
Abstract
Engineering biomaterials for regenerative medicine involves a myriad of aspects to be considered for the successful design, interaction with cells and integration with living tissues (i.e. pore size, mechanical properties, degradation rate, biological activity). Among different technologies used to functionalise synthetic biomaterials and promote cell adhesion, cell growth and cell differentiation, this review focuses on strategies to organise extracellular matrix (ECM) proteins in a biomimetic way, as cells do in natural tissues in vivo (the ECM is a mesh of proteins that surrounds cells, and therefore, constitutes the scaffolding of a tissue), but using functional materials instead of living cells. The authors critically review material-based strategies to organise fibronectin (FN), a core component in the ECM of many tissues, and engineer microenvironments that recapitulate the structure and properties of the ECM. Material-driven organisation of FN in analogy with their natural cell-mediated assembly is a powerful route to engineer the network structure and biological activity of FN fibrillar matrices, seeking to develop biomimetic scaffolds for regenerative medicine. Here, the authors discuss different routes to promote the cell-free formation of FN fibrils as well as the biological impact of these engineered cellular microenvironments.
Introduction
Regenerative medicine aims at promoting the healing of tissues and organs that are damaged as a consequence of a broad variety of diseases. The list includes musculoskeletal degeneration (e.g. osteoarthritis), cardiovascular problems (e.g. myocardial infarct) and neurological disorders (e.g. Parkinson’s). 1–3 Adult stem cells found in different locations within the human body, such as the bone marrow and adipose tissues, are at the centre of these strategies as they can be isolated with relatively simple procedures. Stem cells have the potential to differentiate into a variety of mature cells and generate a new tissue if the appropriate conditions are provided. In vivo, stem cell populations reside in ‘niches’, specific instructive microenvironments that regulate their function and participation in tissue generation, maintenance and repair. 4 On the other hand, the development of technologies to control stem cell differentiation in vitro, encapsulate and transplant stem cells and promote their recruitment from surrounding tissues in vivo is a key for the successful implementation of these novel therapeutical approaches. Engineered biomaterials have indeed emerged as a powerful tool to control stem cell behaviour and repair damaged structures. 4–8
Tissues consist of cells and a surrounding milieu that contains mainly proteins, glycosaminoglycans and water – the extracellular matrix (ECM). Regenerating a tissue involves providing (stem) cells with the adequate stimuli to differentiate, synthesise and secrete these ECM proteins. The main aim of biomaterials in regenerative medicine is to play the role of a synthetic extracellular matrix to support cell growth, differentiation and production of a functional and natural ECM. This particular role of materials in regenerative medicine means that demanding design criteria have to be considered in the engineering of the cellular microenvironment with biomaterials: (i) biomaterials must consist of 3D porous structures with adequate interconnection to allow cells to colonise the whole system. 9–12 This is achieved normally using interconnected scaffolds fabricated using different technologies (e.g. solid-free fabrication techniques, solvent casting/particulate leaching, polymerisation in the presence of templates, etc.), but also using hydrogels that contain degradable fragments upon secretion of cellular products [normally proteins secreted by cells that have the ability to degrade the ECM, such as matrix metalloproteinases (MMPs)]. 13–15 (ii) Also, and importantly for regenerative medicine applications, biomaterials are not permanent structures that will remain in the body forever, but biodegradable materials are used so that the degradation rate matches the rate at which the new tissue is formed. This is particularly important in load-bearing tissues (e.g. bone), so that the mechanical stability of the construct (synthetic biomaterial and newly formed tissue) is guaranteed during the healing process. 16–18 (iii) Finally, synthetic biomaterials consist of inert structures that have to be functionalised with adhesive proteins and other active biomolecules (e.g. growth factors) to enhance the bioactivity of the system. By doing so, the biomaterial becomes biologically active and it is recognised by (stem) cells (see Cell/material interactions section), allowing cell adhesion and differentiation, to promote tissue regeneration. 19–22
Among the variety of strategies available, the challenge to create new synthetic materials able to serve as biologically active materials that support key aspects of tissue regeneration, including the re-establishment of a functional ECM, has driven multidisciplinary efforts for the identification of cell-free routes to engineer biomimetic structures that resemble the properties of the natural ECM. 23 As mentioned, cells within tissues are in fact surrounded by fibrillar matrices that support and regulate interactions, direct cell fate and determine tissue repair. Fibronectin (FN) is a core component of the ECM, whose organisation into fibrillar networks is driven by cells through an integrin-mediated process. 24 During tissue repair, FN promotes cell processes critical to tissue regeneration and regulates the deposition and organisation of other ECM proteins. 25 Within this perspective, the development of artificial biologically active FN networks through cell-free routes gains a distinct bioengineering interest, because it is a way to enhance the biological activity of materials. Indeed, the use of bioinspired novel adhesive coating materials that stimulate cells to produce new, native ECMs would in turn promote cell functions crucial for tissue regeneration and homoeostasis.
The first part of this review describes the fundamentals of the interaction of cells with synthetic biomaterials, a process that must be taken into account for the design of biologically active structures. Then, the authors introduce the organisation of FN in the natural ECM, a biological process that occurs during the formation of new tissues. Afterwards, the authors critically review methods that have been proposed up to now seeking to reproduce FN fibrils in the absence of cells. These methods range from the use of denaturant agents to the use of mechanical force to stretch and assemble the molecules into fibrils, and include the organisation of FN into (nano)networks upon adsorption onto specific surface chemistries. The biological activity of these artificially organised FN matrices is presented in the last section of this paper and includes cell adhesion, ECM organisation and degradation, and higher order cellular functions, such as stem cell differentiation on engineered artificial FN matrices.
Cell/material interaction
Living cells cannot interact directly with foreign materials, but it is now well established that cell adhesion on biomaterials is mediated by a layer of proteins previously adsorbed on the material surface, coming from either the physiological fluids in vivo or intentionally deposited in vitro, e.g. from (competitive) adsorption of serum proteins or even after chemical attachment to the substrate. 26–29 These proteins can be FN, vitronectin (VN), or fibrinogen, representing the so-called soluble matrix proteins in the biological fluids. 28 Upon longer contact with tissues, many other ECM proteins, such as collagens and laminins, will also associate with the surfaces, affecting the cellular interaction. The concentration, distribution, conformation and mobility of the adsorbed protein layer on a surface play a fundamental role in the biofunctionality of a synthetic material, affecting the type and extent of the cell response, and are clue factors to understand the biological response of a substrate. 29,30
Proteins are hence considered to be key factors in mediating the cell/material interactions, and their status (the amount, conformation and strength of interaction) on a material surface determines the biocompatibility of the system. The amount of proteins adsorbed and their conformation depend on the chemical groups of the substrate that they adsorb to, which determine the energetic and entropic interactions. 31 In addition to time-dependent compositional changes, these proteins that finally remain on the surface may undergo conformational and orientational rearrangements to increase the contact area with the substrate and minimise the interaction energy. Moreover, as protein concentration on the surface increases, they may go through ordering transitions. 29,32,33 Different model materials have been prepared to learn more about cell/material interactions, especially in what cell adhesion is concerned. These works are mainly focused on the effect of material properties on the biological performance of the substrate, and only a few investigate this effect by addressing first protein adsorption and conformation on the material surface to correlate this phenomenon with cell behaviour. Upon adsorption, proteins might in fact expose different domains, eliciting specific interactions with cell receptors. Despite the belief that cell/protein/material interactions are critical to the engineering of new biomaterials, clear links between the material properties, the adsorbed protein layer and their influence on cells remain far from being understood; in particular, the behaviour of surface-associated matrix proteins is generally missing.
Even if cells do not interact directly with the surface of synthetic materials, this interaction is mediated by ECM proteins previously adsorbed on the substrate surface; it is said that cells respond to three different kinds of surface parameters: chemical, topographical and mechanical. For example, the influence of surface chemistry on protein adsorption and cell adhesion has been addressed on surfaces with well-controlled chemistry, in order to investigate the role of well-defined chemical groups, e.g. using self-assembly monolayers (SAM). 34–38 Understanding cell/protein/material interactions is fundamental to develop more powerful material-based tools in tissue engineering and regenerative medicine strategies. The design of material substrates with defined physical and chemical properties (chemistry, topography, stiffness, gradients) in three-dimensional environments must lead in the near future to learn more about the specific roles of protein adsorption and the effect of material properties on cell adhesion, cell differentiation, matrix reorganisation, deposition and degradation at the cell/material interface. 39–41
On the other hand, in nature, ECM proteins have a pivotal role in cell adhesion, morphology, and migration; these events underlie the subsequent responses required for tissue repair, with the nature of cell–surface interactions contributing to survival, growth, and differentiation. The interactions of adsorbed proteins with cells are mediated via integrins. These are cell membrane proteins that recognise and bind a variety of cell–surface-associated and ECM-associated proteins. Integrins are heterodimers composed of α and β subunits that occur in distinct combinations and bind specific ligands. 42 In vertebrates, at least 24 integrin heterodimers have been found, composed of 18 types of α subunits and eight types of β subunits in various combinations. Integrins bind cell surface and ECM components, such as FN, VN, collagen, and laminin. 42 The most common integrins found are α5β1 (the classical FN receptor) and α ν β3 (VN receptor). One of the best-known integrin ligands is the RGD peptidic sequence. All five ν integrins, two α integrins (5, 8) and αIIβ3 share the ability to recognise ligands containing an RGD tripeptide active site. 42 This site is located within the type III10 repeat of FN and VN. Another important site of FN is the synergy sequence PHSRN, within the type III9 repeat. This sequence promotes specific α5β1 integrin binding to FN, given that it is the unique amino acidic sequence inside the FN that integrin α5β1 specifically recognises and binds, and raises α5β1 to the status of the main FN receptor. 24,32,42–44
The attachment of the cell to an ECM protein takes place through formation of cell adhesion complexes, so-called focal adhesions. Focal adhesions consist of large supramolecular complexes that provide intracellular binding sites to permit the formation of signalling complexes on the cytoplasmic side of the cell membrane. Integrins bind to the cytoskeleton via adaptor proteins, such as talin, α-actinin, filamin and vinculin. Other focal adhesion proteins are involved in the transmission of signals received from the ECM by integrins and directed to the cell nucleus, such as focal adhesion kinases (FAK). These signalling molecules bind to and associate with integrin-adaptor protein-cytoskeleton complex. 42 After focal adhesion formation, mechanical force and regulatory signals are transmitted that trigger gene and protein expression changes and consequently determine cell fate (including cell growth and cell differentiation) (Fig. 1). This is a sequential process that consists of integrin binding to the ECM, clustering into focal adhesions, and starting of signalling cascades that determine cell fate as a consequence of the extrinsic signals coming from the ECM. Hence, engineering synthetic ECMs (biomaterials) offers a powerful tool to direct integrin binding and eventually cell fate.
