
Abstract
Magnesium and its alloys have a great potential to be used as degradable biomaterials for clinical applications, which only need temporary support. However, the corrosion of Mg under physiological conditions is not well understood yet. The aim of this study was to determine the effect of proteins and complex corrosion environments. The corrosion rates of dysprosium containing magnesium alloys in technical and physiological corrosion set-ups were compared. Moreover, pure magnesium was immersed under cell culture conditions in simple and complex protein solutions. Corrosion rates showed huge variations depending on the corrosion environment. Proteins delayed corrosion and altered the ion composition of the solutions. In conclusion, the choice of appropriate corrosion environment is crucial for in vitro experiments.
Introduction
It has been known since the last century that Mg alloys are favourable in medical applications due to their low elastic modulus, appropriate strength compared to bone, excellent biodegradability and bioresorbability.1,2 The most advanced clinical applications are biodegradable cardiovascular magnesium stents. They were successfully tested in animals,3 and several clinical human trials have been conducted.4,5 Magnesium alloys are also investigated for orthopaedic applications due to their mechanical properties close to bone and the positive effect on bone remodelling.6 Even though Mg and its alloys have been investigated as implants for almost two centuries, commercial implants of Mg and its alloys are still not available.7 One of the key issues is that the corrosion rate of Mg and its alloys is too high or even unpredictable due to their origin from structural applications.1
In response to this problem, many works have been carried out to investigate the corrosion behaviour of Mg. Various simulated physiological fluids such as 0·9wt-%NaCl solution, simulated body fluids, phosphate buffered solution, modified eagle medium, Dulbecco's modified Eagle medium (DMEM), etc. are used.8 – 11 As a result, it is difficult to compare the degradation rates of magnesium alloys determined from different publications. The use of different test solutions and different surface area/electrolyte volume ratios both greatly disturb the measurement.9,12 Moreover, it is reported that the corrosion rate in vivo is much different from that in vitro due to the dynamic body environment and complicated composition of body fluid.13,14 Therefore, to obtain comparable corrosion results between in vitro and in vivo, a new standard test protocol has to be developed. This makes it very essential to investigate the influence of different factors in vitro, such as proteins and corrosion environment, on the corrosion behaviour of Mg. Therefore, the aim of our study was to determine the influence of proteins in different corrosion environments and solutions on the corrosion of Mg.
Experimental
Materials preparation
Metallic Mg (99·94%) was melted in a mild steel crucible under a protective atmosphere (Ar+2%SF6). Pure Dy (99·9%) was added at a melt temperature of 720°C, after which the melt was stirred for 30 min at 200 rev min−1. Permanent mould direct chill casting was used to prepare the ingot.15 The size of the ingot is 20×12×6 cm. The same procedure was used to prepare pure Mg ingot.
Round samples with a diameter of 10 mm and a height of 1·5 mm were prepared by electrical discharge machining. To remove remnants from this process, all the samples were cleaned ultrasonically in ethanol and distilled water.
