
Abstract
Various studies have been performed to find out novel treatment strategies for acute necrotizing pancreatitis (ANP). Inhibition of poly(ADP-ribose) polymerase (PARP) is shown to reduce inflammation in several pathological conditions. We aimed to evaluate the efficacy of benzamide, a PARP inhibitor, in an experimental model of ANP. Thirty Sprague–Dawley rats were divided into three groups: sham-operated, ANP and ANP + benzamide groups. All groups except the sham-operated group were subjected to the ANP procedure, induced by infusing of 1 mL/kg of 3% sodium taurocholate into the common biliopancreatic duct. The ANP + benzamide group received 100 mg/kg/day benzamide intraperitoneally for a total of three days after induction of pancreatitis. The surviving animals were killed at the fourth day and the pancreas was harvested for biochemical, microbiological and histological analysis. Blood samples were also obtained from the animals. In the ANP group, a significant increase was observed in concentrations of serum amylase and neopterin and tissue oxidative stress indices (malondialdehyde, superoxide dismutase and glutathione peroxidase). Almost all of these changes were found to be reversed to near their normal values in the ANP + benzamide group. Histological injury scores were significantly higher in the ANP group than in the sham group (P < 0.05, ANP versus sham), and were significantly lower in the ANP + benzamide group than in the ANP group (P < 0.05, ANP + benzamide versus ANP). Evaluation of bacterial translocation identified significantly fewer infected sites in the ANP + benzamide group than in the ANP animals (P < 0.01). We observed that inhibition of PARP with benzamide reduced the severity, the mortality, the bacterial translocation rates and the neopterin concentrations in an experimental ANP model in rats. These findings suggest that it may be possible to improve the outcome of ANP by using PARP inhibitors.
Introduction
Acute necrotizing pancreatitis (ANP) is a pathological condition with high morbidity and mortality. It is caused by a massive inflammatory reaction, which occurs as a result of an imbalance between the systemic release of pro-inflammatory and anti-inflammatory mediators in the ANP process, which eventually results in septic multiple organ failure. 1 In addition, reactive oxygen species (ROS), such as superoxide anions (·O2 −), hydroxyl radicals (·OH−), hydrogen peroxide (H2O2) and reactive nitrogen species (RNS), such as peroxynitrite (ONOO−), have been identified as potential important factors in the pathogenesis and progress of pancreatitis. 2,3 It is known that recruitment of activated neutrophils and monocytes into the pancreatic interstitium further exacerbates pancreatic damage via the generation of ROS/RNS. 4
Poly(ADP-ribose) polymerase-1 (PARP-1) (also known as poly[ADP-ribose] synthetase or poly[ADP-ribose] transferase) is a nuclear enzyme present in eukaryotic cells. 5 PARP-1 activation is triggered by the presence of nicks and breaks in the DNA strand. This damage can be induced by reactive molecules, such as superoxide, hydroxyl radical or peroxynitrite, which trigger the PARP-1 activation. 6 Rapid depletion of NAD+ due to PARP activation leads to cellular ATP depletion and functional alteration of the cell, and eventually necrotic-type cell death occurs. 7,8 To date, it has been shown that inhibition of PARP attenuates acute inflammation and tissue damage in animal models of various pathological conditions including pleuritis, lung injury, inflammatory bowel diseases and arthritis. 9–11 In our team's previous works, beneficial effects of PARP inhibition were shown in different experimental inflammatory models. 12,13
This study was designed to investigate whether benzamide, a PARP inhibitor, has an ameliorative effect on ANP injury via blockage of cellular ATP depletion by using an experimental model induced by infusing sodium taurocholate into the common biliopancreatic duct.
Materials and methods
Animals
Sprague–Dawley male rats (200–250 g) were used in our study. All animal procedures were approved by the Institutional Committee on the Care and Use of Animals of Gulhane Military Medical School (Issue; 08/31). Before the experiment, animals were fed standard rat chow and water ad libitum and housed in cages with controlled temperature and 12-h light/dark cycle for at least one week.
