
Abstract
Using iron and sulphur oxidising bacteria to catalyse the breakdown of sulphides that host the gold is an important biological method for the pretreatment of refractory gold ores. Following this biological treatment, a combination of chemical and physical methods is used for leaching (such as the cyanide process) and concentration (such as carbon in pulp or electrowinning) of the gold. Although these methods are well accepted by industry, they harbour limitations in the processing of low grade refractory ores and regulatory agency/public acceptance of cyanide use. Thus, it is beneficial for industry to develop environmentally friendly, as well as cost efficient, leaching and concentration techniques that are based on micro-organisms. This may soon be possible by adapting the results of recent multidisciplinary research, which has shown that micro-organisms are capable of driving a biogeochemical cycle of gold dispersion, transport and re-concentration in the supergene environment. The indigenous microbiota in biologically active soil microcosms from a number of Australian sites are capable of solubilising up to 80 wt-% of the gold contained in soils and deeper regolith materials. Studies using molecular microbial techniques have shown that a metallophilic bacterium, Cupriavidus metallidurans, is present in biofilms on gold grains from a number of Australian sites. Cupriavidus metallidurans is capable of actively accumulating gold from solution and therefore contributes to the formation of secondary gold grains and nuggets. Identifying the biochemical and physiological pathways that lead to the dispersion and accumulation of gold in regolith and quantifying the reaction kinetics of these processes may thus lead to the development of industrial bio-processing capacities for gold containing ores.
Introduction
Increasing demand for gold puts pressure on mineral exploration to locate more economic deposits and on processing to extract the metal more efficiently. Currently bullion is produced from different types and grades of ore through a number of mainly physical and chemical steps. High grade ore is pulverised and treated depending on its metallurgical characteristics. Typically, oxide ore is sent directly to a leaching circuit. Refractory ore containing carbon may be roasted at 500°C to burn off sulphide and carbon resulting in an oxide ore. Sulphidic refractory ore without carbon can be oxidised by autoclaving to liberate the gold from sulphide minerals and sent to a circuit where gold is leached out using cyanide. Gold is frequently extracted from very low grade ore by heap and dump leaching in which cyanide solution is percolated through the ore. Gold is adsorbed from the cyanide solution using activated carbon which is then chemically stripped of gold. In a final step, gold is precipitated electrolytically or by chemical substitution. These steps enable both the purification and concentration of the gold.
In the current climate of environmental legislation and with average gold mining and extraction operating costs of approximately US$400–800/ounce and a gold price of over US$1500/ounce, these procedures are economically viable. However, they harbour a number of potential environmental problems, mainly through the use of cyanide and the high consumption of energy and water, which in the perspective of recent political debate on climate change and development of sustainable industries may persuade governments to enact stricter legislation in the future. This development will increase production costs for mining companies, which will have to bear the costs of bioremediation as well as carbon and water trading schemes, and therefore will put especially low grade ore processing under increasing economic pressure. The focus of many companies is now to develop processing technologies that require little maintenance and are economically as well as ecologically sustainable. This may not be achieved by increasing energy and water efficiency of existing techniques alone but may involve the development of entirely new ways of ore processing. One promising approach is to use environmental catalysts found in natural systems that are known to mediate the dispersion and re-concentration of gold (Fig. 1). The main catalysts involved in geochemical cycling of elements in the environment are micro-organisms (Madigan and Martinko, 2006). Therefore, in the following sections, the role of micro-organisms in the geochemical cycling of elements, and in particular gold, is assessed and the potential for a microbial ore processing plant is evaluated in the light of recent advances in gold geomicrobiology.
