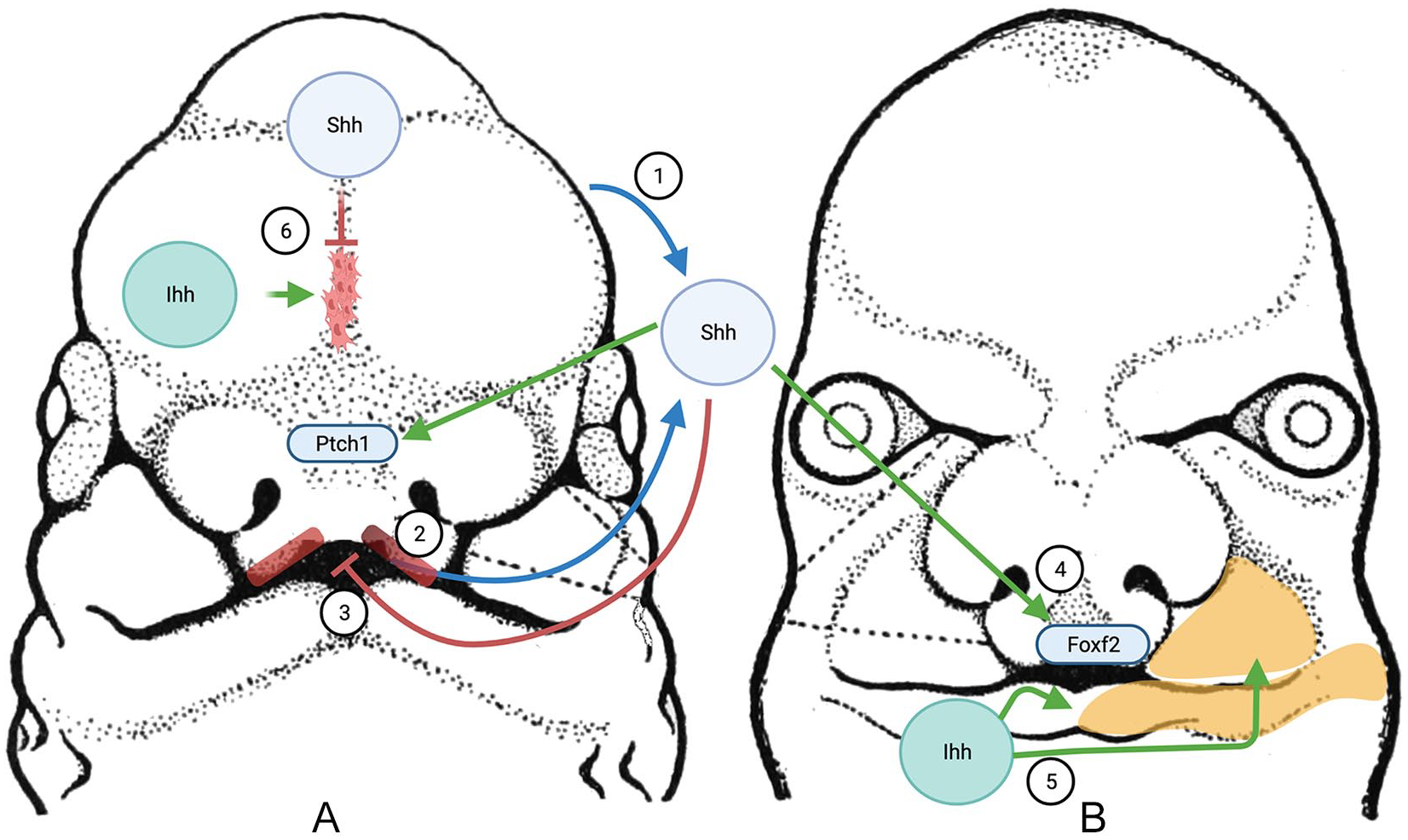
Craniofacial development.
