
Abstract
This study was conducted to explore the effects of epigallocatechin-3-gallate (EGCG) on cognitive performances in psychological stress rats. An animal model of psychological stress was developed by restraint stress for three weeks. Male Wistar rats were randomly assigned to four groups as follows: normal control group, stress control group and two stress groups with green tea polyphenols (GTPs) and EGCG modulation, respectively. The changes of behavioral performances of rats were examined by the open-field test and step-through test. Results showed that behavioral performances of stress control group were changed abnormally, and they were improved in GTPs and EGCG modulation groups. In addition, plasma levels of cortisol, dopamine, norepinephrine, 5-hydroxytryptamine, interleukin-6 and interleukin-2 were detected. Stress control group had increased contents of cortisol, interleukin-6 and interleukin-2, and meanwhile had declined levels of 5-hydroxytryptamine and catecholamines. These changes in GTPs and EGCG modulation groups were similar to that of the normal control group. The expressions of metallothioneins in the hippocampus were detected by reverse transcription polymerase chain reaction. In contrast with the normal control group, their expressions in all the three stress groups were enhanced clearly. The results suggested that GTPs and EGCG modulation could improve the cognitive impairments induced by psychological stress. The related mechanisms may be involved with the changes of catecholamines, 5-hydroxytryptamine, cytokines and expressions of metallothioneins.
Introduction
Green tea polyphenols (GTPs) are the most active constituents in green tea. Studies showed that GTPs could pass through the blood–brain barrier and thus improve the brain's functions. 1,2 GTPs consists of five isoforms, namely epigallocatechin-3-gallate (EGCG), epigallocatechin (EGC), epicatechin gallate (ECG), catechin (CA) and epicatechin (EC). EGCG has been identified as the most active constituents of GTPs. Many studies showed that psychological stress can induce a series of dysfunctions of the central nervous system, such as cognitive impairment, anxiety, amnesia and insomnia. The development of many psychosomatic diseases has also been correlated with psychological stress, for example, tumor, hypertension and diabetes. 3–5 Therefore, more attention was paid to studies of protecting effects of natural potential ingredients on stress-induced injury. Experimental data and epidemiological investigations suggests that polyphenols, especially those derived from green tea and blueberry, can ameliorate age-related cognitive retrogression, and show neuroprotective effects against Parkinson's disease, Alzheimer's disease and cerebral ischemical reperfusion injury. Our previous studies indicated that psychological stress could impair behavioral performances, affect the changes of stress hormones, neurotransmitters, stress-related proteins and some cytokines, ultimately resulting in tissue injury, 6–8 and GTP modulation could improve cognitive impairments induced by psychological stress in rats. 9 To explore the further effects of GTP isoforms on psychological stressed rats, we conducted this study.
Materials and methods
Agents and chemicals
Green tea extract containing more than 99% of GTPs (80% CA and 60% EGCG) and EGCG (purity >98% by high-performance liquid chromatography) were provided by Chengdu Purification Technology Development Co Ltd (Chengdu, Sichuan Province, China). GTPs and EGCG were dissolved in ethanol, and were given to experimental animals by intragastric administration at a dose of 0.1 g/d. The control groups were given equal ethanol at the same time.
Animals
Male Wistar rats weighing 140–160 g were single-housed in cages with access to food and water. Temperature conditions in the animal facility were maintained at 23 ± 1°C. Animals were maintained on a 12:12-h light:dark cycle. After acclimating to laboratory conditions, the rats were divided at random into four groups of 10 animals each: normal control group (CT), stress control group (CS) and two stress groups with GTPs and EGCG modulation, respectively (GS and ES). The rats received humane care throughout the experiment according to the guidelines of the National Institute for Environmental Studies. All efforts were made to minimize both the number of animals used and their suffering.
Psychological stress model
According to the procedure described previously, 10,11 the animal model of psychological stress was developed by restraint for three weeks. Iron cylinder tubes were used to induce restraint stress. The tubes were 8.5 in long and 2.0 in in diameter. Each tube was covered on one end with a Plexiglas cover, which had a small air hole (0.2 in in diameter) in it. These tubes fit closely to the body size of the animals and inhibited movement. Each rat received restraint for six hours every day.
Examination of cognitive performances
Open-field test
The open field was a 100 cm × 100 cm × 50 cm dark gray Plexiglas arena whose floor was divided by black lines into 25 equal squares. The animals were gently placed individually into the center (the central square) of the open field, facing the same wall in all of the tests. They were allowed to explore the arena for 20 min on the first testing day and for 10 min the next day. On the third day, they were exposed once more to the empty open field for five minutes. During this time, the following variables were measured to assess exploration of the empty open field: latency (s), the number of squares crossed with the four paws, frequencies of rearing and modification. 12 The testing room was dimly illuminated with indirect white lighting.