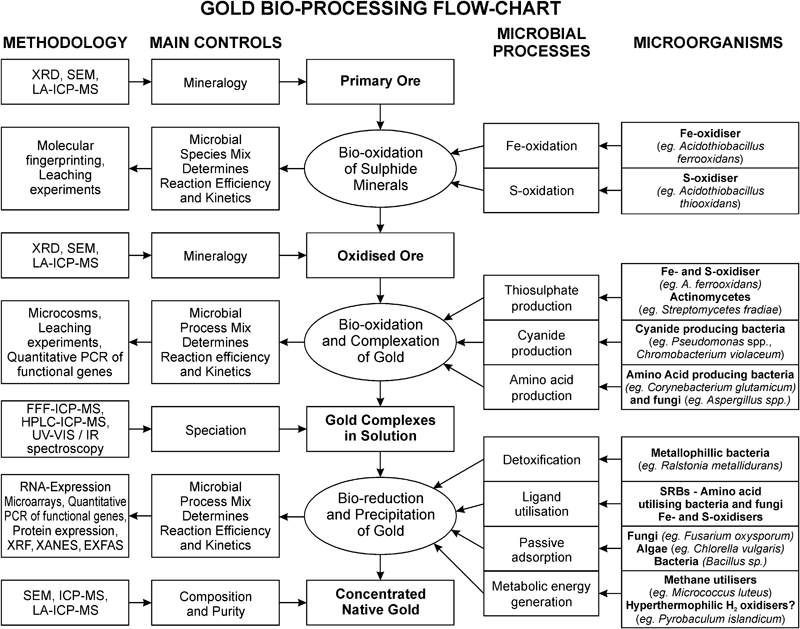
Flow chart linking microbial processes and populations central to the bio-processing of gold with research tools available to study these mechanisms and their influence on the ore. Abbreviations: XRD, X-ray diffraction; SEM, scanning electron microscopy; LA-ICP-MS, laser ablation inductively coupled plasma mass spectrometry; PCR, polymerase chain reaction; FFF-ICP-MS, flow field-flow fractionation inductively coupled plasma mass spectrometry; HPLC-ICP-MS, high performance liquid chromatography inductively coupled plasma mass spectrometry; UV-VIS, ultraviolet visible spectroscopy; IR, infrared spectroscopy; XRF, X-ray fluorescence; XANES, X-ray absorption near edge structure; EXFAS, extended X-ray absorption fine structure
The role of micro-organisms in geochemical cycling of elements
‘Micro-organism’ is a collective term for a large number of genetically, metabolically and ecologically diverse organisms, which have one characteristic in common: they are small, i.e. between several 100 nm to several tens of micrometres, mostly single-celled organisms. Microbes are the oldest and genetically most diverse forms of life on the planet (Ehrlich, 1998, p. 46–47). Fossil records demonstrate that the oldest group of micro-organisms, the single-celled prokaryotes (bacteria and archaea) inhabited the planet as early as 3·8 billion years ago and that these early ancestors of today's prokaryotes display cellular structures similar to their modern descendents (Ehrlich, 1998, p. 47). Other groups of micro-organisms, which are important in environmental systems are fungi and algae which, like ourselves, belong to the eukaryotes and whose earliest ancestors have been around for approximately 2·1 billion years (Han and Runnegar, 1992). Micro-organisms are active in all environments from up to several kilometres high in the atmosphere down to several kilometres depth below the Earth's surface in the lithosphere and the deep hydrosphere (Fyfe, 1996; Madigan and Martinko, 2006). They also occur in ‘extreme’ environments such as deep marine sediments (Parkes et al., 1994), deep sea hydrothermal vents (Juniper and Tebo, 1995), deep rock fractures (Pedersen, 1993), deserts (Adams et al., 1992), polar regions (Vincent and James, 1996) and acidic (pH<1), heavy metal polluted mine wastes (Ledin and Pedersen, 1996; Baker et al., 2004).
Besides being genetically and ecologically extremely versatile, micro-organisms, and in particular bacteria, have developed a wide spectrum of metabolic capabilities, including the ability to utilise inorganic elements in the production of energy (Nealson and Stahl, 1997; Madigan and Martinko, 2006). Elements that bacteria are known to oxidise or reduce in order to gain metabolic energy include H, C, P, S, V, Mn, Fe, Co, Cu, As, Se, Br, Mo, Sn, Sb, Te, Hg, W and U (Woolfolk and Whiteley, 1962; Nealson and Stahl, 1997; Ehrlich, 1998). A number of other elements are directly solubilised or precipitated by micro-organisms, and among these are many heavy and precious metals, e.g. Pb, Cd, Ag, Pt and Au (Southam and Saunders, 2005; Madigan and Martinko, 2006; Reith et al., 2007; Southam et al., 2009).
The biogeochemical cycle of gold in the environment – results from recent Australian studies
Micro-organisms play a key in role in the biogeochemical cycling of major and trace elements (Ehrlich, 1996, 2002). Recent research has produced evidence for a role of micro-organisms in the biogeochemical cycling of gold. Microbiota resident in auriferous Australian soils and other regolith materials are capable of mediating a geomicrobiological cycle of gold leading to the formation of secondary gold as grains and anomalies in calcrete (Fairbrother et al., 2009; Reith et al., 2005, 2006, 2007, 2009a, b, 2010, 2011, 2012a; Reith and McPhail, 2006, 2007; Schmidt Mumm and Reith, 2007; Southam et al., 2009). Secondary gold, which is formed as a result of the biogeochemical solubilisation and re-precipitation of gold under surface conditions, is widespread in surface environments (Reith et al., 2006, 2010, 2012b; Fairbrother et al., 2012). It is highly pure (>99 wt-% Au), finely crystalline (0·01 to <5 μm), and occurs in a variety of morphotypes, including nano- and micro-particles, triangular-plate (pseudo-trigonal octahedral) Au, sheet-like Au, secondary Au grains as well as bacteriomorphic, spheroidal Au commonly observed on the surface of transforming primary gold grains (Reith et al., 2006, 2007, 2010; Falconer and Craw, 2009; Hough et al., 2011). In contrast, primary gold is formed in high temperature, e.g. hydrothermal, systems, and commonly occurs as Ag-rich alloy. Primary grains often consist of large (10 to >100 μm) twinned crystals (Hough et al., 2007).
