
Abstract
Per- and polyfluoroalkyl substances (PFAS) represent a diverse group of synthetic chemicals widely utilized in industrial processes, consumer goods, food-contact materials, and plastics due to their exceptional chemical stability and resistance to degradation. However, their pervasive presence in the environment, persistence, and ability to bioaccumulate have raised increasing concerns about human exposure and associated health risks. Dietary intake, through contaminated food, drinking water, and food packaging materials, serves as a primary route of exposure, placing the gastrointestinal tract at the center of PFAS–host interactions. Recent research suggests that the gut microbiome plays a significant role in modifying PFAS toxicokinetics, bioavailability, and biological effects. This review consolidates current insights into the complex relationship between PFAS and the gut microbiome, detailing mechanisms such as microbial uptake, bioaccumulation, enzymatic defluorination, β-oxidation-like biotransformation processes, and alterations in bile acid and short-chain fatty acid metabolism. Conversely, exposure to PFAS can disrupt microbial diversity, compromise intestinal barrier integrity, and affect various host functions, including immune response, metabolism, liver and kidney health, and neuroendocrine regulation. Experimental studies, data from human cohorts, and findings from multi-omics approaches reveal that microbiome-driven changes contribute to chronic health issues like inflammation, metabolic disorders, insulin resistance, and immune system dysregulation. Additionally, advancements in metagenomics, metabolomics, and transcriptomics are shedding light on the intricate mechanisms underlying PFAS–microbiome interactions. These developments underscore the potential for microbiome-targeted interventions to mitigate the adverse health effects of these enduring environmental pollutants.
Keywords
Highlights
• The gut microbiome plays a central role in PFAS bioaccumulation and biotransformation. • Microbial enzymes mediate partial defluorination and β-oxidation–like PFAS degradation. • PFAS exposure disrupts gut microbial diversity and short-chain fatty acid metabolism. • Multi-omics integration links microbiome shifts to host metabolic and immune pathways. • Microbiome-based therapeutics offer emerging strategies to mitigate PFAS toxicity.
Introduction
Global prevalence and environmental sources of PFAS
Per- and polyfluoroalkyl substances (PFAS) represent a broad group of synthetic fluorinated compounds that have been produced and utilized extensively since the mid-20th century. Due to their remarkable thermal stability, chemical resistance, and surfactant properties, PFAS have found applications in a wide range of industrial and consumer products. 1 However, their extensive production and unparalleled environmental persistence have led to widespread contamination across aquatic, terrestrial, and atmospheric systems on a global scale. These substances have been identified in surface and groundwater, drinking water supplies, soil, sediments, wildlife, and human populations worldwide even in remote areas far removed from direct industrial activity. 2 As a result, PFAS are now categorized as contaminants of global concern and are often called “forever chemicals” because they resist environmental degradation. A key feature of PFAS lies in their multiple carbon–fluorine (C–F) bonds, recognized as some of the strongest covalent bonds in organic chemistry. These bonds’ exceptional strength and stability make PFAS highly resistant to thermal, chemical, and biological breakdown. 3 This resilience is a major factor behind their long-lasting presence in the environment and tendency to accumulate in living organisms. As a result, PFAS are commonly known as “forever chemicals,” given their ability to persist in ecosystems and biological systems for prolonged durations. 4
Key sources of PFAS pollution include fluorochemical manufacturing plants, aqueous film-forming foams (AFFFs), wastewater treatment facilities, landfill leachates, the use of biosolids, consumer products, and food-contact materials. Studies on human exposure have consistently found PFAS in blood, urine, breast milk, umbilical cord blood, and various tissues, underscoring the widespread nature of exposure across all demographics and age groups. These findings have sparked significant concern about the potential health risks associated with prolonged, low-level exposure to these persistent chemicals. 5
PFAS in plastics, food packaging, and emerging alternatives
The widespread use of PFAS in food-contact materials and packaging has garnered significant attention due to its direct link to human exposure. Historically, these chemicals have been incorporated into a variety of products such as paper-based packaging, molded fiber items, fast-food wrappers, microwave popcorn bags, bakery packaging, disposable containers, and certain plastics to impart resistance against oil, grease, and water. Their extensive utilization across the packaging industry has led to broad environmental dissemination. 1
Mounting toxicological and epidemiological findings have driven regulatory bodies around the world to restrict or phase out several older PFAS compounds, including perfluorooctanoic acid (PFOA) and perfluorooctane sulfonate (PFOS). In turn, manufacturers have shifted toward short-chain PFAS and alternate fluorinated substances, such as hexafluoropropylene oxide dimer acid (GenX) and other substitute chemicals. Yet, emerging studies indicate that many of these replacements may share characteristics with legacy PFAS including long-term environmental persistence, high mobility, and biological activity creating growing concerns about their safety and sustainability over time. 6
Entry of PFAS into the food chain and human exposure
PFAS infiltrate the food chain through various environmental pathways. These include industrial discharges, polluted water sources, atmospheric deposition, the application of biosolids, and the breakdown of PFAS-containing consumer products. Once introduced, these harmful compounds can accumulate in agricultural produce, livestock, fish, shellfish, and other food items consumed by humans. Additionally, PFAS can migrate from food packaging materials into the food itself, a process that is particularly exacerbated under conditions of high temperatures, extended storage periods, or the presence of high-fat food products. 7
Among the general population, dietary intake is regarded as one of the primary routes of PFAS exposure. When ingested, these substances are readily absorbed by the gastrointestinal tract and distributed across the body through the bloodstream. Owing to their strong affinity for binding with serum proteins and their resistance to metabolic breakdown, many PFAS tend to have long biological half-lives, leading to their accumulation in tissues such as the blood, liver, and kidneys. Notably, the gastrointestinal tract serves as the first point of contact for PFAS after ingestion. As a result, the gut microbiome plays a pivotal role as a critical interface between external PFAS exposure and the body’s physiological responses. 8
Aim and scope of the review
The gut microbiome is increasingly recognized as a critical factor in mediating the toxicity of environmental chemicals, such as PFAS. While extensive research has explored the toxicological effects and environmental distribution of PFAS, the intricate interplay between these substances and the gut microbiome remains poorly understood. This review aims to address this gap by presenting a detailed synthesis of current knowledge on microbial interactions with PFAS, including processes like bioaccumulation, biotransformation, defluorination, microbiome dysbiosis, changes in microbial metabolites, and their potential impacts on human health. Additionally, it highlights emerging multi-omics approaches that offer deeper insights into PFAS–microbiome dynamics, as well as investigates microbiome-based therapeutic interventions designed to counter PFAS-related toxicity. By drawing upon findings from environmental, microbiological, toxicological, and clinical research, this review strives to establish a holistic perspective on the role of the gut microbiome in mediating PFAS exposure and its associated health consequences.