Immersion test
Specimens for the immersion test were prepared by grinding each side with 2400 grid emery paper and degreasing the surfaces with ethanol before corrosion tests. Weight loss tests were performed under three different conditions: (1) in 0·9 wt-%NaCl solution in atmospheric conditions and at room temperature (RT) as an example for technical corrosion set-ups; (2) in cell culture medium (CCM) consisting of DMEM (Sigma Aldrich Chemie, Taufkirchen, Germany) and 10% fetal bovine serum (FBS Gold; PAA Laboratories, Linz, Austria) under atmospheric conditions (21%O2, 0·043%CO2) at 37°C; and (3) in CCM under cell culture conditions (37°C, 21%O2, 5%CO2, 95% relative humidity). The composition of DMEM solution can be found elsewhere.16 For the corrosion tests in CCM, the samples were sterilised in 70% ethanol for 15 min before the corrosion test, and then all procedures were carried out under sterile conditions. The samples were immersed in NaCl solution for 3 days and immersed in CCM for 14 days. Corrosion products were removed using chromic acid (180 g L−1) by immersion of samples for 20 min at RT. The average corrosion rate was calculated in millimetre per year (mm/year) using the following equation
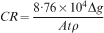
Determination of influence of corrosion solution and proteins
To investigate the influence of protein addition on the corrosion of Mg, two experiments were performed. First, FBS diluted in water was used as corrosion medium. After different time points (0·5, 1, 3, 6, 12, 72, 120 and 196 h), 100 μL of the corrosion solution was sampled for osmolality and pH measurements. The loss of corrosion solution was compensated by adding an equal amount of fresh solution. In a second experiment, FBS was diluted in DMEM as buffered solution. Experiments were terminated after 24 and 72 h. All the solutions are listed in Table 1. The samples were immersed into 3 mL of the respective solutions in a 12 well plate and kept under cell culture conditions. The osmolality and pH of the extracted corrosion medium were measured at different time points using an osmometer (Osmomat 030, Gonotec, Berlin, Germany) and a pH meter (Titan X, Fisher Scientific GmbH, Schwerte, Germany). Owing to the more complex formulation of DMEM, also the amounts of calcium and magnesium in solution were determined using ion selective electrodes (Hanna Instruments, Smithfield, USA). The measured values (mV) were used to determine the ion concentration in solution by plotting against a standard curve. Values were expressed as per cent of control of the initial solutions. The amount of protein in the solution was determined by bicinchoninic acid assay (BCA, Thermo Fisher Scientific, Rockford, USA), according to the manufacturer instructions. In brief, 25 μL of standards and each solution were pipetted into microplate wells. The working reagent (200 μL) was added and mixed for 30 s. After an incubation of the plates for 30 min at 37°C, the absorbance was measured at 562 nm on an ELISA plate reader (Tecan Sunrise, Tecan Deutschland GmbH, Crailsheim, Germany). Concentrations of protein in the samples were determined by plotting against a standard curve (bovine serum albumin).
Solutions used for immersion test: symbols are used as abbreviations for results description

Results
Corrosion rate in different corrosion conditions
Figure 1 shows the corrosion rate of as cast Mg–Dy alloys in 0·9 wt-%NaCl (RT), CCM (cell culture condition) and CCM (37°C). It is clear that the corrosion rate shows a huge variation in different corrosion conditions for all the tested alloys. For example, the corrosion rate in NaCl is ∼7·5 mm/year for Mg–5Dy alloy, while it is reduced to 0·9 and 0·1 mm/year in CCM under cell culture conditions and CCM at 37°C respectively. As a general trend, the immersion in CCM greatly reduces the corrosion rate under atmospheric conditions at 37°C. However, when cell culture conditions are applied, the corrosion rate increases. Furthermore, the trend of corrosion rate is also different in different corrosion conditions. This indicates that both composition of corrosion solution and corrosion environment have significant influence on the corrosion of Mg.
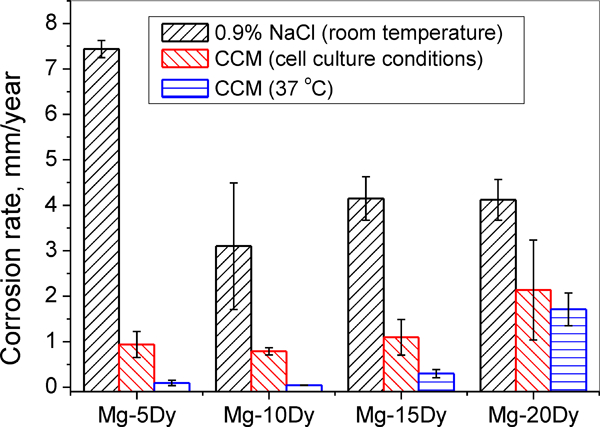
Corrosion rate of as cast Mg–Dy alloys in different corrosion solutions or corrosion environments
Effect of proteins in water
To determine the influence of proteins alone, they were diluted in water. Figure 2a shows the change in osmolality of the respective corrosion solutions as a function of immersion time. In water, the osmolality increases with the increase in immersion time up to 12 h, and then it reduces gradually. A similar trend is also observed in F10 and F50 solution, whereas the F100 solution still increases after 8 days of immersion. Figure 2b shows the increase in osmolality as a function of time. In pure water (F0), the osmolality is stable for 6 h; thereafter, a rapid increase takes place up to 12 h. This is followed by a decrease. The addition of proteins leads to a delay of osmolality increase and to an increase of the peak value. In F10, the peak value was reached after 72 h (0·175) and in F50 after 120 h (0·25). In F100, the peak value was not even reached after 192 h immersion.