The indigenous microbiota in biologically active soil microcosms are capable of solubilising up to 80 wt-% of gold contained in such soils during the first 50 days of incubation, after which solubilised gold is re-adsorbed by mineral and organic soil fractions (Fig. 2; Reith and McPhail, 2006, 2007). In contrast, no gold was solubilised in sterilised microcosms incubated under otherwise identical conditions (Fig. 2). Molecular profiling allowed the differentiation of bacterial communities from auriferous and adjacent non-auriferous soils at the Tomakin Park Gold Mine in south eastern New South Wales, Australia (Reith and Rogers, 2008; Reith et al., 2012a). These results in combination with microcosms amended with dissolved
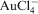 suggest that the presence of highly mobile gold in the regolith, as observed at many Australian sites, may influence the composition of resident microbiota (Reith and Rogers, 2008; Reith et al., 2012a, Usher et al., 2009).
suggest that the presence of highly mobile gold in the regolith, as observed at many Australian sites, may influence the composition of resident microbiota (Reith and Rogers, 2008; Reith et al., 2012a, Usher et al., 2009).
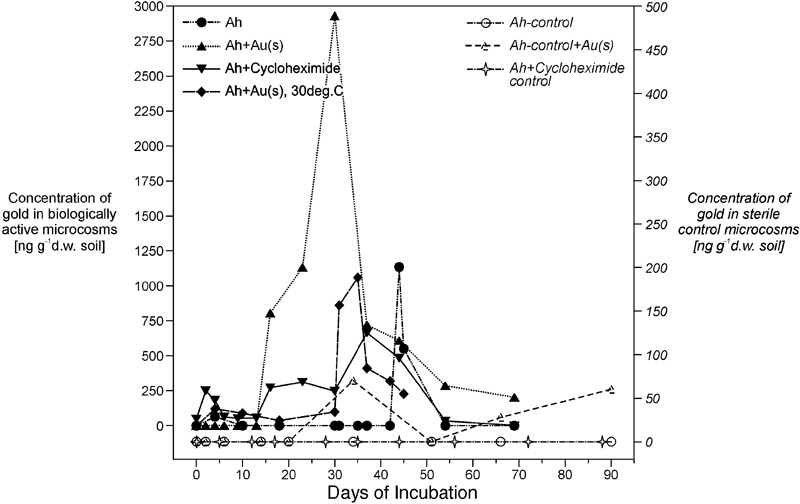
Concentration of solubilised gold in solutions from microcosms, incubated, biologically active or inactive (sterilised) with samples from the Ah horizon at the Tomakin Park gold mine, New South Wales, Australia
Scanning electron microscopy revealed bacterial pseudomorphs on untreated gold grains from the Hit or Miss Gold Mine, Queensland, Australia (Fig. 3a–c). Similar morphologies were also observed on grains from the Prophet Gold Mine, Queensland, and the Tomakin Park Gold Mine, New South Wales, Australia (Reith et al., 2006, 2010). The presence of active bacterial biofilms on the surface of gold grains was confirmed using confocal stereo laser microscopy combined with nucleic acid staining (Reith et al., 2006). Molecular profiling showed that unique, site specific bacterial communities are associated with these gold grains, which differed from those dominating the surrounding soils (Reith et al., 2006, 2010). 16S ribosomal DNA clones belonging to the genus Cupriavidus and bearing 99% similarity to Cupriavidus metallidurans were present on all 16S rDNA positive gold grains from both locations (Reith et al., 2006, 2010). The ability of C. metallidurans to actively accumulate gold from solution was successfully tested, suggesting that C. metallidurans may contribute to the formation of secondary gold in the Earth surface environments (Fig. 3d; Reith et al., 2006, 2009a).
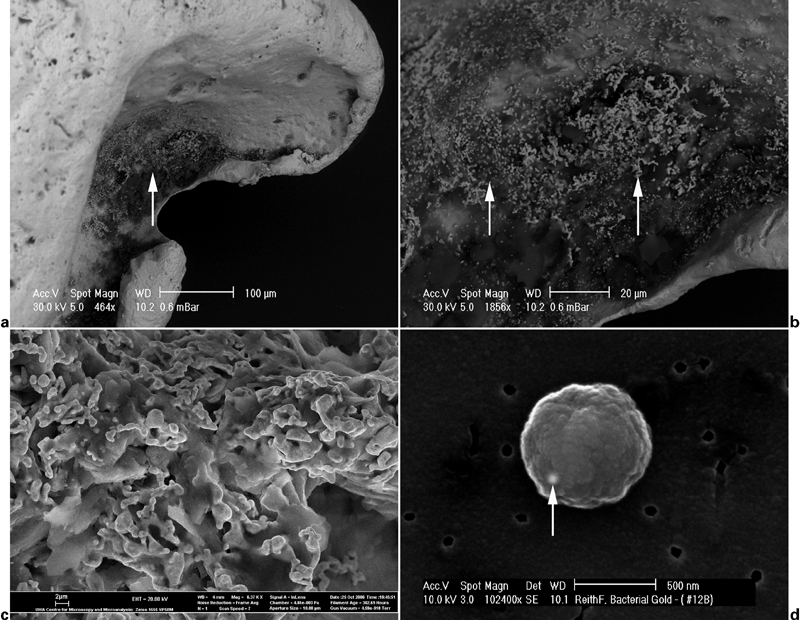
Secondary electron micrographs of surface features of a gold grain from the Hit or Miss Gold Mine in north eastern Queensland, Australia (a–c): a area of secondary ‘bacteriomorphic’ gold on the surface of a gold grain (arrow); b detailed view of branching network of bacteriomorphic gold associated with biofilm-like structures on the surface of a gold grain (arrows); c network of secondary gold sheets made up of budded conglomerates; and d secondary electron micrograph of metallic gold nano-particle (arrow) biomineralised by a spherical C. metallidurans cell
Mimicking the geomicrobial cycling of gold in an integrated biological processing plant
In order to develop microbial processing techniques, microbial processes and populations affecting gold mobility in environmental systems need to be studied (Fig. 1). Fundamental understanding of microbial gold solubilisation and precipitation processes and identification of micro-organisms that facilitate these processes has the potential to provide alternative biologically based processing techniques that have less negative environmental impacts and process management/waste disposal issues compared to current cyanide based processing. To obtain this detailed knowledge, the following key questions will have to be addressed:
Which micro-organisms are responsible for solubilisation and precipitation of gold?
