
Abstract
Acetylation appears suited to provide adequate protection against biological attack for materials derived from non-durable wood species. But still there are unanswered questions related to resistance against fungal decay. The paper summarises existing knowledge related to fungal deterioration of acetic anhydride modified wood and also highlights future research opportunities. In addition, statistical analyses based on previously published decay fungi studies were performed to quantify what factors contribute most to the performance (calculated as test sample/control). The results showed that weight per cent gain can explain approximately 50% of the performance for acetic anhydride treated wood. Others of the applied variables, like wood species or type of fungus, can reduce the variance in performance by additional 15%. Based on the surveyed literature the degree of cell wall bulking in combination with lowering of the equilibrium moisture content seems to be the primary mode of action.
Introduction
Issues of sustainability, carbon sequestration and performance converge in a continuous search for environmentally benign methods of wood protection. The science and technology of wood modification has been comprehensively described by Hill (2006). Most of the commercial developments of modified wood have taken place since 2006 and Hill (2011) gives an update on the recent commercial status. One of the commercialised wood modification processes is acetylation with anhydride. Commercial production of acetylated wood with acetic anhydride started with Titanwood in the Netherlands in 2007, and Eastman Chemical Company commercialised the product in the USA in February in 2012. Derived from non-durable wood species, sufficient treatment concentrations will be capable of achieving adequate Use Classes 3–5 (EN 335 2006) performance (Jones et al. 2006). Even if it is very well established that acetylation is able to impart a significant degree of decay resistance to wood (e.g. Hill et al. 2003; Hill 2006, 2009; Rowell et al. 2009; Papadopoulos 2010), more evidence regarding the mechanism by which acetylated wood is protected from fungal decay is needed for further optimisation of processes. The aim of this paper was to: summarise knowledge related to fungal decay resistance of acetic anhydride modified wood, based on published data use statistical tools to explain what factors contribute most to the performance and highlight future research opportunities.
Literature survey
The history of acetylation of wood is described by e.g. Rowell (1983), Hill (2006) and Rowell et al. (2009). In 1928 three different papers were published on acetylation of wood. Fuchs (1928) used an acetyl weight present gain (WPG) of over 40, decrystalised the cellulose in the process and used it to isolate lignin from pine wood. Horn (1928) used a similar process to remove hemicellulose from beech wood. Suida and Titsch (1928) acetylated powdered beech and pine to yield an acetyl weight gain of 30–35%. Tarkow was the first to demonstrate resistance to fungal decay using acetylated balsa (Tarkow 1945) and the use of acetylation to stabilise wood from swelling in water (Tarkow 1946). According to Hill (2006) the first recorded acetylation reaction of solid wood is attributed to Stamm and Tarkow (1947).
Lab trials
From the 1960s several research groups started testing different treatment levels and wood species in lab against decay fungi, both in monoculture tests and in unsterile soil tests (e.g. Stamm and Baechler 1960; Goldstein et al. 1961; Peterson and Thomas 1978; Kumar and Agarwal 1983; Takahashi et al. 1989; Larsson and Tillman 1989; Militz 1991; Chow et al. 1994, 1996; Beckers et al. 1994, 1995; Goethals and Stevens 1994; Forster et al. 1997, 1998; Suttie et al. 1998; Ohkoshi et al. 1999; Larsson–Brelid et al. 2000; Mohebby and Militz 2002, 2003; Mohebby 2003; Cetin and Ozmen 2005; Papadopoulos et al. 2010, 2011). Hill (2006) gives the following very general summary: ‘In general, most studies of acetylated wood have shown that a WPG of the order of 20% is required to protect the modified wood from attack by brown rot fungi. With attack by white rot fungi, many studies indicate that a WPG of less than 10% appears to be adequate to protect softwoods, whereas a WPG of the order of 20% is necessary to protect hardwoods.’