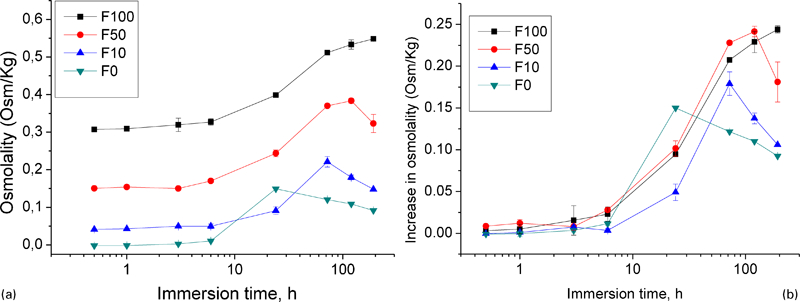
Increase of osmolality in corrosion medium with different amounts of proteins after immersion for different time periods
The according pH values are depicted in Fig. 3. Between 6 and 12 h, the pH for most corrosion media reaches a peak value, except for F100. This peak value reduces drastically with the increase in FBS content, which can be attributed to the buffering effect of FBS under cell culture conditions. The peak value for F0 solution of pH is 9·1, while in F100 solution, only pH 8·2 is reached. After 192 h immersion, the pH of all the corrosion mediums tends to be in a narrow range (8·2-8·4) due to the compromise between corrosion and buffering effect.
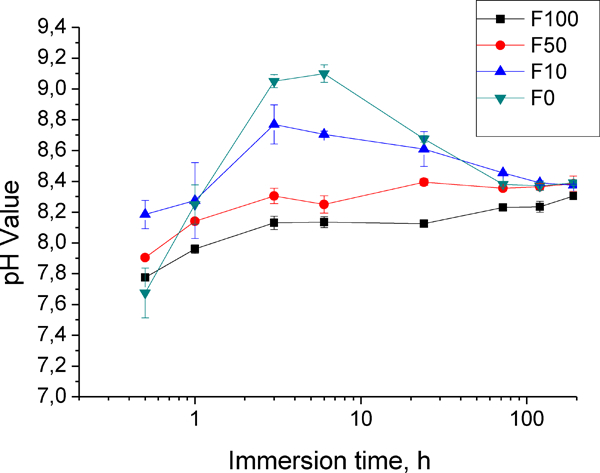
pH change of corrosion medium as function of immersion time
Effects of proteins in buffered solutions
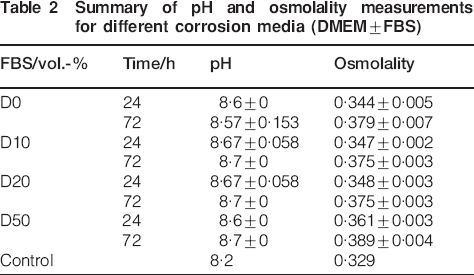
Instead of using water, in this experiment, a buffered solution (standard for cell cultivation) was used to examine whether the type of solution also plays a role. The pH value for all solutions was in the same range, whereas the increase in osmolality was more pronounced than for water diluted protein (Table 2). Between 24 and 72 h, the increase was most pronounced for D0 (0·035), and the solutions with protein exhibited a lower increase and were all in the same range (0·027-0·028).
Summary of pH and osmolality measurements for different corrosion media (DMEM±FBS)
As the osmolality only gives information about the total content of salts in solution, more detailed measurements of calcium and magnesium were performed. It could be observed that after 24 h, the amount of calcium as well as magnesium in comparison to the control was decreased with increasing protein content (Fig. 4). After 72 h, this trend was not observable anymore. Magnesium ions in solution were comparable to control (between 102 and 108%), whereas the amount of solved calcium was drastically reduced (between 57 and 65%). It was also observable that the amount of protein was lower than observed after 24 h (D50: 5250 μg mL−1 after 24 h and 4280 μg mL−1 after 72 h).