Experimental groups and treatment modalities
Thirty rats were randomly assigned to three groups containing 10 rats each: sham-operated, ANP, ANP + benzamide groups. The sham-operated group served as a negative control. This group underwent laparotomy with injection of the 1 mL/kg saline into the common biliopancreatic duct, and then the animals received saline (10 mL/kg) intraperitoneally every 12 h for three consecutive days. The ANP group, as positive control, was also injected with intraperitoneal saline with the same regimen. The treatment group, namely ANP + benzamide, received benzamide instead of saline at a daily dose of 100 mg/kg intraperitoneally. Four days after the induction of pancreatitis, all surviving animals were killed.
Induction of pancreatitis
All animals were fasted overnight and anesthetized by an intramuscular injection of 50 mg/kg ketamine (Ketalar; Parke Davis, Eczacibasi, Istanbul, Turkey) and 10 mg/kg xylazine (Rompun; Bayer AG, Leverkusen, Germany); all operations were performed at room temperature (24°C). Through a midline laparotomy, the common biliopancreatic duct was cannulated with a 28-gauge microfine catheter. An atraumatic microvascular clamp (Bulldog Artery Clamp, Harvard Apparatus, Holliston, MA, USA) was placed on the bile duct below the liver and another around the common biliopancreatic duct at its entry into the duodenum to avoid reflux of enteric contents into the duct. Then, 1 mL/kg of 3% sodium taurocholate (Sigma, St Louis, MO, USA) was slowly infused into the common biliopancreatic duct, and the infusion pressure was kept below 30 mmHg, as measured with a mercury manometer. 14 When the infusion was finished, the microvascular clamps were removed. Finally, the abdominal incision was closed, and the animals were returned to their cages to recover. Water and food were available ad libitum.
Sample collection
All surviving animals were killed on the fourth day of experiment via decapitation; the abdomen was opened and pancreatic tissue samples were harvested for analysis. Pancreatic tissue samples were divided into two pieces, one of them was fixed in 10% formalin solution for histological evaluation and the other was stored at −80°C deep freeze for the assay of antioxidant enzyme activity and tissue lipid peroxidation levels. In addition, samples from the pancreas, visceral peritoneum, mesenteric lymph nodes (MLN), liver and cecum were harvested to evaluate bacterial translocation. Blood samples were also obtained and some additional biochemical analyses were done on the serum samples.
Biochemical analysis
The frozen tissues were homogenized in phosphate buffer (pH 7.4) on an ice cube by using a homogenizator (Heidolph Diax 900; Heidolph Elektro GmbH, Kelhaim, Germany). The supernatant was used for the assay of tissular parameters. Initially, the protein content of tissue homogenates was measured by the method of Lowry using bovine serum albumin as the standard. 15
Lipid peroxidation levels were measured with the thiobarbituric acid (TBA) reaction using the method of Ohkawa et al. 16 This method was used to obtain a spectrophotometric measurement of the color produced during the reaction of TBA with malondialdehyde (MDA) at 535 nm. The calculated MDA concentrations were expressed as nmol/mg-protein.
Superoxide dismutase (SOD) activity was assayed using a modified nitroblue tetrazolium (NBT) method as previously described by Durak et al. 17 In brief, NBT was reduced to blue formazan by ·O2 −, which has a strong absorbance at 560 nm. One unit (U) of SOD is defined as the amount of protein that inhibits the rate of NBT reduction by 50%. The estimated SOD activity was expressed as units per gram of protein.
The glutathione peroxidase (GSH-Px) activity was measured using the method described by Paglia and Valentine 18 in which GSH-Px activity was coupled with the oxidation of NADPH by glutathione reductase. The oxidation of NADPH was spectrophotometrically followed up at 340 nm at 37°C. The absorbance at 340 nm was recorded for 5 min. The activity was the slope of the lines expressed as mmol of NADPH oxidized per minute. GSH-Px activity was expressed as U/g-protein.