Step-through test
The step-through apparatus consists of two chambers, and between them there is a hole that the animal can pass through. The step-through test was used to assess an animal's light–dark passive avoidance response. As it was described previously, 13 a single acquisition (training) trial was performed prior to the test. The trial consisted of a 30 s adaptation in one chamber that was dark at first, followed by the onset of a bright light. To avoid the light, the rat crossed over into the darkened adjacent chamber and received a mild 0.8 mA footshock for 1 s. The time to crossover from the light chamber to the darkened chamber (latency) was recorded with a maximum trial length of 180 s. The rats were tested for recall 24 h after the acquisition period to determine whether they had been trained to criterion. The criterion of rat was remaining in the light chamber for 180 s to avoid the footshock. Any rat that did not meet the criterion was retrained. The latency and number of footshocks were used to measure the ability of rats to remember adverse stimuli.
Determination of plasma cortisol, catecholamines, 5-hydroxytryptamine, interleukin-6 and interleukin-2
Cortisol
Plasma cortisol concentration was measured using the Cortisol Radioimmunoassay kit (Beifang Bio-tech, Beijing, China) according to the manufacturer's instructions.
Catecholamines
The levels of catecholamines were determined according to the method described previously.
14
The samples and phosphate buffer were mixed, and then the tubes were centrifuged at 1500
Norepinephrine was estimated by heating the solution for two minutes in a boiling water bath. The tubes were cooled and the fluorescence of norepinephrine was read in a Shimatzu spectrophotometer (Model RF-500) with excitation and emission wavelengths of 385 and 485 nm, respectively, with a slit width of 10/10 nm.
After the estimation of norepinephrine, the same solution was again heated for five minutes in a boiling water bath. The tubes were then cooled and the fluorescence of dopamine was read with excitation and emission wavelengths of 320 and 370 nm, respectively, with a slit width of 10/10 nm.
5-Hydroxytryptamine
Plasma 5-hydroxytryptamine (5-HT) level was measured by 5-HT enzyme-linked immunosorbent assay (ELISA) kit (Shanghai Shiruike Bio-tech, Shanghai, China) according to the manufacturer's instructions. This assay employs the quantitative sandwich enzyme immunoassay technique. A specific monoclonal antibody to 5-HT has been precoated onto a microplate. Standards and samples were pipetted into the wells and any 5-HT present is bound by the immobilized antibody. An enzyme-linked specific monoclonal antibody to 5-HT was added to the wells. Following a wash to remove any unbound antibody–enzyme reagent, a substrate solution was added to the wells and color develops in proportion to the amount of 5-HT bound in the initial step. The color development is stopped and the intensity of the color is measured.
Interleukin-6 and interleukin-2
Plasma levels of interleukin-6 and interleukin-2 were quantified with TPI Rat interleukin-6 (interleukin-2) ELISA kit (TPI Inc, Lynnwood, WA, USA) according to the manufacturer's instructions.
RT-PCR to test the expressions of metallothioneins
Total RNA was isolated from the hippocampus using a Uniq-10 column total RNA preparation kit (Sangon, Shanghai, China). Purity was confirmed by the A260/A280 ratio. The expressions of metallothionein-1 (MT-1) mRNA and metallothionein-3 (MT-3) mRNA were determined by a semiquantitative reverse transcription polymerase chain reaction (RT-PCR). Gene-specific primers of MT-1 and MT-3 (MT-1: forward, 5′-aca ccg ttg ctc cag att cac-3′; reverse, 3′-cgg agc ctg ttc acg tg-5′; MT-3: forward, 5′-gtt cct gca cct gct cgg ac-3′; reverse, 3′-cgg ata cac tta tca cga cgc-5′) were used with the schedule as follows: 50°C 30 min for RT; 94°C two minutes for deactivation of RTase; 94°C 30 s, 60°C 30 s and 72°C 90 s for 35 cycles; 72°C seven minutes for extension. To ensure that equal amounts of reverse-transcribed RNA were added to the PCR reaction, a parallel amplification of glyceraldehydes-3-phosphate-dehydrogenase (GAPDH) mRNA was performed as an internal reference (GAPDH gene-specific primers: forward, 5′-gcc atc aac gac ccc ttc at-3′; reverse, 3′-tct tcc acc act tcg tcc gc-5′). The ratio of MT-1 (MT-3) mRNA/GAPDH mRNA intensities was used to evaluate the relative levels of mRNA induction.
Statistical analysis
The statistical analysis was performed using SPSS 10.0 software. The results were expressed as mean ± standard deviation. All data were analyzed statistically by one-way analysis of variance, followed by Newman–Keuls test (q-test). Differences with P < 0.05 were considered statistically significant.
Results
Changes of behaviors in the open-field test of rats
Compared with the CT group, the latency and crossing frequency of the CS group in the open-field test showed obvious differences. There were no remarkable differences in the GS and ES groups in contrast to that of the CT group (Table 1).
Behavioral performances of rats in the open-field test*
*Wistar rats were divided into four groups (n = 10 for each): normal control group (CT), stress control group (CS) and two stress groups with green tea polyphenol (GTP) and epigallocatechin-3-gallate (EGCG) modulation, respectively (GS and ES). GTPs and EGCG were given to animals by intragastric administration at a dose of 0.1 g/d. The control groups were given equal ethanol at the same time. All stress groups received restraint for three weeks. At the fourth week, the performances of rats in the open-field were examined. The results were mean ± standard deviation determined by one-way analysis of variance
† P < 0.05 versus CT
Performances in step-through test of rats
The performance of the CS group in the step-through test changed significantly in contrast to that of the CT group (Table 2, P < 0.05). Compared with the CT group, the latencies of the GS and ES groups just had a tendency to increase and had no statistic differences. Moreover, the shock frequencies of GS and ES groups were decreased obviously in contrast to that of the CS group (P < 0.05).
