
Abstract
K+ channels in coronary arterial smooth muscle cells (CASMC) determine the resting membrane potential (E m) and serve as targets of endogenous and therapeutic vasodilators. E m in CASMC is in the voltage range for activation of L-type Ca2+ channels; therefore, when K+ channel activity changes, Ca2+ influx and arterial tone change. This is why both Ca2+ channel blockers and K+ channel openers have such profound effects on coronary blood flow; the former directly inhibits Ca2+ influx through L-type Ca2+ channels, while the latter indirectly inhibits Ca2+ influx by hyperpolarizing E m and reducing Ca2+ channel activity. K+ channels in CASMC play important roles in vasodilation to endothelial, ischemic and metabolic stimuli. The purpose of this article is to review the types of K+ channels expressed in CASMC, discuss the regulation of their activity by physiological mechanisms and examine impairments related to cardiovascular disease.
Ca2+ channels, K+ channels and coronary blood flow
K+ channels dominate the membrane conductance of coronary arterial smooth muscle cells (CASMC) at rest and thus determine E m (Figure 1). That is, E m is closer to the Nernst equilibrium potential for K+ (E K) than for other ions. E K in arterial smooth muscle is estimated to be −83 mV. 1 An asymmetric distribution of K+ across the cell membrane is maintained by the Na+/K+-ATPase and provides a large chemical driving force for K+ efflux by diffusion through open channels. If K+-selective channels were the only type of channels open in CASMC at rest, K+ would diffuse out of the cell until an electrical potential of −83 mV is developed. E m in CASMC, however, is not as negative as E K; generally, values around −50 mV are observed. 2–7 This less negative value of E m, compared with E K, in CASMC is because other ion channels are open at rest (e.g. those permeable to ions with more positive reversal potentials, such as Na+ and Ca2+).
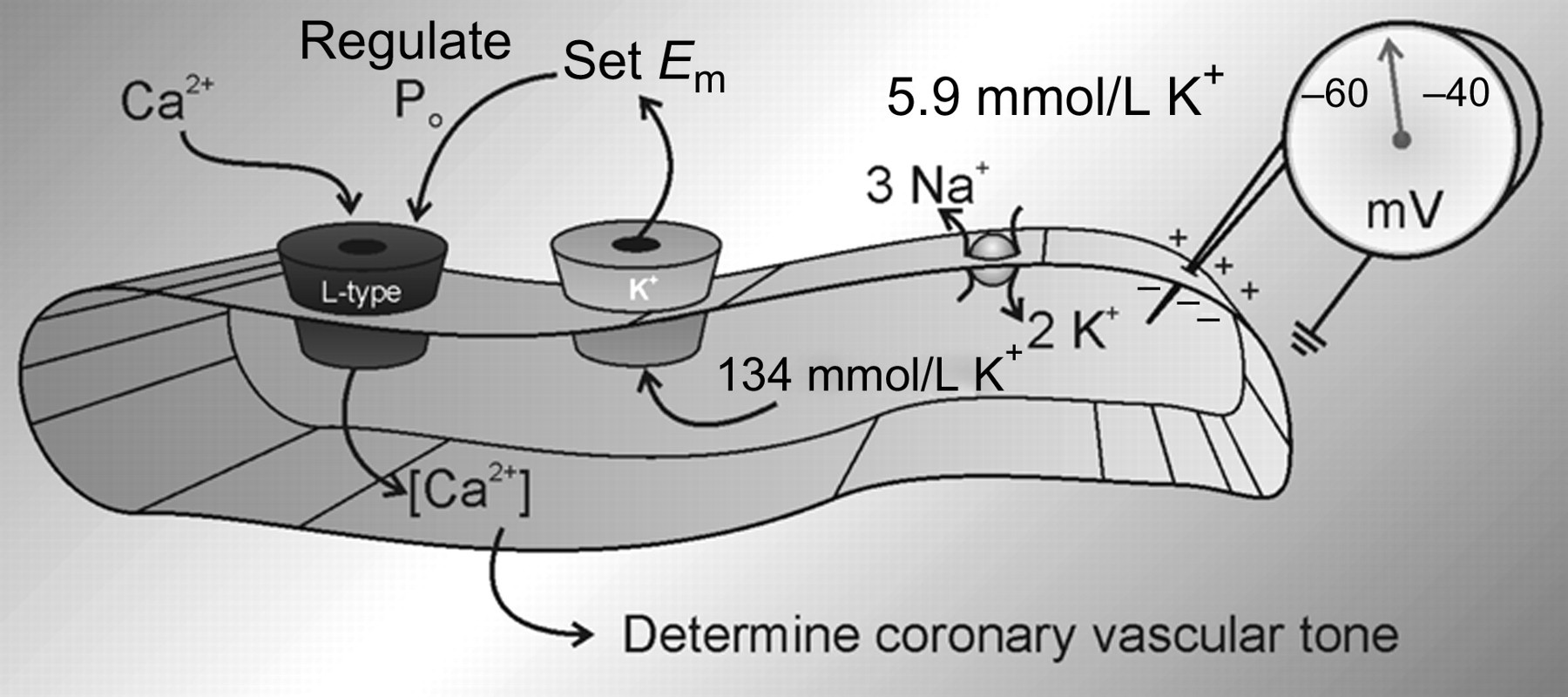
Electromechanical coupling in CASMC. The Na+/K+-ATPase accumulates intracellular K+, providing a large chemical gradient for diffusion of K+ through open channels. Efflux of K+ largely determines E m, which regulates the open probability of L-type Ca2+ channels. The level of free intracellular Ca2+ determines the contractile state of coronary vascular smooth muscle. CASMC, coronary arterial smooth muscle cells
Balancing the hyperpolarizing influence of K+ channels with the depolarizing influence of other channels produces a stable E m in CASMC typically between −60 and −40 mV. The voltage threshold for L-type Ca2+ current activation, and thus contraction, is in this range. 3,6,8,9 CASMC are generally electrically quiescent, respond to stimuli with graded potentials and do not normally display action potentials. However, if K+ channels are inhibited with tetraethylammonium (TEA; a non-selective blocker of K+ channels), Ca2+-dependent action potentials can be observed. 10–12 L-type Ca2+ channel inhibitors and K+ channel openers inhibit phasic contractions induced by TEA and 4-aminopyridine (4-AP; an inhibitor of voltage-dependent K+, KV channels 5,12–16 ). Such findings have led to the hypothesis that coronary vasospasm may be related to reduced K+ conductance in CASMC. 17
Multiple types of K+ channels are expressed in CASMC, but it seems that some have more important roles in determining E m and the state of contraction under resting conditions. For example, 4-AP strongly contracts conduit coronary arteries, 15,18 whereas charybdotoxin, iberiotoxin, apamin and glibenclamide cause little or no contraction. 15 Charybdotoxin and iberiotoxin inhibit large conductance Ca2+-activated K+ (BKCa) channels, apamin inhibits some small conductance Ca2+-activated K+ channels and glibenclamide inhibits ATP-dependent K+ (KATP) channels. There may be complex heterogeneity in ion channel expression and function throughout the coronary tree; while glibenclamide does not contract conduit coronary arteries, it does tend to diminish the balance between coronary blood flow and myocardial metabolism, indicating increased microvascular resistance. 19–22 BKCa, KV and KATP channels have important roles in certain facets of coronary vascular regulation; therefore, the properties of these channels and the modulation of their activity are discussed in the next section.