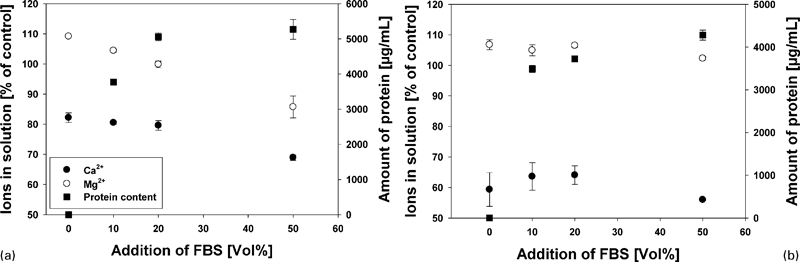
Ion content (circles) and amount of protein (squares) in solution after a 24 h and b 72 h
Discussion
The aim of this study was to determine the influence of proteins and corrosion conditions on the corrosion of pure magnesium and a dysprosium based alloy series. It was performed to determine factors that could be relevant to develop an in vitro system, which is more closely related to the results obtained in vivo. Although no direct comparison with data obtained in vivo was performed, some general trends could be observed: (1) the difference among the technical set-up (0·9% saline at RT), the use of proteins and/or buffered solutions under atmospheric conditions and the use of cell culture conditions all lead to different results; (2) the presence of proteins leads to a different corrosion behaviour of the analysed materials; and (3) the application of a physiological environment is relevant to determine further factors, which could be important to describe the corrosion mechanism.
In detail, for the first point, it is obvious that the results obtained from the technical set-up are contradictory to CCM under atmospheric conditions (ranking by corrosion rate 10Dy<15Dy = 20Dy<5Dy with 0·9% saline; 5Dy = 10Dy<15Dy<20Dy with CCM). By the application of cell culture conditions, the trend obtained using CCM is generally followed but allows some further discrimination (ranking: 10Dy<5Dy<15Dy<20Dy). The more intriguing fact is that there are orders of magnitude between the calculated corrosion rates (5Dy: 7·5 mm/year in NaCl compared to 0·1 mm/year for CCM). This discrepancy could be due to the presence of proteins, which are part of the buffering system in blood.17 The buffering capacity of proteins is due to hydrogen uptake (acidic conditions) or release (basic conditions) by the carboxyl and amino groups of their amino acids. Under basic conditions, the carboxyl group does not play a role (as it is COO−), whereas the amino group is releasing a hydrogen atom (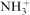 to NH2). The used FBS as a source for proteins was analysed by the manufacturer and contained 42 g mL−1 of protein (30 g albumin, 8 g α-Globulin, 4 g β-Globulin and 5·3 mg IgG). Moreover, it is known that temperature and carbon dioxide are influencing the properties of proteins.18,19 Hence, in further experiments, cell culture conditions were applied to mimic the blood environment.
to NH2). The used FBS as a source for proteins was analysed by the manufacturer and contained 42 g mL−1 of protein (30 g albumin, 8 g α-Globulin, 4 g β-Globulin and 5·3 mg IgG). Moreover, it is known that temperature and carbon dioxide are influencing the properties of proteins.18,19 Hence, in further experiments, cell culture conditions were applied to mimic the blood environment.