Serum amylase, glucose, aspartate aminotransferase (AST), alanine aminotransferase (ALT), alkaline phosphatase (ALP), gamma-glutamyl transferase (GGT), total protein (TPRO) and albumin (ALB) concentrations were measured by an autoanalyzer using commercial kits and routine spectrophotometric techniques (Olympus AU-2700, Hamburg, Germany).
Serum neopterin (NP) concentrations were determined by a high pressure liquid chromatography (Agilent Technologies 1200 Series System, Santa Clara, CA, USA) system with a fluorescence detector as described previously.
19
In brief, to 0.4 mL serum or tissue supernatant, 0.1 mL 2 mmol/L trichloroacetic acid was added and the mixture was left in an ice bath for 10 min. The precipitated protein was removed by centrifugation at 2000
Histological analysis
Tissue specimens were fixed in formalin for 24 h, then embedded in paraffin and cut into 4 µm sections. Slides were stained with hematoxylin and eosin and examined under a light microscope. Each slide was evaluated by two expert investigators blinded to the experiment groups. The treatment protocol scored the tissues for edema, inflammatory infiltration, fat necrosis, parenchymal necrosis and hemorrhage in 20 fields. The scores for each histological parameter were summed up, with a maximum score of 27, as defined by Spormann et al. 20
Bacterial quantitative cultures
Samples from the pancreas, visceral peritoneum, MLN, liver and cecum were taken from each rat under sterile conditions. All samples were weighted and homogenized in cysteine broth immediately for quantitative and qualitative culture of aerobic bacteria. Homogenized samples were diluted serially and cultured onto MacConkey (Merck, Darmstadt, Germany) and 5% sheep blood agars (Merck). All plates were incubated for up to three days at 35–37°C. After incubation, Gram-negative bacteria were identified with the API-20GN system (BioMerieux Vitek, Hazelwood, MO, USA). Gram-positive bacteria were identified to the genus level by means of standard microbiological methods. Bacterial counts were expressed in colony-forming units per gram of tissue (CFU/g). Growth of more than 103 CFU/g of tissue was considered to indicate significant infection.
Statistical analysis
Results were expressed as median (min–max). Translocation incidence was evaluated by chi-square (χ 2) analyses and Fisher's exact test when an expected frequency was less than 5. In the other analyses, differences among the groups were analyzed by the Kruskal–Wallis test. Dual comparisons among groups with significant values were evaluated with the Mann–Whitney U-test; P < 0.05 were considered significant. All analyses were performed with the Statistical Package for the Social Sciences (SPSS) statistical program (Software version 11.0, SPSS Inc, Chicago, IL, USA).
Results
In this study, no complications occurred in the surgical technique. The courses of survival rate and weight for each group are shown in Figure 1. In the ANP group, four of 10 animals died by day 4: two of them died within the first 24 h, and the two others died by 48 and 72 h following induction of pancreatitis. In the ANP + benzamide group, two rats died in first 24 h. Additionally, there were significant differences at the fourth day in terms of the weight of rats. The mean weight of rats in the ANP group was significantly lower than those of sham-operated and ANP + benzamide groups (P < 0.05, ANP versus the other groups). In the ANP + benzamide group, the mean weights were significantly higher than ANP group, but those were still lower than in the sham-operated group (P < 0.05, ANP + benzamide versus ANP and sham-operated).
