
Abstract
Current therapy of a number of neuropsychiatric maladies has only symptomatic modality. Effective treatment of these neuro-degenerative diseases, including amyotrophic lateral sclerosis (ALS), may benefit from combined gene/stem-cell approaches. In this report, mononuclear fraction of human umbilical cord blood cells (hUCBCs) were transfected by electroporation with dual plasmid constructs, simultaneously expressing vascular endothelial growth factor 165 (VEGF165) and human fibroblast growth factor 2 (FGF2) (pBud-VEGF-FGF2). These genetically modified hUCBCs were injected retro-orbitally into presymptomatic ALS transgenic animal models (G93A mice). Lumbar spinal cords of rodents were processed for immunofluoresent staining with antibodies against human nuclear antigen (HNA), oligodendrocyte-specific protein, S100, iba1, neuronal β 3-tubulin and CD34. Co-localization of HNA and S100 was found in the spinal cord of mice after transplantation of genetically modified hUCBCs over-expressing VEGF-FGF2. Double staining in control animals treated with unmodified hUCBCs, however, revealed HNA+ cells expressing iba1 and CD34. Neuron-specific β 3-tubulin or oligodendrocyte-specific protein were not expressed in hUCBCs in either control or experimental mice. These results demonstrate that genetically naïve hUCBCs may differentiate into endothelial (CD34+) and microglial (iba1+) cells; however when over-expressing VEGF-FGF2, hUCBCs transform into astrocytes (S100+). Autocrine regulation of VEGF and FGF2 on hUCBCs, signal molecules from dying motor neurons in spinal cord, as well as self-differentiating potential may provide a unique microenvironment for the transformation of hUCBCs into astrocytes that eventually serve as a source of growth factors to enhance the survive potential of surrounding cells in the diseased regions.
Introduction
Amyotrophic lateral sclerosis (ALS), also known as Lou Gehrig's disease, is a neurodegenerative disease characterized by gradual loss of motor neurons in the brain and spinal cord leading to muscle paralysis throughout the body. The etiology and pathogenic mechanisms of cell death in most forms of ALS are largely unknown. Due to the lack of effective therapy, patients die from respiratory failure within 3–5 y after the onset of paralytic symptoms. Major therapeutic strategies are aimed at inhibition of excitotoxicity and oxidative damage, which are key factors in motor neuronal damage. Stem cell therapy is a potential treatment option in degenerating diseases by which lost parenchymal cells might be replaced. It is unreasonable to expect, however, that stem cells per se could substitute dying neural cells in rigid, highly organized neural networks.
Glial cells may be very effective in increasing survivability of affected neurons by providing a physical scaffolding support and by secreting neurotrophic factors. Transplantation of predifferentiated stem cells with a potential to become glias seems to be a promising strategy for ALS treatment. Predifferentiated glial cells can be obtained from various stem cell sources, such as embryonic or neural stem cells. A more easily accessible alternative is the human umbilical cord blood (hUCB), which contains several types of stem cells, including hematopoietic (CD31, CD34, CD59, CD90, oct4, nanog, oct4, sox2, sca1, fgf4) and mesenchymal stem cells (CD34, CD90, CD105, sca1), as well as endothelial progenitor cells (CD34, gata2, flk1). The rationale for using hUCB cells (hUCBCs) for cellular therapy is based not only on their potential to differentiate into various cell types, such as erythrocytes, leukocytes, megakaryocytes, myoblasts of heart and skeletal muscle tissue, hepatocytes, vascular endothelial cells, neurons, oligodendrocytes, astrocytes, etc. (for a review see reference 1 ), but also on their availability, acceptability, potential for long-term storage, compatibility with both auto- and allogenic transplantation in humans, and their low immunogenicity. Moreover, transplantation of hUCBCs is not subjected to legislative regulations, esthetics and religious prohibition.
Cellular therapy has recently been complemented with the genetic modification of stem cells. Over-expression of cell-adhesion molecules, growth factors and other biologically active molecules may lead to addressed homing of transplanted stem cells that ultimately express therapeutic genes and also differentiate into parenchymal cells at the site of degeneration. In line with this notion, here we generated hUCBCs over-expressing either human vascular endothelial growth factor (VEGF) or human fibroblast growth factor 2 (FGF2), or simultaneously both, for the possible treatment of ALS.
The most widely used animal model of ALS is a G93A substitutional transgenic mouse expressing human mutant copper/zinc super-oxide dismutase-1 (SOD1) gene. These rodents are unique in that over-expression of SOD1 results in an adult-onset phenotype closely recapitulating the human disease at a clinical as well as histopathological level. 2 The G93A model has the following four stages: 3 (i) premuscle weakness stage, during which muscle strength is maintained at the normal level; (ii) rapid declining stage, where muscle strength sharply (usually by >50% within a period of two weeks) declined; (iii) slow declining stage (SD), which lasts for 4–11 weeks; and (iv) a paralytic stage. In this study, SD stage mice were used, immediately prior to the onset of paralysis of the limbs. The effect of gene modification with plasmid expression constructs, encoding both FGF2 and VEGF, on the differentiation of hUCBCs after transplantation into G93A mice was studied. Phenotyping of transplanted hUCBCs was evaluated by double immunofluorescent staining.