Field trials
From 1995 a series of field trial results were published (e.g. Beckers et al. 1995; Rowell et al. 1997; Larsson–Brelid et al. 1997, 2010; Edlund 2004; Papadopoulos 2006, 2009; Edlund and Jermer 2007; Alfredsen and Westin 2009; Mohebby and Militz 2010; Hedley et al. 2011). Still results from long term field trials are sparse, but Larsson-Brelid et al. (2010) found that the resistance to fungal decay, after 18 years in soil contact, of wood with acetyl content of about 20% was of the same magnitude as for CCA treated wood at a high retention level (10·3 kg m−3).
Chemical and anatomical changes during decay
Microscopic examination by Peterson and Thomas (1978) showed that hyphal counts were substantially lower in acetylated wood samples compared to control. Examination of regions where hyphae were in direct contact with the cell wall revealed that there was very little evidence of cell wall degradation. And where it occurred it was confined to the hyphal tips. Penetration of hyphal tips was completely prevented at WPGs in excess of 15%. They stated that acetylation appeared to provide a high level of carbohydrate protection against both white and brown rot fungi.
From the end of the 1990's papers started to look more into the details of chemical and anatomical changes during decay. Ohkoshi et al. (1999) characterised the decay of acetylated wood due to brown-rot and white-rot fungi by analysis of chemical composition, X-ray measurements, and C-13 NMR spectroscopy. The brown-rot fungus decomposed the cellulose in the crystalline region to a large degree when the mass loss was more than 40%, whereas the white-rot fungus decomposed the crystalline region and the noncrystalline regions in acetylated wood to the same degree. The brown-rot fungus preferentially decomposed unsubstituted xylose units in acetylated wood and partly decomposed the mono-substituted xylose units. It was suggested that the mono- and disubstituted cellulose was partly decomposed by brown-rot fungus. Rosenqvist (2000, 2002) used environmental scanning electron microscopy plus microautoradiography and found that introduced acetyl groups showed an even distribution in the wood cell wall at acetylation level of about 5, 15 and 20% (weight gain).
According to Mohebby (2003) fungi could easily penetrate into acetylated wood, preferably vessel lumina and rays at early stages, and then penetrate into fibre cell lumina through interfibre pits or cross fields between rays and fibres. Mohebby and Militz (2002) studied acetylated beech and Scots pine in laboratory soil beds at intervals up to 300 days. For beech changes in chemical composition were measured at lower WPGs while no losses were detected at the highest WPG. For Scots pine minor changes were measured at all WPGs; except in holocellulose. They also found that soft rot fungi could create cavities in the S2 layer, but were not able to remove the highly lignified compound middle lamella and S3 layer. Militz et al. (2003) found that acetylated beech was more resistant against degradation by Trametes versicolor than by soft rot and soil inhabiting organisms. This supports the findings by Takahashi et al. (1989). Their soil bed assays showed that an increase in acetyl content led to a sharper decrease in the degradation of cellulose and lignin when compared to polyoses. Mohebby and Militz (2010) found that after seven years in soil contact soft rot were the dominant rot fungi, decaying non-acetylated wood severely and also attacking moderately acetylated wood slowly. The bacteria cooperated generally with the soft rot fungi to degrade the non-acetylated wood, limiting themselves to pits at the higher degrees of the acetylation. In the moderately acetylated wood, white rot decay was at developing stage, while it was at initial stage at the highest WPGs.
Molecular methods are also convenient tools to study the details in fungal decay of modified wood. Pilgård et al. (2010) used quantitative real-time PCR for quantification of fungal colonisation in modified wood using T. versicolor. For all wood treatments, the maximum fungal DNA level was recorded after an incubation period of two weeks, followed by a decline until the end of the trial at eight weeks. Schmöllerl et al. (2011) studied modified wood samples exposed to Postia placenta at different time intervals (2–26 weeks). Acetylated wood had the lowest mass loss and fungal DNA content. In the beginning of the incubation of all treated wood samples, the genes coding for oxidative metabolic activity had higher expression levels than the untreated control. In the end of the incubation period most of these genes were less expressed than in the untreated control. Pilgård et al. (2011, 2012) studied three acetylation levels of Southern Yellow Pine against P. placenta at time intervals up to 36 weeks. The fungus was able to colonise the wood at an early stage of incubation, before any measurable mass loss was found. The gene expression after four weeks showed that two genes related to carbohydrate metabolism were expressed in a higher amount in P. placenta during growth on untreated wood than during growth on acetylated wood. However, for a third gene, also related to carbohydrate metabolism, the relationship was the opposite. Two genes related to oxidative metabolism were expressed in a higher amount in P. placenta during growth on acetylated wood than during growth on untreated wood. Ringman et al. (2012) reviewed research on cellulose degradation by P. placenta and how it is affected by wood modification.