Sources of human exposure to PFAS
Human exposure to PFAS originates from various environmental and consumer-related sources such as contaminated drinking water, food, indoor dust, occupational environments, and the use of products containing PFAS. Among these, dietary exposure is recognized as one of the primary pathways for PFAS intake in the general population. Beyond contamination of food and water in the environment, food-contact materials have become a significant, ongoing source of exposure due to their widespread application and direct contact with food. 9 Historically, PFAS have been incorporated into a multitude of commercial products because of their remarkable resistance to heat, moisture, oil, and chemical breakdown properties that have made them common in food packaging and plastic materials. Food packaging, in particular, is a notable contributor to PFAS exposure. These substances are often applied to items such as paper, paperboard, molded fiber products, microwave popcorn bags, fast-food wrappers, bakery packaging, and disposable food containers to provide grease-proof and water-resistant properties. 10 Studies have frequently detected both legacy and emerging PFAS compounds in food-contact materials sold worldwide. Critically, these compounds are not always tightly bonded to the packaging materials and can migrate into food during processes like storage, transportation, or heating. Factors such as temperature, storage duration, food type, and packaging attributes influence this migration, with fatty foods generally absorbing more PFAS. Consuming packaged and processed foods regularly may thus lead to a gradual buildup of PFAS in the body over time. 11
The production and disposal of plastic materials are another indirect yet significant source of PFAS exposure through environmental contamination. During the manufacturing of polymers or fluoropolymers for plastic products and packaging, PFAS may either be intentionally added or introduced via processing aids. 12 Over the lifecycle of these materials from usage to weathering, recycling, or degradation PFAS can leach into soils, surface waters, groundwater, or aquatic ecosystems. These contaminants can enter the food chain via crops, seafood, livestock products, or drinking water. Consequently, the contamination associated with plastics poses not only a consumer product challenge but also a pressing environmental health issue, contributing to sustained PFAS exposure across populations. 7
From the lens of host–microbiome interactions, exposure to PFAS derived from food packaging holds particular importance since ingestion is the main pathway for these chemicals to enter the human body. After oral intake, PFAS interact directly with the gastrointestinal tract and gut microbiota before entering systemic circulation. 9 Recent findings suggest that prolonged dietary ingestion of PFAS can alter gut microbial composition, impair short-chain fatty acid production, disrupt bile acid metabolism, and compromise the intestinal barrier’s integrity. These microbiome disruptions can further affect PFAS absorption, bioavailability, toxicokinetics, and vulnerability to negative health effects. Therefore, identifying food packaging and plastic materials as key sources of PFAS exposure is crucial for better understanding how environmental contamination contributes to gut microbiome imbalance and heightened risks for human health complications. 13
Gut microbiome-mediated biotransformation of PFAS: Linking microbial pathways to human metabolic health
A few environmental bacteria have been reported to attach to PFAS compounds. 14 Bacteroides uniformis demonstrated significant PFAS uptake, reaching millimolar (mM) levels inside the cells, while maintaining normal growth. At very low nanomolar concentrations (0.34 nM), Bacteroides uniformis absorbed about 37% of perfluorononanoic acid (PFNA) from the surrounding medium, resulting in an approximately 50-fold increase in its intracellular concentration. At higher concentrations, the rate of PFAS accumulation rose with increasing chain length, with nearly complete uptake observed for longer-chain compounds (>10 carbons). These findings suggest that bacterial cells possess a strong ability to absorb and concentrate PFAS from their environment, even when present at naturally low contamination levels. 14 Cryogenic focused ion beam secondary ion mass spectrometry verified that the PFAS compound PFNA was located inside E. coli cells. The results indicate that E. coli strains use a TolC-dependent mechanism to limit PFAS bioaccumulation. 14 According to one study, mice colonized with human gut bacteria showed higher levels of PFNA in their feces than germ-free controls or mice colonized with low-bioaccumulating bacteria. Taken together, the findings reveal a high bioaccumulation capacity of PFAS in gut bacteria. 14
Certain environmental bacteria, including Pseudomonas strains obtained from areas polluted with PFAS, have been shown to accumulate sulfur-containing PFAS compounds like perfluorohexane sulfonate (PFHxS). Nevertheless, the accumulation occurred at a slow rate, and pretreatment with a solvent was necessary to enable effective separation. 15 PFAS precursors may eventually break down into perfluoroalkyl acids (PFAA). 16 Perfluorooctanoic acid (PFOA) and perfluorooctane sulfonic acid (PFOS) are persistent bioaccumulative chemicals that threaten human health and ecosystems. Their widespread presence and toxic potential raise serious environmental concerns. 17 Hence, finding effective ways to break down these PFAS compounds has become increasingly critical. The C–F bond ranks as the most robust covalent bond in organic molecules. 18 Therefore, breaking down perfluoroalkyl substances through biodegradation is challenging and typically restricted to compounds or molecular segments that lack complete fluorination, such as fluorobenzene, fluoroacetate, perfluorohexylethanol, and perfluorohexyl sulfonate. 19 Some bacteria can convert or defluorinate organic compounds, including PFAS, through various metabolic processes. However, unlike other organic compounds, PFAS are primarily degraded by only a few bacterial species, many of which rely on communal metabolism, where additional growth substrates are provided. 20 A key factor in the degradation of PFAS is the ability of bacteria to tolerate fluoride toxicity. For example, strains of Pseudomonas, such as P. parafulva and P. aeruginosa, are known for their fluoride tolerance and ability to biotransform PFAS. 21
Other bacteria, including strains of Gordonia, Acidimicrobiom, and Pseudomonas plecoglossicida, have also shown potential to biotransform PFAS. 22 Additionally, some strains, such as Pseudomonas PS27 and PDMF10, can bioaccumulate PFAS and reduce PFHxS concentrations by up to 32%. 23 However, to date, no gut-associated bacteria capable of degrading PFAS have been identified, although the possibility of discovering such bacteria in the future remains. 24 The findings suggest that the association of PFAS microbiota may vary across the lifespan. 24 The biodegradation of PFOS by Pseudomonas plecoglossicida was examined, and the results demonstrated that this strain is capable of utilizing PFOS as a sole carbon source, leading to its transformation into perfluoroheptanoic acid (PFHpA) with the concomitant release of fluoride ions. 25 When primary bile acids (BAs) are released into the gastrointestinal tract, they undergo biotransformation (glycine or taurine deconjugation, dehydroxylation, dehydrogenation, and epimerization) by gut microbes to form secondary BAs.26,27 PFOS can interfere with the biosynthesis of BAs by suppressing the expression of 7 alpha hydroxylase (CYP7A1), which is the first and most limiting step in the formation of BAs from cholesterol. 28
Total serum cholesterol levels are positively correlated with levels of the two main PFAS, PFOS and PFOA.
29
The metabolic homeostasis of the host microbiota can be altered by exposure to environmental toxicants (ETs). For example, exposure to dioxins has been shown to disrupt the gut microbiota and induce hepatotoxicity in mice by increasing total BAs and short-chain fatty acids (SCFAs) while decreasing farnesoid X receptor (FXR) signaling in the liver.
30
Exposure to ETs is associated with altered BA metabolism, gut microbiome dysbiosis, insulin resistance, and obesity.
31
These C6-hydroxylated BAs were associated with a gut microbial community composed of Clostridia spp.
31
Exposure to ETs has been associated with increased levels of circulating microbiome-derived lithocholic acid (LCA) and decreased serum ursodeoxycholic acid (UDCA) levels, suggesting that ETs may induce dysbiosis in the gut microbiome that affects the conversion of secondary BAs. Serum BA storage is regulated by hepatic FXR, which plays a pivotal role in regulating BA synthesis and secretion, as well as in regulating lipid and glucose homeostasis in the liver.
32
Men who are particularly exposed to PFAS are more susceptible to metabolic dysregulation. However, mediation analysis of the effect of PFAS on homeostatic model assessment for insulin resistance (HOMA-IR) and gut-derived BAs was stronger in women than in men, suggesting that this could be one of the factors behind sex-specific differences in the impact of PFAS exposure.
33
In a study, it was shown that the LCA/UDCA ratio was increased in a humanized peroxisome proliferator-activated receptor alpha (PPARα) mouse model exposed to PFOA. Furthermore, personalized community modeling of the gut microbiota identified downregulation of the 7α-hydroxysteroid dehydrogenase pathway in the high-chemical exposure groups, which is involved in the bioconversion of chenodeoxycholic acid (CDCA) to UDCA. Several bacterial species from the genera Bacteroidetes, Clostridium, Ruminococcus, and Eubacterium were positively associated with this pathway. In this context, it is interesting that a higher abundance of Ruminococcus and Bacteroidetes has been associated with obesity and other metabolic diseases.