Hedgehog plays an important role in the development and fusion of the facial processes. Shh is initially expressed from the ventral neural tube and diffuses to the developing face. Mesenchymal cells in the frontonasal processes (FNP) receive this Shh ligand which then activates the Hh pathway, which has been shown experimentally by increased Ptch1 expression in these cells. As development proceeds, Shh begins to be expressed in the oral ectoderm. 81 This intraoral Shh-expressing region which lies adjacent to a Fgf8 expressing domain is known as the frontonasal ectodermal zone (FEZ) and plays an important role in the patterning of the FNP and the face.82-85 The correct balance of Shh signaling from the FEZ has been shown to be important in the fusion and formation of the face, with alteration in signaling leading to a variety of clefts.86,87
Interestingly many of the proteins from the pathway that have been linked to craniofacial abnormalities are intraflagellar transport (IFT) proteins, which are an essential link in the hedgehog signaling chain. Mouse genetic screens showed that the IFT protein Kif3 (kinesin superfamily protein 3) is necessary for hedgehog signaling downstream of Ptch and upstream of direct transcription targets. 88 Kif3 is a motor complex that is responsible for importing specific proteins from the cilium to the cytoplasm. Hedgehog signaling passes from Ptch to Smo, and then through the cilium including Kif3 to SUFU/Gli which then activates Gli. 89 Mice with Kif3a deficient cartilage exhibit abnormal hedgehog signaling as well as ectopic cartilage formation, cranial base growth retardation, and dysmorphogenesis. 90 Furthermore, mutations affecting proteins in these pathways, such as DYNC2H1 and IFT44, are associated with craniofacial abnormalities such as HPE and craniosynostosis, demonstrating the role of the intraflagellar transport system in development.91,92
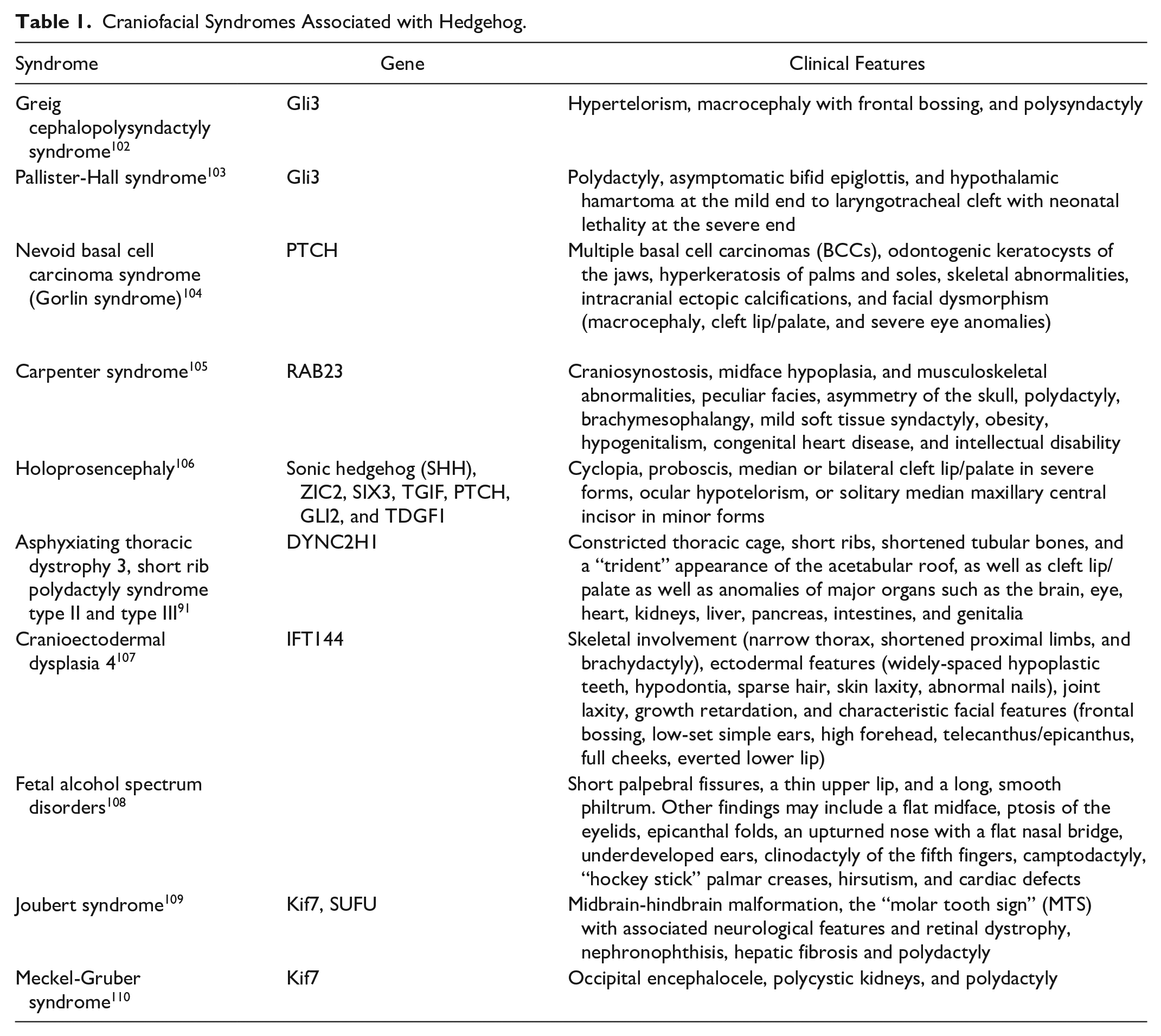
Hedgehog signaling has been implicated in a variety of craniofacial dysmorphisms including Greig cephalosyndactyly and Pallister-Hall Syndrome both of which involve GLI3 mutations.93-95 Many of the syndromes with hedgehog associated mutations are described in Table 1. In addition to the described craniofacial dysmorphisms, Shh has been found to be important in the development of the palate, and mutations in the Shh pathway have been shown to be associated with cleft lip and palate.96,97 Mechanistic details are still under study, but Shh is required for the closure of the upper lip where it induces proliferation of neural crests through the transcription factor FOXF2, a gene which has been found to be associated with cleft lip in genetic screening.98,99 In contrast to lip closure, increased Shh has been shown to inhibit osteogenesis and palatine bone