Mode of action hypotheses
It is well established that acetylation is able to impart a significant degree of decay resistance to wood, but the scientific literature still need to improve the evidences regarding the mechanism by which acetylated wood is protected from microbiological decay (Hill 2006). Another challenge is that the exact mechanism of brown rot decay still is hypothetical and controversial (e.g. Kang et al. 2009). Studies of the genome, transcriptome, and secretome of various decay fungi reveal that there is more diversity than earlier believed, and the border between white and brown rot get more blurry (Martinez et al. 2009). Listed below are hypotheses on why acetylated wood is protected against decay fungi. The hypotheses that have gained most support in the literature are physical blocking and reduction in cell wall moisture content.
Substrate recognition
Hill (2006) provides the following summary: ‘Acetylation changes the chemical nature of the substrate, OH groups are replaced by acetyl moieties. As a consequence it has been hypothesised that various specific enzymes are no longer capable of recognising the substrate and hence cannot break down the cell wall polymers. It is known that such enzymes are not capable of penetrating the cell wall of sound wood, as a consequence, if such mechanism is operative, then this would presumably only be important for erosion attack from the lumen inwards. It may, however, be possible that the action of low molecular wood degrading substances is chemically, or sterically, prevented by substitution of the cell wall polymer OH groups’. According to Rowell et al. (2009) the earliest ideas centred around that the specific enzymatic attack could not take place due to OH substitution and modification of the substrate (e.g. Stamm and Baechler 1960; Takahashi et al. 1989; Takahashi 1996). Takahashi et al. (1989) suggested that the high decay resistance of acetylated wood against white rot decay is associated with high reactivity of lignin with acetylating reagents. Peterson and Thomas (1978) had earlier put forward that substitution of lignin OH groups was not considered to be the primary protection mechanism, due to the relative infrequency of such groups in the lignin. They suggest that a potential unavailability of carbohydrate as an energy source may result in the protection of the lignin component. Rowell et al. (2009) suggest that the mechanism of resistance might be as simple as protecting arabinose from hydrolysis as it might contain a trigger to start degradation. More specific, the modification of the arabinose sugar in hemicelluloses considering that this is a sugar of L-pentose type in the hemicelluloses and it is the only sugar in a strained five-membered ring. However, given cellulose and thermally modified wood are both susceptible to decay, it is clear that even if the hypothesis about this mechanism is true it cannot be the only one. Suttie et al. (1999) showed that with soft rot attack, the WPG level, rather than the extent of hydroxyl substitution is the sole factor determining protection from decay. A series of studies on Corsican pine (Papadopoulos and Hill 2002; Hill et al. 2005a,b, 2006, 2009; Hill and Kwon 2009; Papadopoulos et al. 2010) argues against substrate recognition. Pilgård et al. (2011, 2012) found that gene expression is not inhibited but altered, most likely delayed, in acetylated wood compared to untreated wood. A review by Hill (2009) concludes that ‘Substrate recognition effects are very unlikely to be of importance given the inherent lack of enzyme accessibility.’ The review by Papadopoulos (2009) concludes that the mechanism of protection is not chemical/biological in origin. The degree of cell wall bulking caused by the adduct, rather than the extent of OH substitution is the primary factor controlling the biological durability and water vapour sorption.
Decrease in wood moisture
It is well established that acetylation of wood results in a decrease in the equilibrium moisture content of the wood at a given relative humidity (Hill 2006). It is also well known that fungal attack of wood will only occur if the wood exceeds minimum moisture content (Eaton and Hale 1993). Hence, it is a likely explanation that acetylation may provide decay protection by simply lowering the cell wall moisture content below the level sufficient to support decay (Stamm and Baechler 1960; Rowell 1983; Ibach and Rowell 2000). But this threshold varies between fungal species. Hill (2009) concludes that ‘Reduced cell-wall water content remains the most likely mechanism, but there is clearly much work needed to understand fully this phenomenon.’ It is also important to be aware of that it is difficult to separate the mechanisms of decrease in equilibrium moisture content and physical blocking (Hill 2006).