Antagonists of L-type Ca2+ channels such as nifedipine, verapamil and diltiazem increase coronary blood flow, 23–26 as these drugs relax coronary arteries and arterioles. 27–30 Conversely, Bay K 8644, a dihydropyridine with agonist rather than antagonist properties, elicits coronary vasoconstriction. 31 The coronary vasodilator effects of L-type Ca2+ channel blockers are rather simple to understand; these drugs directly inhibit a major Ca2+ entry pathway in CASMC and thus reduce the intracellular Ca2+ required to maintain contraction. The vasodilatory effects of K+ channel openers are similar; however, their mechanism involves an additional step to hyperpolarize E m. Pinacidil and cromakalim, two K+ channel openers, increase coronary blood flow. 32–34 These agents relax coronary smooth muscle by activating KATP channels. 35–38 In doing so, these drugs render E m more negative and reduce the open probability of L-type Ca2+ channels in CASMC, 8,9 thus limiting Ca2+ influx and promoting vasodilation. This interplay between K+ channels and voltage-dependent Ca2+ channels in CASMC represents a reasonably straightforward example of electromechanical coupling (Figure 2).
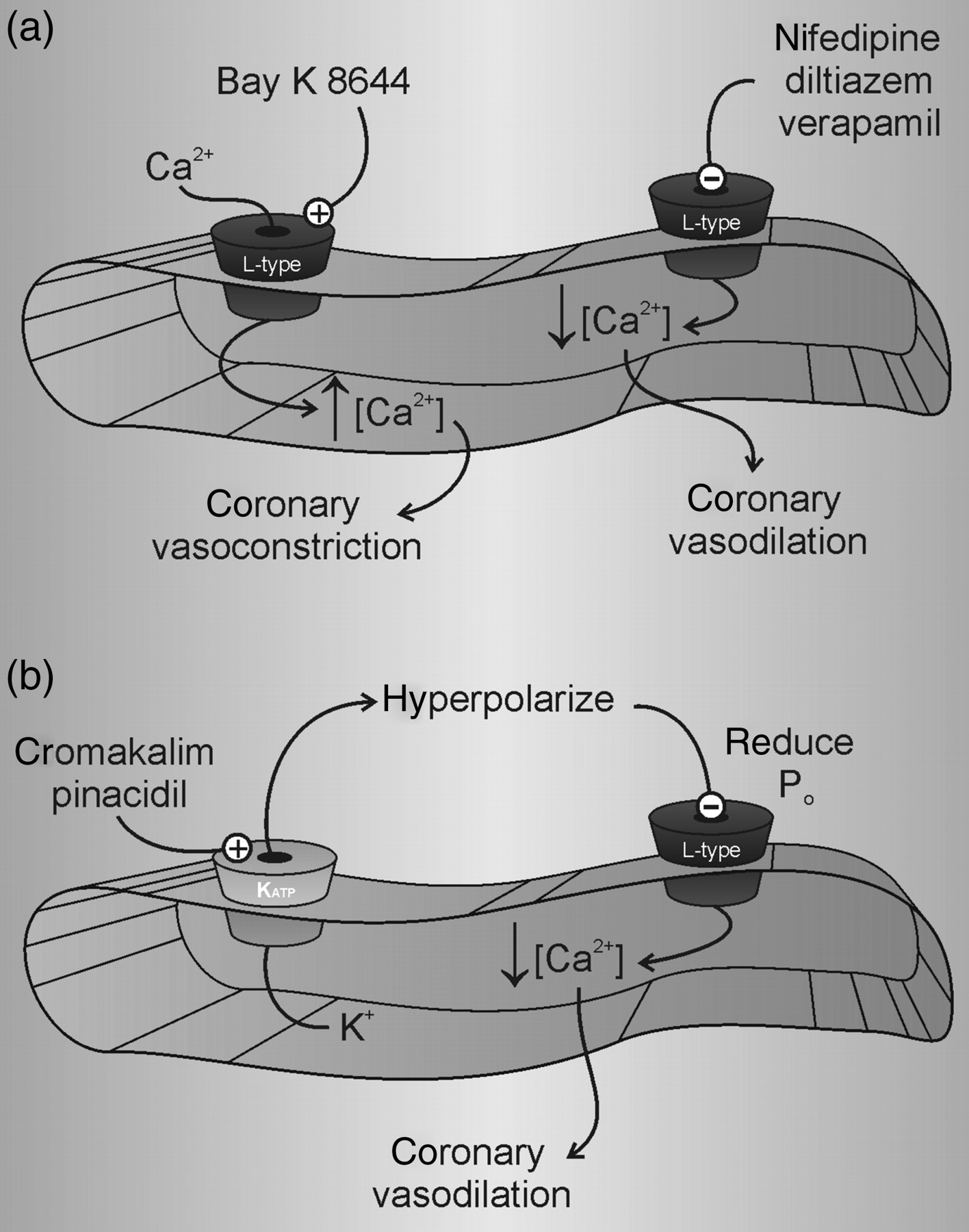
K+ channel openers and Ca2+ channel blockers. (a) Drugs that modulate L-type Ca2+ channel activity affect coronary vascular tone. Nifedipine, diltiazem and verapamil reduce intracellular Ca2+ and lead to vasodilation. In contrast, Bay K 8644 increases intracellular Ca2+ and leads to vasoconstriction. (b) K+ channel openers, e.g. KATP channel agonists pinacidil and cromakalim, hyperpolarize E m. This reduces the open probability of L-type Ca2+ channels, lowers the free intracellular Ca2+ concentration and produces vasodilation. The KATP channel antagonist glibenclamide has an opposite effect
CASMC, of course, express a wide variety of cation and anion channels with important functions that extend far beyond electromechanical coupling (e.g. references 39–44 ); however, this review focuses on just a few of the types of K+ channels involved in coronary vasodilation. While only BKCa, KV and KATP channels are discussed, it is clear that other K+ channels are expressed in CASMC and play important roles (e.g. additional types of Ca2+-activated K+ channels and inwardly rectifying K+ channels 45–47 ). Additionally, it is clear that the ion channels of other cells in the coronary vascular wall function to regulate microcirculatory tone (e.g. KATP channels in endothelial cells regulate NO release; 48,49 ); however, our discussion will be limited to K+ channels in CASMC.
