
Abstract
Antigen presenting cells such as intestinal macrophages are dynamic effector cells that play a critical role in maintaining mucosal homeostasis. However, it is not known how occult intestinal infections alter the response of the intestinal mucosa to subsequent intestinal injury. The aim of this study was to evaluate how persistent subclinical intestinal infection with Mycobacterium avium subsp. paratuberculosis (Map) would influence acute dextran sulfate sodium (DSS)-mediated intestinal inflammation. BALB/c mice were infected intraperitoneally with Map. Following an incubation period of 90 d, mice were administered 2% DSS in the drinking water for six days. Prior to and during treatment with DSS, mice were evaluated for clinical signs of disease and body weights were recorded. At termination of the experiment, body weights, frequency of rectal blood, and gross and histological cecal lesions were evaluated, and tissues were collected for isolation of Map. Subclinical and persistent intestinal Map infection was established based on the absence of both weight loss and rectal blood and the isolation of Map from the small and large intestines in mice infected with Map only. Following treatment with DSS, Map-infected mice had increased weight loss, increased frequency of rectal blood, and exacerbation of gross lesions and increased cecal lesion scores. Also, there was a significant reduction in Map isolated from the small intestines of Map-infected and DSS-treated mice. In conclusion, subclinical Map infection sensitizes the host to enhanced acute DSS-mediated intestinal inflammation.
Introduction
Antigen presenting cells (APCs) such as intestinal macrophages and dendritic cells are continual targets for enteric pathogens, and thus it is imperative to understand mechanisms that balance APC effector function with mucosal homeostasis. For example, in Crohn's disease (CD), a chronic inflammatory disease of the gastrointestinal tract, a disorder of macrophage function resulting in impaired bacterial clearance and cytokine secretion has been described, with some investigators suggesting that CD represents a primary immunodeficiency of macrophages. 1,2 What is less clear is what role, if any, intestinal pathogens that target APCs have in CD and, more specifically, what impact subclinical intestinal infections have on the homeostasis of the intestinal mucosa.
There has been an association between exposure to Mycobacterium avium subsp. paratuberculosis (Map) and CD. 3 Map has been detected by in situ hybridization, polymerase chain reaction and bacterial isolation in patients with CD. 4–6 However, contradictory reports exist on the significance of Map in the pathogenesis of CD. For example, some studies have not detected Map 16s rRNA in patients with CD or ulcerative colitis (UC). Even though a subset of CD patients may have genetic defects that affect innate immune function, namely, the NOD2 mutation, which may predispose patients to an intracellular pathogen such as Map, there is a lack of an association between NOD2 polymorphisms and Map infection. 7–9
Limited studies have examined the role of occult enteric infections on the development of intestinal inflammation. It has been noted that there is an increased prevalence of active cytomegalovirus (CMV) infections in patients with inflammatory bowel disease (IBD). 10 It is speculated that this represents reactivation of a latent infection. In support of a potential role for CMV in IBD, it has been shown that mice with pre-existing subclinical CMV infection have enhanced disease following dextran sulfate sodium (DSS)-mediated colitis. 11 Additional studies have demonstrated that prior exposure to opportunistic pathogens, such as Hymenolepis diminuta, Bacteroides fragilis or adherent-invasive Escherichia coli (AIEC), exacerbates subsequent oxazolone- or DSS-mediated intestinal inflammation. 12–14 In addition to specific pathogens, several studies have indicated that extra-intestinal exposure to bacterial products, such as unmethylated cytosine-guanosine dinucleotides (CpG) or staphylococcal enterotoxin B, may also influence intestinal inflammation. 15,16 Collectively, these studies suggest that specific microbial pathogens or their products can alter intestinal mucosal homeostasis and predispose the host to enhanced intestinal injury.
Considering the ongoing potential for intestinal exposure and occult infection by pathogenic and environmental species of mycobacteria, we set out to understand how exposure to Map would impact mucosal homeostasis. Our hypothesis was that exposure to Map would enhance acute mucosal injury. To address this goal, we utilized a mouse model of intestinal Map infection. 17,18 BALB/c mice were infected with Map for 90 d followed by treatment with a low dose of DSS in the drinking water to induce mild intestinal injury. In these mice, we evaluated clinical signs of disease and development of mucosal pathology before and after intestinal injury. Our results demonstrate that Map established subclinical infection in BALB/c mice. Furthermore, subclinical intestinal mycobacterial infection amplified the severity of DSS-mediated intestinal inflammation as indicated by enhanced clinical disease, exacerbation of gross lesions and increased lesion scores.
Materials and methods
Animals
Conventionally reared 6–8-week-old male BALB/c mice were purchased from The Jackson Laboratory (Bar Harbor, ME, USA) and weighed approximately 22–24 g. Mice were housed in micro-isolation units that were individually ventilated in a positive pressure Thoren unit caging system at the Iowa State University College of Veterinary Medicine biosafety level II animal care facility. All animal experiments were approved by the Institutional Animal Care and Use Committee of Iowa State University.
Bacterial inoculum and infection
The Map strain K10 was a clinical isolate obtained from the National Animal Disease Center (Ames, IA, USA) and maintained in Middlebrook 7H9 broth supplemented with mycobactin J. Logarithmic growth-phase bacteria were washed and resuspended in sterile saline for inoculation. Bacterial concentration was determined by measuring the absorbance at 540 nm, comparing the absorbance optical density with the standard curve and adjusted to a final concentration of 1 × 109 colony-forming units (CFU). Controls were given a sham inoculation of saline. The Map inoculum used in these studies was shown to have greater than 90% viability via fluorescein diacetate staining and flow cytometric analysis prior to inoculation. In addition, challenge inocula were confirmed negative for contaminants by streaking onto sheep blood agar plates 24 h prior to inoculation.