formation leading to a cleft palate phenotype in transgenic mice. 100 These findings show that hedgehog must be kept in a careful balance to allow development to proceed normally. A recent study highlighted the importance of other Hedgehog pathway-related genes in craniofacial development when knockout of the hedgehog co-receptors Boc, Gas1, or Cdon led to craniofacial abnormalities- but deletion of Boc in a Gas1 null background led to rescue of the abnormal phenotype. 101
Craniofacial Syndromes Associated with Hedgehog.
Genetic analysis of families with syndactyly type 1 and craniosynostosis Philadelphia type showed that copy number variations in the conserved non-coding elements of IHH were associated with the developmental abnormalities. 111 In addition to its role in endochondral bone formation a study by Lenton et al 112 showed that Ihh is also responsible for ossification during intramembranous calvarial ossification, and likely regulates Bmp signaling during this process. Conditional, neural crest-specific, Ihh knockouts have an interesting phenotype that includes hypoplastic mandibles, delayed facial cartilage ossification, and a hypoplastic nasomaxillary complex, suggesting that Ihh plays an important role in midface development. 113
A particularly interesting area of research involving hedgehog signaling is the clarification of the pathways involved in cranial suture osteogenesis. The cranial suture system can be thought of as 2 osteogenic fronts with undifferentiated mesenchyme in between, bounded by dura mater inferiorly and pericranium superiorly. Cranial suture closure is regulated by a careful balance between osteoblastogenesis and osteoclastogenesis, and is not yet fully understood. 80 Current research suggests that Shh is expressed in the suture mesenchyme and is responsible for maintaining patency, while Ihh is expressed on the osteogenic bone fronts and is responsible for suture closure. A study of Ihh knockout mice showed decreased calvarial bone size and decreased markers of osteodifferentiation. 112 Other earlier studies similarly showed that Ihh is localized to the bone front and that Ihh promotes osteogenesis in the suture.114,115 In contrast Shh has been found in the suture mesenchyme indicative of a role in maintaining the patency of the suture.95,116 Conflicting data, however, exists. 117
In summary, it is likely that both Ihh and Shh play important roles in craniofacial development as evidenced by the numerous dysmorphisms associated with mutations in either gene. Their complex role in regulating the differentiation of cranial sutures is an active area of investigation. Further understanding of the role of Shh and Ihh in suture homeostasis can potentially be translated to molecular therapies in craniosynostosis and bone tissue engineering.