Types of K+ channels expressed in CASMC
The K+ currents and the electrophysiological properties of CASMC have been demonstrated using patch clamp techniques (e.g. by Wilde and Lee 50 in dog coronary artery). CASMC E m was −51 mV and the current was TEA-sensitive, K+-selective and abolished by intracellular Cs+. Input resistance was very high (>108 Ohms); therefore, small changes in current produce large changes in E m. K+ current increased with elevations in extracellular or intracellular Ca2+, indicating the functional expression of KCa channels. Total whole-cell K+ current consisted of at least two components based on amplitude, time dependence, sensitivity to Ca2+ and pharmacology. Specifically, there was a fast activating current followed by a larger and slower current; the fast component was more sensitive to block by extracellular Cs+, whereas the slow component was more sensitive to block by TEA. The slower component was increased by elevations in intracellular or extracellular Ca2+, but the fast component was little affected. Other patch clamp studies of rabbit and human CASMC elaborated on these two major types of K+ currents (established to be BKCa and KV) and demonstrated a predominance of KV current in the physiological range of E m. 51–54 BKCa channels activate at more depolarized potentials than KV channels in CASMC 52 but increases in intracellular Ca2+ open BKCa channels at physiologically relevant voltages. 55,56 Activating Ca2+ for BKCa channels in CASMC comes from spontaneous release from intracellular stores, 56 L-type Ca2+ channels 57 and receptor stimulation. 58,59
The first patch clamp studies of CASMC K+ channels did not reveal notable KATP channel activity, but this is largely due to the use of conventional dialyzed patch techniques that included mmol/L concentrations of intracellular ATP and effectively inhibited KATP channels. Dart and Standen, 60,61 however, used nystatin- and amphotericin-perforated patch techniques (where the intracellular ATP concentration was not artificially manipulated) to demonstrate adenosine- and hypoxia-activated KATP current in CASMC. Pinacidil activates KATP currents in human CASMC regardless of the intracellular ATP concentration. 38,52 The discussion below concerns the molecular composition and regulation of BKCa, KATP and KV in CASMC (Figure 3).
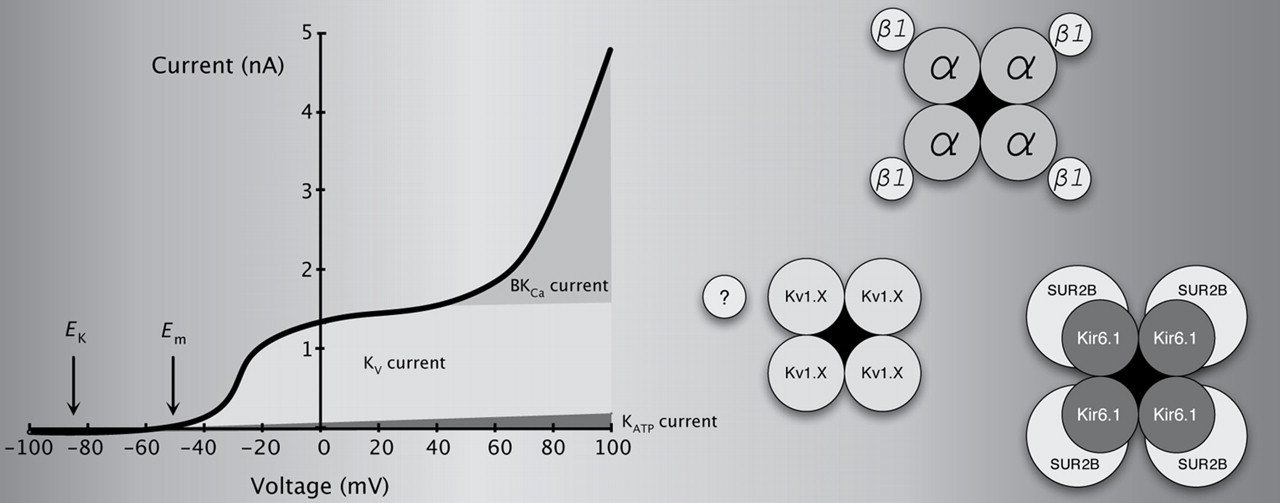
Macroscopic K+ currents in CASMC and molecular composition of the channels responsible. KV and BKCa channels mediate the two most prominent currents, although KATP channels are also expressed. Functional BKCa channels can be formed by the assembly of four pore-forming α subunits, which contain regulatory regions responding to voltage and Ca2+. In CASMC, most BKCa channels are complimented by four accessory β1 subunits, which increase Ca2+/voltage sensitivity and alter biophysical and pharmacological properties. KATP channels are an octameric complex of four pore-forming α subunits and four regulatory sulfonylurea receptors (SUR). The likely KATP subunits in CASMC are Kir6.1 and SUR2B. Functional KV channels are formed by the assembly of four pore-forming α subunits, which are sensitive to voltage. The addition of auxiliary or modifying subunits can change channel properties. The identity of KV channel subunits in CASMC remains an active area of investigation; however, a major portion of the current is sensitive to correolide, indicative of KV1 expression. CASMC, coronary arterial smooth muscle cells
BKCa channels in CASMC
BKCa channels are large conductance (>200 pS), voltage-sensitive K+ channels regulated by Ca2+ in an allosteric fashion. 62 The assembly of four pore-forming α subunits, products of single alternatively spliced gene (KCNMA1), can form functional BKCa channels. Studies have identified four types of modulatory β subunits. The accessory β1 subunit (KCNMB1) increases sensitivity to Ca2+, slows activation and deactivation, and reduces sensitivity to iberiotoxin. 63 In human CASMC, the majority of BKCa channels contain this β1 subunit. 64 Multiple stimuli regulate BKCa channel activity. As can be surmised from the previous section, those coupling to coronary vasoconstriction typically inhibit BKCa channels, while those coupling to coronary vasodilation typically activate BKCa channels.
