
Abstract
The aim of this study was to explore the golden ratio (Φ) based scaling relations existing between the golden ring volume, the golden measure equation (GME) and transversal dynamics of growth ring formation. Magnitudes of the golden ring volume, the golden ring cube and parameters of the GME were experimentally quantified based on X-ray densitometer data of Douglas-fir and white spruce. Thereafter, transversal dynamics of the parameter-fitted equations were analysed. The Φ (1·618) was satisfied by both species with 95% confidence. The first order derivative of the GME agreed with the transversal momentum of growth ring formation. The second order derivative was deduced to have potential in explaining transversal physiological transitions from earlywood to latewood and juvenile to mature wood. The structure and formation of growth rings are shown to be self-optimised based on Φ and this finding may explain why wood has higher strength-to-weight ratio in comparison with other materials.
Keywords
Introduction
Wood has been a material of choice for construction, composite materials, furniture and specialised purposes such as resonance wood. Wood is preferred for construction, composite materials and furniture due to its high strength-to-weight ratio and better aesthetic value when compared with other alternative materials. High strength-to-weight ratio also indicates good resonance properties. Therefore, wood which has a relatively higher strength-to-weight ratio, hereafter called ‘golden wood’, is valued due to its multipurpose use for high value products. The quality of wood, which is intended for construction, furniture, composites or resonance purposes, increases with increasing magnitude of the strength-to-weight ratio. Golden wood can be assumed to have a relation to the golden ratio Φ because a strong relationship has been shown to exist between Φ and the high strength-to-weight ratio (Yadav et al. 2012). Φ is a constant obtained as a ratio of two components of a given whole of physical entity and has a magnitude of ∼1·618 or exactly ((1+51/2)/2) (Stakhov 2005).
Furthermore, trees which grew under hostile environment are believed to yield the golden wood (Schwarze et al. 2008). Harsh growth conditions cause trees to lay down narrow annual rings, a high modulus of elasticity and low density. It is for this reason that timber species of Picea which grew under similar conditions are prized for their golden wood (Green et al. 2010). Moreover, tree species such as bristle cone pines which are capable of adapting to such hostile environment, are observed to have the highest longevity (Currey 1965; Brunstein 1992; Swetnam and Brown 1992). Slow growth and formation of narrow growth rings is observed not only in trees which have been growing in harsh environmental conditions but also when they are long lived and old growth (Black and Colbert 2008; Johnson and Abrams 2009). Wood formed at an advanced tree age can be assumed to be similar to the golden wood since growth generally slows down as maturity phases in. Therefore, Φ may also be associated with tree maturity and longevity in addition to the mechanisms leading to formation of the golden wood. According to Stoel and Borman (2008), the golden wood may owe its noble attributes to the significantly more uniform structure of wood formed both in earlywood and latewood during slow growth. Tree ring width, which is often comprised of widths of earlywood and latewood, is the major parameter appearing in all cases and establishment of its relation to the golden wood and tree longevity could generally contribute to the understanding of dynamics of transversal wood formation. Establishment of a strong relation is particularly helpful to the understanding of mechanisms leading to formation of the golden wood. Since longevity is a result of complex mechanisms taking place in life cycles of trees, it is important to look into its relation to other biological variables.
An investigation by Lanner and Connor (2001) on whether bristle cone pines undergo senesce revealed no manifestation of the common signatures of aging, such as deterioration of meristematic function in embryos, seedlings or mature trees. Nevertheless, the life span of even these long living coniferous trees is finite with no record showing an ability to survive more than 5000 years (Currey 1965; Brunstein 1992; Swetnam and Brown 1992). In earlier works of Ryan and Yoder (1997), Bond et al. (2007) and recently by Enquist et al. (2008), growth limits are shown to have much to do with the size of individual trees. Douglas-fir trees can grow to more than 100 m in height and possess a magnificent straight stem withstanding its own heavy weight and external loads such as snow and transversal wind pressure (Alden 1997). More recently, Kempes et al. (2011) reported tree height to be the major characteristic size which controls physiological traits. Ally et al. (2010) also showed aging to be a factor controlling growth. Since size and age are usually mutually dependent and directly correlated, size is the common parameter controlling growth limit in all cases.