Experimental design
In three separate experiments, 10–11 mice per treatment group were given a single intraperitoneal injection of Map or sham inoculated with saline. Following inoculation, body weights of the mice were measured at weekly intervals and feces were periodically collected for the isolation of Map. At approximately three months postinoculation, acute intestinal injury was chemically induced by administering a suboptimal (as discussed below) concentration of 2% DSS (MP Biomedicals, Solon, OH, USA) in the drinking water for six days. We established this dose by determining the lowest DSS concentration in the drinking water that would induce minimal but perceptible microscopic lesions in the cecal and colonic mucosa of DSS-only-treated mice. Using a pilot study, we tested doses that ranged from 1% to 5% DSS, increasing by 0.5% DSS in each treatment group. Of these, 2% DSS induced consistent, mild, multifocal villus elongation with minimal erosions of the mucosal epithelium. This dose was selected for all experiments in this study. While the mice were administered DSS, they were weighed daily, evaluated for clinical signs of disease, such as the presence of rectal blood and huddling, and on day 6 all of the mice were euthanized with CO2 and necropsies were preformed. At necropsy, body weight and colon and cecal length were measured. Additionally, sections of the spleen, liver, small and large intestine were collected for histopathology and isolation of Map. Additionally, saline washes from the abdominal cavity were collected and inoculated onto 7H10 agar for isolation of Map.
Histopathology
At necropsy, samples of the liver, spleen, small intestine, mesenteric lymph node, cecum and colon were fixed in 10% neutral-buffered formalin, processed by routine methods, embedded within paraffin wax and sectioned. Sections of tissue (5 μm) were stained with hematoxylin and eosin. Cecal histological lesion scores were generated based on the severity of mucosal epithelial erosion and hyperplasia, the degree of lamina propria inflammatory cell infiltrate and edema (Table 1). Additionally, the distribution of each parameter was assessed.
Cecal lesion scoring system
Parameters were scored for each individual animal resulting in a maximal total histological lesion score of 24 per cecum
Acid-fast staining (Ziehl–Neelsen method)
Slides were deparaffinized and rehydrated in distilled water. Carbol fuchsin solution was added for 45 s while microwaving at 450 W. Slides were rinsed in 0.5% acid alcohol for decolorization, rinsed for one minute in tap water and then counterstained in methylene blue for one minute.
Culture of M. avium subsp. paratuberculosis from tissues and feces
Sections of the spleen, liver, small intestine and colon were collected for isolation of Map. Each piece of the tissue was weighed and homogenized in 0.75% hexadecylpyridinium chloride solution and then allowed to stand overnight at room temperature to decontaminate the cultures. Sediments from individual tissue homogenates (100 μL) were inoculated onto Middlebrook 7H10 agar. Additionally, the peritoneum was washed with saline and 100 μL was plated onto Middlebrook 7H10 agar. After 12 weeks of incubation at 37°C, the number of colonies was counted. Fecal pellets were collected from each group of mice at approximately 14-day intervals until the end of the experiment. The fecal pellets were homogenized in 1 mL of 0.05% Tween 80, diluted, decontaminated and plated on supplemented 7H10 agar as described above.
Immunohistochemistry for F4/80
Before staining, slides were deparaffinized, rehydrated and processed for immunohistochemistry. Antigen retrieval was accomplished by incubating the slides with 2% protease in Tris buffer for 20 min at room temperature. Slides were then blocked with 10% normal goat serum at room temperature. The primary antibody was used at 1:1000 (eBioscience, San Diego, CA, USA) and incubated overnight at 4°C. Biotinylated goat anti-rat IgG F(ab')2 fragment secondary antibodies at 1:250 was added and incubated at room temperature for 30 min. HRP-streptavidin diluted 1:200 was added to the slides for 15 min and then DAB was added until optimal staining developed (10–15 min).
Statistical analysis
Changes in body weight were analyzed using a analysis of variance model with repeated measures. Map infection or saline inoculation, DSS or water administration, day and their interactions were considered as fixed effects, with animal being the subject of repeated measures. On each day, the treatment group means were compared first with an overall F-test and then followed by a post hoc Tukey–Kramer's t-test for multiple comparisons if the F-test was significant. A P value of <0.05 was considered significant. For all other parameters, treatment group means were compared first with an overall F-test and then followed by a post hoc Student's t-test with group mean differences considered significant if the P value was <0.05. Data are presented as the mean value ± standard error of the mean except where stated otherwise.
Results
Clinical disease and gross lesions
To establish subclinical intestinal infection, we conducted an initial dose titration study with doses ranging from 1 × 105 to 1 × 108 CFU. A dose of 1 × 108 CFU of Map was selected for subsequent studies. Mice infected with Map transiently lost up to 1 g of their body weight within the first seven days after inoculation relative to mice given a sham inoculation with saline (data not shown). However, mice recovered their body weights and by the end of the 90-day incubation period all treatment groups had similar body weights.
To evaluate clinical disease, we monitored the body weight and rectal bleeding. Other than transient weight loss, inoculation with Map alone did not result in overt signs of clinical disease. This has previously been reported for BALB/c mice. 19 Significant weight loss was identified only in the Map + DSS mice and occurred on day 6 of DSS administration (Figure 1, P < 0.03). Rectal bleeding (Table 2) occurred only in treatment groups given DSS and coincided with the onset of weight loss. There was a greater frequency and severity of rectal blood in Map + DSS mice; 40% of Map + DSS mice had rectal blood compared with 14% of DSS-only mice. In DSS-only mice, there was scant blood limited to the region immediately around the anus. In the Map + DSS group, there was a large amount of blood around the anus extending into the perineum and the ventral base of the tail.