Role in Cancer
The role of hedgehog in cancer is well established. Dysfunction of the Hh signaling pathway has been tied to many cancer types, both directly through genetic mutations directly affecting expression, as well indirectly through overexpression of Hedgehog activating signals. 44 It is estimated that one third of all malignant tumors are caused in part by aberrant Hh signaling. 118 Dysregulation can occur at many steps, but there are classically 3 models for the mechanism of the Hh signaling pathway in cancer: Type I ligand independent, Type II ligand dependent autocrine, and Type III paracrine ligand dependent. 119 There is also a Type IIIb subtype of reverse paracrine ligand dependent, as well as a cancer stem cell model. 120 Recently, there has been increasing evidence for a non-canonical Hh pathway in certain cancers, which involves Smo-independent activation of Gli. 39 Though there is overlap, each model has been observed in specific cancer types. For example, many basal cell carcinomas are associated with a Type I ligand independent mutation to Ptch or Smo.121,122 In contrast, the Type II autocrine model has an association with small-cell lung carcinoma, liver cancer, and breast cancer.119,123-125 The 3 models of hedgehog signaling in cancer are highlighted in Figure 4.
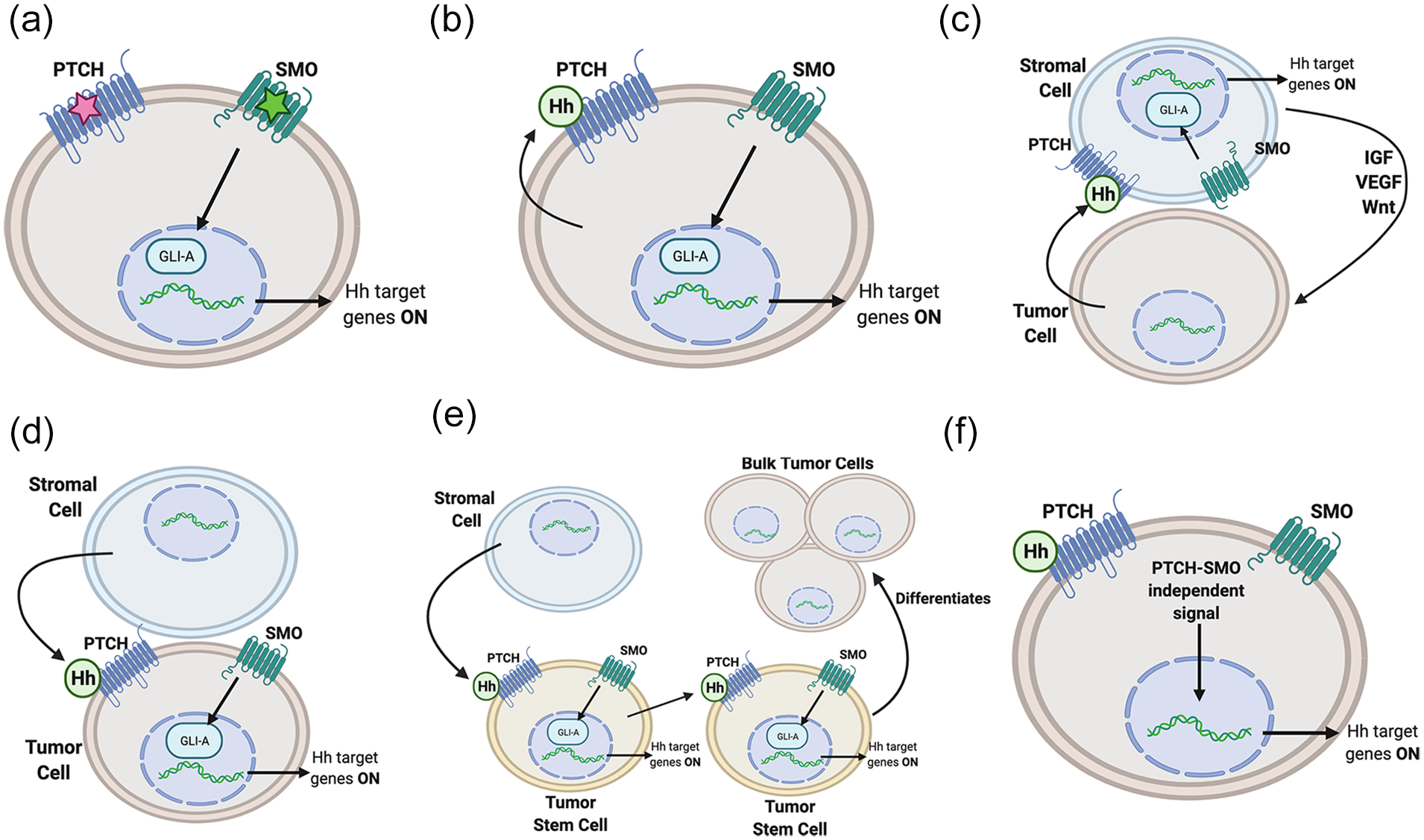
Models of hedgehog (Hh) pathway involvement in cancer: (a) type I: the ligand independent pathway results from either an inactivating mutation (red star) to Patched (PTCH) or an activating mutation (green star), to Smoothened (SMO), bypassing the need for Hh ligand binding, (b) type II: the autocrine/juxtacrine pathway involves an overexpression of Hh, which activates the Hh pathway on the same, or adjacent cells, (c) type III: the paracrine model involves aberrant Hh expression from tumor cells, which activates the Hh pathway on nearby stromal cells. The stromal cells in turn secrete growth/survival signals to the tumor cells, (d) type IIIb: in the reverse paracrine pathway, stromal cells express Hh, which binds the PTCH receptor on nearby tumor cells, (e) cancer stem cell (CSC): like reverse paracrine, stromal cells express Hh, which activates the Hh pathway in CSC’s. CSC’s can then activate the Hh pathway in other CSC’s, or differentiate into bulk tumor cells, and (f) non-canonical: Hh target genes are activated in a pathway independent of PTCH-SMO signaling. Created with BioRender.com.
The Type I ligand independent mechanism of cancer formation involves activation of the Hh pathway without the need for Hh ligand binding, which normally leads to the internalization and degradation of the negative regulator Ptch. This can occur either due to an inactivating mutation to the negative regulators Ptch or Sufu, or an activating mutation in Smo, leading to unregulated expression of GliA target genes. 