Physical blocking
Acetylation might act to physically block the ingress of degradative agents into the cell wall. Blocking of action of fungal catalysts appears to be the primary protection mode of the acetylation technique. Forster et al. (1997) and Forster (1998) supported the theory of the bulking effect of the covalently bounded acetyl group. Mohebby (2003) speculated that there are very small regions in the cell wall that are not acetylated due to the size of the acetate group but that are accessible to free radicals produced by fungi. A series of studies on Corsican pine, later extended to other wood species, promotes the hypothesis of cell wall bulking rather than OH substitution and lack of substrate recognition. Papadopoulos and Hill (2002) worked with different anhydrides with different molecular sizes and stated that decay resistance was a function of WPG rather than extent of hydroxyl substitution. Hill et al. (2005a) were able to demonstrate that the mechanism could be correlated with the volume occupied by the covalently bonded acyl groups in the cell wall and that a reduction in cell wall moisture content was the most likely mechanism for imparting decay resistance. Hill et al. (2006) confirmed decay resistance to be related to WPG. Hill et al. (2009) and Hill and Kwon (2009) expanded this work to include sapwoods and heartwoods of: Japanese larch, Korean pine, European beech and oriental white oak. It showed that the decay resistance due to modification with hexanoic or acetic anhydride is not a function of degree of OH substitution. However, there are variations in behaviour between different species when decay mass loss is plotted against WPG. The reasons for these differences are not known at this time, although two possible explanations were put forward: differences in distribution of the bonded acyl groups at a cell wall level, and there may be more than one mechanism operative, in which case a range of behaviour will be observed depending upon the relative importance of substrate recognition compared to cell wall bulking. They state that further work is required in order to increase the scientific understanding of how acetylation is able to provide decay resistance against microbiological attack.
Hill (2006a) point out that most studies had been concerned with service life predictions rather than investigating the decay protection mechanism(s) in any depth. It may be that the generally lower levels of acetyl substitution are required to provide protection against white rot fungi is a time-delay effect rather than an absolute threshold (Hill 2006a). From a study with the brown rot fungus Coniophora puteana Hill et al. (2006) concludes that the decay protection threshold is dependent on both the exposure time and the fungal strain used. Hill (2006) also states that it is apparent that thresholds are not fixed, but are dependent upon factors other than simply WPG.
Prediction model
From published studies data was collected from papers where it was possible to extract: treatment level, performance data of acetylated and control samples, wood species and test method. The following studies are included: van Acker (2003), Alfredsen and Westin (2009), Beckers et al. (1994), Edlund and Jermer (2007), Larsson–Brelid et al. (1997), Rowell (2010). Both laboratory and field tests were included.
Performance of acetylated wood was calculated as mass loss or decay rating of test sample/control (denoted T/C). It should be pinpointed that T/C measures the performance relative to untreated control samples, and therefore relates more to natural durability classification than to classification of performance of traditional wood preservatives.
Statistical tests were performed as analysis of variance (ANOVA) where only category variables were studied, regression analysis were only continuous variables were studied, and as analysis of covariance (ANCOVA) where category variables were studied together with a continuous variable. Student's t-tests were used for multiple testing of group means. Effects with probability of type 1 error smaller than 0·05 were considered significant. In total 116 test results from the above mentioned studies were included in the statistical analyses. Several characteristics were used to model T/C (Table 1).