Are these processes catalysed by microbial consortia or individual species?
Under which environmental conditions do these reactions occur, e.g. temperature, pH and redox, nutrient availability, etc.?
What are the biochemical and physiological mechanisms involved?
What are the kinetics of solubilisation and precipitation reactions?
How does mobile gold exist in environmental systems, as organic or inorganic complexes and/or nano-particles?
Once the mechanisms are understood in environmental systems, further questions regarding the viability of using these organisms in technical applications need to be addressed, among them are:
How selective are microbial solubilisation and precipitation mechanisms with respect to gold?
How do the organisms deal with high concentrations of other mobile heavy metals commonly found in processing solutions?
How are the organisms maintained?
How much does this way of ore processing cost compared to traditional processing techniques?
What risks are involved in the use of these microbes, e.g. pathogenic characteristics or toxic byproducts?
Once these questions have been satisfactorily answered in laboratory and pilot studies, full scale viability studies can begin at mining and processing sites.
Bio-oxidation of sulphides – an established bio-hydrometallurgical method
Microbial processes are already used in large scale processing operations for the bio-oxidation for gold bearing sulphide minerals (Olson et al., 2003).The bio-oxidation pretreatment process with the longest history of operation is Goldfields’ BIOX plant at the Fairview Mine in South Africa, which has operated since 1986 and treats refractory arsenopyrite/pyrite gold bearing concentrate in large, stirred tank, aerated, continuous flow reactors (Olson et al., 2003). A number of Australian operations (e.g. Beaconsfield, Fosterville, Harbour Lights, Wiluna and Younami) have adopted similar techniques and major international gold companies, such as Barrick Gold Corp. and Newmont Mining Corp., use bio-oxidation in their American heap leaching operations (Olson et al., 2003). In these operations, micro-organisms oxidise refractory ores containing pyrite, arsenian pyrite or arsenopyrite to expose gold occluded within the sulphide minerals matrix, which is then leached by conventional methods (Olson et al., 2003). Micro-organisms commonly used are iron and sulphur oxidising bacteria and archaea, e.g. mesophillic strains such as Acidothiobacillus spp. and Leptospirrillum spp., moderately thermophilic strains such as Sulfobacillus spp. and thermophilic strains of archaea, such as Acidianus spp. and Metallospheara spp. (Bosecker, 1997; Nordstrom and Southam, 1997; Olson et al., 2003; Reith et al., 2007) Using bio-oxidation gold recovery of up to 99% has been documented in association with the use of the thermophilic micro-organisms (Olson et al., 2003).
In the literature, two bio-oxidation mechanisms are discussed, i.e. ‘direct’ and ‘indirect’ bio-oxidation (Ehrlich, 1996; Bosecker, 1997; Sand et al., 2001). ‘Direct’ bio-oxidation assumes that the bacterial cells attached to the mineral directly oxidise the sulphide via its membrane bound enzyme system. However, recent research has shown that no direct pathway exists and that the bio-oxidation of sulphide minerals follows the ‘indirect’ pathway (Sand et al., 2001). In the ‘indirect’ pathway, the oxidative force of Fe(III) ions is used to oxidise sulphides according to the following overall reactions which describe the oxidation of pyrite (Sand et al., 2001):
Microbial oxidation of Fe(II) to Fe(III)
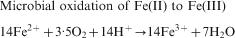
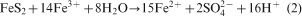
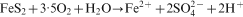
Although the overall reactions (shown above) were soon discovered, the exact microbial processes have only recently been described (Sand et al., 2001; Rawlings, 2002). Iron and sulphur oxidising bacteria have a strong affinity for mineral surfaces like pyrite to which they rapidly attach by forming biofilms. A layer of extracellular polymeric substances that serves as a reaction space is produced by these bacteria, when they attach to a mineral and form biofilms (Banfield and Hamers, 1997; Little et al., 1997; Sand et al., 2001). Biofilms on pyrite contain high concentrations of protons and dissolved and chelated Fe species which are derived from the bacterial oxidation from ferrous to ferric iron (Sand et al., 2001; Rawlings, 2002). The high concentration of Fe3+ and protons mounts the attack on the valence bonds of the pyrite, which is then degraded via the main intermediate thiosulphate (Sand et al., 2001). Fe3+ is reduced to Fe2+ in the process and then re-oxidised to Fe3+ by the iron oxidising bacteria. The general functions of iron and sulphur oxidising bacteria and archaea in the solubilisation of metal sulphides are to: provide sulphuric acid for a proton hydrolysis attack; provide the reactive environment for the chemical reactions to take place; and keep Fe or other metals in the oxidised state.