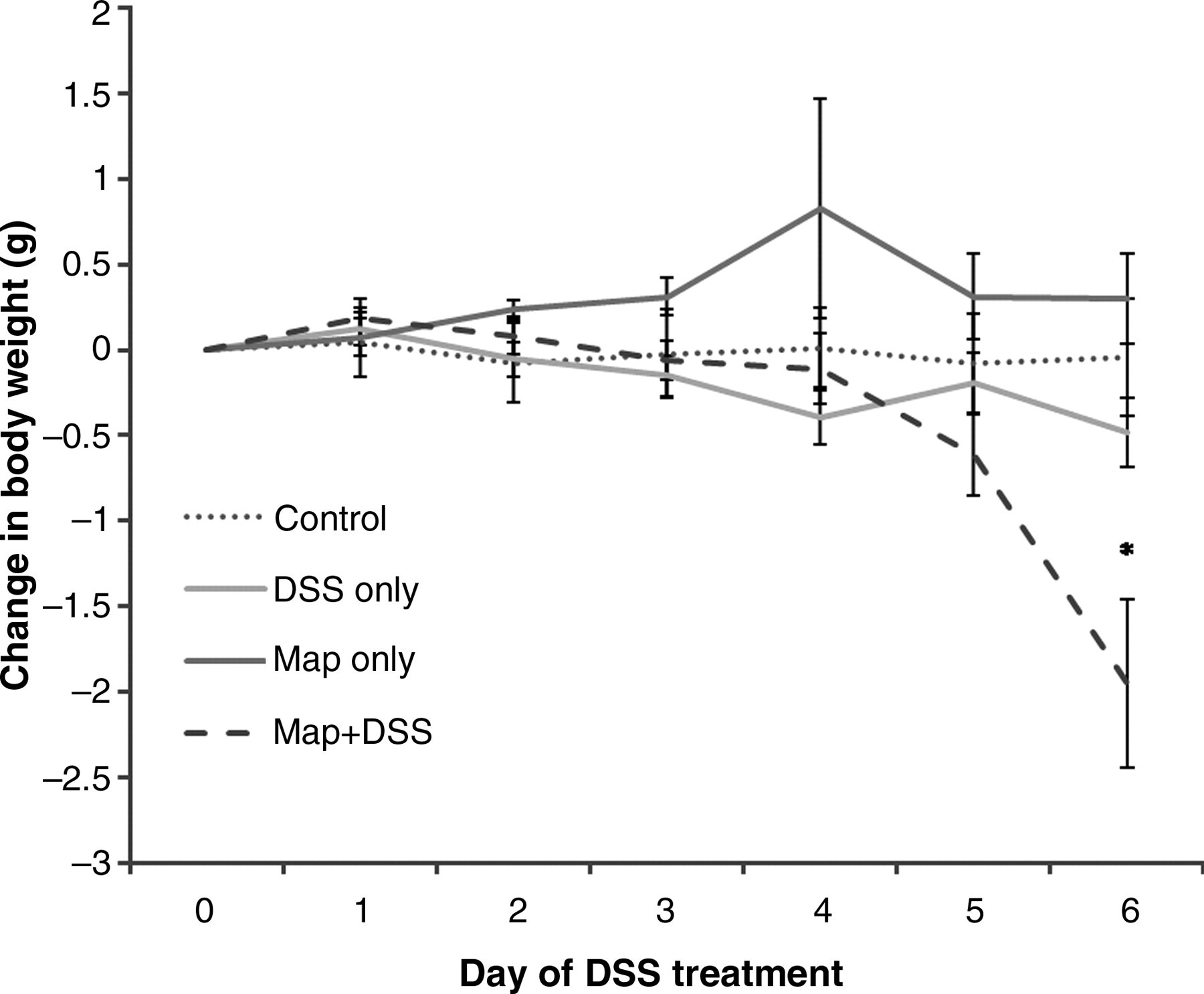
Change in body weight in mice inoculated with saline (control); infected with Map only (Map only); inoculated with saline and treated with DSS (DSS only); and mice infected with Map and treated with DSS (Map + DSS). Map + DSS mice lost a statistically significant amount of body weight relative to all other treatment groups (*P < 0.03). Data are represented as the mean ± standard error of the means from three separate experiments including 22 DSS-only mice, 25 Map + DSS mice, eight Map-only mice and nine control mice. Map, Mycobacterium avium subsp. paratuberculosis; DSS, dextran sulfate sodium
Severity and frequency of rectal blood
Map, Mycobacterium avium subsp. paratuberculosis; DSS, dextran sulfate sodium
Severity and % of mice with rectal blood in mice infected with Map only (Map only); inoculated with saline and treated with DSS (DSS only); and mice infected with Map and treated with DSS (Map + DSS). A greater percentage of Map + DSS mice had rectal blood compared with all other treatment groups. Data are represented as the % of affected mice from three separate experiments including 22 DSS-only mice, 25 Map + DSS mice, eight Map-only mice
A consistent gross lesion in the murine DSS colitis model is cecal and colonic shortening (atrophy) and this was used as a measure of gross pathology in this model (Figure 2). 20 Cecal atrophy was most severe in the Map + DSS group (Figure 2d; Figure 3, **P < 0.0001). DSS-only mice had significant cecal shortening relative to the negative control and the Map-only group (Figure 3, *P < 0.03). Map-only mice had cecal lengths similar to the negative control. We observed similar trends in the colon for gross and histological lesions, with significant shortening of the colon in Map + DSS mice compared with all other treatment groups. We chose to focus our analysis on the cecum because of more consistent lesions with enhanced severity.