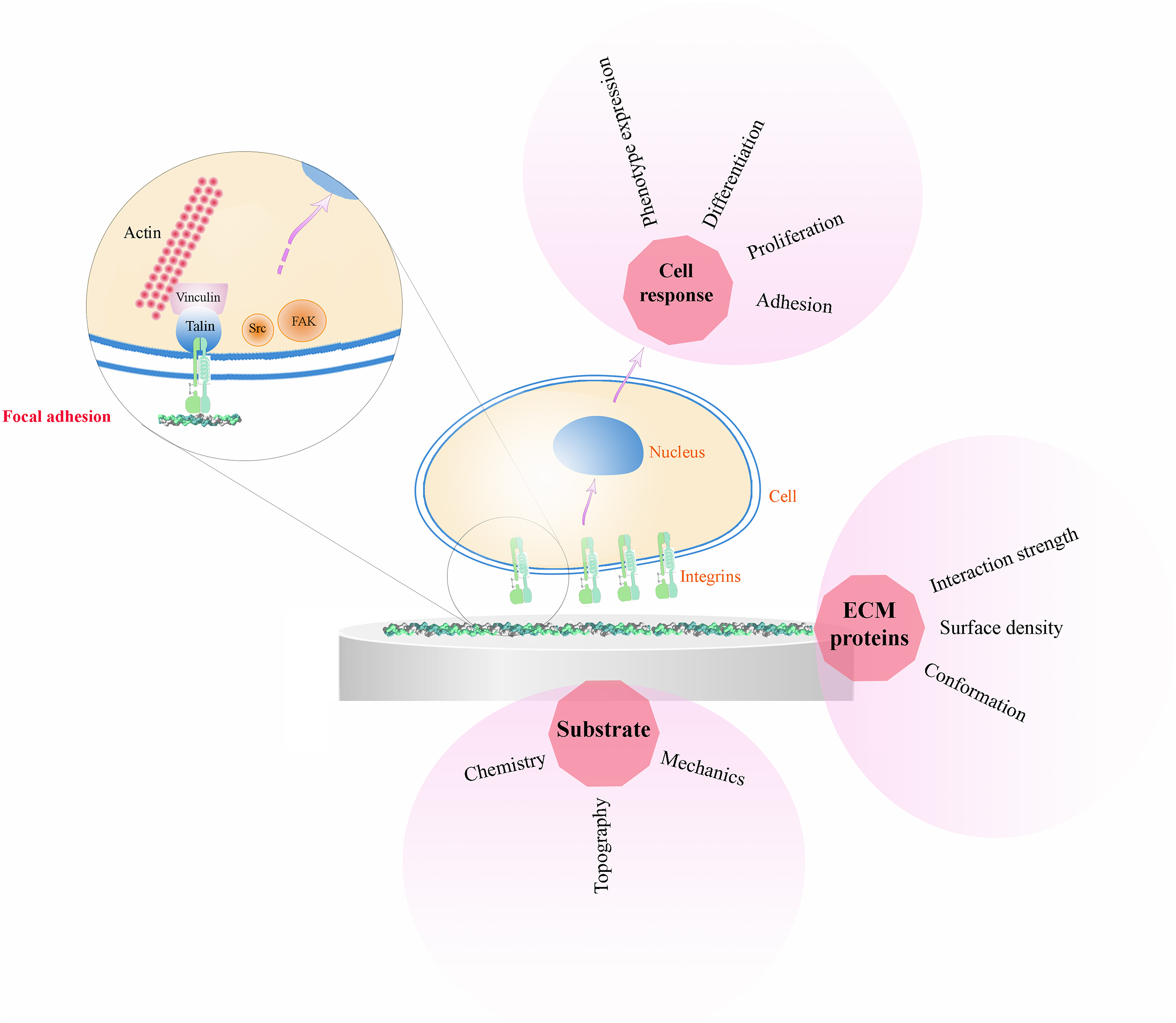
Cell/protein/material interactions. The interaction of biomaterials with living cells occurs through an intermediate layer of ECM proteins. Different properties of the material (chemistry, topography, mechanical properties) influence cell response in dependence of the conformation, surface density and strength of interaction between the material surface and the adsorbed ECM proteins. Cell adhesion to these proteins occurs via integrins that cluster into focal adhesions and link the ECM to the actin cytoskeleton (only one integrin is sketched in the focal adhesion cluster for simplicity of representation)
Natural fibronectin matrices
Fibronectin is a core component of the ECM, synthesised by adherent cells that then assemble it into a fibrillar network in an integrin binding-dependent mechanism. 43 The assembly of FN matrix is the initial step that orchestrates the assembly of other ECM proteins and promotes cell adhesion, migration and signalling. 45 Indeed, FN fibrils possess binding sites for multiple ECM components needed for the assembly of several other ECM proteins. Moreover, they provide support for cell adhesion receptors (most notably integrins) that transduce signals triggering cell fate. 46,47 Therefore, FN matrix has an important role for normal cell adhesion and growth, and plays a critical role in early development. 48
Fibronectin structure and essential domains for FN assembly
Fibronectin is a multidomain protein whose FN monomer ranges in size from 230 to 270 kDa and binds into dimers via two disulphide bonds at the C-terminus of the protein (Fig. 2). Usually protein domains can be incorporated as modules into different proteins. The modular approach to protein architecture is particularly easy to recognise in large proteins, which tend to be mosaics of different domains and thus can perform different functions simultaneously. 42 Fibronectin contains three types of repeating modules: types I, II and III (Fig. 2). The type I and II units contain two intramolecular disulphide bonds to stabilise the folded structure, while type III units lack this kind of bridges. All types of modules are structured in β-sheets, which, in the case of types I and III, are enclosing a hydrophobic core that contains highly conserved aromatic amino acids (which include an aromatic ring, e.g. phenylalanine or tryptophan). 45,49 The most common mechanism of FN production generates the two major different forms of FN: plasma (pFN) and cellular FN (cFN). pFN is produced by hepatocytes in the liver and secreted into the blood, where it remains in a soluble form until the activation of the integrins of haematopoietic cells (blood system cells) by a platelet-mediated mechanism allows pFN to bind and assemble into fibrils. 49,50 On the other hand, cFN is secreted by cells as a dimer in a compact globular structure and then is assembled into fibrils (insoluble form) in a cell-dependent process.
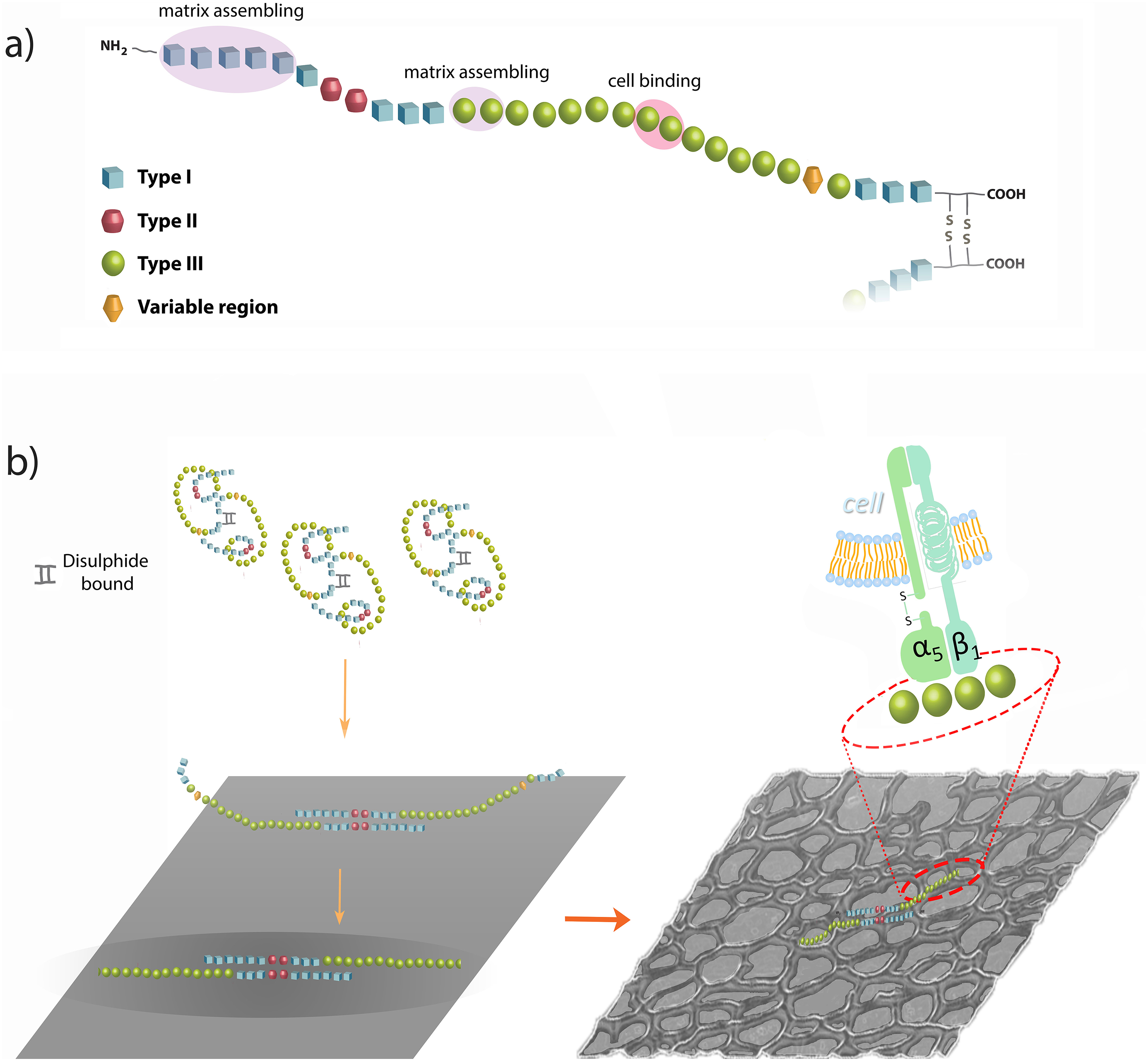
Fibronectin (FN) and fibrillogenesis. a Molecular structure of a monomer of FN, which consists of three different types of modules [type I, blue; type II, red; type III, green; the variable region (V) is also indicated], with indication of the main domains involved in matrix assembly or cell binding; the FN dimer forms via two disulphide bonds at the C-terminus. b The material-driven fibrillogenesis leads to the formation of a FN network through interactions between I1-5 and III1-2 (or III12-14) domains. FN molecules into the network expose cell binding domains, resulting in a biologically active FN network that leads to the activation of cellular signalling pathways after cell-protein binding
Fibronectin contains domains to interact with other ECM proteins, glycosaminoglicans (GAGs), integrins, growth factors, other FN molecules and also pathogens, such as bacteria. 24,25,51 In particular, three domains are essential for FN matrix assembly: the FN dimerisation domain, the 70-kDa fragment and the RGD sequence. Fibronectin dimerisation depends on the covalent association of the subunits mediated by a pair of disulphide bonds at the C-terminus of the FN molecule (Fig. 2). The 70-kDa fragment extends from type I1 to I9, including the N-terminal assembly and the collagen/gelatin binding domains. This fragment binds to cells in monolayer cultures and blocks FN matrix assembly when added in excess. Similarly, the assembly is blocked when antibodies to this region are used. 52,53 Finally, the Arg-Gly-Asp (RGD) sequence is located in the repeat III10, next to the synergy sequence PHSRN in the repeat III9. Its binding to α5β1 integrin is required for the cell-mediated FN assembly to occur. 54 Although both sites are required for fibril formation, the synergy site is not essential because matrix levels are drastically reduced but not ablated with FN lacking this sequence. 55 Other FN binding domains are implicated in matrix assembly: III1–2 and III12–14 can bind FN; in addition, III1 can bind to III7, and III2–3 can interact with III12–14. 24 All these domains can promote FN fibrillogenesis because of the property of binding FN, but they can also participate in intramolecular interactions that keep soluble FN in a compact form. 56 A recent FN fibre model has predicted that the hierarchy of repeat-unfolding within fibrils is controlled by their relative mechanical stabilities (as found for single molecules) and by the strength of interactions between adjacent molecules, which are activated by stretching: the force-induced exposure of cryptic sites regulates the non-linearity of stress–strain curves (strain at which such fibres break). 57
Fibronectin fibrillogenesis and regulation of matrix assembly