Materials and methods
Generation of plasmid constructs
Generation of recombinant plasmid pcDNA-VEGF, expressing human VEGF165, was described previously. 4 The open reading frame (ORF) of human FGF2 was polymerase chain reaction (PCR) amplified using specific primers with engineered restriction sites KpnI and XhoI (forward primer: CGCAGGtACCATGGCAGCCG; reverse primer: CAcTcgAgTCAGCTCTTAGCAGACATTGGAAG). The PCR product was cloned into pcDNA3.1+ plasmid using KpnI and XhoI restriction enzymes. Dual cassette expression vectors were generated using plasmid pBudCE4.1 (Invitrogen, Carlsbad, CA, USA) as a backbone. First, plasmids pBud-EGFP and pBud-VEGF were generated by subcloning enhanced green fluorescent protein (EGFP) and VEGF ORFs from plasmids pcDNA-EGFP and pcDNA-VEGF into pBudCE4.1 using KpnI and XhoI restriction enzymes. Then plasmids pBud-EGFP-FGF2, pBud-VEGF-FGF2 and pBud-VEGF-EGFP were generated by subcloning VEGF, FGF2 and EGFP ORFs from plasmids pcDNA-VEGF, pcDNA-FGF2 and pcDNA-EGFP into plasmids pBud-EGFP and pBud-VEGF using HindIII and XbaI restriction enzymes. Generation of the correct expression plasmids was confirmed by sequencing.
Cell isolation and electroporation
Mononuclear fraction of hUCBCs (hUCB-MC) was isolated by standard technique of sedimentation onto a density barrier (Ficoll, 1.077 g/mL). Isolated cells were transiently transfected by electroporation (Gene Pulser Xcell Total System; Bio-Rad, Hertfordshire, UK) with expression plasmids pBud-EGFP, pBud-EGFP-FGF2, pBud-VEGF-EGFP or pBud-VEGF-FGF2 (20 μg DNA, 0.4 cm cuvette, 300 V, 1000 μF). Cells were allowed to recover for 24 h in RPMI-1640 medium (Sigma, Lyon, France) containing 10% fetal bovine serum. Thereafter, 1 × 106 cells were injected retro-orbitally in presymptomatic G93A mice.
Realtime PCR
Expression level of human VEGF and FGF2 mRNA in transfected cells was examined by realtime PCR. Total RNA from cell cultures was isolated using a HighPure RNA Isolation Kit (Roche, Hertfordshire, UK) according to the manufacturer's instructions. cDNA synthesis was performed utilizing random hexamer primers and moloney murine leukemia virus reverse transcriptase (Promega, Madison, WI, USA) at 37°C for one hour. TaqMan primers and probes (Table 1) were designed using PrimerExpress (Applied Biosystems, Foster City, CA, USA) software. Realtime PCR primers and TaqMan probes, as well as 2.5 × premix for TaqMan realtime PCR, was purchased from Syntol (Moscow, Russia) and used according to the manufacturer's instructions. Amount of RNA was normalized using β-actin. Serial dilutions of cDNA synthesized from mRNA of hUCB-MCs, transfected with pBud-VEGF-FGF2 plasmid, were used to generate a standard curve for relative quantitation of gene expression. Expression level of genes in untransfected control hUCB-MCs was considered as 100%. TaqMan reactions were run in duplicates. Standard error and t-test values were obtained using Microsoft Excel 2007, with P < 0.05 being regarded as a statistically significant difference.
Primers and probes for realtime polymerase chain reaction
*TaqMan probes contain 5′ FAM fluorescent dye and 3′ RTQ-1 quencher
Animals and treatments
Mice, transgenic for hSOD1 (B6SJL-TG(SOD1-G93A)dl1Gur/J), were obtained from Jackson Laboratory (Bar Harbor, ME, USA) and bred at the animal facility branch of Shemyakin and Ovchinnikov Institute of Bioorganic Chemistry (Russian Academy of Science). Mature 21-week-old mice were delivered to Kazan State Medical University, where rodents were housed one/cage, under standard laboratory conditions, with a 12-h light/dark cycle and unlimited access to food and water. Sixteen transgenic female and male mice were randomly assigned into four groups (n = 4) based on transplantation of genetically modified hUCBCs expressing EGFP, EGFP-VEGF, EGFP-FGF2 or VEGF-FGF2. In experimental groups, 1 × 106 cells were injected retro-orbitally in presymptomatic 24–28-week-old G93A mice. Two weeks after transplantation, mice were euthanized under ketamine–xylazine anesthesia. The animal protocol was approved by the Animal Care and Use Committee of Kazan State Medical University.