Width of growth rings is used as characteristic size to gauge the relative degree of growth rate and its narrowness is understood as a good indicator for slow growth and longevity. However, there are no sufficient quantitative explanations helping to directly correlate slow growth, longevity, golden wood and Φ. The two-level system (TLS) coarse graining framework developed by Tekleyohannes (2010) and Tekleyohannes and Avramidis (2011) can be used to establish a quantitative relation between slow growth, longevity and the golden wood. Powerful growth ring parameters were identified in the TLS coarse graining framework which were namely, golden ring volume (GRV), golden ring cube (GRC), β0 and dimensionality d. The GRV was defined in the TLS coarse graining framework as the wood substance contained in unit of volume described by width of earlywood, latewood and the whole growth ring (sum of the widths of earlywood and latewood). The GRC was defined as the wood substance contained in unit of cubic volume described by width of the whole growth ring. The parameter β0 was defined as an equivalent to reciprocal of the GRV in its magnitude. The dimensionality d may be considered as the mean number of potential configurations available to the GRV in given unit of GRC when it behaves as two-component system.
The Φ is shown in Tekleyohannes (2010) and Tekleyohannes and Avramidis (2010) to appear as the limit to the mutual dependence of two scaling exponents describing physical properties and chemical composition of wood as TLS. Water vapour sorption in wood was also shown to scale based on Φ in the vicinity of the transition from mono-layer to poly-molecular adsorption (Tekleyohannes 2010). If Φ appears in the scaling relation between various anatomical and characteristic sizes in the structure and properties of wood, then this may indicate the reason why golden wood has higher strength-to-weight ratio than other materials, such as steel (Yadav et al. 2012). The knowledge can be used to develop engineered wood products such as structural laminates and biocomposites having high strength-to-weight ratios without the inherent defects existing in native wood.
Therefore, the first objective of this study was to present analytical and empirical evidence for the existence of a scaling relation between transversal dynamics of growth rings formation, golden wood, longevity and characteristic sizes such as the GRV, GRC and parameters such as β0 through Φ. The second objective was to explore and gain experimental insight on the dynamics of transversal wood substance formation and growth limits which are useful in developing structurally optimised and efficient wood composites through biomimetics.
Materials and methods
Various scaling relations obtained from combinations of the GRV, GRC and β0, which were derived in Tekleyohannes (2010) and Tekleyohannes and Avramidis (2011) and Φ were tested and validated experimentally and analytically. The scaling relations were derived based on a fundamental assumption stating that a growth ring which is formed during slow growth or at advanced tree age possesses a uniform anatomical structure which is governed by Φ enabling it to become identical to its neighbouring growth ring. The experimental testing and validation were performed using X-ray densitometry data of two softwood species, namely, Douglas-fir (Pseudotsuga menziesii) and white spruce (Picea glauca) which were collected based on a standard method described in Jozsa et al. (1987). The data were gathered using breast height densitometer strips sampled from six second growth naturally regenerated forest sites of Douglas-fir. Basic density data for white spruce were also collected from X-ray densitometry strips sampled from three second growth forest sites in British Columbia. Details describing location, site index, age and stand density of sampling sites are shown in Table 1.
Location, site index, age and stand density of sampling sites for Douglas-fir and white spruce
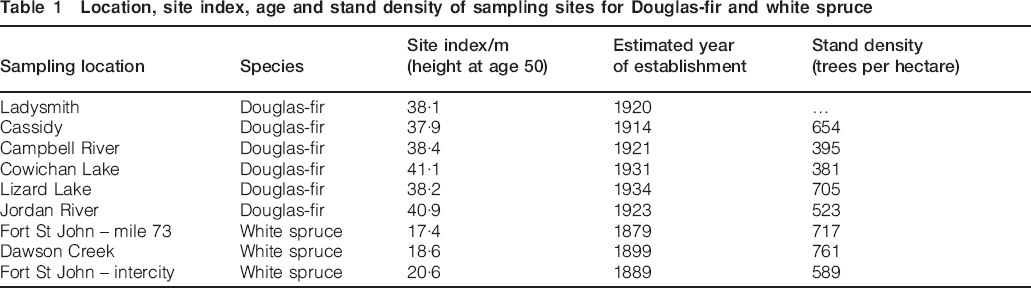
Ten breast height X-ray densitometer strips were obtained from each of the six sampling sites of Douglas-fir while 20 similar strips were obtained from each of the three white spruce sampling sites. Therefore, the total number of samples per species was 60 strips. These data were used in combination with the parameters obtained during validation tests of earlier models given in Tekleyohannes (2010) and Tekleyohannes and Avramidis (2011). This means that β0 values were taken to be 16 (a rounded figure from 16·11) and 32 (a rounded figure from 32·17) for Douglas-fir and white spruce respectively.