Fibronectin in solution has a compact conformation and does not form fibrils even at extremely high concentrations. This compact form is maintained by intramolecular interactions between III2–3 and III12–14 modules. 56 Fibronectin-integrin binding induces integrin clustering, which groups together cytoplasmic molecules, such as FAK, Src Kinase, paxillin and others, promoting the formation of focal complexes. These complexes activate the polymerisation of the actin cytoskeleton and kinase cascades-mediated intracellular signalling pathways. 44 Cell membrane receptor clustering by dimeric FN helps to organise FN into short fibrils. After that, the contractility of the cell cytoskeleton contributes to FN fibril formation. 58,59 The stretching because of cell contractility provokes a progressive extension of the FN molecule and the exposure of binding sites that mediate lateral interactions between FN molecules. Fibronectin has intrinsic protein-disulphide isomerase activity in the C-terminal module I12, and this activity may introduce intermolecular disulphide bonds within fibrils. 26 Initial thin fibrils (5 nm diameter) then grow in length and thickness (25 nm diameter) as the matrix matures, and FN fibrils are converted in an insoluble form (Fig. 3). 45 Proper integration of extracellular signals with active intracellular pathways plays a crucial role in the initiation, progression and regulation of FN matrix assembly. 60
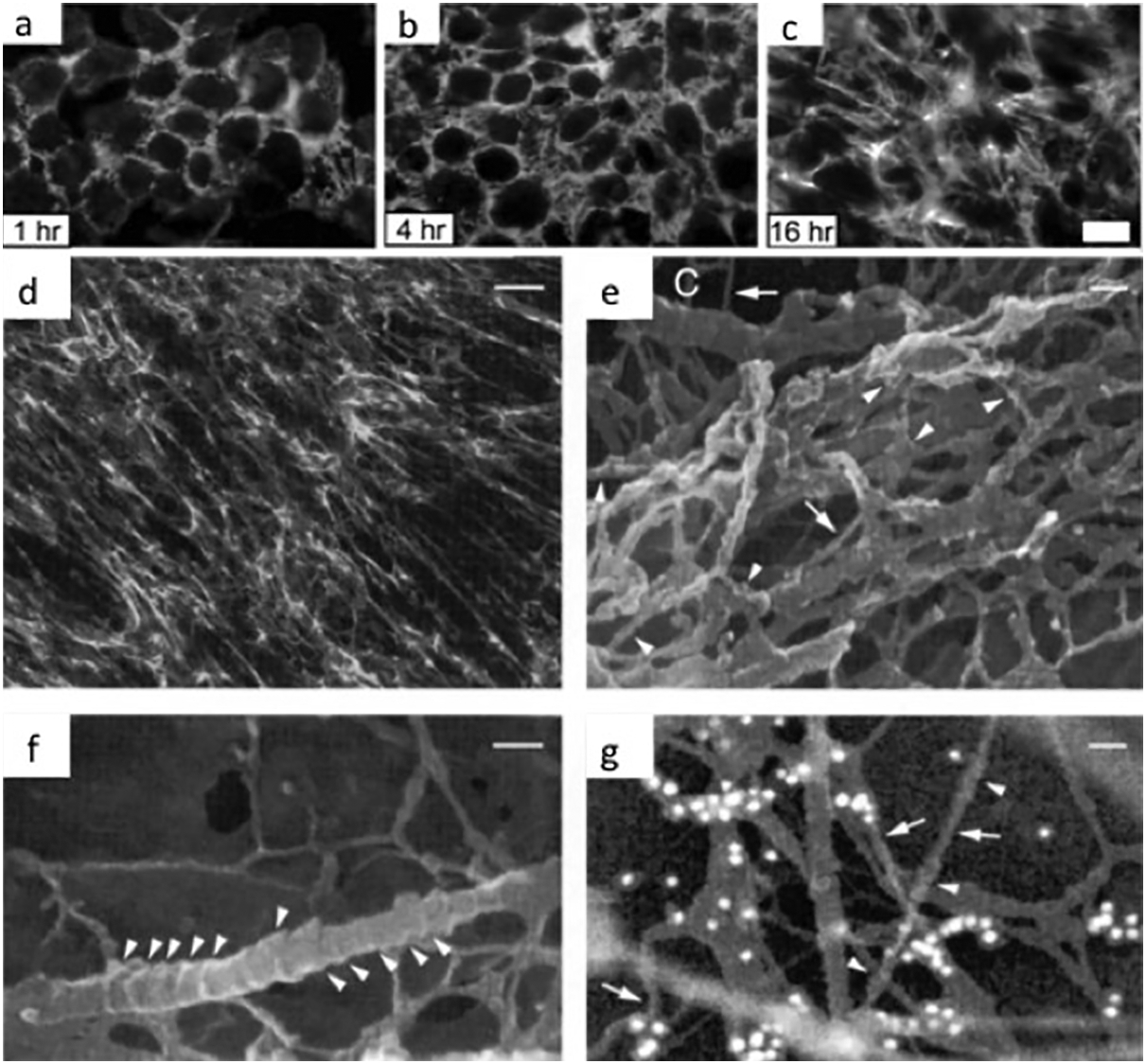
Natural fibronectin (FN) matrices. a–c Time course of fibril formation by CHOα5 cells cultured in medium containing FN-depleted serum and incubated with 25 μg mL−1 pFN for the times indicated. Fibronectin fibrils were visualised by indirect immunofluorescence with monoclonal antibody IC3 specific for rat FN. d–g Immunofluorescence and cryo-HrSEM micrographs of FN fibrils formed in human foreskin fibroblast cultures. Immunofluorescence microscopy d indicates that fibroblast cultures produce an extensive network of FN fibrils. Cryo-high-resolution electron micrographs e–g show that FN matrices in these cultures contain numerous nodular structures (arrowheads, e and g) and smooth regions (arrows in e). Occasionally, collagen fibrils (labelled C in e) are also observed in these cultures. They were readily identified by their distinctive banding pattern (arrowheads, f). Bar 10 μm a–c, 13·3 um d and 45 nm e–g. Reproduced with permission from Ref. 55 a–c and 61 d–g
In a recent study, the onset of fibrillar ordering because of FN fibrils formation and maturation was followed, dynamically and label-free, using sum-frequency-generation (SFG) spectroscopy (Fig. 4a ). 62 Sum-frequency-generation is a technique used to analyse surfaces and interfaces using two laser beams that overlap at a surface, generating an output beam with a frequency equal to the sum of the two input frequencies. This allows deducing the composition, orientation distributions, and some structural information of molecules. Fibronectin fibrils in fact display amyloid structures, implying an ordered conformation detectable with SFG spectroscopy (Fig. 4b ). Sum-frequency-generation results of time-lapse experiments on living cells show the presence of ordered structures as soon as 1 h after cell seeding, and between 4 and 6 h, the major ordering processes in the ECM occurred, as indicated by a large increase in the resonant signal (Fig. 4c ). This same methodology can be applied to analyse artificially ordered protein structures and compare them with their biological counterparts (Fig. 4b , Force-based assembly section). 62
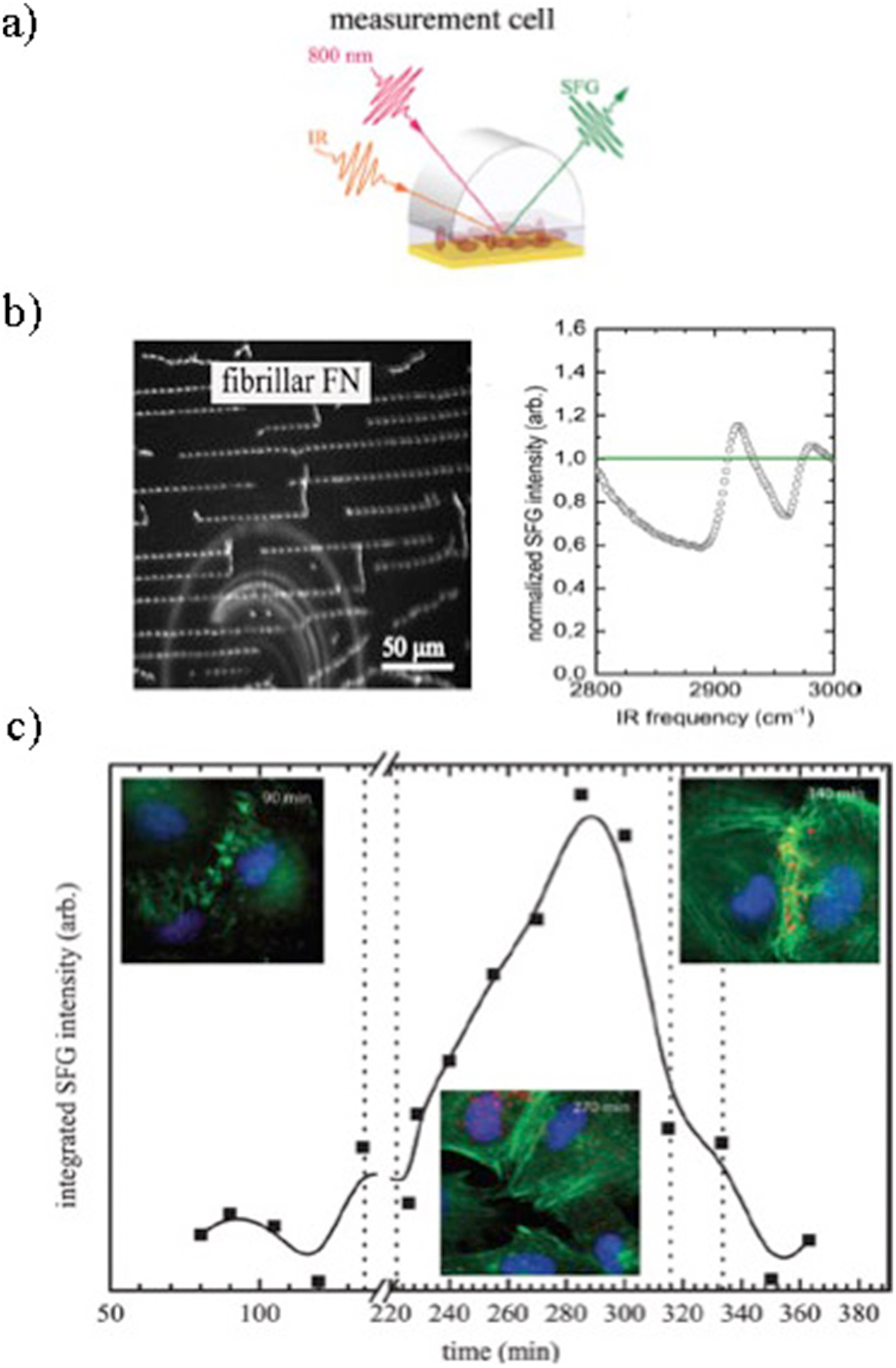
In vitro observation of dynamic ordering processes in the extracellular matrix (ECM) of living, adherent cells. a Schematic depiction of the sum-frequency-generation (SFG) measurement setup used. b Fibrillar fibronectin (FN) (obtained by applying force to globular proteins, promoting dimer unfolding and lateral association) on Au-coated substrate (scale bar 50 μm) and resonant contributions to SFG spectra of fibrillar FN. c Integrated SFG intensity in the CH region over time. Au-coated Si wafers with a confluent layer of REF52wt cells adhering to them were exchanged at the time points indicated by the vertical bars. The curved lines are guides for the eye and do not represent a fit of the data shown. Fluorescent micrographs show REF52wt cells stained for DNA (DAPI, blue), actin (phalloidin, green) and FN (MAB1926, red). The micrographs were taken at the times indicated on the pictures as part of a 30 min time-lapse series. Reproduced with permission from Ref. 62
A complete up-to-date review of in vivo FN fibrillogenesis can be found in Schwarzbauer and DeSimone. 25
Artificial fibronectin matrices
Engineering materials that recapitulate the characteristics of the ECM, such as materials presenting cell-adhesive motifs or protease-degradable cross-links, 4,63 have been a demanding challenge in regenerative medicine research, aiming at controlling and directing cellular responses. Indeed, as anticipated in the previous section, the assembly of a FN matrix in vivo is the initial step that orchestrates the organisation of other ECM proteins; furthermore, it promotes cell adhesion, migration, signalling and controls the availability of other bioactive molecules within the ECM, such as growth factors. Therefore, the assembly of FN matrices has a critical role in normal cell adhesion and growth during tissue homoeostasis and repair. Considering that this initial interaction between cells and the surrounding microenvironment will determine cell fate, engineering biomaterials with the ability to induce cell-free fibrillogenesis of FN provides us with the opportunity to modulate the early interaction of the cells with the materials surface in a physiological-like manner. This strategy would then allow to instruct cells promptly and efficiently towards the desired fate, by imprinting a physiological microenvironment on cells that promotes tissue regeneration. Within