Immunofluorescence
Mice were transcardially perfused with cold phosphate-buffered saline (PBS), followed by cold 4% paraformaldehyde in PBS (pH: 7.4). Entire spinal cords were removed, postfixed with paraformaldehyde overnight at 4°C, and immersed in 15% sucrose solution in 0.1 mol/L cacodylate buffer (pH: 7.3) for one day at 4°C. Spinal cords were subdivided into cervical, thoracic and lumbar parts. Tissues were frozen for two minutes in 2-methyl-butane (iso-pentane) precooled to −30°C. For sectioning, the spinal cord segments were attached to a cryostat specimen holder using TissueTek (Sakura Finetek Europe, Zoeterwoude, Netherlands). Serial longitudinal sections of 20-μm thickness were cut on a cryostat and picked up on SuperFrostPlus glass slides. The sections were air-dried for at least one hour at room temperature (RT) and mounted in antiquenching medium (for the identification of EGFP-transfected cells throughout the spinal cord) or used for immunofluorescence (for the phenotyping of transplanted cells). To detect transplanted cells in the host tissue and to identify their phenotype, spinal cord cryo-sections were subjected to double immunofluorescent staining. For all sections, antigen retrieval was performed by immersion into 0.01 M of sodium citrate solution (pH: 9.0), heated to 70°C in a water bath for 30 min. 5 After cooling of the slides to RT, blocking of non-specific binding sites was performed using PBS containing 0.2% Triton X-100, 0.02% sodium-azide and 5% normal goat serum for one hour at RT. Primary antibodies (Table 2) were applied for three days at 4°C following washing in PBS (3 × 15 min at RT) and incubation with goat anti-mouse IgG conjugated with Cy3 and goat anti-rabbit IgG conjugated with Cy2 fluorescent dyes (dilution 1:200) for two hours at RT. After subsequent washing in PBS, cell nuclei were stained for 20 min at RT with a bis-benzimide solution (Hoechst-33258 dye, 5 μg/mL in PBS) (Sigma), washed again in PBS, mounted in antiquenching medium and examined with a laser scanning microscopy (Zeiss Axiovert 200 M).
Primary antibodies used in immunofluorescent staining
Results
Generation of dual expression cassette plasmids