(a) Survival rates in all groups were shown as a Kaplan–Meier survival curves. The survival rate was significantly lower in the acute necrotizing pancreatitis (ANP) group than in the sham-operated group (log rank = 5.420, P = 0.020, ANP + benzamide group versus ANP group). There was no significant difference between the ANP + benzamide and the ANP group (log rank = 2.111, P = 0.146, ANP + benzamide group versus ANP group). (b) Mean weight of the rats in each group versus time displayed significant weight loss in the ANP group. The treatment with benzamide prevented mean weight loss in the ANP + benzamide group.*P < 0.05 statistically different from sham-operated group; **P < 0.05 statistically different from ANP group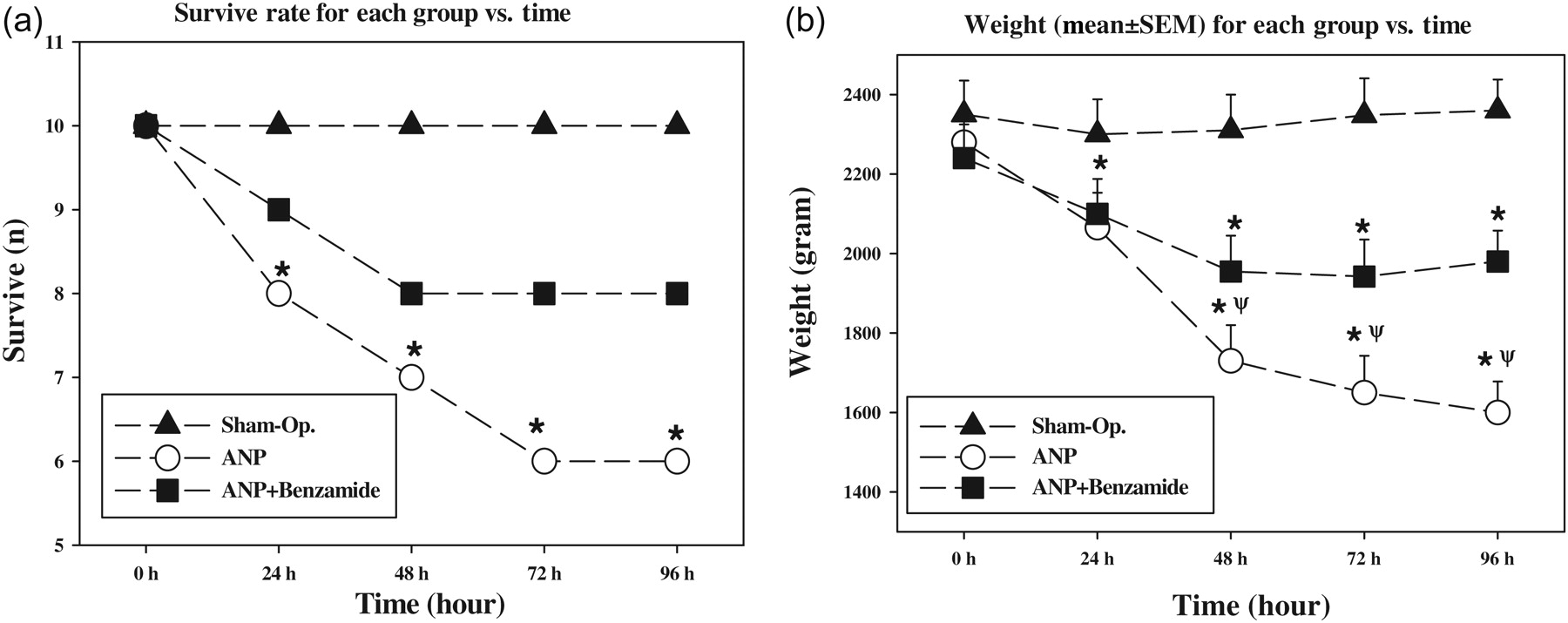
Biochemical results
Serum measurements
Biochemical values in serum
ANP, acute necrotizing pancreatitis; AST, aspartate aminotransferase; ALT, alanine aminotransferase; ALP, alkaline phosphatase; GGT gamma-glutamyl transferase
Data are expressed as median (min–max)
*P < 0.05 statistically different from sham-operated group
**P < 0.05 statistically different from ANP group
Glucose and protein values in serum
ANP, acute necrotizing pancreatitis; TPRO, total protein; ALB, albumin
Data are expressed as median (min–max)
*P < 0.05 statistically different from sham-operated group
**P < 0.05 statistically different from ANP group
Serum NP concentrations were significantly increased in the ANP group (P < 0.05, ANP versus sham group). The increased NP concentrations significantly decreased with benzamide treatment but remained still higher than the sham values (P < 0.05, ANP + benzamide group versus ANP and sham groups). The serum NP concentrations are shown in Figure 2a.