34
Figure 1 provides an overview of the proposed pathways connecting PFAS exposure to changes in the gut microbiome and their impact on host health. The key mechanisms highlighted in the literature include microbial dysbiosis, compromised intestinal barrier integrity, immune system dysregulation, altered metabolite production, and cascading effects on various organ systems. PFAS-induced disruption of intestinal and hepatic metabolic homeostasis.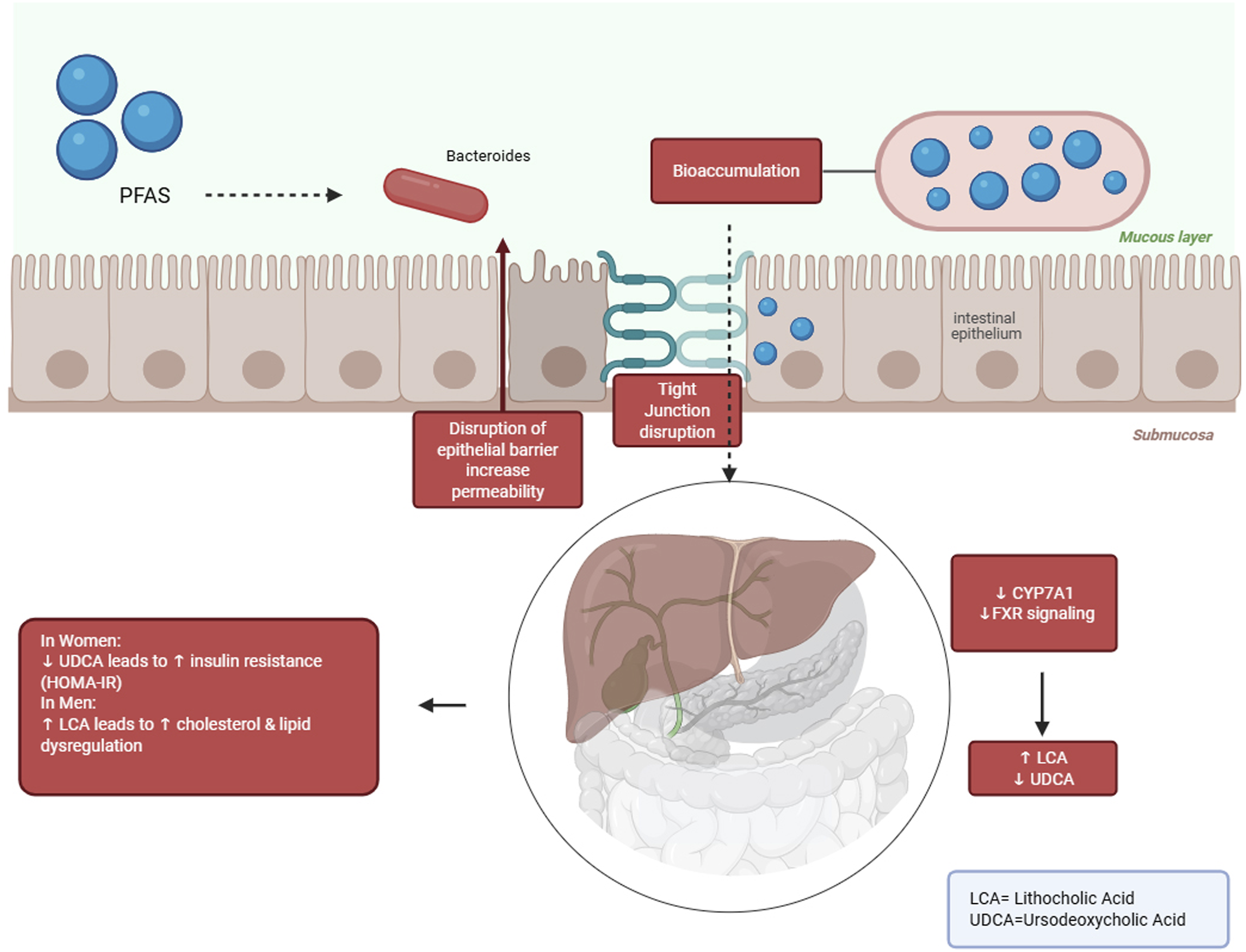
PFAS bioaccumulate in intestinal epithelial cells and gut microbiota (e.g., Bacteroides, Clostridium, Ruminococcus), then through disruption of tight junctions increase the intestinal epithelial barrier permeability. These changes disrupt FXR signaling pathway in liver and also decrease CYP7A1 expression. As a result, bile acids metabolism altered and lithocholic acid (LCA) level rise and ursodeoxycholic acid (UDCA) level decreased. The metabolic consequences of this process are sex-dependent. So, in women increases insulin resistance and in men results in lipid metabolism disruption.
PFAS exposure and the gut microbiome: Mechanistic links to host immune modulation
PFAS exposure can cause neurotoxicity, hepatotoxicity, reproductive toxicity, and immunotoxicity. 35 A cohort study found that serum PFOA concentration was positively correlated with ulcerative colitis, an inflammatory bowel disease (IBD). 36 Perfluorooctane sulfonate disrupted mitochondrial structure, increased the accumulation of reactive oxygen species (ROS), suppressed the levels of superoxide dismutase (SOD) and glutathione peroxidase (GSH PX), and increased the levels of malondialdehyde (MDA) and nitric oxide synthase (NOS). PFOS-induced inflammation was shown to be associated with the activation of NOD-like receptor protein 3 (NLRP3), cleaved cysteine-aspartic acid protease (caspase) 1, tumor necrosis factor (TNF)-α, and interleukin (IL)-1β. In addition, PFOS exposure significantly decreased the ratio of B-cell lymphoma (Bcl2)/Bcl2-associated X (Bax) and increased the expression of cleaved caspase 3 protein Ref. 37.
Currently, the effects of PFSA on the immune system have been shown to reduce antibody responses to vaccines. 38 In vitro, changes in cytokine release, inflammatory activation, and metabolic pathways have been observed in cells exposed to various PFAS. During infancy, PFAS exposure has been associated with several adverse health effects, including reduced response to vaccination, impaired lipid metabolism, and reduced birth weight. 39 Since the composition of the infant’s gut microbiota changes with age, the impact of PFAS on the gut microbiota may also change with age. 24 Exposure to PFAS during pregnancy was associated with lower predicted activity in vitamin B6 metabolism, lipoic acid metabolism, and amino sugar and nucleotide sugar metabolism. 40 Among the studies reviewed, only one focused on SCFA levels in infants. This research found that PFOA exposure was associated with increased concentrations of acetic acid and propionic acid in Norwegian infants at 1 month old. 41 PFAS also affect bacteria by inducing oxidative stress. This occurs when the production of reactive oxygen species (ROS) surpasses the bacteria’s ability to counteract them, leading to tissue breakdown, DNA damage, and ultimately, cell death. Research has demonstrated that exposure to PFOA and PFOS elevates intracellular ROS levels, indicating that PFAS might intensify oxidative stress within bacterial communities. 42
PFAS have been linked to reproductive and developmental toxicity, with evidence suggesting their association with genotoxic effects. Research indicates that exposure to these substances can elevate oxidative stress and produce more reactive oxygen species (ROS). 43 This, in turn, may lead to DNA damage, genomic instability, and hinder the cellular repair processes. Sustained oxidative injury could further result in cellular dysfunction and heighten vulnerability to various adverse health outcomes. Beyond its genotoxic effects, PFAS exposure significantly impacts immune and inflammatory responses. 44 Experimental findings highlight that oxidative stress induced by PFAS activates inflammatory pathways, including the NLRP3 inflammasome, and triggers the release of pro-inflammatory cytokines like TNF-α, IL-1β, and IL-6. These changes can disrupt immune balance and potentially lead to chronic inflammation and progression of related diseases. 45 Considering the pivotal role of NF-κB signaling in intestinal inflammation, growing evidence suggests that heightened activation of the TLR4–NF-κB pathway could exacerbate pro-inflammatory cytokine production, further advancing gut barrier dysfunction linked to chronic inflammatory disorders.46,47
Increased consumption of surfactants may affect gastrointestinal permeability. Disruption of intestinal tight junctions is a common feature of autoimmune intestinal diseases such as IBD. When this barrier is compromised, an immune response to environmental antigens that cross the intestinal mucosa may be elicited, leading to autoimmune intestinal diseases such as celiac disease or food allergies. There is evidence suggesting a link between PFAS and ulcerative colitis in human adults.
48
Increased serum levels of total PFAS (compared to controls) have been observed in Swedish patients with late-onset ulcerative colitis (≥55 years), but not in patients with Crohn’s disease.
49
Experimental data on immunomodulatory effects in the gastrointestinal system show that non-cytotoxic concentrations of PFOS (100 µM, as defined by the authors based on cell viability assays) compromise tight junction integrity in Caco-2 cell monolayers.
50
Increased intestinal permeability can expose subepithelial cells, such as fibroblasts, to luminal contents, including chemicals. Prolonged exposure (96 h) of human colonic myofibroblasts (CCD-18Co) to low levels (≤1 µM) of PFOS and PFOA promotes cell proliferation in vitro, a process linked to intestinal inflammation.
51
Several PFAS are known to be immunosuppressants and, therefore, could be thought to have anti-inflammatory properties. The anti-inflammatory effects of several PFAS have been suggested in association with hepatotoxicity and neurotoxic effects. There is evidence that both PFOA and PFOS exposure suppress the release of proinflammatory chemokines such as CXCL2, IL-8, CXCL9, and CXCL10 in several human cell lines.
52
Growing evidence indicates that PFAS exposure plays a role in disrupting the intestinal barrier and triggering chronic inflammation. Alterations in the composition of gut microbes can impact the host’s ability to resist enteric pathogens by modifying antimicrobial defense mechanisms and disrupting intestinal microbial balance. This highlights the critical role of microbiome integrity in preserving gut health.