Various dual expression cassette plasmids were generated using standard genetic engineering techniques utilizing the plasmid backbone pBudCE4.1. Some of the plasmids were constructed to contain a reporter gene egfp in order to facilitate their tracking. Efficient expression of EGFP by the resulting vectors pBud-EGFP, pBud-EGFP-VEGF and pBud-EGFP-FGF2 was confirmed by fluorescent microscopy (data not shown). Finally, a plasmid pBud-VEGF-FGF2 was constructed, which carries two therapeutic genes, VEGF and FGF2, under the control of two independent constituently active promoters, thus resulting in efficient simultaneous and independent expression of recombinant proteins. To confirm expression of VEGF and FGF2, hUCB-MCs were electroporated with pBud-VEGF-FGF2 plasmid, followed by realtime PCR analysis of mRNA expression (Figure 1). A significant increase in the mRNA expression level was achieved in transfected hUCB-MCs compared with untransfected control cells. An increase in VEGF mRNA expression over 7 × 104-fold and FGF2 mRNA over 1 × 104 times was observed after electroporation with pBud-VEGF-FGF2 plasmid.
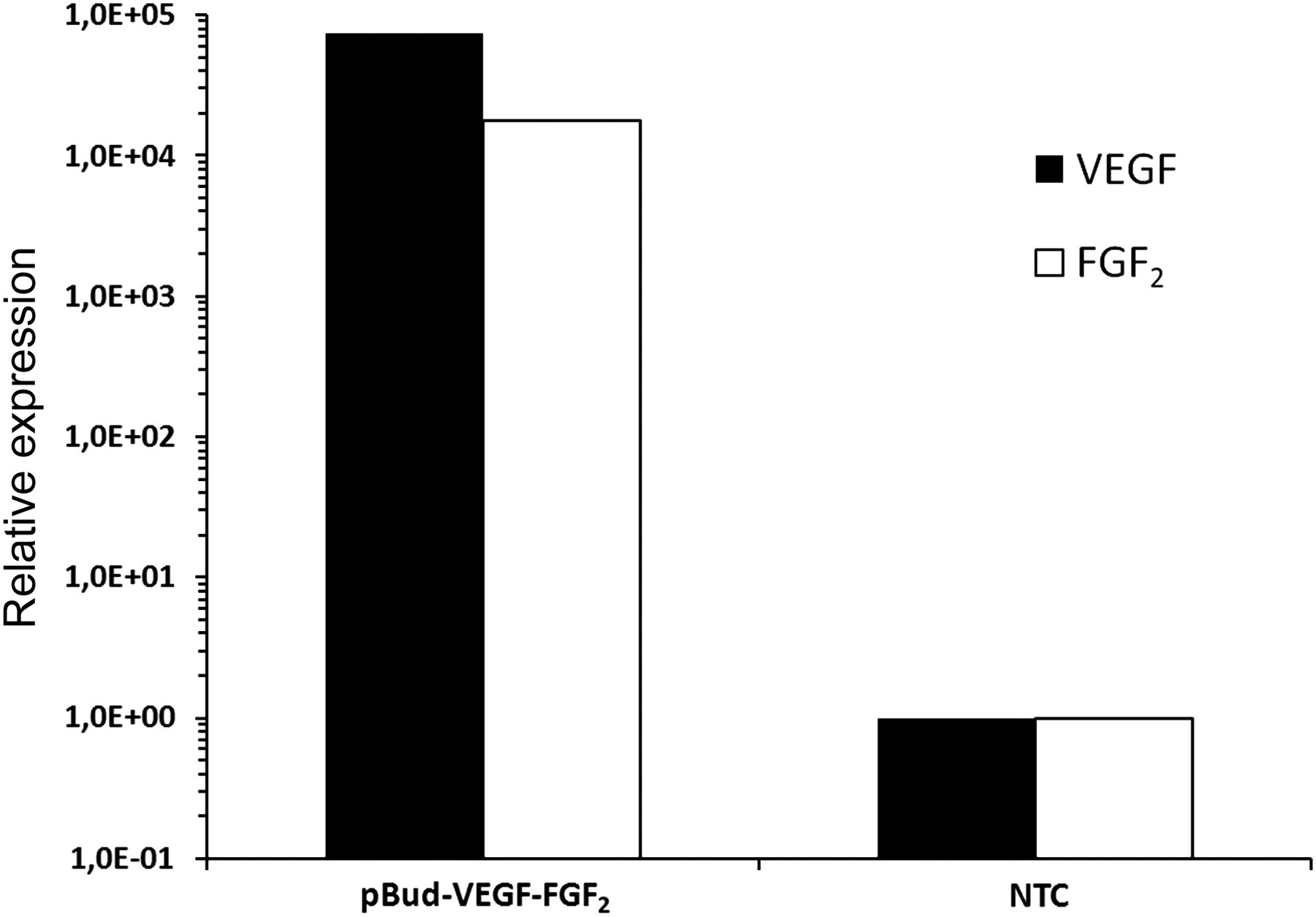
Realtime polymerase chain reaction analysis of human VEGF and FGF2 mRNA expression levels in human umbilical cord blood mononuclear cells (hUCB-MCs) electroporated with plasmid pBud-VEGF-FGF2 or non-transfected control (NTC). The level of expression of VEGF and FGF2 mRNA in NTC was considered 100%. Expression of VEGF mRNA in hUCB-MCs increased over 7 × 104 times and FGF2 mRNA over 1 × 104 times after electroporation with plasmid pBud-VEGF-FGF2. VEGF, vascular endothelial growth factor; FGF2, fibroblast growth factor 2
Identification of hUCBCs in G93A mice spinal cord
To identify transplanted cells in the spinal cords of control and experimental animals, we carried out immunofluorescent staining with an antibody against human nuclear antigen (HNA) as a marker for hUCBCs. To confirm the specific binding of the anti-HNA antibody we also applied standard nuclei staining (see subsection Immunofluorescence). Confocal microscopy has shown co-localization of HNA expression with nuclei staining. HNA+ cells were randomly distributed in the white and gray matter of the spinal cord 14 d after transplantation (Figure 2). Of note, HNA+ cells displayed various nuclear shapes: in particular, both oval and round nuclei of transplanted cells were observed throughout the spinal cord. No HNA+ cells were detected in animals treated with saline used as a negative control in our experiments. As such, these data confirm that retro-orbitally injected hUCBCs survive and migrate through the white and gray matters of the spinal cord in presymptomatic 24–28-week-old G93A mice.