Behavioral performances of rats in step-through test*
*Wistar rats were divided into four groups (n = 10 for each): normal control group (CT), stress control group (CS) and two stress groups with green tea polyphenols (GTPs) and epigallocatechin-3-gallate (EGCG) modulation, respectively (GS and ES). GTPs and EGCG were given to animals by intragastric administration at a dose of 0.1 g/d. The control groups were given equal ethanol at the same time. All stress groups received restraint for three weeks. At the fourth week, the performances of rats in the step-through test were examined. The results were mean ± standard deviation determined by one-way analysis of variance
† P < 0.05 versus CT
‡ P < 0.05 versus CS
Concentrations of plasma cortisol
Compared with the normal control group, the three stress groups all had upregulated levels of plasma cortisol (P < 0.05). Furthermore, the concentrations of plasma cortisol in the GS and ES groups were lowered than that of the CS group (P < 0.05, Figure 1).
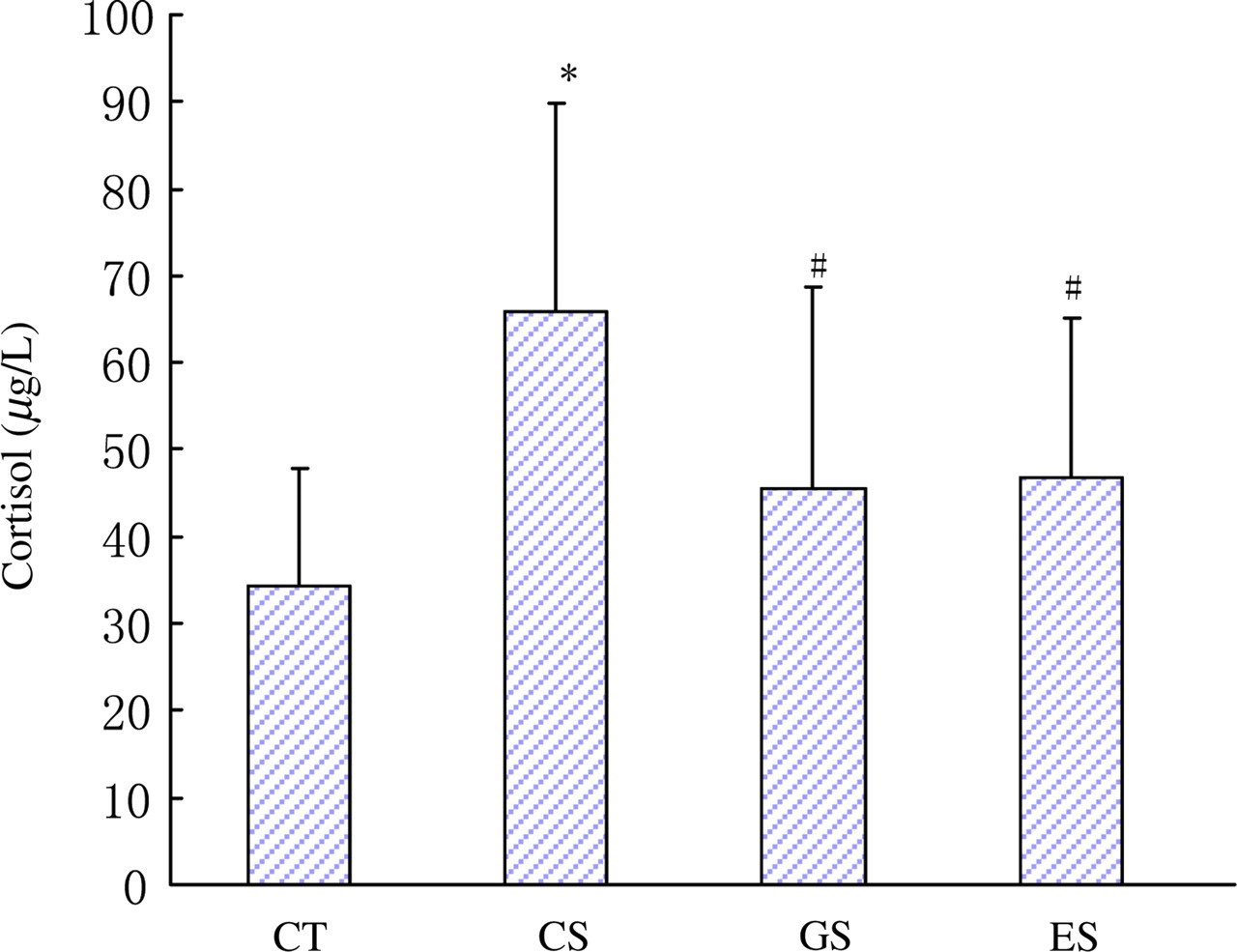
Levels of plasma cortisol in rats. Wistar rats were divided into four groups (n = 10 for each): CT, CS and two stress groups with GTP and EGCG modulation, respectively (GS and ES). GTPs and EGCG were given to animals by intragastric administration at a dose of 0.1 g/d. The control groups were given equal ethanol at the same time. All stress groups received restraint for three weeks. At the end of test, rats were sacrificed and the levels of plasma cortisol were measured. The results were mean ± SD determined by ANOVA. *P < 0.05 versus normal control group; # P < 0.05 versus stress control group. CT, normal control group; CS, stress control group; GTP, green tea polyphenol; EGCG, epigallocatechin-3-gallate; SD, standard deviation; ANOVA, one-way analysis of variance (A color version of this figure is available in the online journal)
Concentrations of plasma catecholamines, 5-HT, interleukin-6 and interleukin-2
Plasma levels of catecholamines and 5-HT were lowered remarkably in the CS group (Figures 2 and 3, P < 0.05). However, the concentration of plasma 5-HT in the GS group was increased in contrast to that of the CS group. Compared with the CT group, plasma levels of norepinephrine and dopamine in the ES group had a potency to decrease, but had no statistic difference. The concentrations of plasma interleukin-6 and interleukin-2 in all the three stress groups were increased significantly compared with the CT group (Figure 4, P < 0.05).