Bio-solubilisation of gold – the next step towards a microbial processing plant
After bio-oxidation gold contained in the pre-treated ore is more accessible to cyanide leaching. In this process, finely ground high grade ore is mixed with cyanide solution in concentration of about 2 kg t−1 of ore; low grade ores are stacked into heaps and sprayed with cyanide solution (up to 1 kg of NaCN per tonne). However, the use of cyanide is not only one of the largest cost factors for running a processing plant but is also fraught with a range of potential environmental problems. These problems include potential surface and groundwater contamination leading to cyanide and heavy metal contamination in catchments which may in turn lead to serious consequences for the surrounding ecosystems. This can result in the loss of potable water and death and extinction of endemic insect, fish and bird species. The contamination of surface and groundwater resources may lead also to socio-economic problems for the towns and settlements along the affected rivers due to loss of livelihood. Thus, it is economically as well as ecologically beneficial for companies, communities and the surrounding ecosystems, if the use of cyanide could in the medium to long term be phased out and replaced by cleaner leaching techniques.
A number of techniques have been assessed for their ability to use on ores that are not accessible to cyanidation. The most promising of these is leaching of gold with thiosulphate in the presence of co-ligands, such as ammonia and oxidants such as Cu(II) (Aylemore and Muir, 2001a, b; Wan and LeVier, 2003). The leaching reaction is described by this equation
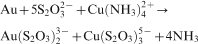
Thiosulphate and ammonium are also produced and excreted by common bacteria and actinomycetes during a number of metabolic reactions (Fig. 1; Sand et al., 2001). Ammonium is commonly produced through the hydrolysis of urea which is mediated by a wide range of yeasts and bacteria among the latter are many alkaliphillic Bacillus spp. (Castanier et al., 1999; Schmidt Mumm and Reith, 2007). Thiosulphate is produced by the Streptomycetes fradiae as a result of the metabolisation of sulphur from the amino acid cystine (Fig. 1; Kunert and Stransky, 1988). In other experiments, the sulphate reducing bacterias have been shown to form thiosulphate during the reduction of sulphite with H2 and formate (Fitz and Cypionka, 1990). A number of sulphate oxidising bacteria such as A. thioparus and A. ferrooxidans have also been shown to produce thiosulphate during the oxidation of sulphide (Fig. 1; Nordstrom and Southam, 1997). Thus, the solubilisation of gold observed in a recent study with gold quartz vein materials containing sub-microscopic gold in arsenopyrite and pyrite may have been mediated by thiosulphate leaching following sulphide degradation. In biologically active (with live microbiota) microcosm experiments, a maximum of 550 ng g−1 of gold (d.w. quartz vein material) were solubilised after 35 days of incubation (Reith and McPhail, 2006). In contrast, in sterile control microcosms, concentrations of solubilised gold were 10 times lower and lay in the range of theoretical solubility for the system (McPhail et al., 2006; Reith and McPhail, 2006).
In the same study biologically active microcosms with organic matter rich auriferous soils displayed up to 80% gold solubilisation in contrast to sterile controls where no gold was solubilised (Fig. 2). DNA fingerprinting and analyses of bacterial community function during the incubation of the microcosms combined with amino acids analyses indicated that changes in the structure of the bacterial community from carbohydrate- to amino acid-utilising populations occurred concurrently with and appeared to be linked to the observed solubilisation and re-precipitation of gold (Reith and McPhail, 2006, 2007). These results suggest the following model of gold solubilisation and re-precipitation. The bacterial community in the early stages of incubation apparently produced surplus amino acids which are known to directly solubilise native gold and stabilise it in solution in the presence of elemental oxygen, suggesting that the rapid microbial turnover of free amino acids in soils may contribute to the solubilisation of gold (Reith and McPhail, 2007). Some amino acids are also precursors for the microbial production of other gold complexing ligands, e.g. cystine a precursor for thiosulphate and glycine has been shown to be converted to cyanide by a number of bacteria, such as Chromobacterium violaceum and a number Pseudomonas spp. (Kunert and Stransky, 1988; Faramarzi et al., 2004; Faramarzi and Brandl, 2006; Fairbrother et al., 2009). In a number of studies, these organisms have been shown to solubilise gold from a range of materials such as gold containing ore and electronics waste (Faramarzi et al., 2004; Faramarzi and Brandl, 2006; Fairbrother et al., 2009). A patent already has been granted for the bio-hydrometallurgical processing of gold containing ores using C. violaceum. Cyanide is produced by the organism at a slow rate which makes it suitable for heap leaching (Krebs et al., 1997).