Serum neopterin concentrations and tissue oxidative stress indices. *P < 0.05 statistically different from sham-operated group; **P < 0.05 statistically different from ANP group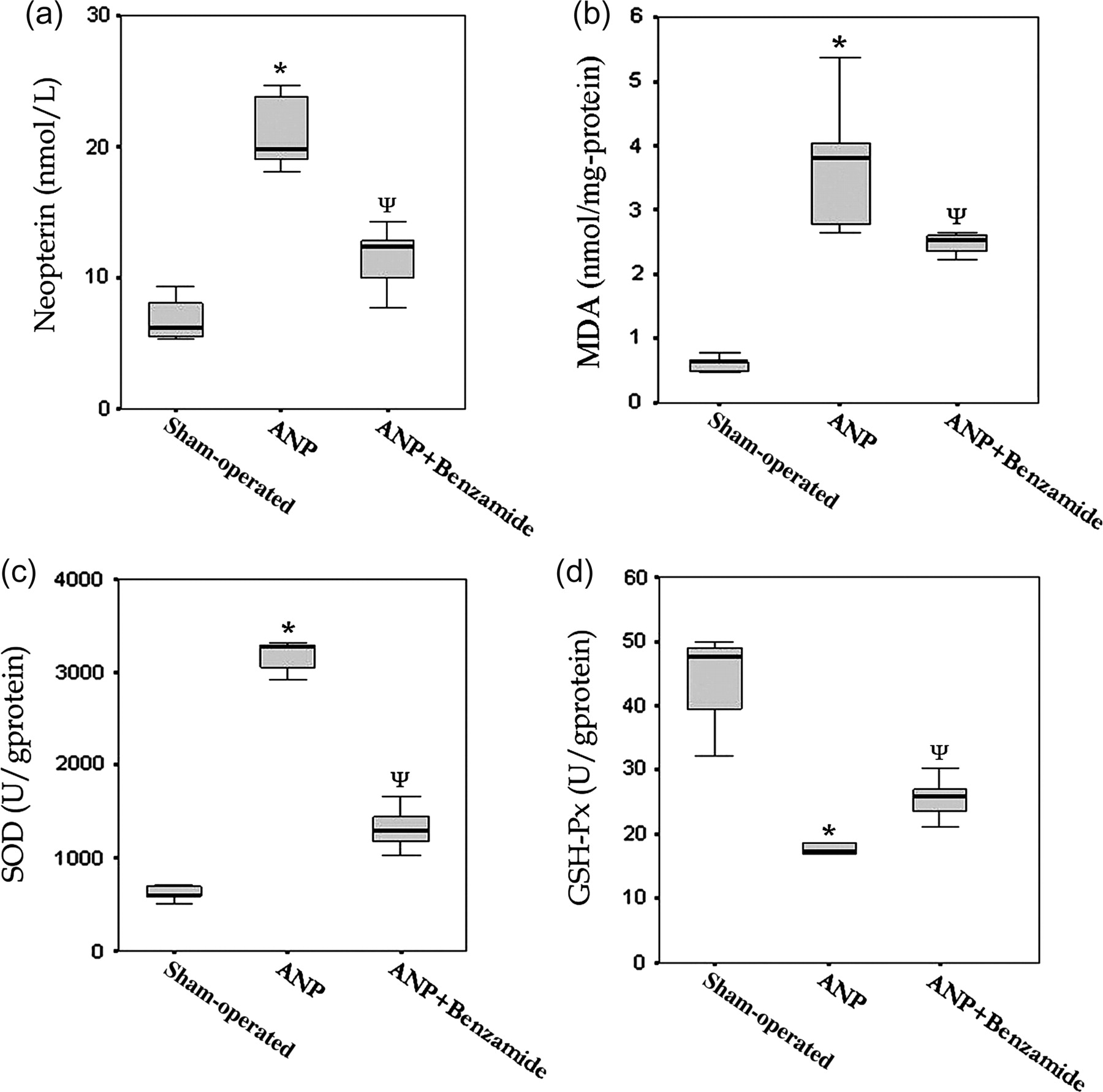
Oxidative stress indices
The pancreatic tissue MDA concentrations in the ANP group were significantly higher compared with other groups (P < 0.05, ANP versus sham and ANP + benzamide groups) reflecting a clear oxidative stress which led to lipid peroxidation. The tissue SOD activity was found to be increased, but GSH-Px decreased significantly in ANP animals (P < 0.05). Although benzamide treatment reversed all changes of these oxidative stress markers significantly, all values remained significantly different from those of the sham-operated animals (P < 0.05, ANP + benzamide versus ANP and sham groups for MDA, SOD and GSH-Px). The outcomes of these oxidative stress parameters are displayed in Figure 2b–d.
Histological evaluation
Histological examination revealed no evidence for necrotizing pancreatitis in the sham-operated group, while all animals in the ANP group showed a severe degree of pancreatitis with marked parenchymal necrosis, hemorrhage and leukocyte infiltration. In the benzamide-treated group, degrees of necrosis and leukocyte infiltration were lower than the ANP group (P < 0.05). Representative photomicrographs of the study groups are presented in Figure 3 and the detailed injury scores are shown in Table 3.
Representative photographs showing the histological changes in pancreatic tissues. Sham-operated group shows normal pancreatic structure. In the acute necrotizing pancreatitis (ANP) group, massive edema (E), parenchymal necrosis (N), pancreatic acinar regeneration (R) and leukocyte infiltration (black arrows) are seen. In the ANP + benzamide group, benzamide treatment normalized edema, parenchymal necrosis and leukocyte infiltration (hematoxylin and eosin, scale bars = 100 μm) (A color version of this figure is available in the online journal) Pathological scores ANP, acute necrotizing pancreatitis Data are expressed as median (min–max) *P < 0.05 statistically different from sham-operated group **P < 0.05 statistically different from ANP group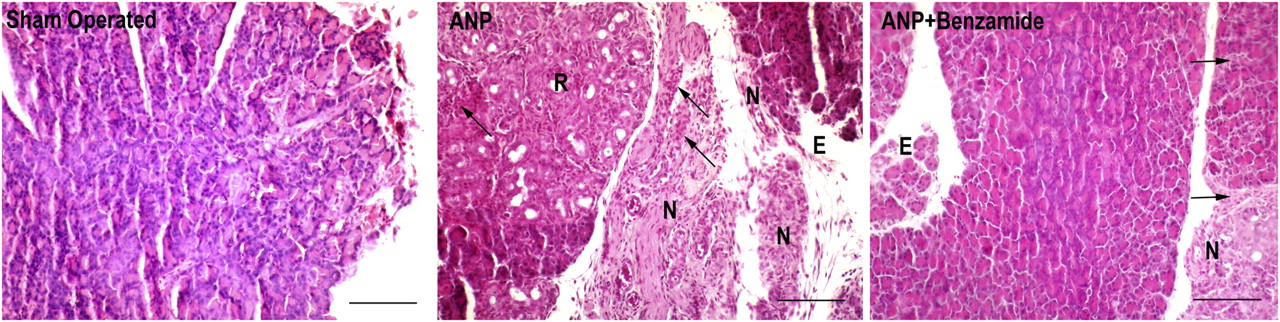