BKCa channels are involved in CASMC responses to endothelial stimulation. The coronary vascular endothelium produces a number of relaxing factors such as nitric oxide (NO), prostacyclin (PGI2) and epoxyeicosatrienoic acids (EETs). While the term endothelium-derived hyperpolarizing factor (EDHF) is typically reserved for the mediator(s) of dilations remaining after inhibition of NO synthase and cyclooxygenase, the term EDHF should be applied to any endothelium-derived factor that activates K+ channels. That is, one can consider NO an EDHF, as it activates BKCa channels in CASMC. 65–68 S-nitrosothiols have similar effects on BKCa channels in CASMC, and the actions of both NO and S-nitrosothiols appear to involve cyclic guanosine monophosphate (cGMP) signaling. 68–70 Some metabolites of arachidonic acid produced by cyclooxygenase, such as PGE2, activate BKCa channels through cAMP and cross-activation of the cGMP-signaling pathway. 71,72 Various metabolites of arachidonic acid produced by other enzyme systems, such as EETs from cytochrome P450 metabolism, activate BKCa channels in CASMC. 73–77
The mediators of ischemic and metabolic vasodilation in the coronary circulation have not been identified conclusively; 78,79 however, K+ channels appear to be end-effectors and several putative dilator mechanisms have been examined. Sympathetic stimulation plays an important role in coronary metabolic vasodilation 80,81 and the β-adrenergic agonist isoproterenol activates BKCa channels in CASMC. 72,82 This appears to be mediated by Gs α and cross-activation of cGMP-dependent protein kinase; dopamine has similar effects. 72,83 Adenosine is proposed as a mediator of ischemic and metabolic vasodilation; however, no patch clamp data show a stimulatory effect of adenosine on BKCa channels in CASMC (such data are available in mesenteric smooth muscle 84 ). H2O2, a putative metabolic vasodilator, activates BKCa channels in CASMC; 85 this may be mediated by activation of phospholipase A2 and production of lipoxygenase metabolites. 86
A variety of substances that constrict coronary vessels inhibit BKCa channels. Angiotensin II, 87,88 endothelin-1 87,89 and thromboxane A2 90 inhibit BKCa channels in CASMC. Common underlying mechanisms for inhibition of BKCa by these G protein-coupled receptors may include activation of protein kinase C 91 or c-Src, a tyrosine kinase. 92
KATP channels in CASMC
Functional KATP channels are composed of eight subunits: four pore-forming subunits (either Kir6.1 or Kir6.2) and four sulfonylurea receptors (SUR1, SUR2A or SUR2B; the 2A and 2B isoforms are splice variants of the same gene, ABCC9). In situ hybridization and RT-PCR show Kir6.1 and SUR2B mRNA in coronary arteries and arterioles 93,94 and immunohistochemistry indicates Kir6.1 and SUR2 proteins in coronary arterioles <100 μm. 94,95 Kir6.1-null mice demonstrate coronary vasospasm, 96 but Kir6.2-null mice do not. 97 Genetic ablation of ABCC9 (encoding both SUR2A and SUR2B) also produces coronary vasospasm. 98 These studies support the idea that CASMC KATP channels are composed of Kir6.1 and SUR2A/B subunits, as knockout of these subunits produce coronary vasospasm. The mechanism may be much more complex, however, as smooth muscle-specific restoration of SUR2 expression in knockout mice does not correct coronary vasospasm. 99 Thus, coronary vasospasm in Kir6.1 and SUR2A/B mice may be secondary to some other change and unrelated to channels in CASMC.
KATP channels in CASMC may be the targets of endothelium-dependent relaxing factors, but there are few patch clamp studies to provide direct evidence. For example, no patch clamp study has demonstrated the activation of KATP channels by NO in CASMC (such data are available in bladder smooth muscle 100 ). With regard to ischemic vasodilation, it has been demonstrated that adenosine and hypoxia activate KATP channels in CASMC. 60,61 In the context of metabolic vasodilation, dopamine, an endogenous catecholamine, has been shown to activate KATP in CASMC. 101 In contrast, coronary vasoconstrictors including endothelin-1, vasopressin and angiotensin II inhibit KATP channels. 102–104
KV channels in CASMC
More than 40 KV channel subunits have been identified; because of this diversity, the molecular identity of KV channels in CASMC remains unclear. Complexity arises not only from the sheer number of genes involved, but is compounded by the alternative splicing of mRNA encoding individual subunits, assembly of different subunits into heteromultimeric channels and a variety of post-translational modifications. Functional KV channels are a tetrameric assembly of pore-forming α subunits, but multiple types of accessory subunits modify KV channel properties. The KV current in CASMC is a delayed rectifier with little or no time-dependent inactivation. Its biophysical behaviors share many properties with some channels cloned from the KV1 (Shaker; KCNA), KV2 (Shab; KCNB) and KV3 (Shal; KCNC) gene families. Sections of the human heart demonstrate KV1.5 immunoreactivity in CASMC. 105 Further, KV1 family genes, multiple KV1 proteins and current sensitive to correolide (a blocker of Shaker-type KV1 channels) are expressed in canine and rat coronary artery. 106,107 KV3 gene transcripts and a highly TEA-sensitive KV current are expressed in pig coronary arteries. 108 More studies will be required to determine the molecular composition of delayed rectifier channels in CASMC, as well as differences that may be due to species, vessel caliber and other factors (e.g. side of the heart from which the vessels originate 107 ).