With a number of micro-organisms and microbial processes that lead to the solubilisation of gold already known research into using these individually or combined in a bioleaching is clearly important. Should these processes be suitable for technical application, economic ways of sourcing nutrients for maintaining the microbiota have to be found. These may include waste from food processing and water recycling plants that are often high in proteins and urea.
Gold accumulation by microbiota – promising results in bio-accumulation of gold
A number of comprehensive studies have been conducted into the accumulation of gold by micro-organisms (summarized in Reith et al., 2007; Southam et al., 2009). These studies suggest that selective microbial processes for gold bio-accumulation exist and that further research into the physiology, biochemistry and genetics of these processes needs to be undertaken to assess their value for industrial bio-accumulation of gold.
In early studies with Bacillus subtilis cell walls, Beveridge and Murray (1976) showed that the reduction and precipitation of gold from
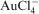 solution was selective because other metals, such as Ag+, were not reduced. In solutions containing
solution was selective because other metals, such as Ag+, were not reduced. In solutions containing
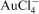 , Cu2+, Fe2+ and Zn2+ gold was selectively adsorbed by Bacillus subtilis and the cyanobacterium Spirulina platensis (Gee and Dudeney, 1988). The accumulation of complexed and colloidal gold was further investigated by Karamushka et al. (1987a, b) and Ulberg et al. (1992). They found that the accumulation of gold by a strain of Bacillus cereus was dependent on the chemical structure of the cell envelopes and involved functional groups of proteins and carbohydrates. In addition, they discovered that the accumulation of gold was directly dependent on the metabolic activity of the cells and especially on metabolic reactions on the plasma membrane, in particular the hydrolysis of ATP (adenosine triphosphate) by the enzyme ATPase (Karamushka et al., 1990a, b; Ulberg et al., 1992).
, Cu2+, Fe2+ and Zn2+ gold was selectively adsorbed by Bacillus subtilis and the cyanobacterium Spirulina platensis (Gee and Dudeney, 1988). The accumulation of complexed and colloidal gold was further investigated by Karamushka et al. (1987a, b) and Ulberg et al. (1992). They found that the accumulation of gold by a strain of Bacillus cereus was dependent on the chemical structure of the cell envelopes and involved functional groups of proteins and carbohydrates. In addition, they discovered that the accumulation of gold was directly dependent on the metabolic activity of the cells and especially on metabolic reactions on the plasma membrane, in particular the hydrolysis of ATP (adenosine triphosphate) by the enzyme ATPase (Karamushka et al., 1990a, b; Ulberg et al., 1992).
Energy dependent uptake of gold has also been shown for other bacteria such as Spirulina platensis (Savvaidis et al., 1998). Addition of
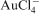 solution to cell suspensions of Geobacter metallireducens oxidised c-type cytochromes (Kashefi et al., 2001), which are thought to be involved in electron transfer to metals (Lovely and Phillips, 1988). Further experiments conducted with mesothermophilic and hyperthermophilic Fe(III)-reducing bacteria and archaea demonstrated that some of these organisms, such as Pyrobaculum islandicum, Shewanella algae, Desulfuromonas vulgaris and Geovibrio ferrireducens, are capable of precipitating gold by reducing Au(III) complexes to Au(0) with hydrogen as electron donor (Kashefi et al., 2001).
solution to cell suspensions of Geobacter metallireducens oxidised c-type cytochromes (Kashefi et al., 2001), which are thought to be involved in electron transfer to metals (Lovely and Phillips, 1988). Further experiments conducted with mesothermophilic and hyperthermophilic Fe(III)-reducing bacteria and archaea demonstrated that some of these organisms, such as Pyrobaculum islandicum, Shewanella algae, Desulfuromonas vulgaris and Geovibrio ferrireducens, are capable of precipitating gold by reducing Au(III) complexes to Au(0) with hydrogen as electron donor (Kashefi et al., 2001).
The bacterium Hyphomnas adhaerens has been shown to function as a hyper-accumulator for gold (Quintero et al., 2001). H. adhaerens was able to bind colloidal gold to the polar polysaccharide capsule of the prosthecate cell. However, a mutant without the capsule was not able to do so (Quintero et al., 2001). A further study of 30 different species of micro-organisms (8 bacteria, 9 actinomycetes, 8 fungi and 5 yeasts) has shown that bacteria were especially efficient in removing Au(I/III) complexes from solution (Nakajima, 2003). In another study, Keim and Farina (2005) reported the reductive precipitation and accumulation of
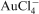 by uncultured magnetotactic cocci.
by uncultured magnetotactic cocci.