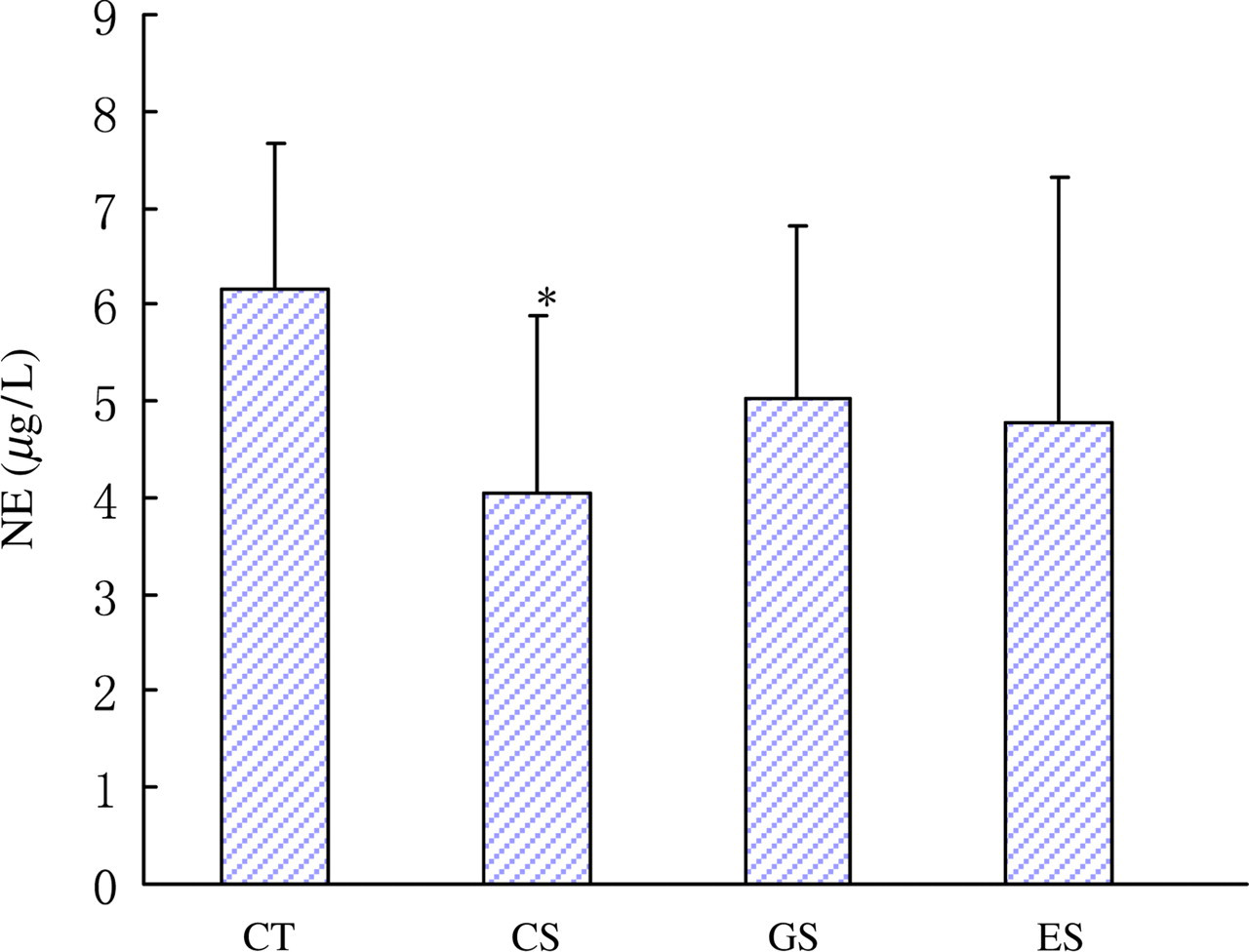
Levels of plasma NE in rats. Wistar rats were divided into four groups (n = 10 for each): CT, CS and two stress groups with GTP and EGCG modulation, respectively (GS and ES). GTPs and EGCG were given to animals by intragastric administration at a dose of 0.1 g/d. The control groups were given equal ethanol at the same time. All stress groups received restraint for three weeks. At the end of test, rats were sacrificed and the levels of plasma NE were measured. The results were mean ± SD determined by one-way ANOVA. *P < 0.05 versus normal control group. NE, norepinephrine; CT, normal control group; CS, stress control group; GTP, green tea polyphenol; EGCG, epigallocatechin-3-gallate; SD, standard deviation; ANOVA, one-way analysis of variance (A color version of this figure is available in the online journal)
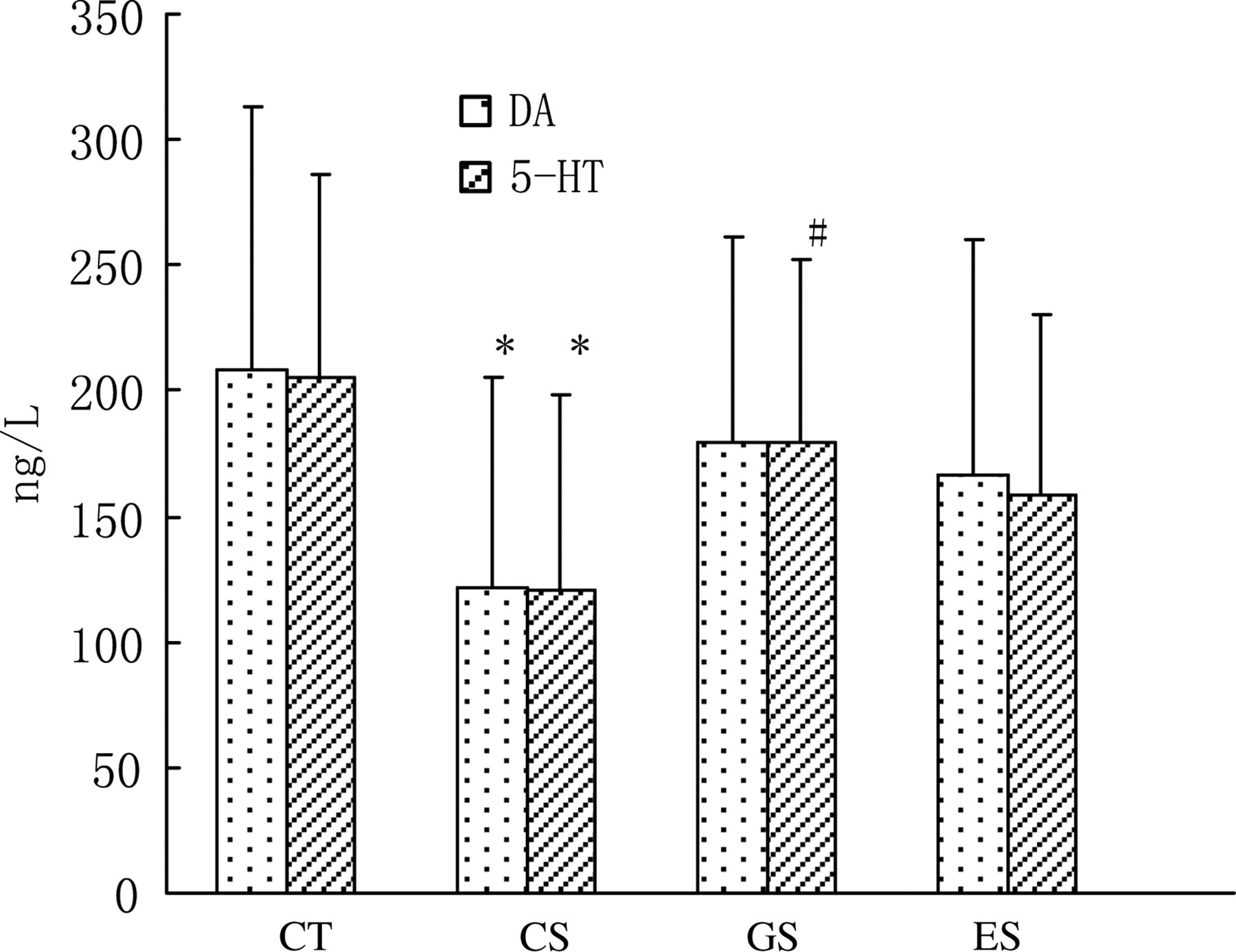