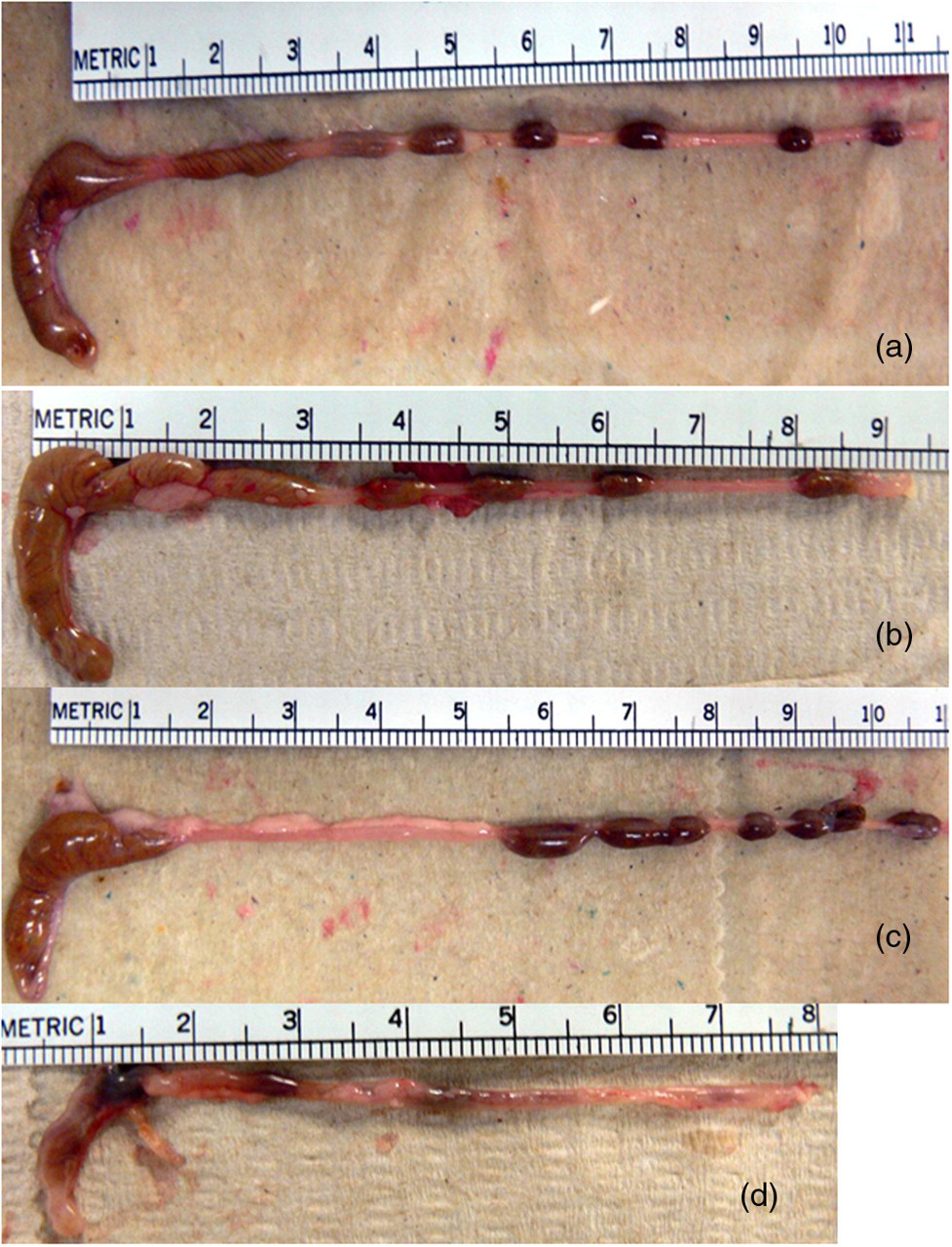
Representative pictures demonstrating cecal atrophy. Control (a); Map only (b); DSS only (c); and Map + DSS (d). Note that there is also shortening of the colon and a reduction in the number of formed fecal pellets in the mice administered DSS. Map, Mycobacterium avium subsp. paratuberculosis; DSS, dextran sulfate sodium
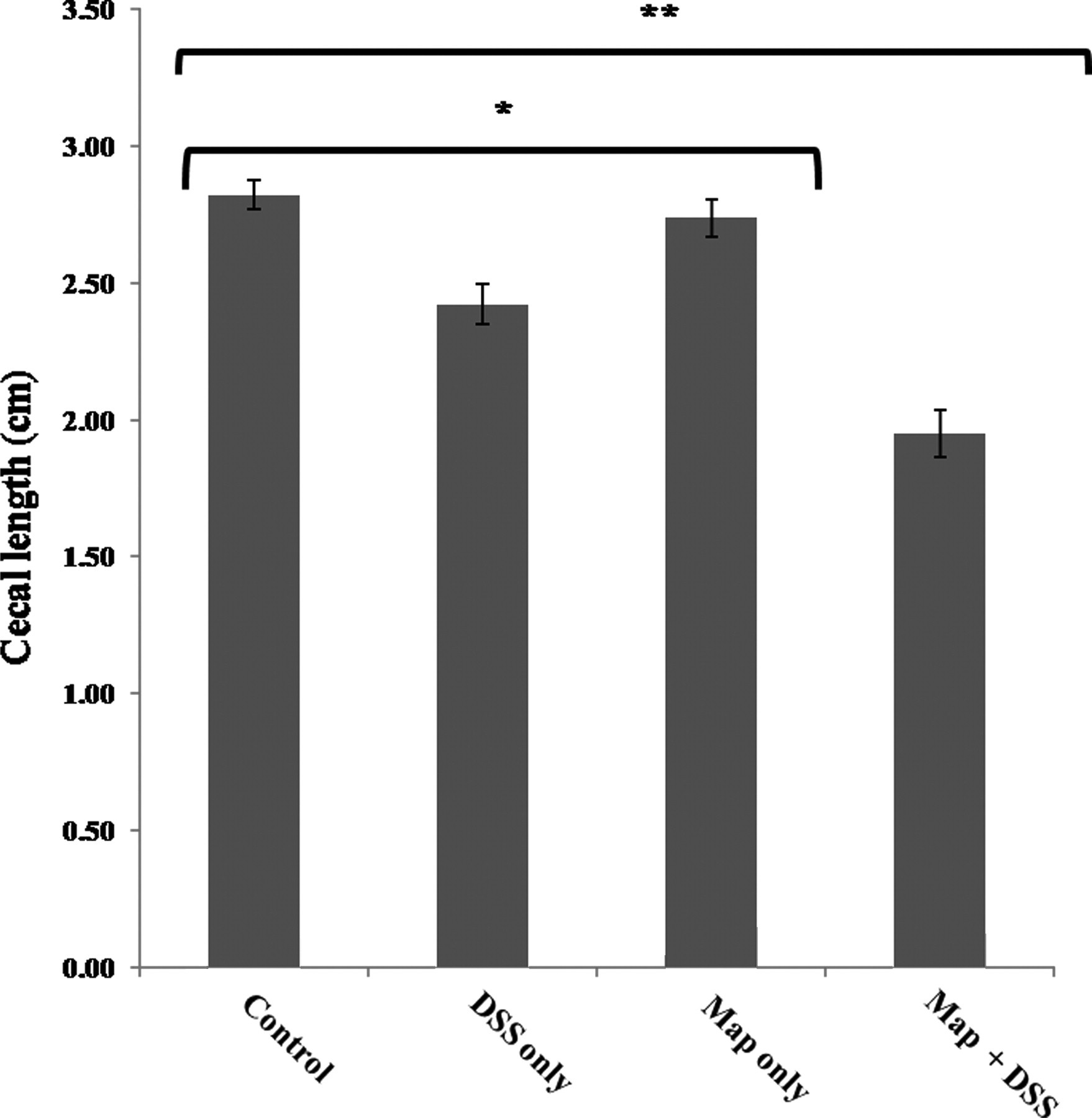
Cecal lengths in mice inoculated with saline (control); infected with Map only (Map only); inoculated with saline and treated with DSS (DSS only); and mice infected with Map and treated with DSS (Map + DSS). The average cecal length in Map + DSS mice was shorter relative to all other treatment groups (**P < 0.0001). The average cecal length for DSS-only mice was shorter relative to the control and Map-only mice (*P < 0.03). Data are represented as the mean ± standard error of the means from three separate experiments including 22 DSS-only mice, 25 Map + DSS mice, eight Map-only mice and nine control mice. Map, Mycobacterium avium subsp. paratuberculosis; DSS, dextran sulfate sodium
Cecal histological lesion scores