53
Figure 2 demonstrates how PFAS can build up within intestinal epithelial and immune cells, stimulate inflammasome signaling pathways, and facilitate the secretion of pro-inflammatory cytokines. Together, these processes enhance intestinal permeability, disrupt microbial balance, and perpetuate ongoing inflammatory reactions. Mechanistic illustration of PFAS-induced intestinal barrier dysfunction and immune dysregulation.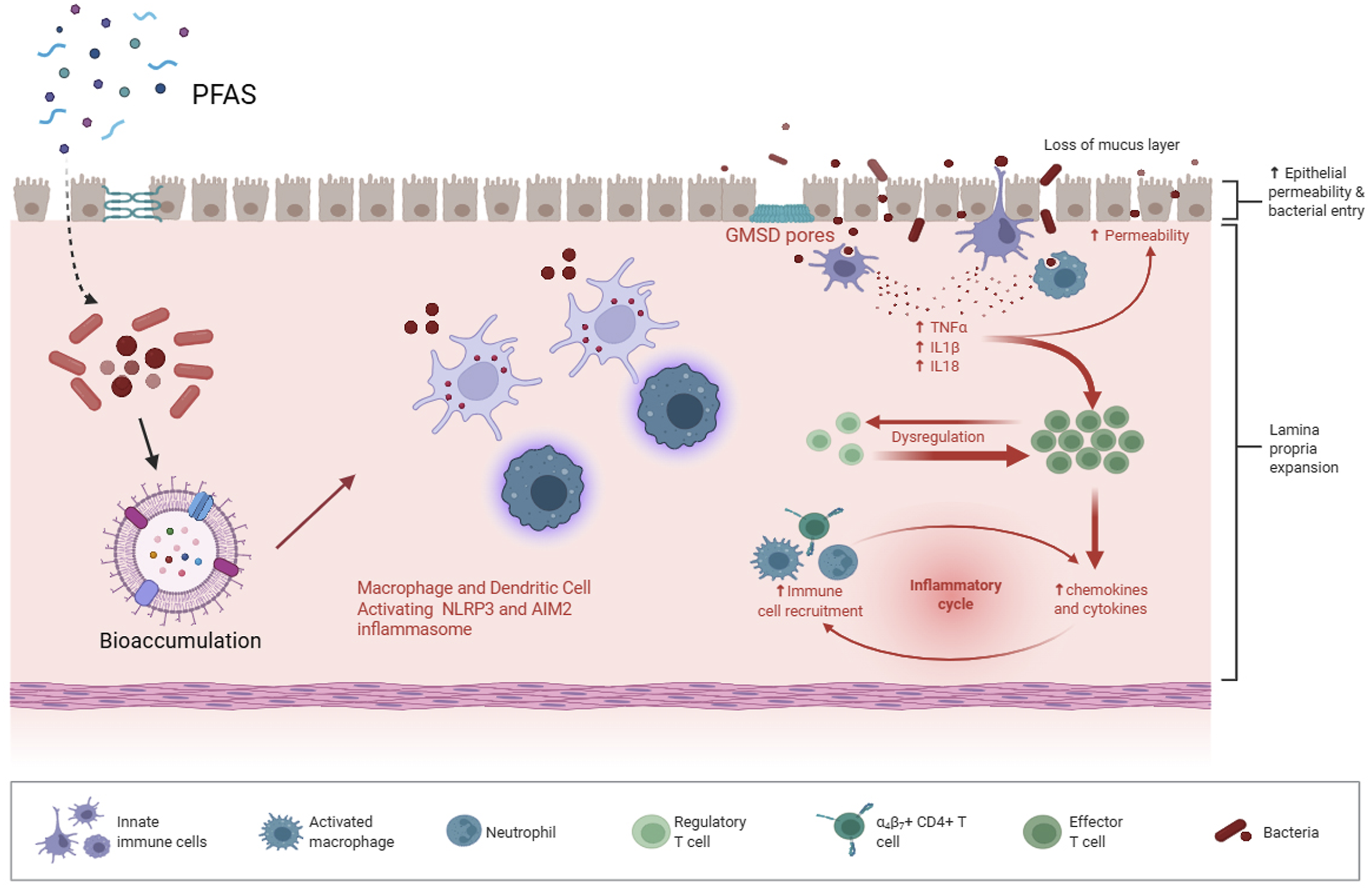
PFAS trigger immune responses via intestinal epithelial and immune cells bioaccumulation and macrophage and dendritic cell activation by NLPR3 and AIM2 inflammasomes. This process leads to proinflammatory cytokines (TNFα، IL1β، IL18) and chemokines release that results in immune cells recruitment and lamina propria expansion. Disruption of mucosal layer integrity and GMSD pores formation, increase permeability and bacterial entrance. So, a chronic inflammatory cycle has been formed that results in dysbiosis and epithelial barrier dysfunction.
The role of gut microbial enzymes in PFAS biotransformation
Previous studies have focused on fluorine atoms, which are often incorporated into pharmaceuticals and agricultural chemicals because of their ability to improve bioavailability and increase the half-life of the compounds. Fluorine atoms reach our gut microbes indirectly through, for example, highly fluorinated compounds such as PFAS which can accumulate in our gut bacteria. 54 Although certain soil bacteria possess enzymes that enable defluoridation, it remains unclear whether such enzymes exist in the human gut microbiome. Enzymes typically involved in defluoridation include hydrolases from the α/β-hydrolase superfamily and members of the haloacid dehalogenase-like (HAD) superfamily. While many HAD enzymes catalyze only dechlorination reactions, the specific structural features responsible for defluoridation activity in some members of this group are still not fully understood. 55
Aerobic biotransformation of PFAS has been observed for 5:3 polyfluorinated carboxylic acid (PFCA) and n:2 FTOHs, with evidence of fluorine removal indicated by the formation of shorter-chain perfluorinated acids. 56 Pathways such as beta-oxidation were proposed for the biodefluorination of short-chain PFAS by aerobic sludge communities based on the removal of the parent compound, identification of the transformation product, as well as fluoride release. 57 Enzymes capable of cleaving the carbon-fluorine bond have been found to be widely present across bacterial phyla commonly found in the human gut. Earlier research demonstrated that HADs, originating from human gut microbiota, possess the ability to facilitate this reaction. 58 Studies of fluorinated substrate analogs that inhibit alanine racemases of pyridoxal phosphate (PLP)-dependent enzymes, as well as other classes of enzymes, have advanced the discovery and characterization of enzyme catalytic mechanisms and their physiological roles. A notable example is the discovery of an (S) α-fluoromethyltyrosine inhibitor of a microbial PLP-dependent tyrosine decarboxylase. This previously unknown microbial enzyme converts levodopa, a drug used to treat Parkinson’s disease, to dopamine, thereby contributing to interindividual variation in drug efficacy. 59
Activation of short-chain PFAS initiates biotransformation by adding coenzyme A to the molecules, followed by beta-oxidation, which shortens the carbon chain of the polyfluorinated compounds. Fluorine on the terminal carbon was removed via oxidative HF elimination and hydrolytic defluorination. 57 The 1:5, 2:4, and 3:3 FTCAs were converted into FTCA-CoAs by a long-chain acyl-CoA synthetase derived from Gordonia sp. strain NB4−1Y, providing evidence for the proposed β-oxidation–like defluorination pathway that begins with CoA activation. 60 It was suggested that 3-(trifluoromethoxy) propanoic acid is defluorinated through the same beta-oxidation pathways as short-chain PFAS. 57 This process was subsequently accompanied by similar hydrolysis and spontaneous defluorination. The PFAS compound di (polyfluoroether thioether(S)-oate) sulfonate (diFESOS), which contains both ether and thioether linkages, was shown to degrade into 2-((1,1,2-trifluoro-2-(heptafluoropropyloxy)ethyl)thio)ethan-1-ol and 2-((1,1,2-trifluoro-2-(heptafluoropropoxy)ethyl)thio)acetic acid (FESCA) when exposed to activated sludge in a sulfate-free defined medium. The discovery of the fluorinating enzyme as well as the proposed reductive C F aryl bond cleavage via a Birch-like reductive mechanism sheds light on the understanding of PFAS biodefluorination. 61
Molecular dynamics simulations revealed that the C-terminus of gut microbial HAD proteins exhibits greater flexibility than other protein regions. It was suggested that the C-terminus of HAD proteins, along with “function enhancement” through the design of a C-terminal chimera, plays an important and previously unreported role in defluorination. This knowledge lays the foundation for engineering gut microbial proteins with broad substrate domains for carbon-fluorine bonds, opening up new applications in human health and the environment. This adds a prokaryotic dimension to the “C-terminus” to more accurately predict and modulate enzyme functions in microbiomes and across domains of life. 62 In a study investigating enzymes that catalyze the cleavage of the carbon-fluorine (CF) bond, the focus was primarily on a limited number of native microbial hydrolases that are reactive with the natural product fluoroacetate. Identification and engineering of new enzymes is essential for the biodegradation of commercially available fluorinated compounds, many of which are known as per- and polyfluorinated alkyl substances. For example, some hydrolases react with –CF2 moieties, a common function group of PFAS compounds. Enzymatic degradation mechanisms for excess CF catalyzed by reductases, lyases, and oxygenases have been identified through screening. Screening and evolution of PFAS defluorination in bacteria are inhibited by the forced release of toxic fluoride from CF fission. Engineering greater fluoride tolerance in bacteria is a problem that needs to be addressed, along with enzyme improvement. 63