Experimental magnitudes of the GRV and GRC were established as given in Tekleyohannes (2010), and Tekleyohannes and Avramidis (2011). Quantification of the GRV, GRC and the dimensionality along with validation of equations (1) and (2) was carried out by separating ring series into early juvenile, late juvenile, mature heartwood and sapwood based on the new framework of classification for transversal wood types as given in Tekleyohannes (2010). The mean number of GRV (ng) in a corresponding unit of GRC was quantified based on equation (1) as the average obtained for each of the transversal sections and wood types
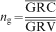
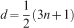
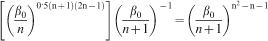
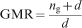
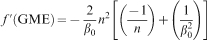
Results
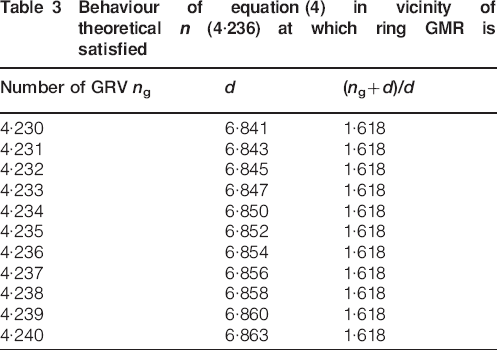
Summaries of results for the GMR obtained by separating ring series into early juvenile, late juvenile, mature heartwood and sapwood of Douglas-fir and white spruce are listed in Table 2. Both species displayed greater dispersion for juvenile wood, though white spruce was observed to have the highest one probably owing to its narrower latewood which is more difficult for instrumental measurement. Table 3 shows the behaviour of equation (4) in the vicinity of theoretical ng (4·24) at which the ring GMR is satisfied. It can be asserted from Tables 1 and 2 that equations (1) and (2) sufficiently describe ng (the number of mean GRV in a given unit of GRC) and the dimensionality d. The numerical magnitude of ng (4·24) can be deduced to be equivalent to [(1+51/2)/2]3 corresponds to which a dimensionality of 6·85 or equivalently [(1+51/2)/2]4. Computed magnitude of n is termed as ng to indicate its representation of the mean number of GRVs in a single unit of GRC.
Dimensionality of GRC to GRV ratio ng and corresponding GMR based on equation (10)*
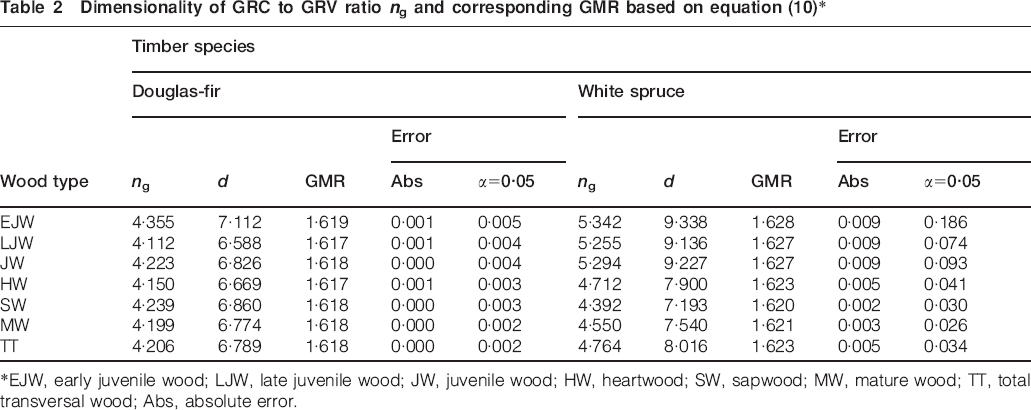
*EJW, early juvenile wood; LJW, late juvenile wood; JW, juvenile wood; HW, heartwood; SW, sapwood; MW, mature wood; TT, total transversal wood; Abs, absolute error.
Behaviour of equation (4) in vicinity of theoretical n (4·236) at which ring GMR is satisfied
The reciprocal of β0 is equivalent to the mean GRV (the mean wood substance content) which corresponds to a unit of mass, although it was non-dimensionalised. The variable n is a radial increment (displacement) of the wood substance in a unit of time. Therefore, the preterm given in equation (5) imparts a sense of motion from which readily follows the fact that the equation describes the momentum of wood substance formation. In this sense, the first order derivative may result in a physically meaningful equation.