More recent research by Lengke et al. (2006a, b, c) focused on the precipitation of gold(III) chloride by cyanobacteria. They show that the reduction mechanism of Au(III) chloride to elemental gold by cyanobacteria involves the formation of an intermediate Au(I) species, i.e. Au(I) sulphide, with sulphur originating from the cysteine or methionine in cyanobacterial proteins. In another study, they assessed the precipitation of gold thiosulphate complexes
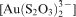 by a strain of the sulphur oxidising bacterium (Acidithiobacillus thiooxidans) isolated from deep subsurface gold mines (Lengke and Southam, 2005). They found that the
by a strain of the sulphur oxidising bacterium (Acidithiobacillus thiooxidans) isolated from deep subsurface gold mines (Lengke and Southam, 2005). They found that the
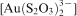 complex was stable in the bacterial systems until the oxidation of free thiosulphate was complete, after which the bacteria oxidised thiosulphate bound in
complex was stable in the bacterial systems until the oxidation of free thiosulphate was complete, after which the bacteria oxidised thiosulphate bound in
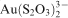 complexes, leading to the precipitation of native gold (Lengke and Southam, 2005). The gold was precipitated inside the bacterial cells as fine grained colloids ranging between 5 and 10 nm in diameter and in the bulk fluid phase as crystalline micrometre scale gold. Precipitation of gold was also observed in the presence of active, i.e. live, sulphate reducing bacterias due to the formation and release of hydrogen sulphide as an end product of metabolism (Lengke and Southam, 2006). The presence of iron sulphides, formed by biological processes, caused significant removal of gold from solutions by adsorption and reduction processes on the iron sulphide surfaces (Lengke and Southam, 2006). The presence of gold nano-particles within and immediately surrounding the bacterial cell envelope highlights the presence of localised reducing conditions produced by the bacterial electron transport chain via energy generating reactions within the cell.
complexes, leading to the precipitation of native gold (Lengke and Southam, 2005). The gold was precipitated inside the bacterial cells as fine grained colloids ranging between 5 and 10 nm in diameter and in the bulk fluid phase as crystalline micrometre scale gold. Precipitation of gold was also observed in the presence of active, i.e. live, sulphate reducing bacterias due to the formation and release of hydrogen sulphide as an end product of metabolism (Lengke and Southam, 2006). The presence of iron sulphides, formed by biological processes, caused significant removal of gold from solutions by adsorption and reduction processes on the iron sulphide surfaces (Lengke and Southam, 2006). The presence of gold nano-particles within and immediately surrounding the bacterial cell envelope highlights the presence of localised reducing conditions produced by the bacterial electron transport chain via energy generating reactions within the cell.
Reith et al. (2006, 2010) found that the bacterium, Cupriavidus metallidurans, is associated with secondary gold grains and has the ability to precipitate gold, suggesting that it may contribute to the formation of secondary gold grains in the environment via active precipitation of mobile gold complexes (Fig. 3). An explanation for the presence of active precipitation of gold(I/III) complexes in C. metallidurans and other bacteria may lie in the toxicity of these complexes to the organisms (Southam and Beveridge, 1996; Mossman et al., 1999; Karthikeyan and Beveridge, 2002). Even at low concentration Au(I/III) complexes can cleave peptide and protein disulphide bonds, disrupt cell walls and membranes and cause cell death (Witkiewicz and Shaw, 1981). Thus, for C. metallidurans reducing the toxic complexes and accumulating the native metal forms an effective mechanism to avoid gold toxicity and detoxify their immediate environment (Reith et al., 2009a, 2010).
C. metallidurans is able to withstand high concentrations of heavy metals such as Cu, Cd, Pb, Zn, Cd and Ag as well as gold better than any other well studied micro-organism. C. metallidurans is a gram negative facultative chemo-litho-autotrophic β-proteobacterium, which was first identified in 1976, when it was isolated from industrial sediments, soils and wastes that were polluted with high concentrations of various heavy metals (Mergeay et al., 2003). The extraordinary heavy metal resistance of C. metallidurans and its ability to accumulate these metals on its surface arises from multiple layers of efflux pumps (Nies, 2000; Mergeay et al., 2003). Nine hundred and thirty-two putative transporter proteins (13%) were recognised in its proteome. This is the highest percentage proteins involved in transfer of metal ions and organic molecules across the cell membrane found in any fully sequenced micro-organism (von Rozycki et al., 2005). This makes it an ideal model organism for understanding gold resistance and accumulation processes as well as a potential agent for bio-processing of gold.
A complementary multidisciplinary approach for assessing geomicrobial processes for use in gold processing plants
Understanding the geomicrobiology of gold in the environment and assessing its uses for processing applications requires expertise and experimental techniques from a wide range of disciplines. To evaluate the mechanisms and populations responsible for the environmental cycling of gold, a combination of techniques derived from classical and molecular microbiology, geochemistry, geochemical modelling, soil science, biochemistry, spectroscopy, hydrometallurgy and process engineering has to be applied (Fig. 1).