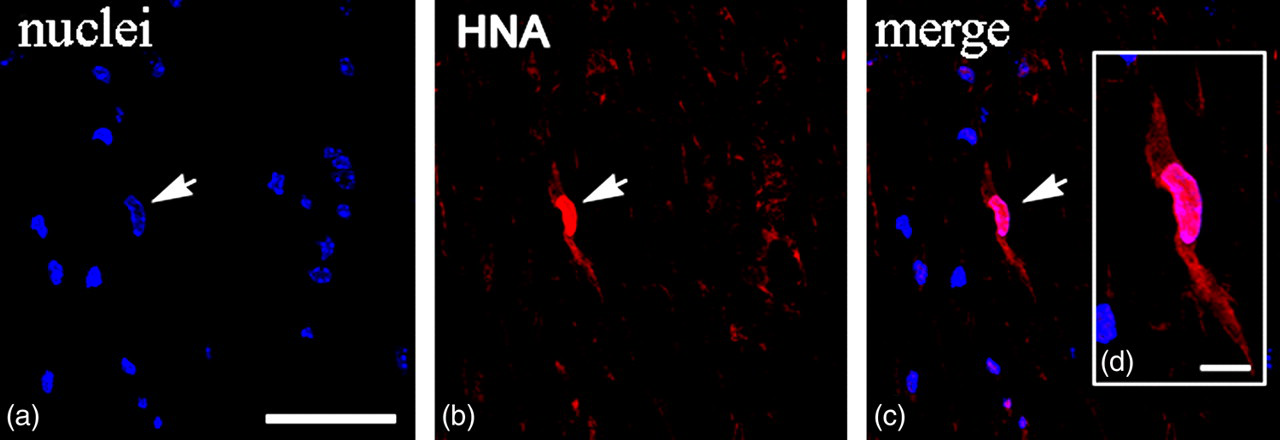
Spinal cord of presymptomatic 24-week-old G93A mice injected retro-orbitally with enhanced green fluorescent protein-transfected human umbilical cord blood cells (hUCBCs). Fourteen days after transplantation, hUCBCs were identified with antibodies against human nuclei antigen (HNA). (a) Nuclei staining (blue); (b) HNA labeling (red); (c) merged image representing HNA-positive hUCBC; and (
Expression of plasmid vectors in hUCBCs after transplantation
The transcriptional activity of plasmid vectors was evaluated by examining the expression of reporter EGFP plasmid vector in the transplanted cells. To visualize EGFP expression, an appropriate filter set and laser-491 were used in the confocal microscope. EGFP-expressing cells were observed in the spinal cords of mice transplanted with hUCBCs transfected with plasmids encoding EGFP alone, or in combination with FGF2 or VEGF (Figure 3). These EGFP-expressing cells are roughly 10 times smaller when compared with neurons. These cells were infrequently also distributed both in the white and gray matter of the spinal cord. Even though the expression of EGFP in cells was detected 14 d after transplantation, EGFP expression may not always be detectable after two weeks due to the elimination of plasmid vectors.
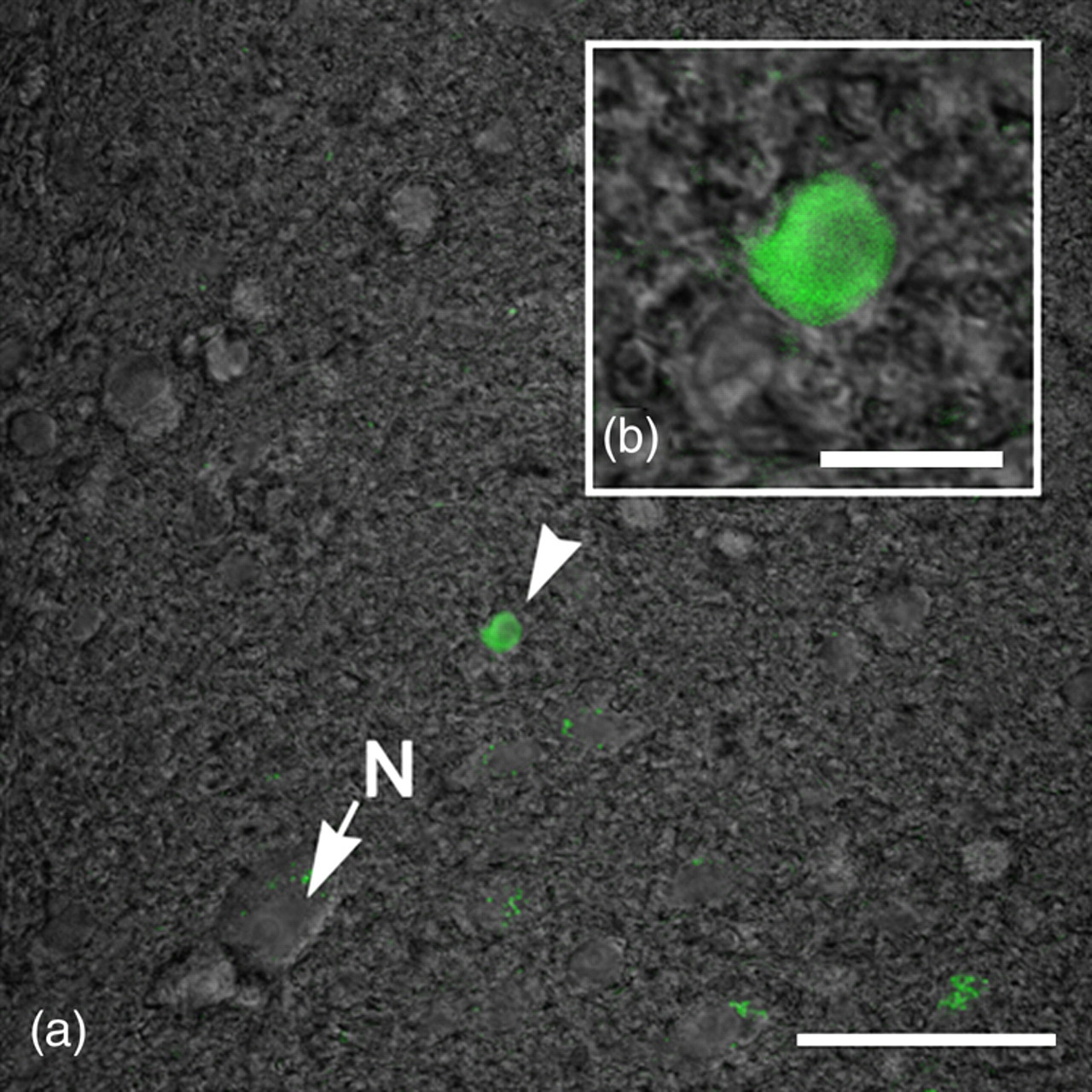
Enhanced green fluorescent protein (EGFP)-expressing human umbilical cord blood cells (hUCBCs) in the spinal cord of G93A mice 14 d after transplantation. EGFP expression is indicated by arrow-head (green): EGFP-transfected cell. Neuron is indicated by arrow (N). Scale bars: (a) 50 μm; (b) 10 μm (A color version of this figure is available in the online journal)
Phenotyping gene-modified hUCBCs in the spinal cord 14 d after transplantation
In order to evaluate which cell types the transplanted cells had differentiated into after 14 d, phenotyping of HNA+ cells with markers for neurons, glial, microglial and epithelial cells was performed. To identify hUCBC phenotype, spinal cord sections were subjected to double immunofluorescent staining. Anti-HNA antibody was used in combination with one of the following markers: neuron-specific β 3-tubulin, oligodendrocyte-specific protein, S100, iba1 and CD34. In the control group of mice subjected to transplantation with genetically modified hUCBCs expressing EGFP, double staining revealed