Microbiome-driven mechanisms of PFAS metabolism and their impact on chronic disease risk
PFAS, traditionally regarded as highly persistent contaminants with minimal biodegradability, are now increasingly recognized for their potential interactions with the gut microbiome, which may affect their environmental behavior and biological impact. 64 Before investigating into the specific microbial taxa and enzymatic pathways involved in PFAS transformation, it is essential to acknowledge the gut microbiome’s role as a dynamic metabolic interface between environmental factors and host physiology. By influencing bioaccumulation, adsorption, biotransformation, and host metabolic pathways, intestinal microorganisms can potentially modify PFAS bioavailability, persistence, and toxicity. Consequently, exploring these microbiome-mediated interactions is critical for understanding the potential health implications associated with PFAS exposure. 65
Microbial enzymes can bio transform environmental chemical substances and affect their absorption, defecation, and toxicity, although xenobiotics also can change combination and function of microbiome reciprocally. 66 Recent evidences show that PFAS can interact with gut microbial ecosystem and raising the possibility that PFAS-microbiome interactions can influence host metabolic, immunological, and organ-specific pathways. 54 Recent research suggests that plant-derived tannins contribute to improved antioxidant levels, bolster immune function, fortify the intestinal barrier, and influence gut microbiota composition. These findings emphasize the critical role dietary polyphenols play in supporting the balance between the host and its microbiome. 67 These interactions may occur through several pathways including bioaccumulation, active transport and efflux pumps, biotransformation of PFAS or their precursors, and modulation of host homeostasis. 54
PFAS exposure can act as a selective pressure on gut microbial ecosystems and can change microbiome structure and function over time. In rodent models, PFOS and PFOA exposure result in microbial diversity reduction. This diversity reduction usually accompanied with decline in population of beneficial species such as Bifidobacterium and Lactobacillus and also increase in opportunist species that can grow in imbalance conditions. 68 Human cohort studies showing exposure-correlated shifts in microbiome composition and change in their metabolite profiles. 69
The microbiome changes caused by PFAS exposure have widespread consequences for organs and systems of the body. Metabolic disruptions are one of the main concerns. The rodent models show that dysbiosis induced by PFAS correlates with insulin resistance, increased adiposity, and lipid metabolism disruption.
68
These changes probably occur because of reduction in SCFAs production, changes in bile acids composition, and dysregulation of signaling pathways related to energy metabolism. Recent advancements in receptor biology have shed light on how signals from the extracellular microenvironment can drive biased GPCR signaling. This discovery offers a mechanistic basis for understanding how metabolites produced by the microbiome might impact immune function and metabolic responses in the host.
70
Recent findings indicate that the loss of certain beneficial gut microbes and their bioactive metabolites may play a role in metabolic dysfunction. This underscores the idea that microbiome-derived metabolites serve as crucial mediators connecting environmental factors to the host’s metabolic health.
71
In humans, epidemiologic evidences show that prenatal exposure to PFAS can alter the maternal microbiome during pregnancy. These changes can affect fetal development and early-life metabolic programming which in turn may increase risk of diabetes and obesity in life.69,72 Figure 3 summarize the proposed mechanism by which maternal exposure to PFAS may impact fetal development through changes in the gut microbiome. PFAS-induced microbial imbalances result in reduced SCFA production and heightened inflammatory signaling, potentially leading to placental dysfunction and disruptions in the developmental programming of the offspring. Conceptual model of maternal PFAS exposure and microbiome-mediated developmental effects.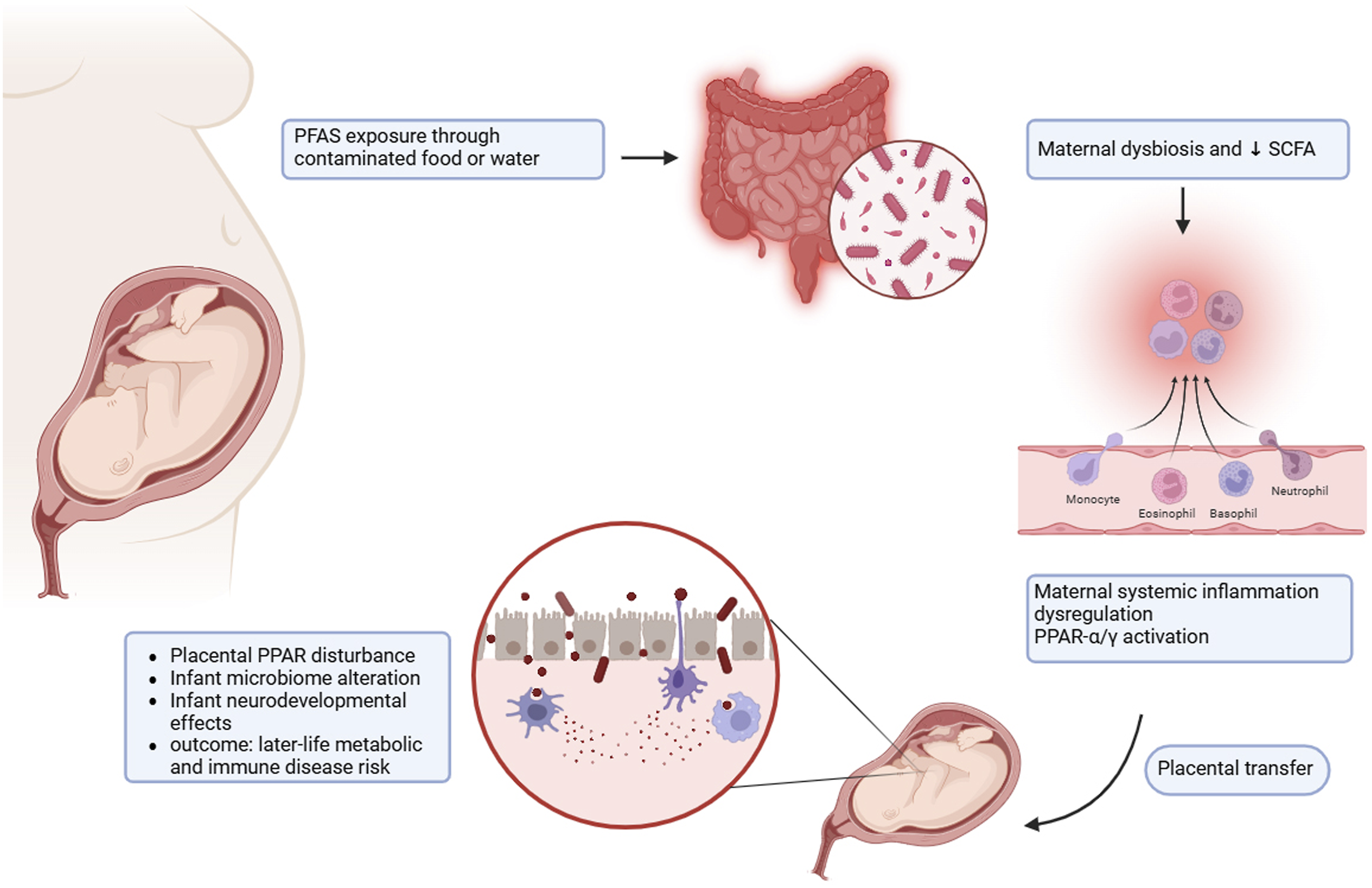
PFAS exposure through contaminated food or water leads to maternal gut microbiome disruption and reduce SCFAs production. This process causes systemic inflammation and also activation of PPAR-α/γ signaling pathways. PFAS and inflammatory mediators pass through placenta and result in placental dysfunction, infant dysbiosis, and infant’s neural and metabolic growth disruption. This changes in early years of life increase the risk of chronic diseases in later years.