Bacterial translocation
Total incidence of bacterial translocation (evaluated by χ 2 analyses)
ANP, acute necrotizing pancreatitis; MLN, mesenteric lymph nodes
*P < 0.05 statistically different from sham-operated group
**P < 0.05 statistically different from ANP group
Discussion
In the current study, we aimed to test effects of PARP inhibition via benzamide administration on various pathological parameters, especially bacterial translocation rates and NP concentrations, in a rat model of ANP. The experiment showed that benzamide had a beneficial effect on pancreatic necrosis. Moreover, benzamide also had an ameliorating effect on oxidative stress indices of the pancreatic tissue in correlation with histological findings. Although oxidative stress is the initial step in the pathogenesis of pancreatic injury, overexpression of inducible nitric oxide synthase changes the nature of this stress from oxidative to nitrosative stress, because the combination of nitric oxide with superoxide results in peroxynitrite formation. Peroxynitrite then induces both apoptosis and necrosis of cells. 21,22 In this mechanism, activation of the DNA repair enzyme PARP-1, a member of PARP enzyme family, mediates ONOO−-induced necrosis. PARP-1 detects and signals DNA strand breaks induced by binding to DNA of a variety of genotoxic insults, including ionizing radiation, alkylating agents, oxidants and free radicals. 23–26
Upon binding to DNA, strand breaks occur, and PARP transfers ADP-ribose units from the respiratory coenzyme NAD+ to various nuclear proteins. From a physiological viewpoint, PARP-1 activity and poly(ADP-ribosyl)ation reactions are implicated in DNA repair processes, the maintenance of genomic stability, the regulation of gene transcription and DNA replication. An important function of PARP-1 is to allow DNA repair and cell recovery under conditions associated with a low level of DNA damage. In case of severe DNA injury, overactivation of PARP-1 depletes the cellular stores of NAD+, an essential co-factor in principal energy production mechanisms, including the glycolytic pathway, the