Microscopic lesions were typical of the DSS mucosal injury model and were defined by a multifocal proliferative to ulcerative typhlitis and colitis (Figure 4). The histological grading system used in this study is outlined in Table 1. In the DSS-only group, there was mild multifocal gland proliferation and rare mucosal ulceration. The histological score was not significantly different between the DSS-only, negative control and Map-only groups. In contrast, the lesions were more severe and widespread in the Map + DSS group (Figures 4 and 5, *P < 0.0013). Inoculation with Map alone did not induce histological lesions. Lesions within the large intestine have been reported for BALB/c mice infected with Map 19698; however, lesions were not prominent until 32 weeks after infection and it was not specified whether or not they were located in the cecum. 21
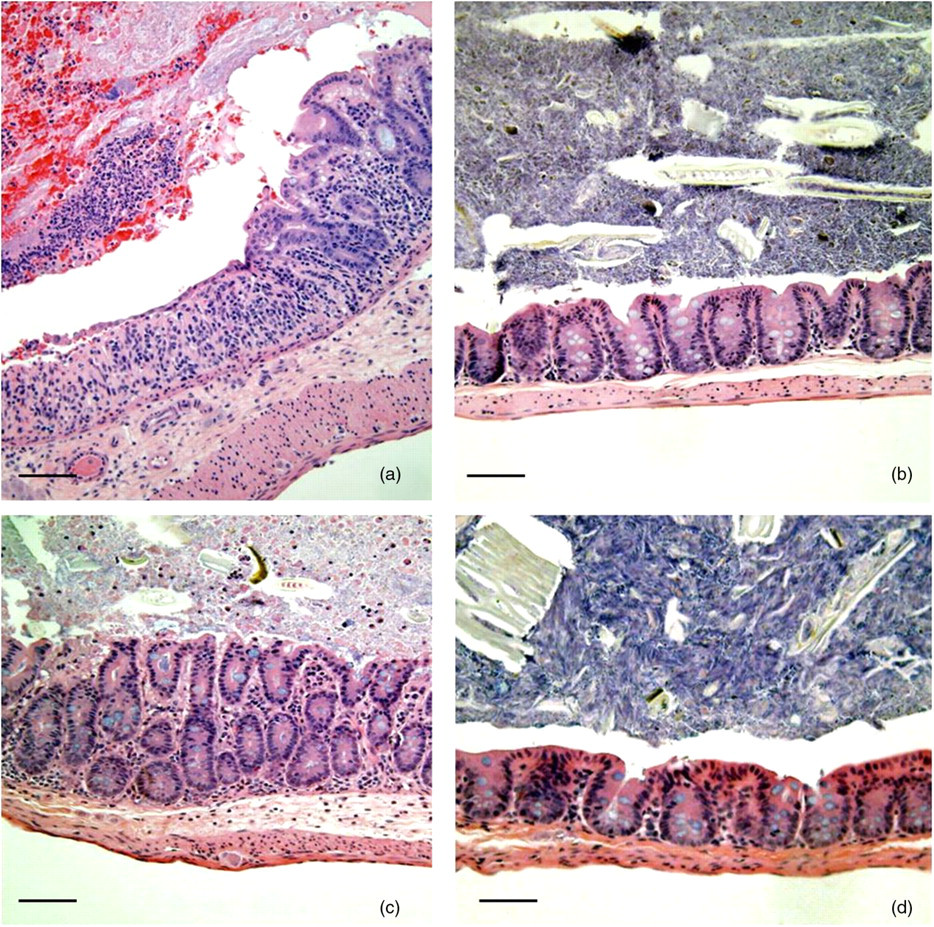
Representative photomicrographs of ceca from Map + DSS (a), Map only (b), DSS only (c) and control (d) mice. Note the ulceration of the mucosa and the cecal lumen containing neutrophils intermixed with blood in the Map + DSS mouse. There is moderate hyperplasia and submucosal edema in the DSS-only mouse. Lesions were not identified in the mucosa of the Map-only and control mice. HE; Bar = 100 μm. Map, Mycobacterium avium subsp. paratuberculosis; DSS, dextran sulfate sodium; HE, hematoxylin–eosin
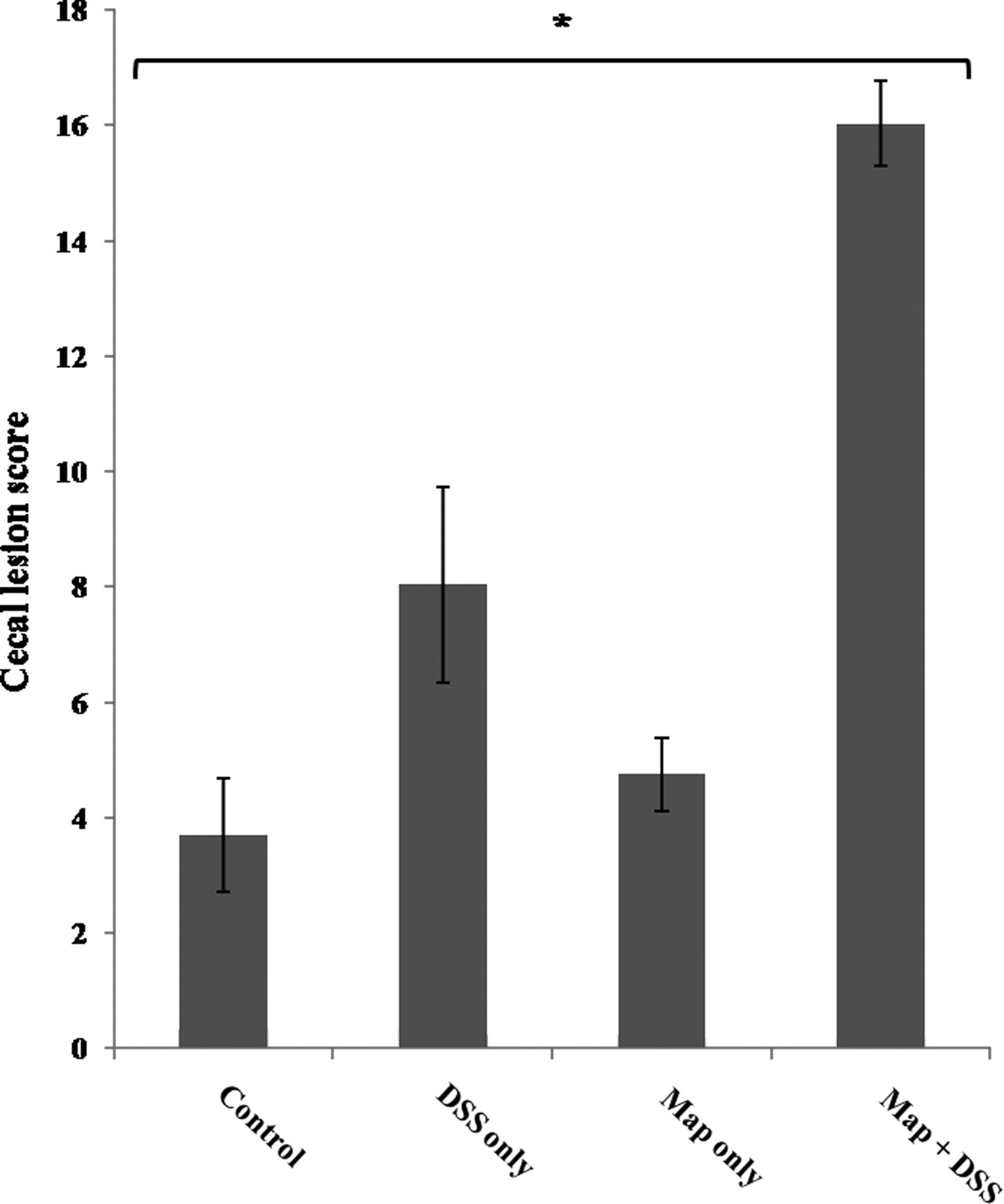
Cecal histological lesion scores in mice inoculated with saline (control); infected with Map only (Map only); inoculated with saline and treated with DSS (DSS only); and mice infected with Map and treated with DSS (Map + DSS). Cecal lesion scores were significantly higher in Map + DSS mice relative to all other treatment groups (*P < 0.0013). Data are represented as the mean ± standard error of the means from three separate experiments including 22 DSS-only mice, 25 Map + DSS mice, eight Map-only mice and nine control mice. Map, Mycobacterium avium subsp. paratuberculosis; DSS, dextran sulfate sodium
M. avium subsp. paratuberculosis tissue colonization
The distribution of Map in tissues was similar to that reported previously for Map-infected BALB/c mice. 18 Map was isolated from the spleen, liver, peritoneum, small intestine and large intestine, with the greatest concentration of bacteria being isolated from the spleen (Figure 6). Map was not isolated from uninfected control mice. DSS administration had a significant effect on Map recovery only in the small intestine where DSS administration corresponded to lower CFU in Map-infected mice (Figure 6,*P = 0.043). There was a trend for decreased numbers of Map CFU in the colon of mice given DSS, but this did not reach statistical significance. Map was not isolated from the feces from any of the treatment groups.