Immune system modulation is another main pathway that is affected. PFAS can disrupt the interconnection between the gut microbiome and immune system. Especially, SCFAs reduction weakening the gut microbiome ability to induce regulatory T cell differentiation. These cells have a key role in inflammatory response inhibition and their reduction leads to rise in inflammation, autoimmune defects, and even exacerbation of allergic diseases. 73 In addition, changes in microbiome composition and their metabolic products can participate in carcinogenesis. For example, rise in secondary bile acids production or increase in oxidative stress caused by microbiome result in DNA damage, altered cellular signaling, and facilitate tumor growth and development. 74 Microbiota-derived metabolites possess the ability to provide systemic protective effects by influencing inflammatory and cell death pathways. This underscores the broader significance of gut microbial products in reducing chronic disease risks linked to environmental toxicant exposure. 75 Dietary bioactive compounds play a vital role in boosting antioxidant capacity, strengthening immune function, and positively influencing gut microbial communities. This highlights the intricate connection between microbiota composition and the metabolic health of the host. 76
Kidney and liver outcome are also significant. PFAS has a high chemical affinity to plasma proteins and cell structures that lead to its accumulation in liver tissue. This accumulation results in changes in the flow of microbiome-derived metabolites like lipids and bile acids that in animal models exacerbate hepatotoxicity. 77 In human studies, the interconnection between high levels of PFSA and changes in the microbiome composition and reduced function of kidneys have been reported. These findings show that the gut microbiome can act as a mediator for PFSA toxic effects and also can use as a biomarker of individual sensitivity. 78 The combination of these evidences highlight gut-liver-kidney axis through which PFAS can affect chronic disease risk. A pathway in which the microbiome changes can determine the severity, type, and duration of consequences causing by accumulation of these components in the body. Understanding of mechanism of PFAS influence on microbiome and host has been achieved via a set of complementary experimental approaches. In animal models, using germ-free mice and also antibiotic-treated animals show that in absence of gut microbiome, PFAS exposure because more severe metabolic effects such as energy dysregulation, increase in insulin resistance, and severe changes in lipid profiles. These findings indicate that the microbiome have an influence on PFAS toxic effects modulation and can change or decrease their toxic load. 68 Ex vivo gut fermentation models show that PFAS can directly affect growth and function of the microbiome. These models indicate that some microbial species are capable of metabolizing PFAS and their precursors and also, PFAS exposure can cause direct toxicity to some bacterial groups. 13
In human cohort studies, although most of the data are observational and do not fully prove a causal relationship, but there are strong evidences that show the correlation between higher level of PFAS in blood or urine and changes in the gut microbiome composition and also in metabolites profiles like SCFAs and bile acids.13,78 These results show the significant convergence between human and in vivo studies. Despite growing evidence, causal mechanisms of PFAS-microbiome interaction are poorly defined. Intestinal enzymes of biotransformation are mostly uncharacterized and microbiome composition role in inter-individual variability in PFAS kinetics is remain vague. Clarifying these pathways need integration of mechanistic models, longitudinal cohorts, and multi-omics. Figure 4 highlights how PFAS-induced changes in the gut microbiome may play a role in systemic toxicity. These effects occur through various interrelated pathways, including immune system disruption, metabolic imbalances, liver toxicity, kidney damage, and neuroinflammation. Mechanistic model of gut microbiome-mediated PFAS toxicity across multiple organ systems.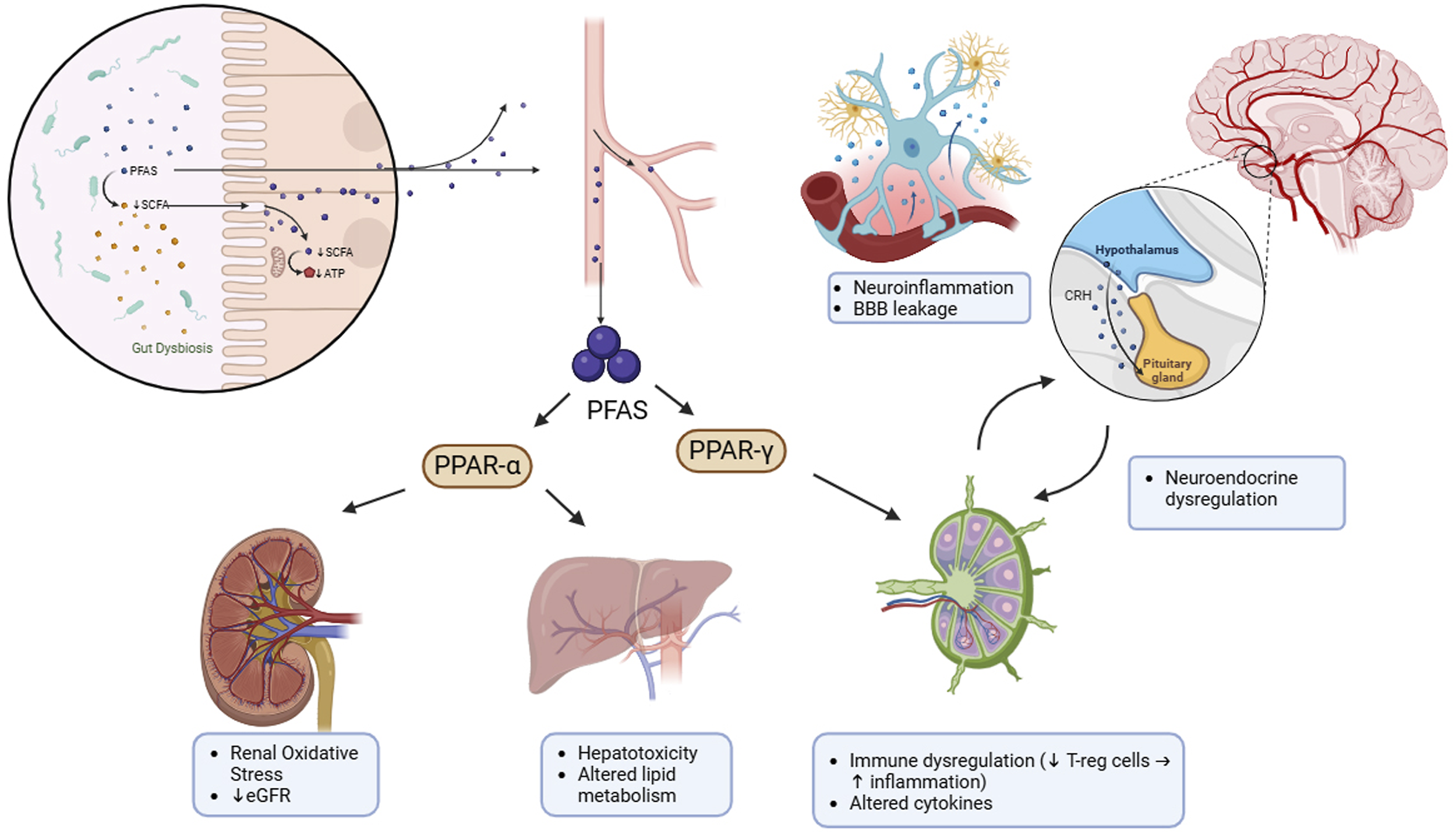
PFAS exposure leads to gut microbiome disruption and via SCFAs reduction exacerbates the dysbiosis. Increase in absorption of PFAS into bloodstream, activates PPAR-α and PPAR-γ signaling pathways that results in renal oxidative stress, eGFR reduction, hepatotoxicity, change in lipid metabolism, and also immune regulation disruption that characterized by regulatory T cells reduction and increase in inflammation. Circulatory cytokines and PFAS cause neuroinflammation, increase in blood brain barrier permeability, and also change in hypothalamus-pituitary axis neuroendocrine responses. Finally, these interactions after PFAS exposure can increase the risk of chronic diseases effectively.