HNA+ cells expressing microglial marker iba1 (Figure 4) and the endothelial cell marker CD34 (Figure 5). The iba1+ hUCBCs possess a small oval cell body with numerous processes of small diameter: a characteristic feature of microglial cells. Expression of the astrocyte marker S100 by HNA+ cells was found in the spinal cord of mice after transplantation with the genetically modified hUCBCs over-expressing VEGF-FGF2 (Figure 6). S100+ cells displayed astrocyte-like morphology with many branching processes. Neither the neuronal β 3-tubulin nor the oligodendrocyte-specific protein were expressed in the hUCBCs in any control or experimental rodents.
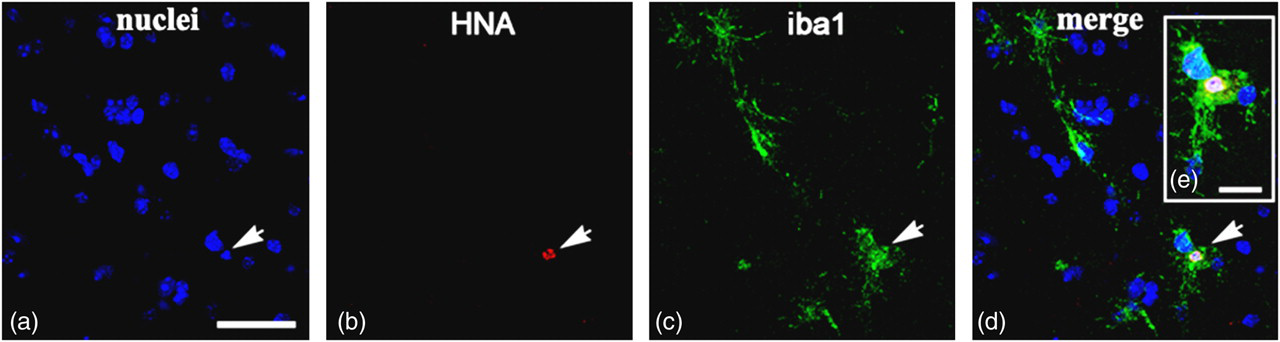
Enhanced green fluorescent protein (EGFP)-transfected human umbilical cord blood cells (hUCBCs) in the spinal cord of G93A mice 14 d after transplantation. Co-localization of human nuclei antigen (HNA) and iba1 in hUCBCs is shown. The iba1+ cells with ramified processes are depicted, which are characteristic features of microglial cells. Arrows on panels a–d indicate HNA+ nucleus in iba1+ cells. (a) Nuclei staining (blue); (
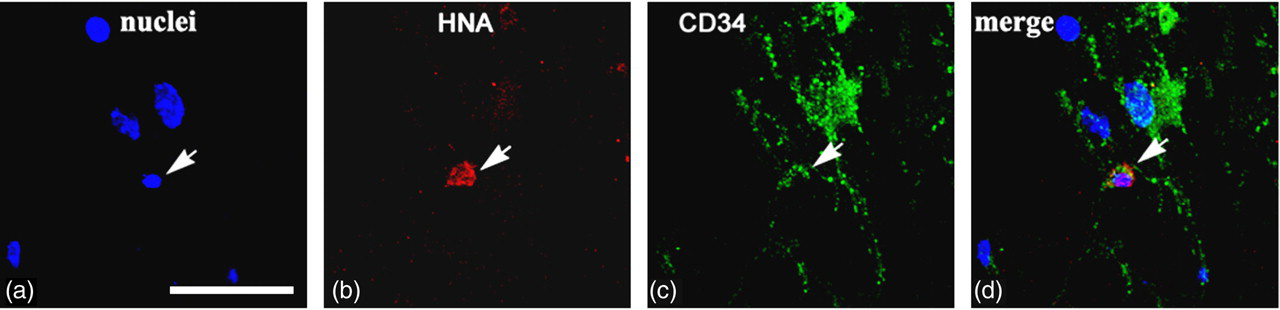
Enhanced green fluorescent protein (EGFP)-transfected human umbilical cord blood cells (hUCBCs) in the spinal cord of G93A mice 14 d after transplantation. Co-localization of human nuclei antigen (HNA) and CD34 (marker for endothelial cells) is shown. Arrows on panels a–d depict HNA + /CD34+ cells. Scale bar: (a
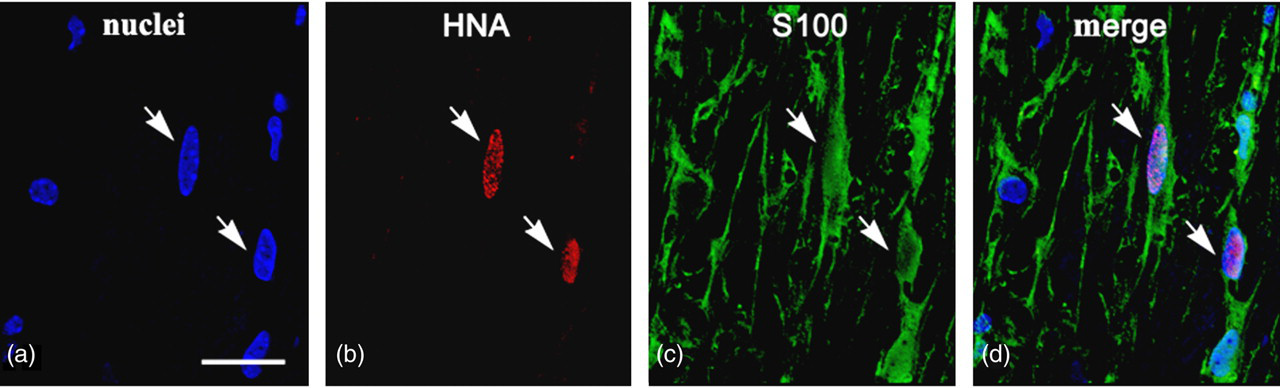
VEGF-FGF2-transfected human umbilical cord blood cells (hUCBCs) in the spinal cord of G93A mice 14 d after transplantation. Co-localization of human nuclei antigen (HNA) and S100 is presented. HNA + /S100+ cells have numerous projections: a common feature of astrocytes. Arrows on panels a–d show HNA + /S100+ cells. Scale bar: a–d 25 μm (A color version of this figure is available in the online journal)
Discussion
Recent advances in biotechnology have provided the potential to use combined gene and stem cell therapy in various neurodegenerative disorders, such as Alzheimer's, Parkinson's, Huntington's, Lou Gehrig's disease, as well as several others caused by multiple genetic and environmental factors resulting in neuronal cell death. Cells for transplantation in basic and clinical trials may be obtained from embryonic, fetal or adult tissues that can be predifferentiated into neurons or glias. 