Equation (5) becomes dependent only on n once β0 is determined. This dependence is demonstrated on plots given in Fig. 1 both for Douglas-fir and white spruce. The profiles are similar to inverted parabolas having maxima, and n-intercepts as the minimum positive value. The maximum value for Douglas-fir was at about n = 128 while it intercepted the n-axis at n = 256. On the other hand, the maximum value for white spruce was located at about n = 512 with an n-intercept at 1024.
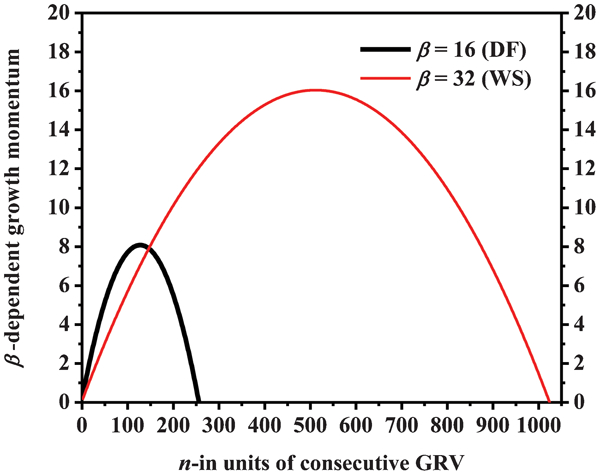
Plot of first order total derivative of equation (3) as given by equation (5) showing parabolic projectile of growth momentum for Douglas-fir (DF) and white spruce (WS)
Discussion
Douglas-fir displayed closer-to-theoretical magnitudes of experimental ng than white spruce as the results in Table 2 demonstrate. Furthermore, the data in Tables 2 and 3 support the assertion made by Tekleyohannes (2010) and Tekleyohannes and Avramidis (2011) in which Douglas-fir was recommended as the standard or baseline species of comparison during investigation of others since it had β0 corresponding to the theoretical unit of ng. This finding may justify why coastal Douglas-fir grows more in height than spruce species while both of them have comparatively close magnitudes of the strength-to-weight ratio as strength tests made on small clear specimens indicate (Green et al. 2010). Therefore, the taller straight stem of Douglas-fir trees, which is capable of withstanding its own heavy weight and external loads such as snow and transversal wind pressure, can be justified by its golden wood or higher degree of golden ratio based transversal self-optimisation in the structure of its growth rings.
Table 2 further shows that magnitudes of ng and d obtained for sapwood of both species were closer to the theoretical results than those which were obtained for early juvenile wood, late juvenile wood and heartwood. Hence, sapwood is more similar to golden wood than other transversal wood types. This finding supports the assumptions made during derivation of the equations which propounded growth ring formation at an advanced tree age to be governed by Φ since sapwood contains growth rings formed at older age. Equations (2) and (4) and Tables 2 and 3 additionally demonstrate existence of a power law relationship between Φ, d and ng. Consequently, if Φ is raised to the power of some natural numbers then it yields useful discrete quantities, such as ng and the dimensionality d of growth increments, which can be used to characterise given series of growth increments. The natural numbers observed for Douglas-fir and white spruce were 3 and 4 for ng and d respectively.
The rate of wood substance formation increases with increasing tree age as is evident from equation (5) and Fig. 1 which may be owing to the increase in the capacity to photosynthesise. However, the reciprocal of n in equation (5) works against an increase in the rate of wood substance formation, apparently by simulating aging. The elegance of equation (5) is consisted in β0 which is the only free parameter that must be known to quantify the first order derivative of GME. The parameter β0 has a unit corresponding to an energy quantity and it can be safely assumed to have equivalence to the thermodynamic temperature (Tekleyohannes 2010; Tekleyohannes and Avramidis 2011). The parameter β0 is also the reciprocal of the mean wood substance content in the GRV. Consequently, it is possible to establish a direct connection between the molecular level wood cell metabolism and the amount of mean annual wood substance formation in terms of energy requirements through the classical Boltzmann equation.
Width of sapwood may be regarded as a good indicator to the vigour in a living tree, but various studies did not provide conclusive evidence on the dependence of width of sapwood on tree age (Hazenberg and Yang 1991). Indeed, width of sapwood is usually assumed to vary depending on factors such as site, climate, elevation and tree vigour itself. Hence, there is a difficulty to obtain a clear dependence of the sapwood on tree age. However, Hazenberg and Yang (1991) and Nawrot et al. (2008) obtained a curvilinear relation on the dependence of the number of growth rings in sapwood on age. The observed dependence shows the same trend like the parabolas given in Fig. 1 in agreement with equation (5).