Characteristics used to motel performance as test sample/control. For decay tests it was distinguished between sterile and unsterile laboratory tests and field trials above ground (use class 3) and in soil contact (use class 4)
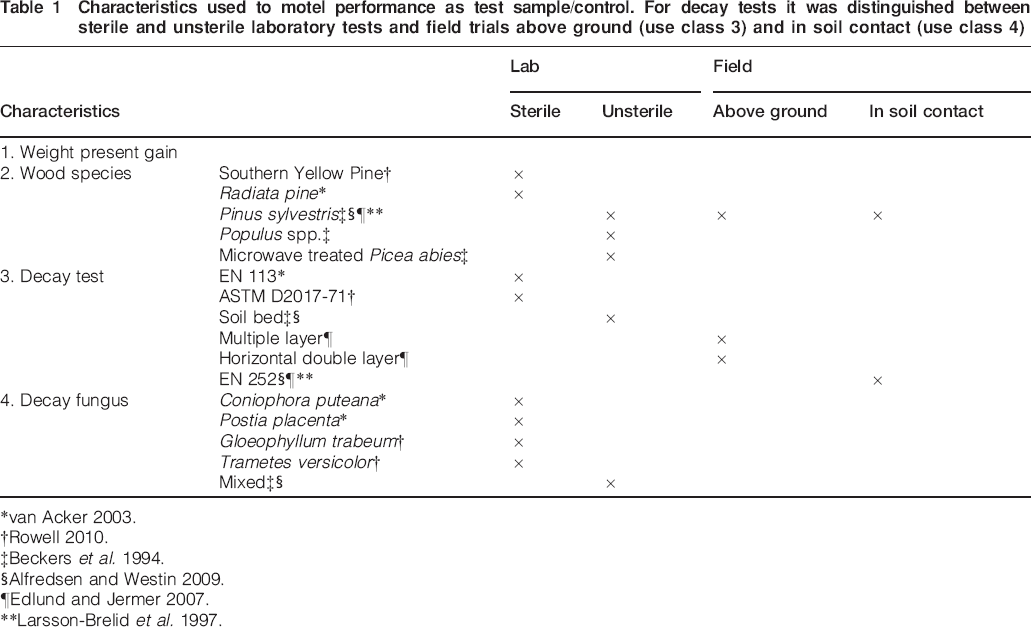
*van Acker 2003.
†Rowell 2010.
‡Beckers et al. 1994.
§Alfredsen and Westin 2009.
¶Edlund and Jermer 2007.
**Larsson-Brelid et al. 1997.
The model with the best performance in describing T/C totally for all studies included WPG (P<0·0001; F ratio, 103·6) and wood species (P<0·0001; F ratio, 7·8). The model described 62% (R2 = 0·62) of the variation in T/C. Results from multiple testing with WPG as covariate are shown in Table 2.
Multiple test with WPG as covariate. Levels not connected by same letter are significantly different
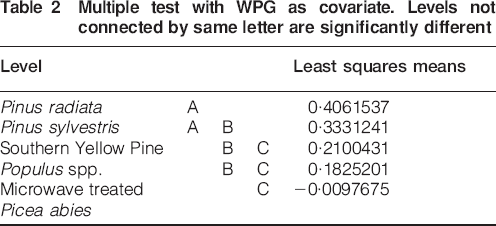
Figure 1 show predicted T/C values of various wood species over the range of WPG applied in the original decay studies.
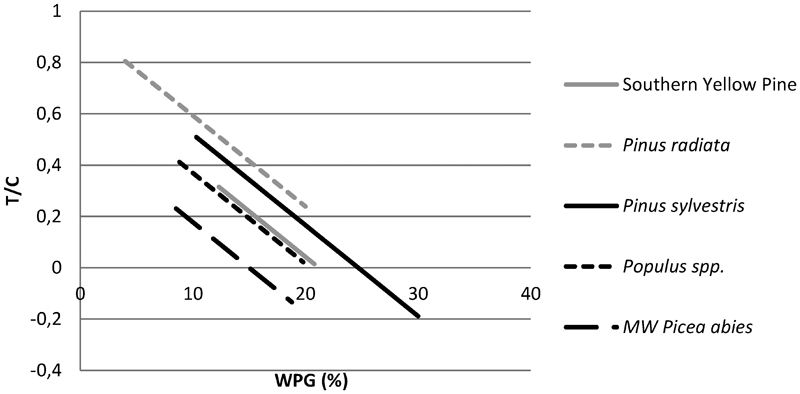
Predicted performance, T/C, of various acetylated wood species versus WPG
There was no significant effect of type of decay test when WPG was used as covariate in a model. Weight present gain described the largest share of variance in all models, and a linear regression model showed that WPG alone described 52% of the variance of T/C.