Multi-omics approaches for investigating PFAS–microbiome interactions
Several epidemiologic studies show the relationship between PFAS exposure and adverse outcomes like metabolic disorders, immune and renal impairments. However, the exact mechanistic pathways, especially where the gut microbiome act as mediator or modulator, are still unclear. 78 Determining causal relationship between PFAS exposure, microbiome changes, and host’s implications need multi-omics data. Multi-omics (metagenomics, metabolomics, and transcriptomics) has the capacity to simultaneously record and connect changes in microbial composition, microbe/host metabolic function, and host’s gene response, and so it can suggest potential causal pathways. 79 Multi-omics approaches analyzing two key points in PFAS effects on the human health: first, determining the role of gut microbiome changes as a PFAS-effect mediator on specific clinical outcomes. Second, identifying metabolic and gene signaling pathways that mediate this relationship. 80 Integrative studies that merge 16S rRNA sequencing with multi-compartment metabolomics have shown that changes in gut microbial composition are strongly connected to systemic metabolic shifts across various tissues. These findings underscore the importance of multi-omics approaches in uncovering microbiome-driven mechanisms underlying diseases. 81
Recent review and experimental studies show that use metagenomics, metabolomics, and transcriptomics in combination is necessarily, metagenomics to identifying taxonomic changes and functional capacity of bacteria, metabolomic to clarifying functional pathways and metabolic changes, transcriptomics to track host’s cellular, immune, and metabolic responses. Combination of these data allows tracking key functional pathways such as change in SCFAs production, bile acids conversion, and lipid pathways and also can adapt with changes in immune and metabolic genes expression. 79 At the same time, available studies show that for Accurate causal inference or mediating, study should be trackable base on time changes and dynamics of responses. 82 However, in review articles no true longitudinal and interventional sampling is possible, but summarizing previous studies results show that recording key metadata such as age, sex, body mass index (BMI), diet, drug or antibiotic consumption, time and the method of sampling, and sample storage condition has an important role in interpretation of results. Because these agents can influence both PFAS level and microbe composition. 54 In addition, review articles suggest that conceptual controls such as reference samples, examination of microbe diversity, and correlation with host’s metabolome and gene data have an important role in understanding mechanisms and reducing errors caused by individual or technical differences. So, also in review articles, presenting a conceptual framework simplify understanding of multi-omics role in clarifying cause and effect relationships and also human health-microbiome-PFAS mediator pathways. 83
In the field of PFAS, shotgun metagenomics is necessary not only for describing microbe species diversity but also for recognizing genes and metabolic pathways related to absorption, accumulation, and transformation of PFAS. Main goal of this approach is providing taxonomic and functional data that can match with metabolome changes and gene response. 84 For example, a new study shows that 38 strains of gut bacteria have the ability to bioaccumulate PFAS from nanomolar to 500 micromolar, and this capacity was demonstrated in Bacteroides uniformis. Proteomics and metabolomics analysis in this study also shown significant changes in metabolic pathways after PFAS exposure. 54 Proposed technical progression for performing metagenomics includes DNA extraction with kits with high spectral coverage, library preparation with minimal PCR bias, removal of host reads, and sequencing depth appropriate for study’s goal (5–20 million paired-end reads for species describing and at least 20 million for MAG reconstruction). Suggested tools for taxonomic and functional profiling including Kraken2/Bracken or MetaPhlAn for taxonomy and HUMAnN3 for functional pathways. In addition, MEGAHIT or metaSPAdes assembly and binning for MAG reconstruction is necessary when the goal of study is recognizing species that are responsible for metabolite transformation and interaction with PFAS. 85
Metabolomics is a key functional part in PFAS studies that can show the relationship between microbe changes and host’s response. Especially, metabolite that have microbial origin like SCFAs, secondary amines, and microbially modified bile acids or lipid metabolites that involved in PFAS- related metabolic pathways. Several studies show that PFAS exposure causes significant changes in fecal, serum, and liver metabolome profile; such as SCFA reduction and lipid or phospholipid pathways disruption. 86 In addition, both untargeted and targeted metabolomics approaches can identify known and unknown metabolic pathways affected by PFAS and through direct matching PFAS plasma level and certain metabolites provides evidences for affected pathways. 87 Recent researches also shown that PFAS exposure can affect arachidonic pathways, serotonergic synaptic signaling, and tryptophan metabolic pathway (TRP) and lead to changes in behavioral outcomes. Although, some studies concentrate on metabolomic biomarkers such as glycerophospholipids that have been suggested as effective indicators of PFAS exposure. 88 For more detailed analysis, combination of Liquid Chromatography–Mass Spectrometry (HILIC for polar metabolites, and RP for lipids) with Nuclear Magnetic Resonance spectroscopy (NMR) for validation has been suggested. Internal standards, pooled QC and batch correcting strategies (QC-based LOESS or ComBat) should be applied to reduce technical artifacts. 79
Host’s transcriptomics in PFAS exposure can show how gene expression response to microbe and metabolites changes. Gene expression analyzing in gut mucosal tissue by using bulk RNA-seq or single-cell/spatial transcriptomics when needed, clarify that how microbe and metabolite changes affected immune pathways, epithelial barrier, and metabolic signaling. For samples with high microbial content, rRNA depletion usually preferred over polyA selection, and a sequencing depth of 30–50 million paired-end reads is sufficient for bulk RNA-seq. preprocessing (trimming and alignment), differential expression analysis (DESeq2/edgeR), and biological pathways analysis (GSEA/Reactome) are used to understand biological effects. 78 Recent studies have been provided direct evidences of gene expression changes caused by PFAS. For example, in silkworms, PFAS exposure results in significant changes in genes related to metabolism and gastrointestinal function, and correlation analysis shows that the gut microbiome act as a mediator in these effects. Also, a recent preprint study on bacteria shows that RNA-seq can reveal gene expression direct changes caused by PFAS at the genomic level. 89 Combining host’s transcriptomics data with metabolomics and microbiome allows researchers to clarify PFAS effects on immune pathways like NF-κB, inflammatory pathways, and hepatic metabolism paths. 90
Multi-omics integration in PFAS studies is accompanied with several biostatistics challenges such as data scale differences, missing values, batch effects, and imbalance between the number of features and samples. There are three approaches for combining data: first, early integration that enters all of the features in to the model simultaneously, second is intermediate integration which includes dimensionality reduction or choosing features before data combination, and third, late integration in which each omics part analyzing independently and then results have been integrated at the network or pathway level. Choosing the strategy is dependent on data structures. 82 For PFAS studies standardize analytical workflow is suggested, that has 3 stages: first, layer-specific preprocessing, second, feature filtering and dimensionality reduction, third, multivariate feature selection across omics layers. Layer-specific preprocessing is using CLR transformation for microbe data, log/z-score, or VST for metabolomics and transcriptomics datasets for homogenizing scales. Feature filtering and dimensionality reduction is removing low-variance and low-abundance features for statistically improve. Multivariate feature selection across omics layers is applying strategies like DIABLO for recognizing a collection of common features between omics layers that have the highest correlation with PFAS levels or health consequences.91,92
After recognizing key features multilevel correlation networks can be made, for example, using SparCC for the microbiome and spearman or partial correlation for linking microbes, metabolites, and host’s genes. Then, performing causal mediating analysis in longitudinal or interventional studies for examining metabolite or microbe pathways mediator role in PFAS effects. 78 In recent years, new statistic approaches have been recommended. For example, in one analysis, metabolites have been modeled in pathway-level groups to estimate pathway-mediated effects. This strategy also has been applied in a new study for examining prenatal PFAS exposure effects on gestational age using metabolomic datasets. In addition, metabolic flux and pathway modeling by using metabolomics data and microbe functional genes can clarify possible operative pathways for PFAS biotransformation or their effects on host’s metabolism. 82 This model-base integration especially when combined with genome-scale metabolic models (GEMs) allows prediction of active pathways in response to PFAS. 93 The multi-omics approach that combined metabolomics, metagenomics, and host’s transcriptomics is the best current solution for discovering microbial-mediated mechanisms underlying PFAS effects. Strict implementation of sampling protocols, rigorous quality control, and using advanced integration strategies like DIABLO and causal network formation in combination with mediation analysis, can provide reliable and interpretable results and facilitate development in preventive and treatment approaches. 80
Therapeutic modulation of the gut microbiome to counteract PFAS-induced toxicity