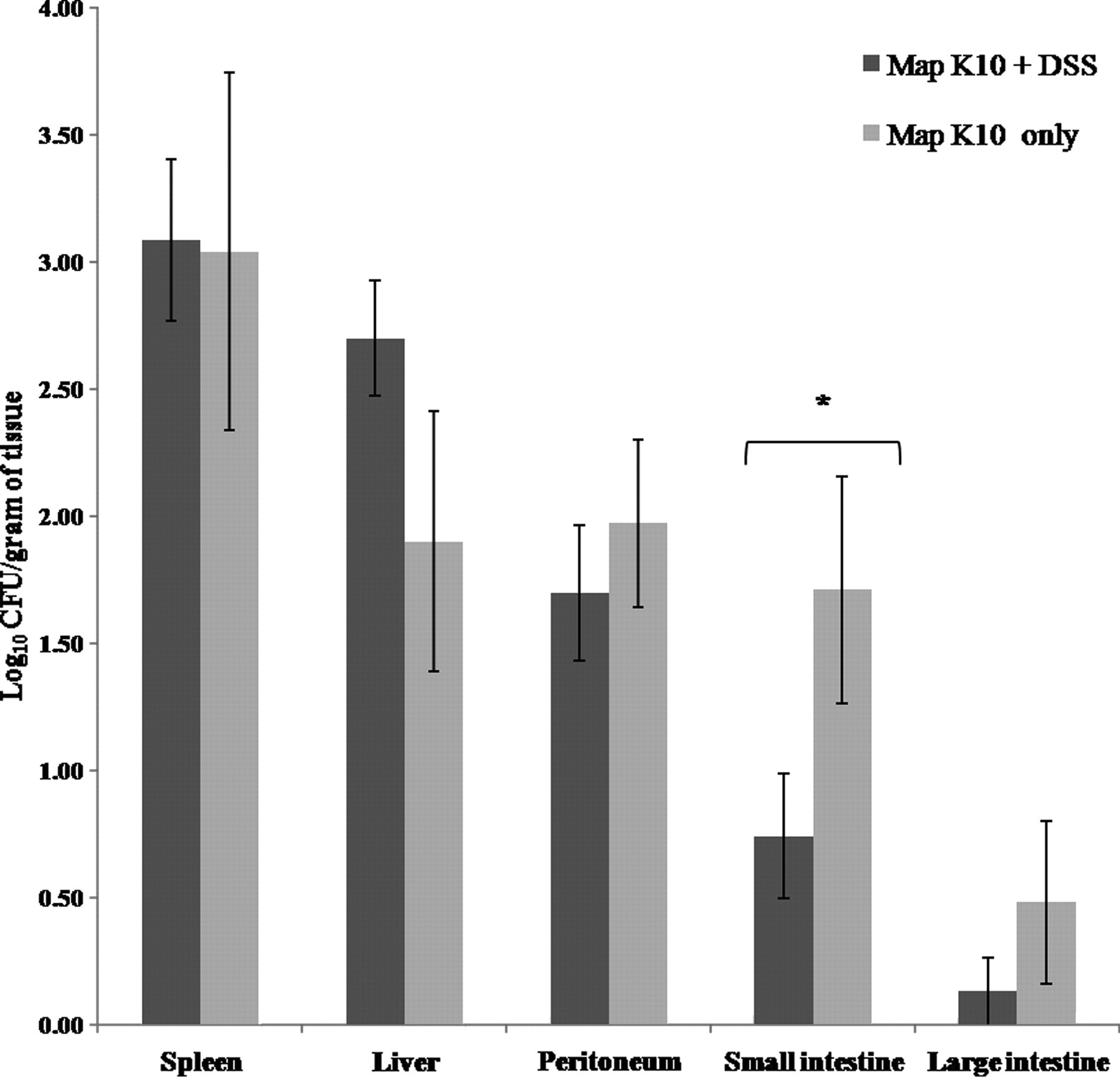
Log10 colony-forming units per gram of tissue in mice infected with K10 Map and given either DSS or water. A greater concentration of K10 Map was isolated from the small intestine from Map-K10-only mice relative to Map K10 + DSS mice (*P = 0.043) . Map, Mycobacterium avium subsp. paratuberculosis; DSS, dextran sulfate sodium
Acid-fast stains of tissues
Acid-fast bacilli were present within the lamina propria of the cecum and small intestine and foci of granulomatous inflammation in the liver, spleen and peritoneum. Consistent with the isolation results, there was a greater density of acid-fast bacilli in the spleen, liver and at the site of inoculation in the peritoneum compared with the small and large intestine. In the small and large intestine, low numbers of acid-fast bacilli were identified within the mucosa. In the large intestine, acid-fast bacilli were most commonly associated with gut-associated lymphoid tissue (GALT). Acid-fast bacilli were present in cells with macrophage morphology, which is consistent with Map being an obligate pathogen of macrophages. 21 To confirm that cells containing acid-fast bacilli were macrophages, immunohistochemistry for F4/80 antigen, a marker for mouse macrophages, was performed on serial sections of the cecal tonsil. F4/80-positive cells were associated with foci of cells containing acid-fast bacilli (Figure 7).
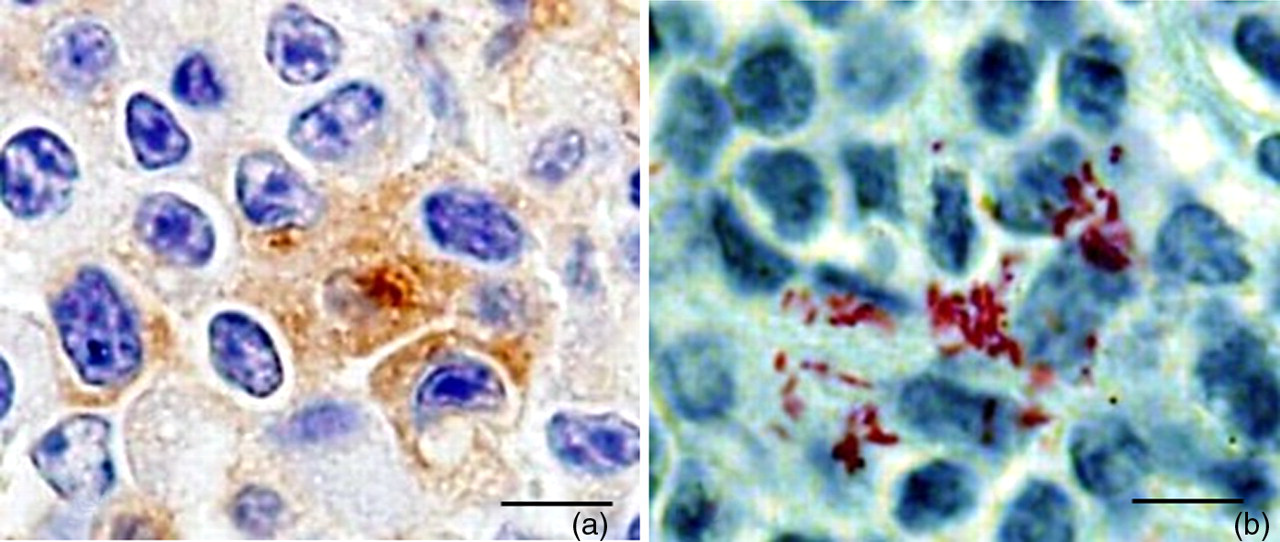
Serial sections of cecal tonsil demonstrating acid-fast bacilli (a) associated with F4/80 immunopositive cells (b). Note F4/80 immunopositive cells are brown. Bar = 20 μm
Discussion
We set out to explore how occult intestinal mycobacterial infection would influence intestinal homeostasis. This is relevant considering the frequency of enteric mycobacterial infection in immunocompromised patients, and the potential for opportunistic infections in individuals with persistent intestinal inflammation, such as those with IBD. 22 In this study, we demonstrated that in our model, subclinical Map infection sensitizes mice to DSS-mediated intestinal injury. Mice infected with Map and treated with DSS had increased weight loss and frequency of rectal blood, increased cecal lesion scores and more severe cecal atrophy. These observations suggest that mycobacterial infection alters the mucosal response to injury and potentially influences repair mechanisms.