NO and iloprost (a PGI2 mimetic), but not 11,12-EET (an EDHF from the cytochrome P450 pathway), activate KV channels in CASMC. 67 No patch clamp study demonstrates effects of adenosine or β-adrenergic agonists on KV current in CASMC; however, a common signaling pathway may be cAMP-dependent protein kinase, which has been shown to activate KV channels in portal vein smooth muscle. 109,110 H2O2, a putative metabolic vasodilator, activates KV channels in CASMC through unknown signaling mechanisms. 111 No patch clamp studies demonstrate inhibition of KV channels by vasoconstrictors in CASMC; however, data are available from other vascular beds. Endothelin-1, angiotensin II and thromboxane A2 inhibit KV current in the mesenteric artery, pulmonary artery and portal vein smooth muscle and a common mechanism appears to be activation of protein kinase C. 112–115 In cerebral arterial smooth muscle, Rho kinase and tyrosine kinases, involved in CASMC Ca2+ regulation and contraction, inhibit KV current. 116,117
Regulation of CASMC K+ channels by endogenous vasodilators
Many mechanisms regulate coronary vascular resistance, 79,118,119 several of which were introduced in the above section. Factors from the endothelium influence coronary vascular tone, ischemia is an important pathophysiological stimulus and metabolism is the key regulator of coronary blood flow on a beat-to-beat basis. Importantly, K+ channels play a central role in producing vasodilation to all of these stimuli and their endogenous mediators. At present, the study of K+ channels in coronary blood flow depends almost entirely upon pharmacological blockers (e.g. iberiotoxin, glibenclamide and 4-AP) and very few investigators have been able to take advantage of newer, and perhaps more specific, molecular tools. Hopefully, models that incorporate knockout, transgenic and RNA interference approaches will receive more attention in the future. The following section summarizes studies of K+ channels in coronary vascular relaxation (in vitro studies of arteries and arterioles) and the regulation of coronary blood flow (in vivo). For simplicity, the role of K+ channels in various coronary vascular responses are discussed one at a time; however, it is important to recognize that key vasoregulatory factors modulate coronary microvascular tone via multiple K+ channels subtypes. For example, coronary endothelial-dependent dilation to bradykinin is mediated, to varying degrees, by BKCa, KATP and Kv channels (Figure 4a). Another example is how ischemic coronary vasodilation involves both KATP and Kv channels (Figure 4b). Dilators appear to regulate coronary blood flow through mechanisms involving multiple types of K+ channels; therefore, if one K+ channel type is inhibited/impaired, another may compensate. Such redundancy may act to preserve myocardial perfusion. 120 The precise roles of redundant K+ channel pathways and signaling cross-talk in the overall control of coronary blood flow merit further investigation.
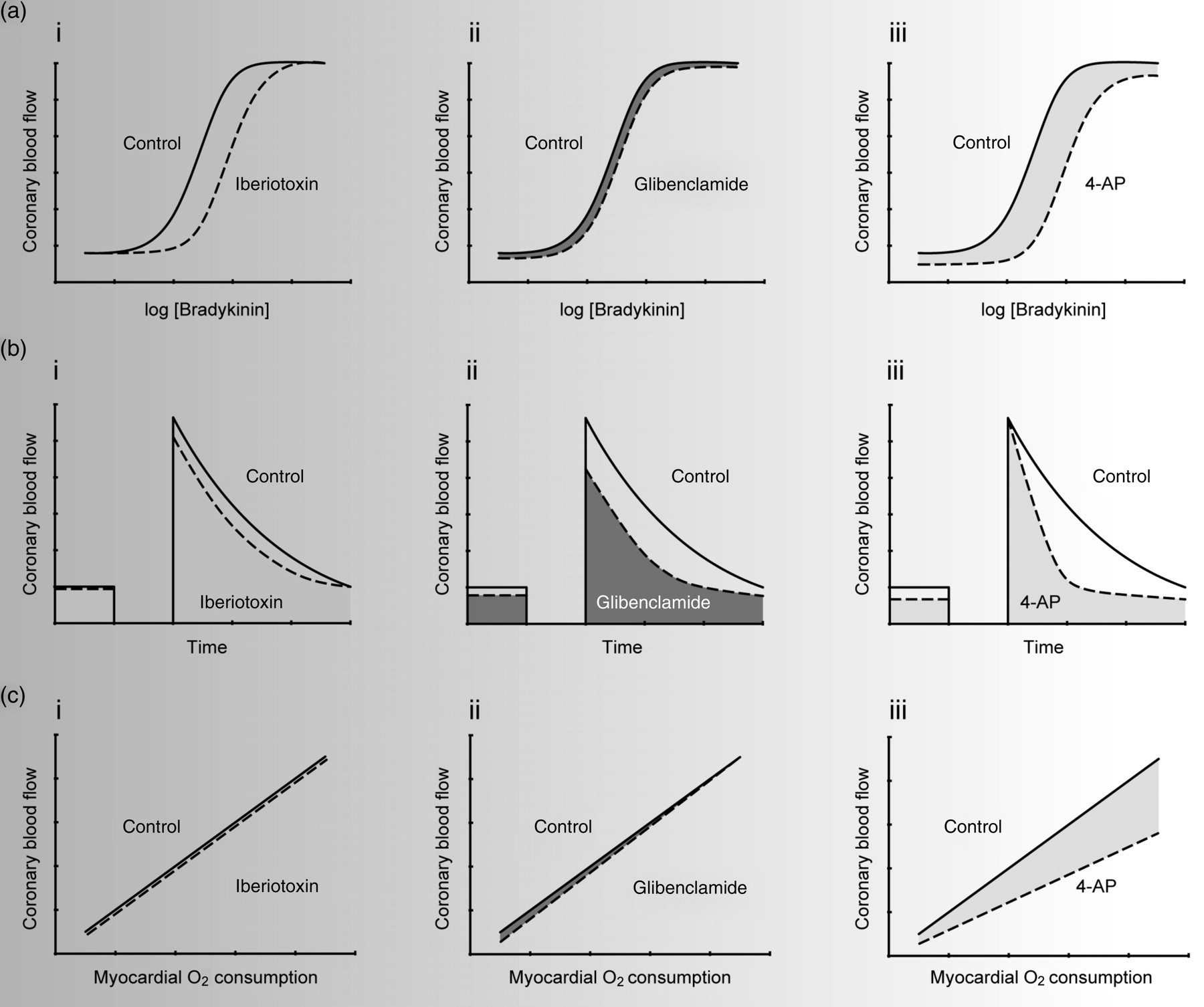
K+ channels in the regulation of coronary blood flow. (a) Effects of iberiotoxin (i), glibenclamide (ii) and 4-AP (iii) on endothelium-dependent coronary vasodilation. (b) Roles of BKCa (i), KATP (ii) and KV (iii) channels in coronary reactive hyperemia. (c) Coronary metabolic vasodilation does not involve BKCa (i) or KATP (ii) channels, but does depend upon the opening of KV channels (iii). 4-AP, 4-aminopyridine
Endothelium-dependent vasodilation