this respect, different approaches to reconstitute the network structure and bioactivity of FN fibrillar matrices have been proposed. Cell-free routes to induce FN fibrillogenesis are based on the assumption that unfolding of soluble FN dimers from their globular conformation is needed for FN–FN interactions to occur, leading eventually to FN polymerisation and fibril formation. Methods proposed in literature so far include assembly via chemical agents (addition of reducing 63,64 or oxidising 65 agents to the protein solution; use of denaturing, 61,66–68 cationic 69,70 or anionic 71,72 compounds), use of peptidic FN fragments; 73–75 force-based assembly, via application of mechanical tension 76–80 or shear forces; 81–90 and surface-initiated assembly 91–95 (see Table 1 for a complete classification of published methods). In these studies, FN used for the production of fibrillar matrices is generally obtained from human plasma.
Methods for the assembly of artificial fibronectin matrices
A list of different cell-free routes used in literature to induce fibronectin (FN) fibrillogenesis, with emphasis on the agent responsible for the polymerisation of FN fibrils, the mechanisms involved and their similarity with cell-produced FN matrices.
Assembly via chemical agents
Reducing agents, such as dithiothreitol (DTT), have been shown to induce the self-assembly of plasma FN via unfolding of disulphide-stabilised globular domains and subsequent promotion of noncovalent binding interactions among FN molecules. 64 At low DTT concentration, the formation of FN multimers involves a disulphide exchange mechanism from intramolecular to intermolecular bonds and interactions through the terminal region of the FN chains, as well as type I and type II repeats. 63 Low concentrations of strong oxidants, such as FeCl3 and CuSO4, were also reported to induce disulphide-bonded FN polymerisation. 65
Another cell-free method to induce FN fibrillogenesis in vitro involves the addition of denaturants to a protein solution. Chemical denaturants provoke the alteration and loss of the native structure of a protein, leading, in the case of FN, to the unfolding of its globular conformation in solution. Mosher and coworkers first used guanidine as a denaturing agent; 66–68 denatured FN dimers partially expose their free sulphydryl groups, allowing multimeric FN to form. This multimeric FN forms fibrillar structures similar to the ones produced in culture by human skin fibroblasts, as observed using high-resolution cryo-scanning electron microscopy (cryo-HRSEM) (Fig. 3e–g ). 61,68 Two distinct structural arrangements (a rope-like structure with one free end, consisting of 10- to 13-nm diameter nodules, and a constrained straight conformation with smooth surface) were observed, depending on the tension applied to the fibrils. 61 The nodules were proposed to represent discrete domains of three to four type III repeats, as they could be labelled with the monoclonal antibody IST-2 to the III13–14 repeats in FN, and they were observed in FN fragments that only contain type III repeats. 61
Also, anionic molecules, specifically heparin, were found to induce FN fibrillogenesis in the absence of cells. 71,72 Heparin induces the transition of FN from a globular to an elongated form, exposing masked binding sites responsible for self-association, and therefore, leading to the formation of fibrillar precipitates. 71,72 Precipitation was considerably dependent on the ionic strength, indicating that electrostatic forces played a major role in the aggregation of FN. Electrostatic interactions were also involved in FN polymerisation induced by polyamines (cationic molecules). 69,70
Assembly via recombinant FN fragments
Interaction of FN dimers with purified recombinant fragments of FN obtained by bacterial protein expression (this is a standard biotechnological procedure based on the genetic modification of bacteria to produce proteins with specific sequences of amino acids, in this case FN fragments) was shown to induce fibrillogenesis in the absence of cells. 73–75 Treatment of FN with anastellin, a C-terminal recombinant fragment from the first type III repeat of FN (III1-C), converted FN into high-relative molecular mass multimers resembling matrix fibrils (termed superfibronectin as a result of their greatly enhanced adhesive properties), probably through disulphide exchange. 73 The main structural features of anastellin resemble those of amyloid fibril precursors, implying some similarity between FN and amyloid fibril formation. 75 Engineered FN mutants (full-length FN molecules with specific mutations along their sequence, obtained by site-directed mutagenesis) revealed that the unfolding of the III2 domain is one of the key factors in anastellin-induced FN aggregation. 96 Another aggregation-inducing fragment is the recombinant III10 FN module: 74 the interaction of the III10 fragment with the III1 domain in intact FN promotes a conformational change within the III1 module, unmasking the amino-terminal binding site and triggering the self-polymerisation of FN.
Force-based assembly
The importance of molecular unfolding for FN fibrillogenesis was confirmed by Zhong et al. by applying mechanical tension to expose the self-assembly sites within FN, thereby enhancing the binding of soluble FN in the absence of cells. 76 The role of mechanical tension for the formation of FN fibrils was further investigated by Baneyx and Vogel, who observed FN assembly into extended fibrillar networks after adsorption to a dipalmitoyl phosphatidylcholine (DPPC) monolayer in contact with physiological buffer. 77 The mechanical tension caused by domain separation of the lipid monolayer pulled the proteins into an extended conformation, exposing cryptic self-assembly sites and allowing the formation of networks stabilised by disulphide cross-linking.