Levels of plasma dopamine and 5-HT in rats. Wistar rats were divided into four groups (n = 10 for each): CT, CS and two stress groups with GTP and EGCG modulation, respectively (GS and ES). GTPs and EGCG were given to animals by intragastric administration at a dose of 0.1 g/d. The control groups were given equal ethanol at the same time. All stress groups received restraint for three weeks. At the end of test, rats were sacrificed and the levels of plasma dopamine and 5-hydroxytryptamine were measured. The results were mean ± SD determined by one-way ANOVA. *P < 0.05 versus normal control group; # P < 0.05 versus stress control group. CT, normal control group; CS, stress control group; GTP, green tea polyphenol; EGCG, epigallocatechin-3-gallate; SD, standard deviation; ANOVA, one-way analysis of variance; 5-HT, 5-hydroxytryptamine
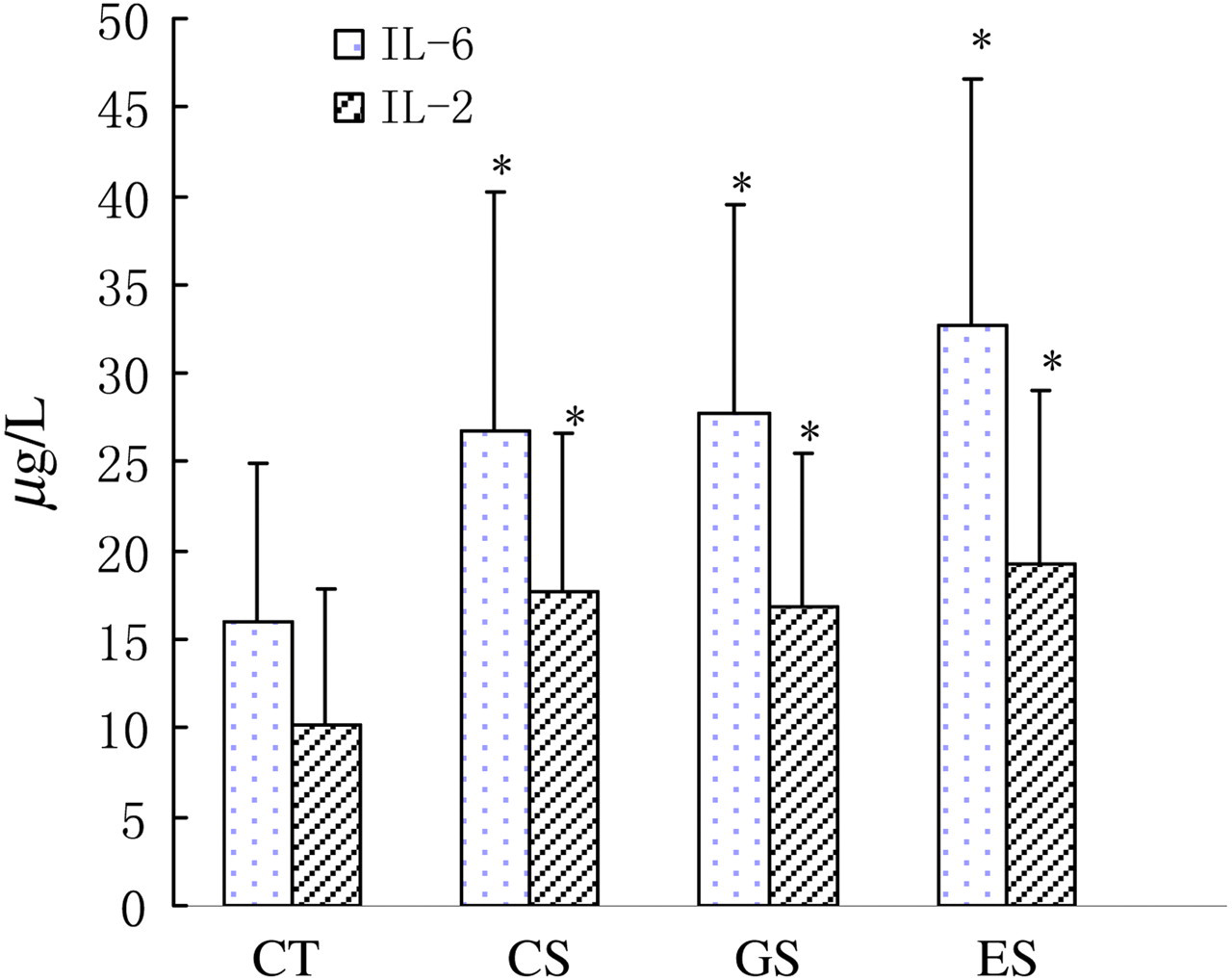
Levels of plasma interleukin (IL)-6 and IL-2 in rats. Wistar rats were divided into four groups (n = 10 for each): CT, CS and two stress groups with GTP and EGCG modulation, respectively (GS and ES). GTPs and EGCG were given to animals by intragastric administration at a dose of 0.1 g/d. The control groups were given equal ethanol at the same time. All stress groups received restraint for three weeks. At the end of test, rats were sacrificed and the levels of plasma IL-6 and IL-2 were measured. The results were mean ± SD determined by ANOVA. *P < 0.05 vs. normal control group. IL, interleukin; CT, normal control group; CS, stress control group; GTP, green tea polyphenol; EGCG, epigallocatechin-3-gallate; SD, standard deviation; ANOVA, one-way analysis of variance
Expressions of MTs in the hippocampus
In this study, the productions of MT-1 mRNA and MT-3 mRNA in the hippocampus of rats were determined by RT-PCR. The results showed that their expressions in all stress groups were increased obviously in contrast to that of the CT group (Figure 5).