6–10 However, the use of embryonic or fetal organs is highly restricted based on ethical and religious controversies. Stem cells prepared for transplantation must have predictable and reproducible characteristics, but at the same time the high risk of tumor transformation or immunological conflict must also be taken into consideration. Alternative sources of stem and progenitor cells are mesenchymal stem cells of the bone marrow, 11,12 hematopoietic and mesenchymal stem cells from hUCB, 13,14 stem cells of tooth soft tissues, 15 and olfactory ensheating cells. 16 Gene modification of stem cells may significantly enhance their therapeutic effect by addressed homing, increased survivability, secretion of growth and trophic factors and directed terminal differentiation. Therapeutic growth factors transiently over-expressed by transplanted cells may have autocrine, paracrine and endocrine regulatory mechanisms. Autocrine effects may increase cell survivability, enable control of addressed homing and migration, and modulation of terminal differentiation. Paracrine and endocrine regulation of the transplanted cells may have trophic and cytoprotective effects on the affected parenchymal cells at the site of degeneration by delivering growth and trophic factors to the damaged tissue in order to support cell survival and tissue regeneration. 17,18
In our previous investigations we have demonstrated that genetically modified (VEGF) hUCBCs migrate in the spinal cord parenchyma, proliferate and differentiate into endothelial cells to form new blood vessels in ALS mice. Several similar data have shown that hUCBCs, genetically modified with human VEGF, can induce angiogenesis in limb ischemia rat models, 19 as well as in myocardial infraction in mice. 20 In the present study we generated hUCBCs over-expressing human VEGF, human FGF2, or simultaneously both factors. Our results demonstrate that hUCBCs injected retro-orbitally in presymptomatic 24-week-old G93A mice can undergo guided differentiation into microglial, astrocytic or endothelial cells, but not neurons, based on their pretransplantational gene-modification. Thus, EGFP-transfected cells might be transformed into microglia-like (iba1+) or endothelial-like (CD34+) cells; however VEGF-FGF2 transfection stimulates them to differentiate into astrocyte-like (S100+) cells.