Agonists such as acetylcholine and bradykinin couple to NO, PGI2 and EDHF production and thus coronary vasodilation. Many other endothelium-dependent agonists have been tested and adenosine-induced coronary dilation also has an endothelial component. 48,49 The contribution of specific K+ channel types to endothelium-dependent coronary vasodilation has been studied in numerous species. Iberiotoxin, a selective BKCa channel antagonist, inhibits bradykinin-induced coronary vasodilation in dogs in vivo. 121 Similarly, charybdotoxin, a BKCa channel antagonist, inhibits bradykinin-induced vasodilation of human coronary arterioles. 122 Iberiotoxin attenuates NO- and cGMP-induced relaxation of human 66 and animal 123–125 coronary arteries, indicating a role for BKCa channels in endothelium-dependent NO-induced hyperpolarization. 126 The role of BKCa channels in PGI2-induced coronary artery relaxation may depend on species, as iberiotoxin inhibits hyperpolarization induced by iloprost, a stable PGI2 mimetic, in porcine CASMC, 127 but iloprost has no effect on BKCa channels in bovine CASMC. 67 No in vivo study has addressed the role of BKCa channels in iloprost-induced coronary vasodilation. When NO and PGI2 production are blocked, the remaining relaxation is attributed to EDHF (which may be a cytochrome P450 metabolite). Iberiotoxin inhibits EDHF-induced coronary relaxation in vitro in pigs 128 and EDHF-induced coronary vasodilation in vivo in dogs. 129 Some investigators present data indicating no significant role for BKCa channels in endothelium-dependent relaxation of the coronary arteries of guinea pigs, 130 dogs and monkeys, 131 as well as humans. 132 However, the general consensus seems to be that BKCa channels play at least some role in endothelium-dependent coronary vasodilation (Figure 4ai).
It is generally reported that there is no effect of glibenclamide on endothelium-dependent vasodilation to bradykinin or acetylcholine in the coronary circulation, suggesting no role for KATP channels in this response. Negative findings for glibenclamide in endothelium-dependent vasodilation have been found for coronary blood flow in vivo 19,133–135 and for isolated hearts and arterioles in vitro. 122,136 Also, glibenclamide does not affect the coronary vascular smooth muscle response to NO donors. 20 However, KATP channels have been reported as mediators of bradykinin- and PGI2-, but not NO- induced vasodilation in rabbit hearts. 137 Glibenclamide has no effect on EDHF responses in pig 138 or human 139 coronary arteries. Thus, the general consensus is that KATP channels play little, if any, role in endothelium-dependent coronary vasodilation (Figure 4aii).
There has been some conflicting evidence regarding the role of KV channels in endothelium-dependent vasodilation; however, most studies have demonstrated that these channels contribute significantly. Using pig coronary arteries, Cowan and Cohen 140 demonstrated no effect of 4-AP on bradykinin-induced relaxation; however, Thollon et al. 141 showed that 4-AP inhibits NO-induced hyperpolarization. The latter is compatible with the effect of NO and iloprost to activate KV channels in bovine coronary smooth muscle. 67 While Illiano et al. 142 have shown that 4-AP has no effect on EDFH-induced relaxation in canine coronary arteries, Nakashima et al. 143 have observed an inhibitory effect of 4-AP in the same tissue. Three studies agree that 4-AP inhibits EDHF-induced relaxation in guinea pig coronary arteries. 130,144,145 Our group has demonstrated the role of 4-AP-sensitive KV channels in regulating basal coronary flow 106,111,146 and our preliminary studies indicate that KV channels play a large role in endothelium-dependent coronary vasodilation in vivo in dogs. These data do not necessarily suggest that KV channels have a more prominent role in the microcirculation, as 4-AP-induced vasospasm is observed throughout the coronary tree from conduit arteries to resistance arterioles. 147 The overall consensus for KV channels in endothelium-dependent coronary vasodilation is shown in Figure 4aiii.