Spatz et al. also induced fibrillogenesis by applying mechanical tension to FN. 78–80 This group developed a system where cell-free FN fibrillogenesis could be achieved through a two-stage process initiated by the shear-stress independent partitioning of globular FN molecules to the air/liquid interface, where they formed an insoluble two-dimensional sheet, followed by force-dependent fibrillogenesis along a superhydrophobic surface made of elastic micropillars. Fibronectin fibre formation requires proper alignment of the outer type I domains, enabling specific intermolecular interactions between the type I domains of different chains within the air/water interface sheet. 97,98 Subsequent application of force to this FN sheet attached to a micropillar array resulted in the formation of rough fibres with globular subdomains, whose sizes were affected by the tension applied to the FN layer. The average force applied to the FN particles by micropillar bending was approximately 8 pN/molecule, which is within the range necessary for FN unfolding. 99–101 This force-induced FN fibrillogenesis was adapted to obtain a method for the production of highly regular arrays of nanofibrils from other ECM proteins besides FN, including collagen I (COL I) and laminin. 80 These results were congruent with the model proposed by Guan et al. for the fabrication of DNA nanowires, indicating a common mechanism of polymer fibrillogenesis induced by dewetting. 102 The necessity for self-association sites was suggested by the observation that bovine serum albumin (BSA), even at high concentrations, did not yield nanofibrils, despite its ability to concentrate at the air/buffer interface. 78,80 As anticipated in Fibronectin fibrillogenesis and regulation of matrix assembly section, the well-ordered conformation of the FN fibrils obtained through this methodology could be detected via SFG spectroscopy (Fig. 4b ), confirming that the application of mechanical stress to FN is similar to fibrillogenesis in vivo. 62
Other cell-free routes for the force-induced assembly of FN were based on the application of shear forces to a FN solution in different set-ups that included manual pulling of single fibrils out of a concentrated drop of FN, 81,83,84,87,89,90 stirring of a FN solution in a ultrafiltration cell, 81,82,85,88 and wet extrusion of a concentrated FN solution. 86,88,103 In the first method, FN was allowed to self-associate under the influence of a directional shear force. 81 These manually deposited fibres resembled in vivo FN fibres in diameter and composition. 89 Mechanical characterisation of these fibres revealed that they show signs of breakage only when stretched over 5–6 times their resting length that high extensions involve unfolding of type III modules, and that the mechanically strained fibres clearly bind more N-terminal 70 kD FN fragment than the relaxed ones, suggesting that mechanical tension exposes cryptic binding sites for this fragment. 89
Instead of fibres, concentration of a FN solution under a continuous unidirectional stirring motion was used to produce oriented FN mats. 81,82,85,88 The yield of FN mat was drastically improved in the presence of urea, which denatures FN by unfolding it into an elongated conformation 104 that favours lateral association and fibril growth. 82 Finally, shear-dependent fibrillogenesis was achieved via wet extrusion of urea-denatured concentrated protein solutions, 86,88,103 leading to large FN cables with parallel fibril alignment. 88
Material-driven assembly
The last cell-free route of FN fibrillogenesis is based on the unfolding of soluble FN as a result of its adsorption onto a material surface. 91–95 These methods rely upon the effects of surface chemistry on passive protein adsorption, as anticipated in Cell/material interaction section. It is in fact known that surface chemistry plays a pivotal role in determining the conformation of proteins adsorbed from serum, mixture or single protein solutions, as well as the rate of adsorption and the amount and composition of the adsorbed protein layer. The process depends on the interactions between the functional groups on the substrate and those of the proteins themselves. 105,106 In the case of FN, hydrophilic and negatively charged surfaces were shown to promote the extension of its dimer arms upon adsorption, while hydrophobic surfaces provoked the disruption of the secondary structures of the protein. 37,38,107–109 Nelea and Kaartinen found that FN filaments could be formed by adsorbing the protein on a surface with negative potential (polysulphonated substrate) in the absence of cells. 93 From the height profile analysis of AFM scans, the filaments were revealed to be formed of a chain with periodic arrangement of connected beads giving a ‘bead-on-a-string’ appearance. 93 This is in accordance with the model of FN adsorption onto a high negative charge density surface proposed by Pernodet et al.: electrostatic interaction of the surface with the III12–14 modules of FN would open the molecule, allowing its arms to extend. 91 Then, interactions between I1–5 and III12–14 modules of two FN molecules would create a staggered alignment, 110–112 and position two type III1–7 modules (reported to drive FN fibrillogenesis 113 ) interacting with each other. This would create a thicker stretch in the filament with the appearance of a bead.
Feinberg and Parker demonstrated that surface-initiated assembly could be used to engineer multiscale, free-standing nanofabrics using a variety of ECM proteins (FN, laminin, fibrinogen, collagens). 94 The process involved adsorbing nanometre-thick layers of ECM proteins from a solution onto a hydrophobic surface at high density to partially unfold them and expose cryptic binding domains, 107,114 transferring the ECM proteins in the unfolded state to a relatively hydrophilic, dissolvable surface, and thermally triggering surface dissolution to synchronise matrix assembly and its non-destructive release (Fig. 5). The analysis of the response of the FN nanofabrics to mechanical load revealed that the elastic properties of FN depend on the conformational extension of the protein, while the plastic deformation depends on domain unfolding (Fig. 5). 115 According to this study, cell-generated forces, being well below the average value associated with plastic deformation, would not be large enough to trigger domain unfolding and plastic deformation of the ECM, accounting for the elastic behaviour of ECM in vivo.

Stretched fibronectin (FN) nanofabrics are highly extensible with a nodular/fibrillar change in nanostructure under strain. a Schematic of a nanofabric composed of FN 20×20 lines at 90° before release and b stretched post-release. c SEM image of a FN nanofabric stretched longitudinally demonstrating that large strains are supported. d Zoomed region from c shows that the horizontal lines (yellow ▾) have contracted to 3 μm wide, whereas the vertical lines (yellow) have been stretched to 45 μm wide, a 15-fold difference. e Optical image of a stretched nanofabric, a section of which was imaged by AFM. f Region (1) shows the high-density nodular nanostructure of the horizontal lines that have contracted laterally. Region (2) shows the higher density nodules, where the horizontal and vertical lines overlap. Region (3) shows the low-density nodules and fibrillar appearance of the vertical lines stretched between two lateral lines. g Zoomed region from f reveals the transition in nodular structure from the overlap region to the more fibrillar region. h Zoomed region from g reveals what appears to be stretched fibrils (white ▾), connecting two larger nodules with a smaller nodule in-between. i Zoomed region from g illustrates the linear arrangement of nodules like ‘beads-on-a-string’ aligned in the direction of stretch suggesting these nodules are coupled by unfolded FN. j A 3D rendering from the region in i emphasises the nodular structure (white ▾) and a region (as denoted by white ‘*’) that contains a meshwork of smaller fibrils running between nodules. Nodule diameter is consistent with folded and partially refolded FN dimers, and the height of the fibrils connecting the nodules is consistent with unfolded FN dimers. k Prestress in the stretched FN nanofabrics is confirmed by examining the morphology at points of failure. l Rupture of a FN 20×20 line (yellow) reveals an apparent 3-fold contraction from an initial length of 15–5 μm. The compressive stress from the failure is sufficient to cause periodic buckling (yellow ▾) in the underlying, orthogonal nanofabric. m An intact orthogonal junction. Scale bars are 100 μm for c; 10 μm for d,e,k; 1 μm for l,m. Reproduced with permission from Ref. 94
In our group, the authors hypothesised that adsorption of individual FN molecules onto particular surface chemistries would induce exposure of self-assembly sites, driving FN fibril assembly, and they identified poly(ethyl acrylate) (PEA) as a potential surface chemistry suitable for this purpose. 116,117 The authors investigated the organisation of FN molecules at the material interface and studied its analogy with the physiological cell-induced FN fibrillogenesis. In particular, it was observed that a FN solution onto a PEA surface gave rise to protein fibrils, as revealed by AFM: an interconnected network of FN fibrils was identified (Fig. 6a ). To further address the mechanism underlying this material-driven fibrillogenesis, protein organisation at the material interface was investigated on two similar chemistries: PEA and poly(methyl acrylate) (PMA), which differ in one single carbon in the side chain (Fig. 6b ). These surface chemistries show similar wettability and total amount of adsorbed FN; however, only PEA induces material-driven fibrillogenesis, whereas dispersed globular molecules are present on PMA. Strikingly, material-driven fibrillogenesis absolutely requires the 70 kDa amino-terminal region of FN, known to be essential for cell-mediated assembly, 24,118 as reported in previous paragraphs. Indeed, addition of the 70 kDa fragment completely blocks the organisation of FN at the material interface and only discrete molecular aggregates can be observed, resembling the protein adsorbed onto the control PMA polymer (Fig. 6b ).
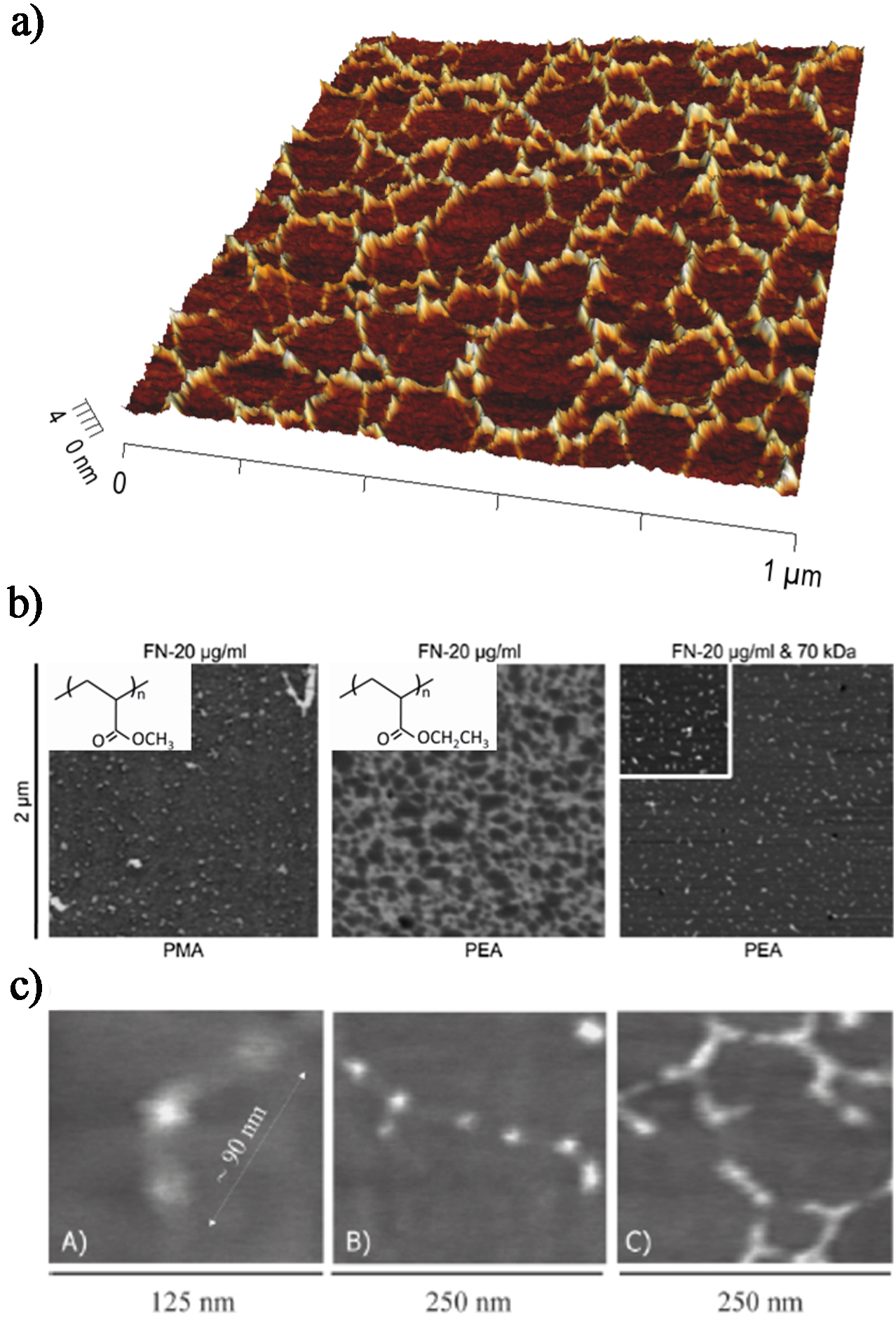
Organisation of fibronectin (FN) molecules in solution upon adsorption onto poly(ethyl acrylate) (PEA) and poly(methyl acrylate) (PMA) surfaces as observed via tapping mode AFM. a Combined 3D height image and phase image of a FN network formed after adsorption from a 20 μg mL−1 protein solution on PEA. b FN distribution on material substrates as obtained by phase imaging: globular aggregates on PMA and FN network on PEA after adsorption from a solution of concentration 20 μg mL−1. Fibronectin fibrillogenesis is blocked on PEA in the presence of the amino-terminal 70 kDa FN fragment, leading to dispersed molecular aggregates. The inset squares show the chemical structure of the polymers. Reproduced with permission from Ref. 95 cA. Isolated FN molecule on PEA, cB two FN molecules interacting through the amino-terminal (I1-5 and III1-2) domains, cC assembly of FN molecules into an incipient network. Reproduced with permission from Ref. 117
The material-driven FN fibrillogenesis onto PEA was found to be a time- and concentration-dependent process, whose dynamics were followed via AFM at different adsorption times, 119 or at increasing protein concentration (for a fixed adsorption time). 117 Based on these observations, the proposed model of material-driven FN fibrillogenesis includes the following sequence of events:
Conformational change of FN upon adsorption onto PEA. Fibronectin has a compact folded structure in physiological buffer, stabilised through ionic interactions between its arms. 56 Its interaction with chemical groups of the substrate (a vinyl backbone with -COOCH2CH3 side chain) provokes conformational changes in the molecule that lead to the extension of the protein arms (Fig. 6c A). Adsorption onto slightly charged surfaces (net negative charge in the -COO− group) gives rise to elongated structures of the molecule, as obtained for SiO2 and glass. 107,114 Fibronectin likely orients at the surface, so that its hydrophobic segments interact with the methyl groups of PEA, maybe throughout the heparin-binding fragment, as proposed for the FN–DPPC interaction, 77 but with more efficient arm extension because of the net negative charge of the surface.