In the first experiment, only the osmolality as a measure for the totally dissolved ions in solutions and the pH value was measured. It could be shown that the addition of proteins induced a buffering effect under cell culture conditions, which was correlated to the amount of protein present (Fig. 3). An increase in the pH value normally leads to the formation of a protective corrosion layer under atmospheric conditions.20 However, in the used physiological environment, this seems not to be the case, as the main ion release for pure water starts after 6 h (Fig. 2), while the pH is decreasing during that time (Fig. 3). The addition of proteins led to an increase in osmolality, which means that more ions are dissolved from the sample. Therefore, under cell culture conditions, the addition of proteins leads to an increase in the corrosion rate. This is in contrast to other studies, where the addition of proteins leads to a decrease in corrosion rate.21,22 In these studies, HEPES was used as buffering agent, which was recently described to also have an influence on the corrosion rate.23
A general drawback of studies under static conditions is the rise of pH above physiological conditions (Table 2 and Fig. 3). This remains the main obstacle for predicting in vivo results from in vitro data.24 However, this study revealed the very interesting fact that the application of cell culture conditions (mainly characterised by the introduction of a continuous supply of carbon dioxide) seems to be one of the key points towards more predictable results. This can be deduced from the results obtained by using pure water (Figs. 2 and 3, F0). As expected, pH is rising during the initial corrosion (up to 6 h, peak value 9·2). This would induce the formation of a corrosion protective layer of magnesium hydroxide and hence to a low corrosion rate, which is also indicated by the low increase in osmolality (Fig. 2). However, between 6 and 12 h, a sudden increase in osmolality is observable, accompanied by pH decrease. This indicates that even deionised water acquires buffering capacity in this environment due to the release of magnesium ions from the samples in combination with the presence of CO2. This is due to the higher solubility of magnesium carbonate (220 mg L−1) compared to magnesium hydroxide (12 mg L−1), leading to a (super)saturation of the solvent.16 Under atmospheric conditions, this effect is not observable, and pH can rise up to 10 (unpublished results).
To shed further light on the corrosion mechanisms, we performed a further experiment in complex buffered solutions. Here, we choose only two time points because the previous experiment showed that in the presence of proteins, the main ion release occurred between 24 and 72 h. Here, it could be shown that the measurement of osmolality and pH did not elucidate differences (Table 2). Therefore, we determined the amounts of magnesium, calcium and protein in the solution. The measurement at 24 hours indicated that the presence of proteins reduced the solved ions in solution, whereas at 72 hours no such effect of the proteins could be determined. This is probably due to the binding of both ions (Mg2+ and Ca2+) to proteins, especially serum albumin.25 After 72 h, it is supposed that the ions are in equilibrium state, so no effect of protein is observable anymore. However, the depletion of calcium (which is derived from CCM) to ∼60% indicates the formation of calcium containing corrosion products. This was also observed in other studies.16,24
Although the amount of proteins in solution was reduced, still ∼80% of the initial protein could be measured. It is generally believed that proteins interact and alter the corrosion behaviour of metals via adsorption, chelation and formation of a protective layer.26,27 The adsorption of proteins on surfaces is a complex process that involves van der Waals, hydrophobic and electrostatic interactions and hydrogen bonding.28 On surfaces, for example, albumin molecules will adhere in a flattened triangular shape with a length of 13 nm and 1·2 nm average thickness.29 As a result, these proteins would accelerate the dissolution rate of metals through their chelating effects.30 On the other hand, a protein containing layer would be formed, inhibiting the contact between electrolyte and metal surface, thus reducing the further corrosion.26,27 However, previous studies indicate that this mechanism may not occur when degradable metals are used.16 Protein detection by bovine serum albumin (Fig. 4) indicated a non-linear relationship. The amount of protein in solution was similar between 24 and 72 h with the substitution of 10%FBS, whereas at higher concentration of FBS, the content decreased. Thus, at physiologically relevant conditions, the proteins stay in solution and seem not to be incorporated into the corrosion layer. In contrast, at higher concentrations, a depletion of proteins from solution was observable. This could be an indication of protein incorporation into the corrosion layer. However, as this phenomenon was not extensively studied, it will be the aim to analyse the role of proteins in further research.
In conclusion, this study shows that the choice of corrosion environment is crucial for the experimental results. From the technical point of view, a lot of corrosion mechanisms of magnesium have been identified. However, physiological corrosion is still an obstacle and lacking a systematic approach requiring a multidisciplinary investigation. As a first approach, the application of cell culture conditions seems to be appropriate towards a more predictive in vitro environment. Under these conditions, many buffering systems are present, which alter the corrosion rate, corrosion products and maybe even corrosion mechanisms of magnesium alloys. Whether this is just due to pH effects or if proteins are an active component should be elucidated in further studies.
Footnotes
Acknowledgements
The authors would like to acknowledge G. Salamon and D. Lange for expert technical assistance. Financial supports from CSC-Helmholtz scholarship for the PhD study of L. Yangs in HZG are gratefully acknowledged.