The magnitudes obtained as intercepts (solutions) and maxima based on equation (5) for Douglas-fir and white spruce agree with age of maturity and advanced age at which the rates of wood substance formation are maximal and minimal respectively (Szeicz and MacDonald 1996; Alden 1997; Nesom 2006; Johnson and Swanson 2009). This finding entails an interpretation for n-max to be the average year at which a given tree with parameter β0 enters age of maturity, while n-min indicates the average year beyond which growth inhibition can be expected. The dependence of maturity and advanced age on the square of β0 is evident from equation (5). Since β0 is the reciprocal of GRV, mature and advanced ages are inversely related to the square of the GRV. The bigger the β0, the higher the mature and advanced ages are, which directly translates to the smaller the GRV, the longer the mature and advanced ages will be. Consequently, the transversal growth momentum is expected to decelerate once the mean number of growth years passes a magnitude which is equivalent to half of the square of β0 causing formation of narrower growth rings, and finally becomes null if the mean number of growth years attains a magnitude of
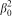 . Therefore, growth slows down in direct proportion to the square of β0 as tree enters the phase of advanced tree age which is an essential condition for properties of the final GRV to converge to that of the golden wood. The full physical and biological significance of the first order derivatives with regard to transversal formation of growth rings and the golden wood should be investigated and explained further using multi-species experimental data in the future.
. Therefore, growth slows down in direct proportion to the square of β0 as tree enters the phase of advanced tree age which is an essential condition for properties of the final GRV to converge to that of the golden wood. The full physical and biological significance of the first order derivatives with regard to transversal formation of growth rings and the golden wood should be investigated and explained further using multi-species experimental data in the future.
The second order derivative of the GME will be reported in a separate paper owing to the complexity and vastness of analytical issues associated with it. However, some of its interesting aspects could be apparent from the first order derivative. For instance, if the first order derivative of the GME yielded an equation agreeing with growth momentum, then its second order derivative can be deduced to yield a force, in physical analogy. The force may be likened to growth vigour or longevity. The second order derivative could also be expected to be governed by the single parameter β0 with more singularity points (in addition to zero) which will potentially indicate anatomical and physiological transitions like the one existing between earlywood and latewood, juvenile wood and mature wood and probably to terminal growth inhibition or death. These assertions shall be analytically and experimentally validated in a separate and detailed report.
Earlywood and latewood are anatomical building units of wood and this work demonstrated that their layering does not go on indefinitely. Transversal growth is finite; hence the number of layers of growth rings should be correspondingly finite. No tree is observed to grow infinitely in height but there are trees such as coastal Douglas-fir which are known for their tallness. The analytical and experimental evidence obtained in this study implied the reason for tall growth of Douglas-fir to be self-optimisation of its transversal growth ring formation based on Φ. Hence, defect free round timber having transversal structure of growth rings similar to Douglas-fir is naturally self-optimised based on Φ and capable of performing well, for instance as a column. Nevertheless, wood and wood products are used in segments and finite forms in the majority of structural and functional applications, such as for columns, sides and load bearing partitions. Generally, round timber is rarely used in its native form and it is commonly sawn into boards and remanufactured into less optimal structural elements which are prone to warping accompanied by a corresponding reduction in properties of the golden wood. However, the scaling relations tested in this study can be used to engineer the remanufactured structural members, so that it is structurally as efficient as the golden wood or defect free and self-optimised round timber.
Conclusion
The GME, its first order derivative and ng showed a capability to predict and explain the dynamics and mean magnitude of the transversal growth momentum in softwood timber species with conspicuous growth rings, particularly in Douglas-fir and white spruce. The GRV, its reciprocal β0 and GRC were the major controlling parameters in all tested equations. They are also shown to be characteristic sizes which may potentially control transversal dynamics of growth ring formation, formation of golden wood and longevity. The scaling relationship exhibited between the various characteristic sizes through Φ will help to develop high performance engineered wood composites with high strength-to-weight ratio. It is recommended to test and validate the findings of this study on more softwood and hardwood species with less conspicuous growth rings, and transfer the findings obtained for growth rings to microscopic, nanoscale and molecular level.
Footnotes
Acknowledgements
The authors would like to express their great appreciations to Dave Munro of FPInnovations-Forintek Division for providing densitometry data of Douglas-fir and white spruce. They would also like to greatly thank the anonymous reviewers whose valuable comments and feedbacks helped to make the report better. This project was funded by a Discovery Grant from the Natural Sciences and Engineering Research Council of Canada.