Microbial communities and physiological mechanisms functions associated with gold solubilisation and precipitation can be identified using DNA and RNA fingerprints obtained from gold grains, soils, deeper regolith materials and soil microcosms. Microcosms can be used to simulate in situ conditions in the laboratory and are used to establish the mechanisms and kinetics of the biologically mediated solubilisation of gold. Having established the kinetics for gold solubilisation and combining them with the precipitation kinetics measured in culture studies, they can be incorporated into a biogeochemical model for gold. Obtaining DNA and RNA fingerprints of microbial communities at the time of gold solubilisation will lead to the identification of key gold solubilising organisms which can then be cultured. Identifying gold complexes in soil solutions that were formed during the processes of microbial gold solubilisation may point to physiological mechanisms underlying microbially mediated gold solubilisation. Techniques such as high performance liquid chromatography coupled with an inductively coupled plasma mass spectrometry are highly sensitive techniques that can be used to determine the presence of gold complexes, such as gold amino acid and cyanide complexes in soil solution and bacterial growth media (Fig. 1; Lobinski, 1998). If gold solubilisation is linked to amino acid or cyanide biosynthesis, identification of pathways and functional genes important for production of these ligands is possible. Molecular probes for these genes can be constructed and their activity during gold solubilisation in soils using real time polymerase chain reactions can be assessed.
The fundamental physiological, genetic and biochemical mechanisms that C. metallidurans uses to precipitate gold can be assessed using techniques such as synchtroton spectrospcopy [i.e. micro-X-ray fluorescence (μXRF) and micro-X-ray absorption near edge spectroscopy (μXANES) mapping] and transcriptome micro-arrays (Reith et al., 2009a; Brugger et al., 2010). Synchrotron radiation X-ray adsorption spectroscopy allows a highly resolved measurement of the distribution and speciation of trace elements in natural materials. It also allows the measurement of trace element distribution within individual cells (Fig. 1; Kemner et al., 2004; Brugger et al., 2010). Micro-X-ray fluorescence and absorption spectroscopy (μ-XANES) was successfully used to assess the distribution and oxidation state of gold precipitated by C. metallidurans (Reith et al., 2009a). Using μ-XRF and μ-XANES has enabled us to measure the distribution and speciation of gold in individual bacterial cells. In addition, gold distribution and speciation around and within individual bacterial cells amended with different gold complexes, e.g. Au(III) chloride, Au(I) thiosulphate, Au(I) sulphide and Au(I) thiomalate), can be successfully measured.
In recent research with the cyanobacteria (Plectonema boryanum) mechanisms of gold bioaccumulation were studied at three gold concentrations (0·8, 1·7 and 7·6 mM final gold) from gold(III) chloride solutions using real time synchrotron radiation X-ray absorption spectroscopy (Lengke et al., 2006b). Interaction of cyanobacteria with aqueous gold(III) chloride initially promoted the precipitation of nano-particles of amorphous gold(I) sulphide at the cell walls and finally formed octahedral (111) gold platelets (∼10 nm to 6 μm) near cell surfaces and in two solutions. The X-ray absorption spectroscopy results confirm that the reduction mechanism of Au(III) chloride to elemental gold by cyanobacteria involves the formation of an intermediate Au(I) species, namely Au(I) sulphide.
Whole genome transcriptome micro-arrays are an exciting new tool used in molecular microbiology. Micro-arrays are gene chips on which several hundred thousand nucleic acid probes can be placed and used simultaneously (Widada et al., 2002; Zhou, 2003). This allows researchers to study the total genomic reaction of bacterial species, such as C metallidurans, towards changing environmental conditions, such as mobile gold in bacterial growth medium (Reith et al., 2009a). Using this technique, it can be shown which genes are up regulated (more active) and which are down regulated (less active) if mobile gold is present. This will allow a detailed insight into which fundamental mechanisms the organism uses to deal with gold toxicity.
Conclusion
The results of our recent studies and the work of other groups have shown that further research into the development of economically sustainable and environmentally friendly processing techniques for gold is warranted. The focus of this research has to lie in using novel techniques such as synchrotron spectroscopy and whole genome expression micro-arrays to establish the fundamental mechanisms of gold turnover in micro-organisms. A number of processes that warrant further investigations are: the production of thiosulphate by iron and sulphur oxidising bacteria during the breakdown of sulphide which may be used to directly solubilise gold during bio-oxidation; the biogenic production of free amino acids and cyanide for the solubilisation of gold; and the utilisation of C. metallidurans and selected sulphate reducing bacteria for the biogenic concentration of gold from the leaching solution.
Footnotes
Acknowledgements
The authors acknowledge the following individuals and institutions for their contributions: the Australian Research Council (LP100200102 to FR and JB); Flinders University, The University of Adelaide, Centre for Tectonics, Resources and Exploration (TRaX), CSIRO Land and Water, The South Australian Museum; Newmont Asia Pacific, Barrick Gold of Australia Limited; Adelaide Microscopy. This paper was originally presented at World Gold Conference, 22–24 October 2007, Cairns, QLD. It has been subsequently peer-reviewed and revised for publication in Mineral Processing and Extractive Metallurgy with the kind permission of the Australasian Institute of Mining and Metallurgy.
This paper is part of a special issue on Australian gold processing