When analysing data across several studies, with different experimental designs, there is a risk that the model is affected by uncharacterised confounding variables or by imbalance of the experimental designs.
In the following analyses are based on the data from a round robin EN 113 test of acetylated Pinus radiata performed at eight different test laboratories in Europe (van Acker 2003). A model including WPG (P<0·0001; F ratio, 111·3) and type of fungus (C. puteana or P. placenta) (P = 0·0012; F ratio, 11·6) described 67% of the variance of T/C. Using WPG as covariate T/C was significantly higher for C. puteana than for P. placenta.
The statistical analyses above show that WPG can explain approximately 50% of the performance, measured as T/C, for acetic anhydride treated wood. Other of the applied variables, like wood species or type of fungus can reduce the variance in T/C by additional 15%. The model performance can probably be improved, but some level of residual variation is unavoidable, especially since T/C is measured based on the activity of living organisms.
Future research opportunities
From the above knowledge main further research opportunities with regard to acetic anhydride modified wood and decay fungi can be defined.
How well is acetic anhydride modified wood protected against different decay fungi?
There seems to be a difference in the WPG needed to protect wood against different fungal groups (white-, brown and soft rot). Is that really the case and if so why?
Relatively few fungal species have been tested so far, is there a huge variation between different fungi? Or between different strains of the same fungus?
What is the colonisation and succession pattern of different wood decaying fungi?
Does fungal diversity vary between acetic anhydride modified wood and other wood modification, preservative treatments or untreated wood species?
What are the anatomical and chemical changes in the wood during decay in the points listed above?
What is the mode of action of acetic anhydride modified wood against decay fungi?
Is moisture exclusion and cell wall blocking the main mode of action against decay fungi?
How does wood moisture vary in the cell wall wood during decay?
Can experiments where modified wood is exposed to enzymes help understand the mode of action?
Many wood-destroying organisms are capable of expressing acetyl esterase enzymes, which can de-acetylate modified wood (Tsujiyama and Nakano 1996), what fungi express this enzyme and how important is it for fungal deterioration of acetic anhydride modified wood?
What are the changes in the entire transcriptome and secretome of relevant decay fungi during decay?
Since fungi are able to penetrate and colonise modified wood at early stages of incubation, how to optimise homogenous treatment and service life?
How to characterise the protective zone?
Does the zone vary between WPSs and tree species?
Does it affect different fungi differently?
What are the main test method challenges?
Does a threshold exist or is just a matter of delayed decay?
What is the effect of heartwood content and juvenile wood on decay resistance?
How to create a better way to compare tests between labs and field test sites?
It is worth to keep in mind that several of the bullet points above are highly relevant also for other wood materials than acetylated wood. For example the use of the term decay protection threshold is not an exact term, neither for untreated, preservative treated or modified wood. It will always be time dependent. But little literature is so far available. The point of initiation of decay and the rate of decay in different exposure situations is also the key to better service life prediction data. Another issue is potential differences in the reactivity of heartwood and sapwood, or between juvenile and mature wood in acetylation reactions. Considering the importance of this to any commercial operation, it is very little literature dealing with this topic (Hill 2006a). The presence of extractives in the wood can have a significant effect upon the results obtained during acetylation studies, with leaching of extractives into the acetylation medium leading to lower WPG values being recorded than have actually been obtained. It would clearly be impractical to use extractive free wood on an industrial scale, but alternative methods for determining the extent of the reaction (i.e. not gravimetric) are required (Hill 2006a). For acetylated wood, but also other treatments, it is difficult to draw comparisons between laboratory studies, even for the same treatment at similar treatment level, since they vary in test methods, fungal strains, virulence of the test strain and wood species. And it is even harder to summarise the results of field tests than for monoculture lab studies. Climate, exposure time, wood species and biological inoculum potential at the test site vary. The challenges with regard to service life prediction from field trials are discussed in more detail by e.g. Brischke et al. (2006, 2012) and Larsson–Brelid et al. (2011).
Footnotes