Enhanced FN–FN interaction on the PEA substrate. As the adsorption process continues, the conformation of individual FN molecules favours FN–FN interactions involving the amino-terminal 70 kDa fragment, 107 probably throughout the interaction between I1–5 and III1–2 domains located near the amino side 51 (Fig. 6c B).
Formation of the FN network. New FN molecules are preferentially adsorbed in close contact to the proteins already present in the substrate (Fig. 6c C), probably as a consequence of the presence of polar-oriented FN molecules; this enhances the collision rate of FN self-assembly sites, 77 eventually leading to a well-interconnected network of FN on the surface of the substrate (Fig. 6a ). 117 The formation of a network on PEA is not a property unique to FN. For example, a similar network was found for fibrinogen 120 and collagen IV, 121 but only globular-isolated molecules were observed after laminin 122 and VN adsorption. 123
In agreement with the proposed model of material-driven fibrillogenesis, faster dynamics of FN network formation were observed on polymer surfaces similar to PEA, but with higher molecular mobility [poly(butyl acrylate), PBA, and poly(hexyl acrylate), PHA]. 124,125 On these polymers, which differ only in the number of methyl groups in the side chain compared to PEA, FN adsorption from solutions of increasing concentration leads to the formation of a well-developed protein network at a lower concentration of the solution as the number of methyl groups in the side chain of the polymer increases. The glass transition temperature of PBA and PHA is lower than that of PEA, which means that surface mobility is enhanced on these polymers. Hence, the mobility of the protein adsorbed at the material interface is enhanced, increasing the collision rate of FN self-assembly sites and the probability of FN–FN interaction as new molecules are adsorbed onto the surface. By contrast, only globular FN molecules are adsorbed onto PMA, which supports the idea that minute variations in polymer chemistry can alter FN conformation during adsorption.
Similarly, the coadsorption of FN with small amounts of a lower molecular weight protein, such as VN, which does not contain any specific binding domain for FN or other VN molecules, resulted in the entrapment of VN within the FN network, conferring it higher mobility and allowing for faster dynamics of network formation onto PEA. 126
Biological activity of artificial fibronectin matrices
The cell-free routes reported in the previous section to induce polymerisation of FN dimers generate fibrils that are claimed to be morphologically and/or functionally similar to physiological matrix fibrils. Within this respect, these methods lead to fibrillogenesis, and not sole aggregation, of the protein, involving the unfolding of globular dimers and their lateral association. The biological activity and functionality of these fibrils may be assessed on the one hand by comparing its structure with fibrils produced by cells (e.g. through microscopy techniques or antibody binding, as previously described), on the other by evaluating the response of cells cultured onto them (as will be explained in the following paragraphs). Key aspects confirming the similarity of artificial fibrils to physiological ones are for example the enhanced exposure of antigenic epitopes otherwise hindered, including cell-adhesive domains, and the involvement of the 70 kDa fragment in the lateral association between dimers. 45
Cell adhesion and matrix secretion
Given its similarities with the physiological assembly of FN, the material-driven FN fibrillogenesis presented in Material-driven assembly section yielded a protein network with enhanced biological activity. The conformation adopted by the protein promoted the exposure of domains that favour the interaction with cells and other proteins (Fig. 2). Indeed, the binding of HFN7·1 antibody (a receptor-mimetic probe for integrin binding and cell adhesion) 127 was enhanced on the material-driven network (Fig. 7a ). 128 It is known that differences in the availability of FN adhesion domains after protein adsorption influence the initial cell–material interaction, in terms of signalling (Fig. 7b ), integrin expression, focal adhesion formation and F-actin cytoskeleton development (as explained in Cell/material interaction section). When protein and gene expression were investigated in the absence of any additional serum proteins (serum contains large amounts of FN and VN, which might perturb the effect of the provisional FN matrix previously assembled), α5 was shown to be the main receptor involved during the initial cell interaction with the material-driven FN network on PEA, as it happens for the interaction of cells with natural-physiological matrices. 54 α5β1 binding has been related to the simultaneous availability of the synergy and RGD sequences within FN. 129 On the contrary, when FN was adsorbed as discrete aggregates on material surfaces, the availability of the whole integrin binding site of FN was reduced and cell adhesion mostly occurred through the αv receptor (Fig. 7b ), which only needs the exposure of the RGD sequence. The phosphorylation of FAK (non-receptor protein-tyrosine kinase that becomes activated in response to cell-matrix adhesion) was also enhanced on the material-driven FN network compared to the same amount of FN adsorbed as globular aggregates on other surfaces. 128 Overall, the assembly of FN fibrillar matrices at the material interface involves the formation of larger focal adhesions as compared with the globular adsorption of FN. 130
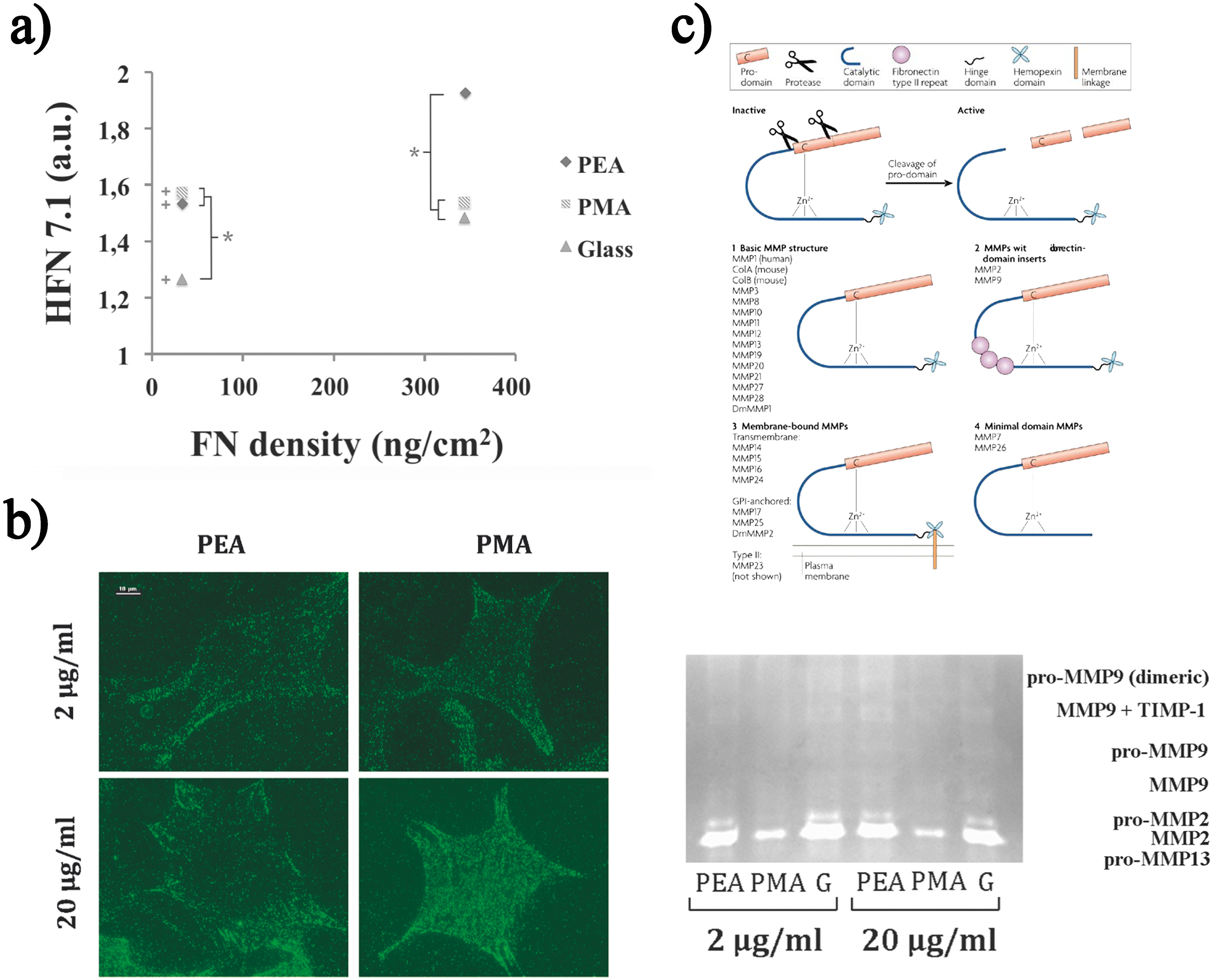
Effect of material-driven fibrillogenesis on signalling, integrin expression and matrix protein degradation. a Activity of the adsorbed fibronectin (FN) on different substrates obtained by plotting the binding of HFN7·1 monoclonal antibody versus FN surface density measured on different substrates from solutions of different concentrations. b α5 integrins bound to adsorbed FN at the material interface are observed after cross-linking and extraction of cellular components (scale 10 μm). c Gelatin zymography shows the activity of MMP2, 9 and 13 and of their various forms (pro-MMP13, MMP2, pro-MMP2, MMP9, pro-MMP9, MMP9+TIMP1, dimeric pro-MMP9). Error bars represent the standard deviation of three independent experiments. (+) Indicates whether there is a statistically significant difference between the conditions of 2–20 μg mL−1 and (*) shows conditions with significant difference (p<0·05). Reproduced with permission from Ref. 128
Another evidence of the biological activity of this FN network was obtained by investigating cell adhesion on electrospun fibres of PEA. 117 Fibroblast adhesion studies demonstrated that the FN network is biologically active, more than the FN adsorbed on the underlying glass, supporting the hypothesis that the material-driven FN network shares some similarities with the physiological one. Cells tended to interact with PEA fibres rather than glass and started to orient and migrate, modifying their characteristic spread morphology, following the fibres direction; cells adhering to the material-driven FN network showed well-developed focal adhesion complexes and actin stress fibres. 117