Ischemic vasodilation
Large and transient compensatory increases in coronary blood flow follow a brief period of ischemia. This reactive hyperemia represents a repayment of blood flow debt due to the accumulation and washout of ischemic vasodilators. Mediators of ischemic coronary vasodilation have not been identified conclusively. Regardless of what the signaling mechanism might be, it is clear that K+ channels in CASMC play a large role in reactive hyperemia. Node et al. demonstrate that BKCa channels regulate blood flow to ischemic myocardium in dogs; 121 however, our preliminary studies show that penitrem A, another inhibitor of BKCa channels, has no effect on reactive hyperemia in pigs. 148 A modest role for BKCa channels in reactive hyperemia is shown in Figure 4bi. Most studies have focused on the contribution of KATP channels to ischemic vasodilation. In dogs and pigs, glibenclamide reduces the peak and duration of blood flow repayment following a brief coronary occlusion. 19,20,149–152 No study has addressed whether KATP channels regulate reactive hyperemia in the human coronary circulation; however, tolbutamide-sensitive channels may regulate this response in the human forearm. 153 The contribution of KATP channels to reactive hyperemia is summarized in Figure 4bii. We have demonstrated that KV channels regulate the duration of reactive hyperemia in the canine coronary circulation in vivo. 106 4-AP (300 μmol/L) reduced resting coronary blood flow by approximately 33%. Higher doses of 4-AP reduced coronary flow to the point of causing S-T segment depression, a sign of myocardial ischemia. KV channels regulate the duration of reactive hyperemia and reduce the repayment of flow debt. Additionally, 4-AP inhibited coronary vasodilation to NO and adenosine, two putative mediators of reactive hyperemia. The role of KV channels in coronary reactive hyperemia is shown in Figure 4biii.
Metabolic vasodilation
Myocardial metabolism is the key regulator of coronary blood flow on a beat-to-beat basis, from rest to climbing a flight of stairs or to maximal exercise. Coronary blood flow increases in an almost linear fashion with myocardial oxygen consumption. This very tight coupling between oxygen supply and demand is necessary because the heart has very little capacity for anaerobic metabolism and the myocardium, even at the resting heart rate, consumes approximately 75% of the oxygen delivered to it. Thus, there is very little reserve for oxygen extraction from coronary arterial blood and increases in work done by the heart must be met with parallel increases in coronary blood flow. Unfortunately, at present, we know more about which mechanisms are not responsible for coronary metabolic vasodilation. Further, complexity is added by species-specific differences in responses to exercise and mechanisms underlying them. 79 Thus, much research remains to be done on this central question of coronary physiology. Studies in the last two decades have focused on what K+ channels may be responsible for this critical physiological phenomenon. BKCa channels appear to play little, if any, role in coronary metabolic vasodilation. 154 Merkus and colleagues measured coronary blood flow and myocardial oxygen consumption in pigs exercising on a treadmill before and after intravenous treatment with TEA. 154 When one examines the relationship between coronary flow and myocardial oxygen consumption, there is no difference; however, there was a significant reduction in coronary venous oxygen tension. 154 TEA, even at an estimated plasma concentration of 0.6 mmol/L, is not ideal as an inhibitor of BKCa in exercise hyperemia studies, as TEA is also a ganglionic blocker 155 and reduces heart rate and blood pressure (major determinants of myocardial oxygen consumption) as well as coronary blood flow. We have conducted preliminary experiments with penitrem A, a more selective inhibitor of BKCa channels, in exercising pigs. Penitrem A has no effect on exercise-induced coronary metabolic vasodilation. 156 The lack of effect of BKCa channel blockers on the coronary blood flow versus myocardial oxygen consumption is shown in Figure 4ci.
Most studies in this area have focused on the contribution of KATP channels to coronary metabolic vasodilation. The general consensus is that KATP channels are not responsible for exercise-induced coronary hyperemia (Figure 4cii). Glibenclamide decreases coronary blood flow at rest in dogs, but has no effect on exercise-induced coronary vasodilation. 19–21,157 In humans, however, glibenclamide attenuates pacing-induced hyperemia. 158 KATP channels also appear to play a role in pacing-induced hyperemia in dogs. 159 However, comparing pacing and exercise-induced hyperemia is tenuous, as there are fundamental differences in the nature of the vasodilator mechanisms activated (e.g. pacing does not include the sympathetic control mechanisms elicited by exercise 160 ). Examples of discrepancies between pacing- and exercise-induced hyperemia have included conclusions made about BKCa and KATP channels. 20,156,159,161 If one considers exercise the most physiologically relevant stimulus, then the contribution of KATP channels to metabolic vasodilation is negligible. In humans performing handgrip exercise, glibenclamide has no effect on exercise-induced forearm blood flow, 162 an observation identical to the lack of effect of glibenclamide on coronary blood flow in exercising animals.
We have investigated the role of KV channels in coronary metabolic vasodilation. 163 Measuring coronary blood flow and myocardial oxygen consumption in dogs, we demonstrated that intracoronary infusion of 300 μmol/L 4-AP reduced blood flow at rest and attenuated pacing- and catecholamine-induced vasodilation. 163 The inhibitory effect of 4-AP on the relationship between myocardial metabolism and coronary blood flow is significantly more pronounced than has been observed with any other inhibitor of CASMC K+ channels such as glibenclamide, TEA or penitrem A. These data suggest that KV channels may be the most important channel type linking myocardial metabolism to coronary blood flow. Unfortunately, however, no study has yet addressed the role of KV channels in exercise-induced coronary vasodilation. We have suggested that H2O2 may be a signaling molecule linking myocardial metabolism to coronary blood flow, 163,164 and others support this idea. 161,165 H2O2 activates 4-AP-sensitive KV channels in CASMC 111 and H2O2-induced coronary vasodilation is blocked by 4-AP. 146 The role of KV channels in coronary metabolic vasodilation is shown in Figure 4ciii.
Dysfunction of CASMC K+ channels in disease
K+ channels in CASMC are involved in and affected by many processes related to the pathogeneses of cardiovascular diseases.