tricarboxylic acid cycle and the mitochondrial electron transport chain. As a result, the loss of NAD+ leads to a marked reduction in the cellular pools of ATP, resulting in cellular dysfunction and death via the necrotic pathway. This is known as the ‘suicide hypothesis’ of PARP activation and seems to be a regulatory mechanism to eliminate cells after irreversible DNA injury. 21,22 A vast amount of experimental evidence has established that the PARP-1 pathway of cell death plays a pivotal role in tissue injury and organ dysfunction in numerous disease processes. 12,24,27,28
NP is a marker associated with cell-mediated immunity and produced in monocytes/macrophages via the stimulation of interferon-γ. It has been shown that NP concentrations not only provide knowledge about the state of cell-mediated immunity, but also allow monitoring of the disease progression. 29 Also, it has been shown that enhanced NP concentrations reflect the severity of the acute pancreatitis. 30 Another study reported that NP concentrations are higher in acute pancreatitis patients with poor prognosis and are more specific for predicting disease progression than levels of C-reactive protein. 31 A recent study by our team, testing another treatment modality in the same experimental model, also found enhanced NP concentrations in ANP-induced animals, which were in correlation with oxidative stress and histological injury. 32 Thus, in the current study, due to its promising reports as a biochemical parameter to diagnose and follow-up pancreatic inflammation, serum NP concentrations were measured. NP concentrations in animals receiving benzamide came close to values in sham group. The inhibition of PARP by benzamide improved pancreatic functions, as evidenced by the reduction in amylase and NP concentrations in animals with ANP (Table 1, Figure 2). By using sources of cellular energy, namely ATP, to repair injured tissue, it seems likely that treatment with benzamide prevented pancreatic necrosis and allowed the preservation of the cellular stores of NAD. This in turn would have led to decreased activation of monocytes/macrophages, and as a consequence, reduction in levels of NP.