Fibronectin fibrils and matrices obtained through other cell-free routes also enhance adhesion of different cell types. For example, when different cell types were seeded on FN fibrils obtained by the application of a directional shear force, cells adhered and aligned along the axis of the fibres, confirming that cell adhesion sites are exposed on their surface. 81,83,84,88,89,94 Moreover, the presence of FN fibrils significantly promoted the migration of human dermal fibroblasts and the proliferation of Schwann cells. 87–88 In other studies, the focal contacts and actin microfilaments of Schwann cells, macrophages and fibroblasts were found to be oriented in the direction of the fibrils, 83,84 which is consistent with the fact that formation of focal adhesions within certain directions predetermines the orientation and guidance of the whole cell. Therefore, these studies reveal the potential of these shear force-generated FN fibrils to guide and direct cellular behaviour by contact guidance. Ulmer et al. showed that fibrils generated by applying mechanical tension to FN were quite robust and could support cell attachment and focal adhesion assembly. 78 Surface-initiated FN nanofabrics were also shown to be able to sustain and direct the adhesion of cardiomyocytes, and even to support cyclic contraction over several days of culture. 94 On the other hand, FN fibrils assembled via recombinant FN fragments were shown to greatly enhance cell adhesion and spreading, in such a way that reduced or even suppressed cell migration. 73
There are a few examples in the literature that shows how material-driven FN fibrils can direct the spatial organisation of cell-secreted FN matrices, which reflects the high biological activity of the pre-organised FN at the material interface. 117 The FN network assembled on PEA electrospun fibres influenced the secretion of FN that was preferentially deposited following the direction of the PEA fibres. 117 Moreover, FN was shown to assemble into network-like patterns on electrospun poly-acrylonitrile methylacrylate (PAN-MA) fibres. These fibrils afterwards promoted that Schwann cell-generated FN matrix was also organised in a topographically sensitive manner. 131 Thus, one can assume that, by tailoring the fibre orientation, the organisation of the provisional FN matrix secreted by the cells can be tuned, which is a way to engineer the matrix and imprint on the matrix assembled by cells on the dorsal side, the spatial-topographical information previously read from the ventral material-driven FN network.
Matrix remodelling
Besides organisation, the ECM undergoes proteolytic degradation, which is a mechanism for the removal of the excess matrix usually associated with remodelling and mainly operated by a family of proteases known as MMPs. The proteolytic remodelling of matrix proteins at the biomaterials interface has received attention in the last 10 years. 132–136 This led to the engineering of material systems, which incorporate protease-degradable sequences in addition to cell adhesion motifs. 15,136 In our group, the authors have shown that the activation of proteolytic routes in MC3T3-E1 cells is a MMP-dependent phenomenon sensitive to surface chemistry by making use of FN-coated SAMs with a controlled ratio of methyl/hydroxyl groups. 34 To investigate the existence of a link between material-driven FN assembly and matrix protein remodelling (degradation) at the cell–material interface, FN was adsorbed onto PEA and PMA, which result in different protein distribution and conformation as previously explained. 128 MMP9 activity was higher on the assembled FN fibrils (Fig. 7c ), as FN activity was (Fig. 7a ). Moreover, MMP2 and MMP13 activities (zymography) were much higher on the material-assembled FN matrix. MMP2 has FN type II repeats inserted into the catalytic domain and has been found to cleave FN and VN into small fragments in vivo, leading to increased cell adhesion and migration. 132,133 In this system, degradation of the assembled fibrils at the material interface would enhance the exposure of adhesion sites, which would in term support the enhanced integrin (α5) expression and focal adhesion formation explained in the previous section. This also uncovers the relationship between FN activity at the cell–material interface, the ability to reorganise the underlying layer of proteins at the material interface and proteolytic cascades. Higher MMPs activity (Fig. 7c ) is required to remodel the provisional matrix when cells are not able to reorganise this layer of proteins at the material interface, as it occurs in the case of the material-driven FN network. 124,137 It is precisely the initial proteolytic activity (after 4 h) that enhances cellular behaviour on the FN fibrils assembled on PEA.
In summary, cell adhesion on material-driven FN fibrils occurs primarily through the specific α5β1 receptor occupancy, with enhanced focal adhesion formation and FAK signalling. MMP activity is also enhanced, through a mechanism that is dependent on the ability of cells to reorganise the adjacent layer of proteins at the material interface.
Cell differentiation
Besides supporting enhanced cell adhesion, matrix secretion and degradation, material-driven FN fibrillogenesis was also shown to affect higher order cellular functions, such as cell differentiation. Particularly, the biological activity of material-driven FN networks was examined using the ability to induce myogenic differentiation (Fig. 8). The material-induced FN network supported significantly higher sarcomeric myosin expression and cell bipolar alignment and fusion into myotubes, markers of myogenesis (Fig. 8b , PEA-20), as compared to the same material coated with a lower density of FN that does not lead to the fibrillar organisation of the protein at the material interface, and other polymers lacking any fibrillar organisation (Fig. 8b ). 95,108
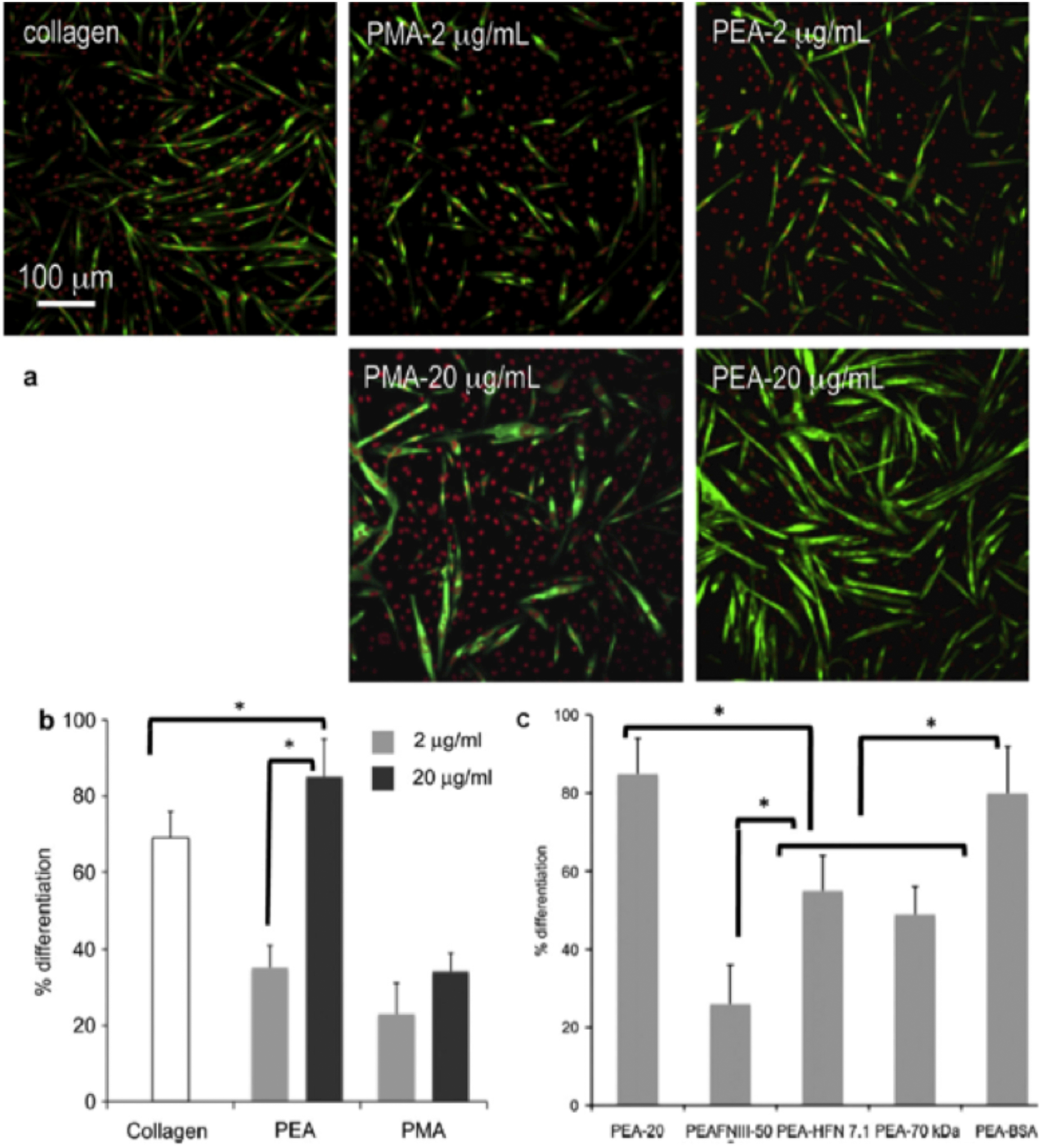
Myogenic differentiation on different substrates. a Fluorescence staining showing sarcomeric myosin-positive cells (green) and cell nuclei (red) on fibronectin (FN)-coated surfaces (from solutions of concentration 2 and 20 mg mL−1) and control collagen. b Myogenic differentiation as determined by the percentage of sarcomeric myosin-positive cells on different substrates after adsorbing FN from solutions of concentrations 2 and 20 μg mL−1. c Both the central FN domain (FNIII7–10) and the amino-terminal fragment (70-kDa) involved in FN fibrillogenesis enhance myogenic differentiation on the material-driven FN network on PEA. Myogenic differentiation as determined by the percentage of sarcomeric myosin-positive cells on the substrate-induced FN network (PEA-20), after coating PEA with a recombinant fragment of FN (FNIII7-10), blocking the central FN domain with HFN7·1 antibody (PEA-HFN7·1), after adsorbing FN altogether with the 70 kDa fragment, which blocked the formation of the FN network (PEA-70 kDa), and control experiment for the 70 kDa fragment using bovine serum albumin (BSA) instead (PEA-BSA). Statistically significant differences (p<0·05) are indicated with *. Reproduced with permission from Ref. 95