126 This mechanism was one of the first discovered associations of the Hh pathway and cancer, noted in reduced expression of the negative Hh pathway regulator patched gene in nevoid basal cell carcinoma syndrome (Gorlin syndrome).127,128 Gorlin syndrome is known to cause multiple basal cell carcinomas, among other tumor types. 129 Subsequently, a murine model showed that a sustained Hh signal is not only correlated to basal cell carcinoma, but a requirement for its proliferation. 130 The type I ligand independent mechanism is associated with many other cancers, including medulloblastomas (also associated with Gorlin syndrome), breast cancers, esophageal carcinomas, gastric carcinomas, trichoepitheliomas, bladder carcinomas, and rhabdomyosarcoma.44,131-136
The type II ligand dependent cancer mechanism is an autocrine/juxtacrine pathway that involves overexpression of Hh itself, leading to aberrant stimulation of the Hh pathway. 120 The Ptch receptor is bound, ultimately leading to degradation, by Hh ligands released by either the same cell, or adjacent cells. Many cancers have been found to be associated with this mechanism, including lung, pancreatic, upper and lower gastrointestinal, prostate, breast, melanoma, stomach, esophageal, ovarian, endometrial, and glial tumors.123,137-146 Characterized by Hh overexpression, the type II mechanism can be leveraged as a prognostic marker. For example, in SHH protein-upregulated gastric cancers, SHH expression is correlated to age, tumor differentiation, depth of invasion, staging, and nodal metastasis. 147
The type III mechanism of aberrant Hh signaling is also ligand dependent, but uses paracrine, rather than autocrine/juxtacrine signaling. 120 This involves the secretion of Hh from tumor cells, which binds to Ptch on stromal cells, activating the Hh signaling pathway within the stromal cells; the stromal cells in turn secrete growth and survival signals (IGF, VEGF, Wnt) which signal the tumor cell. This mechanism was first observed in the LNCaP xenograft tumor model, in which tumor cells expressed Shh and activated tumor stroma in a paracrine manner, 148 a finding later corroborated by other mouse and human studies. One such study observed the inability of Hh ligands to activate tumor epithelial cells directly, relying instead on a paracrine feedback loop with the stromal microenvironment.149-151 The paracrine Hh signaling pathway has been shown to be involved with prostate, pancreatic, and colon cancers. 44 In pancreatic cancer, SHH from tumor cells activates pancreatic stellate cells in the stroma to secrete signals that promote, and are possibly necessary for, perineural invasion. 152 In the Type IIIb reverse paracrine model, Hh is secreted from surrounding stromal cells to tumor cells. This subtype has only been described in certain hematological cancers, such as B-cell lymphoma, multiple myeloma, and leukemia.153,154