Necrotic pancreatic tissue derived from animals that were treated with benzamide demonstrated a decrease in the extent of necrotic sites, restoration of acinus structure and inhibition of the neutrophil recruitment. A major requirement for the initiation of the cell survival and regeneration process is the repletion of cellular ATP levels. Therefore, the enhanced restoration of the injured pancreas in the setting of PARP inhibition may partly be due to the protection of pancreatic cells as evidence of decreased suicide hypothesis of PARP activation.
We evaluated the tissue oxidative stress marker (MDA) and the tissue antioxidant system (SOD and GSH-Px). Data revealed that the inhibition of PARP significantly ameliorated oxidative stress; in other words, PARP activation contributed to pancreatic injury. Higher SOD levels were observed in all groups except sham (Figure 2). On the other hand, GSH-Px activities in the control group were found to be reduced. These data collectively suggest that the tissue is undergoing oxidative stress. Higher SOD accompanied by lower GSH-Px can be more dangerous; SOD can reduce superoxide to hydrogen peroxide, but if hydrogen peroxide accumulates because of reduced GSH-Px levels, it can undergo a Fenton reaction to produce the hydroxyl radical. This may have occurred in the ANP group. However, in the ANP + benzamide group, both higher SOD and GSH-Px might reduce such an oxidative risk as evidenced in Figure 2.
PARP activation may contribute to pancreatic injury via several other mechanisms: PARP activation is likely to upregulate the activities of both extramitochondrial NADPH oxidase and mitochondrial NADH oxidase, which are the important superoxide-generating enzymes. 33 In addition, PARP activation alters transcriptional regulation and activates nuclear factor κ-B and activator protein-1, a transcription factor that regulates proinflammatory gene expression. 34 Such activation leads to increased formation of endothelin-1 and inflammatory cytokines (e.g. tumor necrosis factor-α, interleukin-6 and interleukin-1β, all known to contribute to superoxide generation). 34,35
Another important outcome was the finding that bacterial translocation was significantly increased in all of the rats' tissues harvested from the pancreas, visceral peritoneum, MLN, liver and cecum after induction of ANP. Bacterial translocation is known to have critical importance for the mortality and morbidity in patients with pancreatitis. 36 Nevertheless, the pathogenesis of pancreatic superinfection is not completely understood. 37 Our findings are in agreement with the hypothesis that intra-abdominal spread by lymphatics is the pathway most likely to be involved in this process. 38 It has been shown that peroxynitrite is capable of mimicking many of the pathophysiological alterations associated with shock, i.e. endothelial and epithelial dysfunction, vascular hyporeactivity and cellular dysfunction; all these conditions are mediated by PARP activation. 39 Similarly, it has also been shown that different PARP inhibitors show a protective effect against shock-induced intestinal epithelial permeability changes. 40 Moreover, Cuzzocrea et al. 41 have investigated the role of PARP in intestinal tight junction permeability, and this study has demonstrated that the alteration of permselectivity is most likely induced by PARP activation. Thus, it seems likely that benzamide treatment prevented bacterial translocation in the peritoneum, MLN, liver and cecum by inhibiting activation of PARP, which consequently led to an alteration in permselectivity of the intestinal mucosa. However, benzamide was not able to prevent bacterial translocation in the pancreatic tissue. The possible reason for this diversity may be that the pancreas is itself the main source of the inflammatory process.
In conclusion, inhibition of PARP with benzamide reduced severity, septic complications and mortality in the experimental ANP rat model. These findings suggest that it may be possible to improve the outcome of ANP by using novel and more effective PARP inhibitors as a promising therapeutic strategy.
Footnotes
ACKNOWLEDGEMENTS
We thank epidemiologist Selim Kilic, MD, PhD, for statistical evaluation. This work is a part of studies supported by the ‘Gulhane Military Medical Academy Research and Progress Center’ by Grant AR-2007/3.