Gut microbiota have been shown to interact directly with PFAS with modulating their absorption and distribution. 94 Certain gut microbes can bind or change PFAS compounds, with decreasing their systemic bioavailability or, conversely, producing secondary metabolites with enhanced toxicity. Using engineered microbial communities, several PFAS compounds, including PFOA and PFNA, were found to undergo substantial microbial uptake or transformation. 54 This microbial processing can modulate systemic exposure to PFAS and change their toxicological effect on the host. Additionally, microbial metabolism may influence PFAS-induced oxidative stress, inflammation, and immune dysregulation mechanisms largely responsible for their toxic effects. 95 Individual bacterial strains showed different abilities for PFAS depletion which highlights the role of specific gut microbes in chemical metabolism. These interactions suggest that the composition and functional activity of the gut microbiome may directly affect PFAS bioavailability, endocrine disruption, and subsequent health outcomes. 94 Understanding these microbiota-mediated mechanisms is therefore critical for evaluating PFAS-related risks in humans. Restoring a healthy balance of gut microbiota has been linked to enhanced intestinal barrier function and better immune regulation. This highlights the potential of microbiome-focused therapies as a promising approach to address gastrointestinal and metabolic issues caused by PFAS exposure. 96
Human gut bacterial strains interact with PFAS with considerable variability, particularly PFNA, a long-chain compound primarily excreted via feces. Due to a study with 89 microbial strains which surrounded major gut phyla, probiotics, and kefir-derived microbes, distinct differences in PFNA sequestration was noticed. 54 Accumulation patterns were detected as strongly phylum-dependent, with Bacteroidota demonstrating the highest uptake, and a significant division was shown between low-accumulating Gram-positive strains and high-accumulating Gram-negative strains. Experiments with lipid-rich yeast indicate that, in addition to the higher lipid content of Gram-negative bacteria, other microbial traits contribute to enhanced PFNA accumulation. 97 These findings emphasize that PFAS maintenance can be modulated by gut microbial composition and metabolic diversity. Evidence from experimental studies suggests that modulating the gut microbiota may enhance the integrity of the intestinal mucosal barrier and support metabolic balance. This highlights the potential of microbiome-targeted approaches in addressing PFAS-related metabolic and gastrointestinal issues. Interventions aimed at reshaping gut microbial communities have demonstrated their ability to mitigate metabolic dysfunction and repair the integrity of the intestinal mucosal barrier. This highlights the promising therapeutic potential of microbiome modulation in addressing conditions linked to exposure to environmental toxicants.98,99 These factors eventually influence their bioavailability and toxicity in the host. Understanding such strain-specific interactions is crucial for assessing the affection of microbiome on PFAS toxicity. 100
Recent evidence highlights the critical role of gut microbiota in modulating the absorption, distribution, and excretion of PFAS compounds. Studies in germ-free or pseudo germ-free models prove that according to the impairment detected in gut barrier function, disruption of the gut microbiota may lead to increased PFAS. Certain PFAS, such as HFPO-TA, HFPO-DA, and PFOA, have shown to be resistant to microbial metabolism. But other environmental contaminants could possibly go through microbial changes which could eventually produce toxic metabolites in the colon. So in conclusion, the toxicological effects of PFAS depend on individual gut microbiome profile and functional potential as well as their inherent molecular stability. These findings emphasize that gut microbiota composition and activity are fundamental factors influencing PFAS bioavailability and toxicity. Therefore, considering the gut microbiome is important for properly assessing the risks of PFAS and similar chemicals. 101
Beyond persistence: How gut microbes influence the bioavailability and toxicokinetics of PFAS in humans
Toxicokinetics and bioavailability are significant factors in understanding why PFAS affect people differently. They decide how these compounds are absorbed, distributed, metabolized, and eliminated, and that determines their levels inside the body and their toxic effects. Chemical factors such as chain length and biological factors affect these mechanisms; but gut microbiota has been considered an interesting factor modulating PFAS transformation in a direct way. These microbes modify how they cross the intestinal barrier, or interact with hepato-intestinal cycle; this ultimately affects the extent to which these substances stay in the body. 13 The gut microbiome helps regulate human metabolism which influences how different compounds are processed. Certain microbial species, specially archaea because of its contact with PFAS, could negatively be associated with glycine-conjugated bile acids such as GDCA, GUDCA, and GUDCAS. 69 This indicates that gut microbes may modulate bile acid metabolism, which in turn could affect PFAS absorption and bioavailability. Additionally, due to evidence based on recent research, PFAS can disrupt the intestinal reabsorption of bile acids; which could possibly change their Toxicokinetics. Studying these interactions could result in better understanding of how the gut microbiome might affect PFAS metabolism in humans. 102
How methanogenic archaea affects the metabolism of bile acids, has not yet been fully elucidated. Evident contact with PFAS affected the composition and variety of gut microbes as predicted; which eventually affected the metabolism of bile acids, or the opposite. 69 According to strong connections between particular microbial changes and conjugated bile acids, archaea could be an important mediator in the interactions between bile acid metabolism, reuptake, and PFAS exposure. It is also important to note that even with longitudinal designs, modest sample sizes in previous studies could affect our understanding of how chemical exposures influence the gut microbiome in mother-infant settings. Still, the result suggests that PFAS exposure is likely a contributing factor in shaping gut microbial composition. 69
Recent studies have been focused on the quick uptake and retention of PFAS in gut bacteria, especially in Bacteroides uniformis, a common gut bacterium. 54 Due to those studies, PFNA uptake happened within minutes in growing and stationary-phase cultures, and after being taken up, the compound remained within bacterial cells for several days. The amount of PFAS retained seemed stable across a broad range of exposure concentrations, and uptake increased with chain length, reaching higher levels for longer-chain PFAS such as PFDA. Importantly, bacteria continued to grow normally even at micro molar concentrations much higher than those usually found in the environment. These observations indicate that gut microbes can retain PFAS efficiently, which possibly affects their bioavailability and persistence in humans. 103 Analysis of secreted metabolites has shown that contact with PFAS could regulate the profiles of numerous microbial metabolites, including carnosine, acetophenone, and creatinine, depending on the level of exposure. 104 PFAS influence the amount of certain types of gut microbes, with increases in opportunistic pathogens like Morganella and Enterobacterales, and also decreases in beneficial SCFA-producing genera such as Lachnoclostridium. 105 These microbial shifts match the changes in SCFA production, with higher PFAS concentrations that sometimes enhances SCFA levels, which could be an adaptive response. Collectively, these findings indicate that PFAS-induced microbial imbalance can disrupt critical metabolic pathways, which may influence PFAS bioavailability, host metabolism, and gut health. 106
Conclusion
PFAS, known for their persistence in the environment, are commonly encountered through sources such as contaminated food, drinking water, food packaging, and various consumer products. Emerging research highlights the significant role of the gut microbiome in shaping PFAS bioavailability, toxicokinetics, and biological impacts. Exposure to PFAS has been shown to disrupt the composition and metabolic activity of gut microbes, interfere with bile acid and short-chain fatty acid pathways, weaken intestinal barrier function, and contribute to immune system imbalances and metabolic disorders. Evidence suggests that microbiome-related mechanisms may influence an individual’s vulnerability to health effects associated with PFAS, including metabolic, liver-related, and inflammatory conditions. Additionally, certain microbial groups appear capable of sequestering or transforming PFAS, emphasizing the dual role of the microbiome as both a target and a potential modulator of PFAS toxicity. Advances in multi-omics technologies are enhancing our understanding of these intricate interactions and paving the way for microbiome-centered strategies to mitigate the health risks posed by PFAS exposure. However, further detailed mechanistic and longitudinal research is needed to establish clear causal links and aid in developing robust risk assessment and intervention measures.
Footnotes
Acknowledgements
We would like to thank the Clinical Research Development Unit of Kowsar Educational and Research and Therapeutic Center of Semnan University of Medical Sciences for providing facilities to this work.
Consent to participate
This article is a review and does not involve human participants or animal experiments.
Author contributions
All authors contributed to the study’s conception and design. Material preparation, data collection and analysis, S.T., and N.A.; the first draft of the manuscript, M.E., S.A., and S.T.; Writing—original draft preparation, S.T., N.A., S. S. and S.A.; writing—review and editing, M.E.; Software, S.T.; visualization and supervision, M.E.; project administration, M.E., All authors have read and agreed to the published version of the manuscript.
Funding
The authors received no financial support for the research, authorship, and/or publication of this article.
Declaration of conflicting interests
The authors declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Data Availability Statement
All data discussed are derived from previously published studies, as cited in the manuscript.