Diabetes, hyperglycemia and metabolic syndrome
A number of studies have demonstrated defects in CASMC BKCa channels in diabetes, hyperglycemia and metabolic syndrome. The activation of BKCa channels by arachidonic acid in CASMC of Zucker diabetic fatty rats is reduced compared with lean rats; this appears to be secondary to reduced activity of PGI2 synthase. 166 BKCa channels in CASMC from Zucker diabetic fatty rats also have a lower Ca2+/voltage-sensitivity due to reduced expression of the regulatory β1 subunit. 167 Diabetes increases BKCa current in CASMC from conduit arteries of pigs, 168 while spontaneous transient outward currents mediated by BKCa channels are reduced in arteriolar CASMC from diabetic pigs. 169 We have shown that metabolic syndrome reduces whole-cell penitrem A-sensitive and NS1619-activated BKCa current in CASMC. 170 KATP channels in CASMC are also affected by diabetes, as vasodilation due to KATP channel activation is impaired in human coronary arterioles from patients with type 1 and type 2 diabetes. 94 KV channel function is negatively impacted by high glucose concentrations in vitro. Elevated glucose impairs cAMP-mediated dilation by reducing KV channel activity in rat CASMC; 171,172 one mechanism appears to be nitration of the channels. 173 Another mechanism by which diabetes may impair K+ channel function in CAMSC is through the activation of protein kinase C. 174
Atherosclerosis and ischemia
Hypercholesterolemia impairs coronary vasodilation mediated by K+ channels. 175 In pigs, high blood cholesterol has been reported to increase 168 and decrease 176 BKCa current in CASMC. The contribution of KV channels to adenosine-mediated dilation of pig coronary arterioles is abolished by hypercholesterolemia. 108,176,177 Further, KV currents in porcine CASMC are reduced by high blood cholesterol. 108,176–178 Homocysteine, an independent risk factor for atherosclerosis, inhibits BKCa channels in pig CASMC. 179 BKCa channels in smooth muscle cells from coronary plaque have a higher activity than those in normal CASMC. 180 Further, compared with normal CASMC, the BKCa channels of CASMC from atherosclerotic plaques respond differently to growth factors. 181 BKCa channels contribute significantly to endothelium-dependent vasodilation in the ischemic myocardium of dogs. 121,182 KATP and KV channels compensate to preserve hypercapnic coronary flow when NO production is absent. 183 Acute hypoxia dilates guinea pig hearts 184 and pig coronary arteries and arterioles 185,186 through the activation of KATP channels; however, in rabbit coronary arteries, KATP channels are not involved. 187 Chronic hypoxia has opposite effects on KV current magnitude in right and left CASMC from rats, suggesting differences in KV channel isoforms or signaling mechanisms. 188 Acidosis increases KV, but not BKCa or KATP, current in rat CASMC; 189 however, acidosis dilates pig coronary arterioles through a glibenclamide-sensitive mechanism. 190,191 Lactate, a metabolite produced during myocardial ischemia, activates BKCa channels in pig CASMC. 192
Hypertension, cardiac hypertrophy and cardiomyopathy
Aldosterone, a mineralocorticoid with relevance to hypertension, reduces BKCa channel expression and iberiotoxin-sensitive relaxations in murine coronary arteries. 193 BKCa channels compensate for a loss of endothelial function in coronary arteries of rats made hypertensive with ouabain. 194 Genetic ablation of KATP channels causes hypertension and coronary vasospasm in mice. 96,98 BKCa current is reduced in CASMC from rats and rabbits with left ventricular hypertrophy, attributable to a reduction in single channel amplitude and Ca2+/voltage sensitivity. 195 KATP channels are normally not involved in coronary metabolic vasodilation; however, KATP channels become important for exercise-induced hyperemia in dogs with left ventricular hypertrophy. 196 KV current is increased in CASMC from rats with right ventricular hypertrophy due to pulmonary hypertension. 188 BKCa channels compensate for the loss of NO-dependent coronary artery relaxation in cardiomyopathic hamsters. 197 The role of KATP channels in coronary vascular regulation changes in the remodeled left ventricle of swine with a 2–3-week-old myocardial infarction. 198 Specifically, in normal pigs, KATP channels are not required for exercise-induced coronary vasodilation; however, following an infarction, KATP channels are responsible for a significant portion of exercise hyperemia.
Aging and reactive oxygen species
Aging is the main risk factor for coronary heart disease and spontaneous contractile activity is higher in coronary arteries of the aged, consistent with the idea that reduced K+ channel expression or activity is related to coronary vasospasm. 17 Aging reduces BKCa channel current and pore-forming α subunit protein in CASMC from rats and humans. 199 Similarly, the expression of the regulatory β1 subunit of BKCa channels is reduced in CASMC from aged rats. 200 Menopause, part of the aging process in women, reduces estrogen levels; this may also influence BKCa channel activity and coronary artery reactivity. Estrogen increases BKCa channel activity in human CASMC and iberiotoxin inhibits estrogen-induced coronary artery relaxation. 201,202 Reactive oxygen species are signaling molecules involved in normal coronary physiology as well as pathophysiology. H2O2 relaxes pig coronary arteries and arterioles via BKCa channels 85,203 and dilates the canine coronary circulation through the activation of KV channels. 111,146 Peroxynitrite, the product of a reaction between NO and superoxide anion, inhibits BKCa channels in human CASMC; superoxide alone was without effect. 204 Peroxynitrite also inhibits KV current in rat CASMC, 173 as does superoxide, which inhibits cAMP-mediated dilation by reducing KV channel function in the coronary arteries of diabetic rats. 205
Summary of K+ channels in coronary blood flow
BKCa, KATP and KV channels are important in various aspects of coronary vascular regulation. KATP and KV channels are important in determining basal coronary microvascular tone, whereas BKCa channels do not appear to contribute to resting coronary flow. BKCa and KV channels participate in endothelium-dependent coronary vasodilation, while KATP channels appear to play no role. All three channels seem to contribute in some way to ischemic coronary vasodilation. Only KV channels are mediators of coronary metabolic vasodilation, as inhibitors of KATP and BKCa channels have no effect on coronary blood flow elicited by metabolic demands. Diabetic conditions impair the function of BKCa, KATP and KV channels in CASMC. The expression and activity of CASMC BKCa, KATP and KV are altered by hypercholesterolemia, ischemia, hypoxia, hypertension, myocardial hypertrophy and cardiomyopathy. Aging decreases the function and expression of BKCa channels, while reactive oxygen species have physiological and pathophysiological effects on the activity of BKCa and KV channels. Overall, KV channels may have the most central role in regulating coronary blood flow, as these channels are involved in resting vascular tone, as well as endothelium-dependent, ischemic and metabolic vasodilation.
Footnotes
Acknowledgements
This work was supported by NIH HL-67804 and Research Funds Development Grant from West Virginia University.