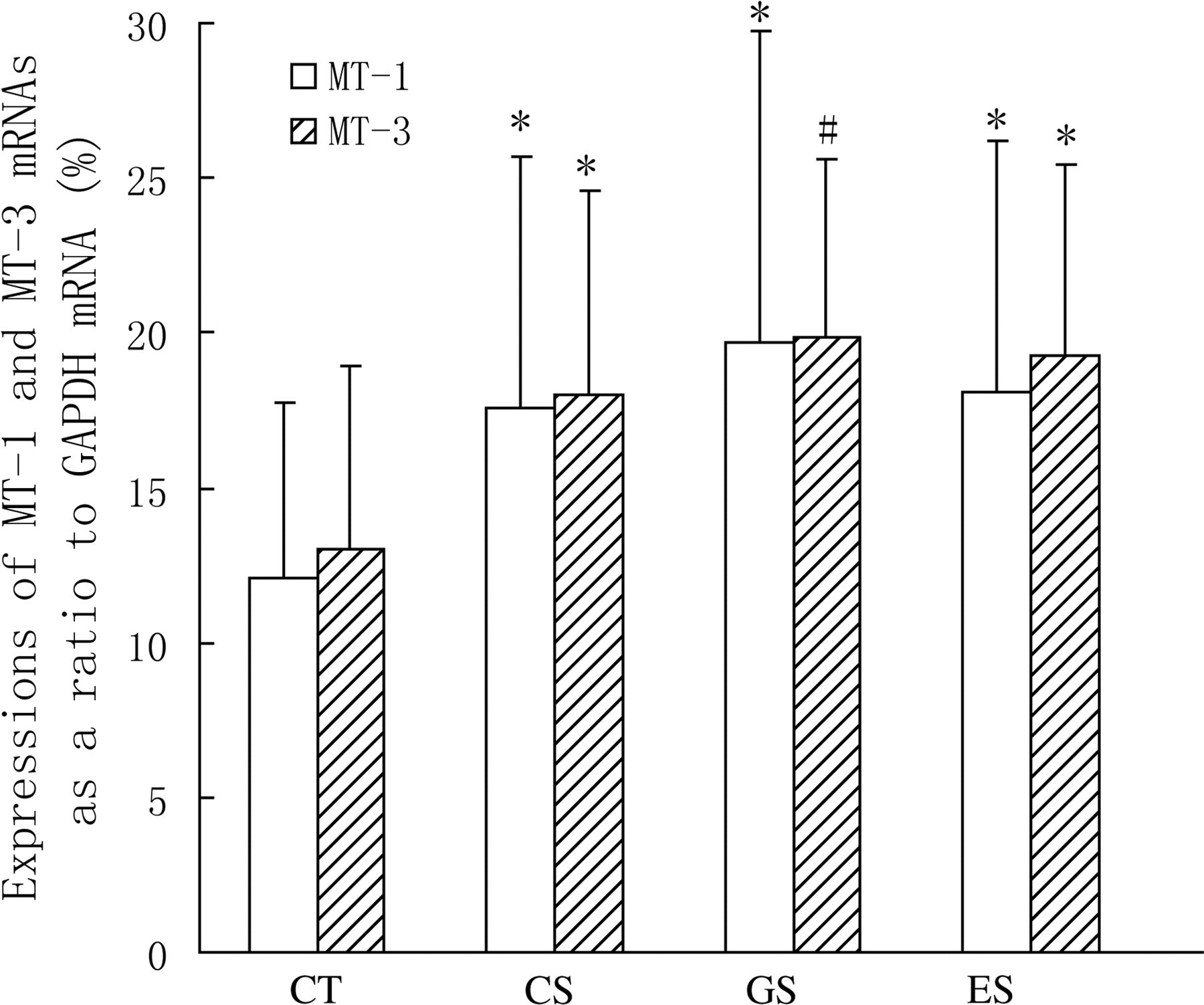
Expressions of hippocampal MT-1 and MT-3 mRNAs in rats. Wistar rats were divided into four groups (n = 10 for each): CT, CS and two stress groups with GTP and EGCG modulation, respectively (GS and ES). GTPs and EGCG were given to animals by intragastric administration at a dose of 0.1 g/d. The control groups were given equal ethanol at the same time. All stress groups received restraint for three weeks. At the end of test, rats were sacrificed and the expressions of MT-1 mRNA and MT-3 mRNA were measured. The results were mean ± SD determined by one-way ANOVA. *P < 0.05 versus normal control group; # P < 0.01 versus normal control group. MT, metallothionein; CT, normal control group; CS, stress control group; GTP, green tea polyphenol; EGCG, epigallocatechin-3-gallate; SD, standard deviation; ANOVA, one-way analysis of variance
Discussion
In recent years, there has been considerable interest in investigating the neuroprotective effects of phenolic compounds from different botanical sources. Green tea is currently considered a source of dietary constituents endowed with biological and pharmacological activities relevant to human health. Indeed, tea consumption is inversely correlated with the incidence of dementia, Alzheimer's and Parkinson's diseases. Green tea derivatives are very attractive drug candidates because they are naturally occurring phytopharmaceuticals with known neuroprotective effects. In particular, as the most effective ingredient in GTPs, EGCG has been shown to exert neuroprotective/neurorescue activities in a wide array of cellular and animal models of neurological disorders, 15,16 and to be safe for humans when tested in clinical studies. 17 Moreover, EGCG also could reduce oxidative stress and neurotoxicity in different model systems of Alzheimer's and Parkinson's disease and modulate the expression of cell survival and cell death genes; 18,19 EGCG potently modulates early steps in the aggregation process of an amyloidogenic polyglutamine-containing protein. 20 So EGCG may have considerable potential as a drug candidate for the development of treatments for Huntington's disease and for protein misfolding and amyloid diseases in general. Some studies demonstrated that EGCG also protects against ischemia- and reperfusion-induced injury in the brain. 21,22 A recent study showed EGCG could suppress matrix metalloproteinase-9 activation and reduce the development of delayed neuronal death after transient global cerebral ischemia in C57BL/6 mouse brain. 23 Long-term administration of a preparation of green tea CAs (polyphenol E) or EGCG has been demonstrated to improve spatial cognition and learning ability in rats. 24 The present study showed that EGCG also could ameliorate the cognitive impairments induced by psychological stress.