In order to gain a deeper comprehension of the phenomena underlying the biological activity of the substrate-induced FN fibrils, different blocking antibodies and FN fragments were used to assess the role of different parts of the FN molecule in the myogenic differentiation process (Fig. 8c ): (1) Addition of the adhesion-blocking HFN7·1 antibody inhibited differentiation to levels found for the substrates on which FN is not organised into a network (Fig. 8c , PEA-HFN7·1). HFN7·1 is specific for human FN (adsorbed on the substrate) and does not cross-react with mouse FN (cell secreted), indicating that the human FN adsorbed onto the substrate before cell seeding provided the dominant signal for differentiation; (2) Use of a recombinant fragment of FN spanning the seventh–tenth type III repeats (FNIII7–10) does not recapitulate the material-dependent differences in myogenic differentiation. 95 FNIII7–10 encompasses the integrin-binding domain of FN, but does not contain the domains involved in FN–FN interactions (I1–5 and III1–2 or III12–14 domains). Adsorption of this fragment onto PEA was found to yield minimal levels of myogenic differentiation (Fig. 8c , PEA-FNIII7–10), demonstrating that both integrin-binding domains and domains involved in FN–FN interactions are required for enhanced myogenic differentiation on PEA-driven FN matrices. Consistent with our observations for the FN network formation (Fig. 6b ), (3) addition of the 70 kDa amino-terminal fragment during the adsorption process of FN blocked the differentiation of myoblasts on PEA (Fig. 8c , PEA-70 kDa). 95
The effect of material-driven FN fibrillogenesis on myoblast differentiation was further studied by making use of gradients of adsorbed FN onto flat substrates of PEA and PMA. 138 The first polymer yields a gradient of interconnected fibrils, whereas on the second one, a higher amount of globular FN molecules was found along the sample. This kind of gradient wherein the sole amount of FN was varied, but where the globular-like conformation of the protein was maintained (PMA), did not yield any differential effect on myoblast differentiation and, still, supported cell migration. On the other hand, a gradient characterised by a gradual formation of FN fibrils (PEA) led to cell migration and to a gradient of myogenic differentiation. The authors attributed the increase in cell differentiation along this gradient to the gradual formation of a network of FN fibrils and a progressively higher cell density. 139
Beyond myogenic differentiation, the role of material-driven FN fibrillogenesis in skeletal mesenchymal stem cells differentiation has been investigated: 125 in this case, it was found that the differentiation towards the osteoblastic lineage, determined by quantifying protein levels for osteocalcin, osteopontin and Runx2, was enhanced on the substrate that does not induce the formation of a FN network. It was suggested that the FN adsorbed in a globular conformation targets skeletal stem cell differentiation along the osteoblastic lineage with greater efficiency compared to FN fibrils, as shown by the upregulation of characteristic osteoblastic markers osteocalcin and osteopontin. 125
Also in the field of osteogenesis, FN fibres prepared by manually pulling out of a concentrated protein drop (Force-based assembly section) were shown to upregulate osteogenic differentiation of hMCSs when stretched (Fig. 9). 140 Moreover, osteogenesis was increased when α ν β3 integrins were blocked on relaxed FN fibres, and decreased when α5β1 integrins were blocked (Fig. 9a ) or when epidermal growth factor (EGF) receptor signalling was inhibited on stretched FN fibres (Fig. 9b ), suggesting that the conformational change of FN caused by stretching might regulate integrin binding and growth factor signalling pathways, impacting hMSCs osteogenesis. 140
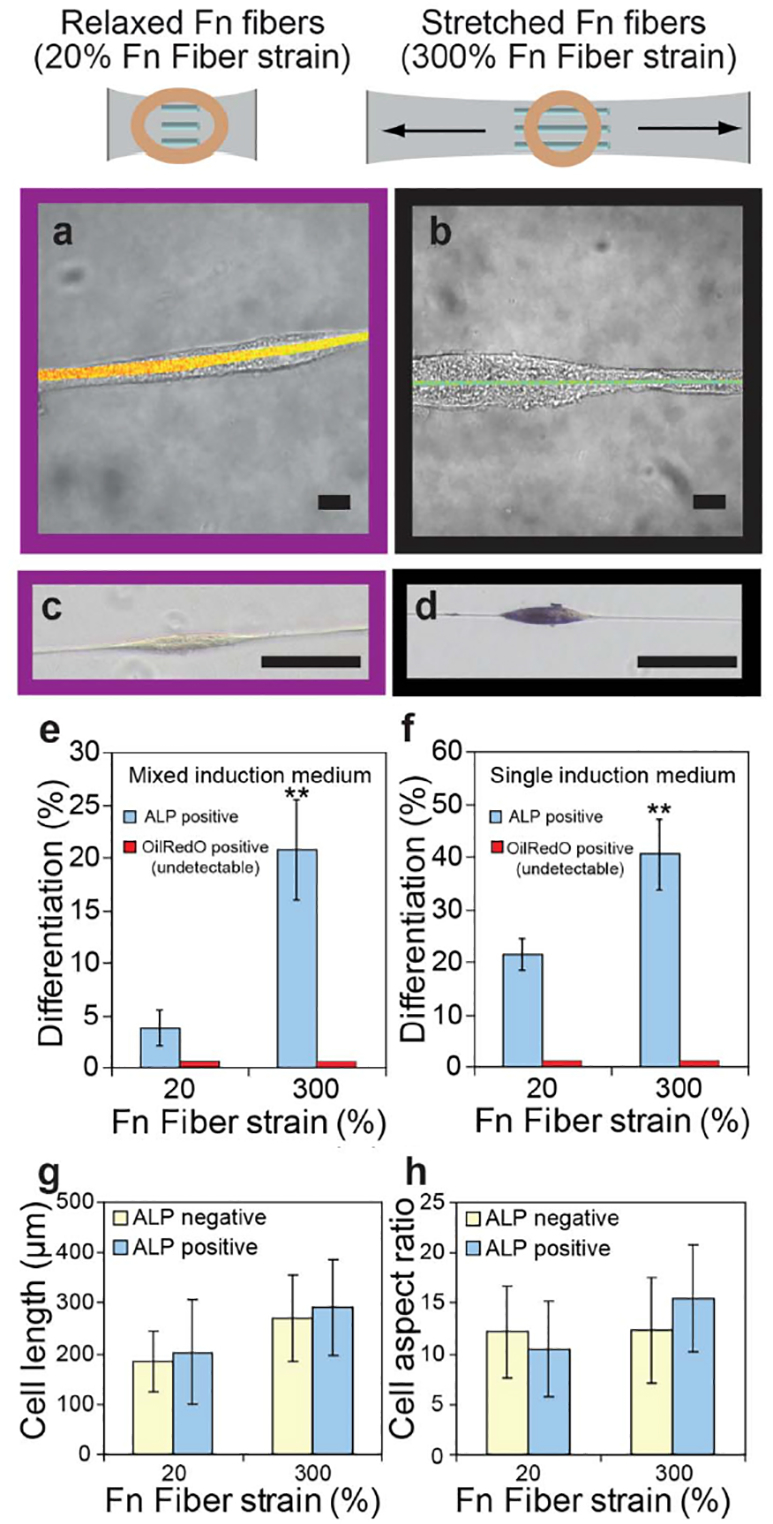
Osteogenic differentiation of hMSCs is upregulated on stretched single fibronectin (FN) fibres and regulates differential integrin and EGFR signalling. a Brightfield images of hMSCs merged with FN-FRET images of relaxed a or stretched b single FN fibres after cell attachment and culture for 2 days (scale bars: 10 mm). c and d Brightfield images of hMSCs cultured on cross-linked, relaxed (c, 20% fibre strain) or stretched (d, 300% fibre strain) single FN fibres for 7 days, with histochemical staining for ALP (blue). Scale bars: 100 mm. e and f Percentage of OilRedO and ALP positive hMSCs when cultured on single FN fibres in mixed e or single f induction medium for 7 days. Two asterisks: p<0·01 versus 20% Fn fibre strain. g and h Analysis of cell length g and aspect ratio h of ALP positive (blue) and ALP negative (yellow) hMSCs on relaxed and stretched single FN fibres. b hMSCs cultured on single FN fibres in osteogenic induction medium for 7 days with or without constant exposure to function-blocking antibodies. Percentage of ALP positive hMSCs is shown in the presence of function-blocking antibodies against integrin α5β1 (red), integrin α ν β3 (blue) or without antibodies (yellow). b Percentage of ALP positive hMSCs when cultured for 7 days on single FN fibres in the presence (green) or absence (yellow) of the EGFR inhibitor GW572016 in osteogenic induction medium is shown. Reproduced with permission from Ref. 140
Conclusion
Fibronectin matrices produced via cell-free routes represent promising routes to engineer novel biomaterials that direct cell response and promote tissue regeneration, being endowed with a distinctive biological activity that derives from their similarity with physiological FN fibrils. In particular, the FN network formed upon passive adsorption onto specific surface chemistries was shown to enhance cell adhesion, matrix secretion and degradation, but also to affect higher order cellular functions, such as cell differentiation. The improved biological properties of this artificial FN network confirm the similarity of this cell-free fibrillogenesis process and of the resulting network structure with the physiological FN fibrils, paving the way for its exploitation to control and direct cell behaviour in more complex systems that would include other ECM components mimicking natural tissue and cell microenvironments.
Acknowledgement
The support from the European Research Council (ERC) through HealInSynergy 306990 is acknowledged.