Map infection alone resulted in persistent infection but did not induce clinical disease as indicated by the absence of weight loss and lack of rectal blood. This is supported by previous observations in Map-infected BALB/c mice. 19 Infection was demonstrated by the isolation of Map from the spleen, liver, small and large intestines, and peritoneum. Acid-fast staining localized Map to the GALT in the cecum and foci of granulomatous inflammation in the spleen, liver and peritoneum. Even though bacteria were isolated and acid-fast bacilli were demonstrated in both the small and large intestines, distinct foci of granulomatous inflammation were not present in these tissues. The bacterial numbers recovered from the small and large intestine were low, which is consistent with prior studies in the BALB/c mouse. 23 This condition bears some similarities to occult abdominal M. avium infection in humans where clinical disease is absent and bacteria difficult to identify. 22
When enteric pathogens are recognized, the mucosal immune response must control pathogen proliferation and dissemination, yet minimize inflammation and injury. It has become clear that APCs such as intestinal macrophages and dendritic cells play a critical role in maintaining the balance between mucosal defense and homeostasis. 24 With respect to their homeostatic role, resident intestinal macrophages have a unique functional phenotype. They can eliminate potential pathogens while at the same time preventing excessive inflammation. 25 This anti-inflammatory phenotype of human intestinal macrophages is defined by the absence of innate response receptors such as receptors for LPS (CD14) and CR4 (CD11c/CD18) and the inability to produce proinflammatory cytokines in response to inflammatory stimuli. 25 Despite this dual protective role of intestinal macrophages, it is clear that macrophages can contribute to the pathogenesis of mucosal injury, such as in IBD where it has been documented that large amounts of interleukin (IL)-1β are produced almost exclusively by intestinal macrophages. 26,27 Although studies demonstrate that intestinal macrophages have important homeostatic or potentially pathogenic roles in the intestinal mucosa, it is unclear whether or not pathogens that target APCs such as macrophages can cause a shift in macrophage effector function and impact mucosal homeostasis. In this study, we have demonstrated that an occult infection with a pathogen that has a tropism for intestinal macrophages is capable of augmenting subsequent intestinal injury. In addition, our previous studies have shown that Map-infected DC may also contribute to altered innate immune responses in this model. 28
To the authors' knowledge, only one other study, similar to the current one, has been described in which latent CMV infection, a virus known to infect intestinal macrophages, 29 sensitized mice to a worsening of disease following treatment with DSS. 11 Although infection was established in these mice based on the detection of viral DNA in various tissues, including the intestines, clinical signs of disease such as weight loss and gross or occult colonic or rectal blood were not observed, in addition to the absence of viral-induced histological lesions. After the administration of a similar dose of DSS used in the current study, enhanced disease was demonstrated in those mice subclinically infected with CMV, as evidenced by increased weight loss and enhanced gross and histological lesion scores. One other study has evaluated the effect of Map in a model of intestinal injury. Increased weight loss and increased colonic lesion scores following oral challenge with live Map, strain Ben (ATCC 43544), were demonstrated in an IL-10-deficient mouse model of spontaneous colitis. 30 Map infection of IL-10-deficient mice resulted in enhanced production of Ag85B/MPT59 peptide 25 specific tumor necrosis factor-α and interferonγ from both serum and cells isolated from the Peyer's patches and mesenteric lymph nodes. The authors proposed that since Ag85B/MPT59 peptide 25 is conserved across several mycobacterial species, Map may function as an immunomodulator that stimulates T-cells in a manner similar to a superantigen. This suggests that the enhanced disease observed in these mice is not unique to Map, but may occur following infection by certain pathogenic and non-pathogenic species of mycobacteria.
In our study, intestinal colonization was demonstrated by isolation of bacteria from both the small and large intestines and by demonstration of acid-fast bacilli that were often present in GALT. In addition to the intestines, and consistent with previous studies in BALB/c mice, the greatest concentration of bacteria was isolated from the liver and spleen and was not significantly affected by treatment with DSS. 21 A smaller number of bacteria were isolated from the small intestines in mice treated with DSS. In contrast, smaller numbers of bacteria were isolated from the large intestine, but this was not statistically different. This result may partially be explained by an enhanced non-specific immune response induced by DSS, which enhances bacterial killing. Dextran sulfate has been demonstrated in colonic macrophages in a DSS-mediated mouse model of colitis. 31 Moreover, it has been shown that DSS-pulsed macrophages promote T-cell responses in colitic mice. 32
In summary, we propose that our model can be utilized to better understand how persistent enteric infection with an opportunistic pathogen, such as Map, disrupts the regulation of intestinal homeostasis and tips the balance in favor of enhanced inflammation following injury to the intestine. Enhanced disease in our model may be associated with Map-mediated exacerbation of inflammation, disruption of repair mechanisms or a combination of both. Future studies with this model will define the immune profile of the locally injured intestinal mucosa and attempt to characterize how chronic mycobacterial infection modifies the function and phenotype of intestinal macrophages, which are critical components of an elaborate regulatory system that maintains the intestine in a non-inflammatory state despite the presence of large numbers of bacteria in the adjacent intestinal lumen.
Footnotes
ACKNOWLEDGEMENTS
We thank Elise Huffman and Dr Brandon Plattner for technical assistance. This work was funded through an Iowa State University research grant and the Iowa Healthy Livestock Initiative.