It was manifested that two neuroendocrinology systems were involved in the regulation of stress response, namely the hypothalamic–pituitary–adrenal (HPA) axis and sympathetic nervous system (SNS). Psychological stress activated HPA which resulted in the increasing release of glucocorticoids (GCs). Many studies inferred that endogenic GCs played an important role in pathological impairments induced by stress. 25,26 As an essential stress hormone, GC levels can show the body's stress intensity. 27 The present study demonstrated that plasma levels of cortisol were increased dramatically in stress rats. This suggested that the animal model of psychological stress was developed successfully. Furthermore, the present study showed that the performances of psychological stress rats in the open-field test and step-through test were all changed abnormally. Also, appropriate doses of GTP and EGCG modulations could improve the cognitive impairments induced by stress. This may be related with the declining GC levels and the increasing contents of catecholamines and 5-HT, as psychological stress also activated SNS, and then catecholamines were released from the sympathetic nerve terminal and the adrenal medulla. Catecholamines consist of epinephrine, norepinephrine and dopamine; they are involved with the modulation of the body's cognition, awareness, attention, emotional state and other psychoactivities. The physiological functions of dopaminergic neurons were enhanced when exposed to psychological stress. If the intensity and duration of stress overtopped the body's coping capacity, the cognitive impairments appeared. The behavioral disturbances induced by psychological stress may be related with the declines of norepinephrine and dopamine. 28,29
Central serotonin (5-HT) dysregulation contributes to the susceptibility to mental disorders, including depression, anxiety, post-traumatic stress disorder, and learning and memory deficits. 5-HT-deficient mice exhibited no gross abnormality in brain structures and had normal locomotor activity, but their spatial learning memory was impaired. Serotonergic neurotransmission has been reported to play a prominent role in a different kind of inhibition, namely, the capacity to withhold over time a response to affectively charged stimuli and to delay gratification, 30–33 and thus be more implicated in cognitive – rather than motor – inhibition. The present study showed that plasma 5-HT was decreased in psychological stress rats that had poor cognitive performances. GTP and EGCG modulation could upregulate the levels of 5-HT, and had improved behaviors compared with stress rats.
Psychological stress can influence the regulation of many genes' expressions, such as stress-related proteins and some signal transducers. In the present study, we detected the expressions of MTs in the hippocampus. The results showed that their mRNA expressions were increased obviously in stress rats. MT-3 and MT-1 are important stress proteins in the brain, especially MT-3, which has tissue specificity, and is only expressed in the brain. Our previous study showed that stress could induce the expressions of MT-1 and MT-3 in the hippocampus. 6 The present study also suggested increasing productions of MTs in stress and GTP modulation rats. The enhanced expression of MTs may be helpful to the body's adaptability to psychological stress.
In addition, the changes of plasma interleukin-6 and interleukin-2 were also examined in this study. The results showed that psychological stress aroused the secretions of interleukin-6 and interleukin-2. Psychological stress provoked a series of regulations of neuroendocrine and immune activities. Interleukin-6 is a pleiotropic cytokine with a wide range of biological activities, including immunoregulation and mediation of acute-phase responses. 34 In the central nervous system, upregulation of transgenic excretion of interleukin-6 induced the changes of many genes' expression, including stress-related proteins, such as MTs and hot-shock proteins. 35 The former study demonstrated that GTPs could enhance humoral immunity in vivo by increasing the expressions of interleukin-2 and interleukin-2 mRNA, accordingly to promote the proliferation of T-leukomonocyte. The increasing productions of interleukin-6 and interleukin-2 could motivate the related alternations so as to enhance the body's stress reaction.
In summary, the present study suggests that moderate EGCG modulation plays an important role in improving cognitive performances in psychological stress rats. The related mechanisms may involve the secretion of stress hormones, production of catecholamines and 5-HT, regulation of cytokines and the expressions of stress-related proteins.
Footnotes
ACKNOWLEDGEMENTS
This work was supported by the grant ‘Effects of green tea polyphenols on body's adaptation to psychological stress’ (No. 30500406) from the Nature Science Fund of China (NSFC). The corresponding author verified that all individuals who made contributions to this study were included.