The cancer stem cell (CSC) model of is yet another mechanism of Hh involvement in cancer. CSCs are a small subset of cells within a tumor that are typically quiescent, but after radiation can differentiate and proliferate, causing tumor recurrence. 155 Hh signaling has been found to be involved in the maintenance of CSCs. 120 Hh signaling in CSCs has been found to be involved in the tumorigenesis of many hematologic and solid tumor types including CML, gastric cancer, breast cancer, colon cancer, small cell lung cancer, and pancreatic cancer.156-162 Multiple myeloma specifically has been found to express Hh ligands to stimulate CSC expansion, and to then undergo Hh pathway blockade for terminal differentiation of CSCs. 163
Non-canonical Hh signaling in cancer is of increasing interest. A Smo inhibitor, such as cyclopamine which inhibit Smo, is commonly used in studies to rule out the canonical pathway when observing non-canonical Hh pathway tumor pathogenesis. 164 Multiple co-carcinogenic pathways, such as the RAS/RAF/MEK/ERK pathway in colon and pancreatic cancer, and PI3K/AKT/mTOR in breast cancer have been shown to overlap with non-canonical hedgehog signaling.165-167 Furthermore, non-canonical signaling has been shown to be involved in multiple steps of cancer progression. One study demonstrated that in human gastric cancer, the Smo-independent Hh pathway was modulated to promote the epithelial mesenchymal transition and invasion. 168
Given the role of the hedgehog pathway in the pathogenesis of many cancers, it is a natural target for cancer therapeutics. The aforementioned role in basal cell carcinoma is targeted by vismodegib and sonidegib (approved in 2012 and 2015, respectively). These SMO inhibitors are used to treat metastatic and locally advanced basal cell carcinomas, as many basal cell carcinomas involve a loss of function of PTCH1, the normal negative regulator of SMO.1,2 While these small molecule inhibitors target the same step in the hedgehog pathway, they have potential differences in pharmacokinetics; however, given differences in their clinical trials (such as criteria for BCC severity) further research is warranted to determine the clinical significance of their differences. 3
Conclusions
Hedgehog signaling plays diverse and important roles throughout the body. First established in embryonic patterning, Hh signaling has been elucidated in cancer, limb and craniofacial development, and osteogenesis. Its intricate tissue- and spatiotemporally-specific regulation of gene networks remain yet to be fully understood. Further investigation into hedgehog signaling is likely to result in exciting new developments in tissue engineering, nonsurgical craniofacial therapies, and cancer therapeutics.
Footnotes
Appendix
Declaration of Conflicting Interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author(s) received no financial support for the research, authorship, and/or publication of this article.