In the central nervous system, VEGF is produced by neurons, glias and neural stem cells, 21–23 and has neuro-trophic effects. Axotomy increases expression of VEGF and its nuclear receptor flt-1 in lumbar spinal cord motor neurons. 24 Recent studies have also shown links between VEGF and ALS. 25 Reduced expression of VEGF results in ALS-like progressive degeneration of motor neurons. 26,27 Injection of VEGF-expressing lentiviral vector into various muscles delayed onset and slowed progression of ALS in G93A mice. 28 The rationale for FGF2 gene modification of hUCBCs is motivated by its involvement in the differentiation of various cell types via specialized signaling systems. Moreover, FGF2 is a growth factor with a significant neuroprotective effect. Following neurotrauma, the number of FGF2-positive cells in the spinal cord is increased up to 15 times, 29 suggesting that FGF2 is an important factor in molecular and cellular mechanisms of neuroregeneration. The positive influence of FGF2 is partially due to its ability to block caspase-3 gene expression and cell apoptosis at the stage of secondary degeneration, 30 which supports regeneration of neurites and restoration of spinal cord function. FGF2 is strongly implicated in neuro- and gliogenesis from stem cells. Up-regulation of neurogenic gene expression with FGF2 promotes the migration and maturation of neurons derived from transplanted neural stem cells. 31 FGF2 and its receptors contribute to the creation of an important microenvironmental niche that promotes neurogenesis in the adult and aged brain. 32 Exposure to FGF2 selectively induces astrocyte precursors to proliferate, forming clusters of vimentin-positive cells, which, within two weeks, differentiate into astrocytes. 33 FGF2 promotes conversion of preoligodendrocyte progenitor cells 34 and inducts oligodendrocyte progenitors in the dorsal forebrain. 35 Neural stem cells cultured with FGF2/epithelial growth factor (EGF) generate neurospheres, which include cells expressing glial markers. 36
Various stem and progenitor cells are present in hUCB-MCs. Culturing or transplantation of hUCBCs results in their differentiation into various cell types; they may give rise to any blood cells, macrophages, endothelial or several mesenchymal cells, etc. 37 Gene modification may result in directed differentiation of hUCBCs into any needed cells. Indeed, we have previously demonstrated differentiation of gene-modified hUCBCs into endothelial cells in G93A mice. Plasmid L1 cell-adhesion molecule (L1CAM) and VEGF vectors resulted in addressed homing and transformation of transplanted cells into endothelial cells. In the present investigation we have documented differentiation of hUCBCs into astrocytes: 14 d after transplantation into G93A mice, hUCBCs over-expressing VEGF and FGF2 turned into S100+ cells with elongated bodies and extended processes. Self-differentiating potential, modeling the effects of VEGF and FGF2 on hUCBCs via autocrine regulation and signal molecules from dying motor neurons in spinal cord possibly created the unique condition and microenvironment for differentiation of hUCBCs into astrocytes. In terms of gene–stem cell therapy of ALS, astrocytic enrichment is known to have neuroprotective effects in motor neuron diseases as it has been demonstrated that astrocyte dysfunction occurs in human ALS and G93A mice and transplantation of lineage-restricted astrocyte precursors provides therapeutic effect in animal models of ALS. 38 Apart from the beneficial effects of the elevated number of astocytes per se, our gene-modified newly formed astrocytes also enriched the local concentration of VEGF and FGF2 at the site of neurodegeneration, which may further increase survivability of spinal motor neurons. Although our findings have a clinically exploitable potential, further studies are required to address survival of motor neurons, as well as functional benefits of hUCBC transplantation.
Footnotes
ACKNOWLEDGEMENTS
This study was supported by the Russian Foundation for Basic Research grants 08-04-01680 and 10-04-01423, Russian Federal Agency for Science and Innovations government contract FCP 02.740.11.0302, and by Asklepios-Med (Hungary). Some of the experiments were conducted with support of the Regional Center of Collective Use, Kazan State University (Kazan, Russia). AAR was supported in part by a NATO reintegration grant NR.RIG.983007.