
Abstract
Autologous chondrocyte implantation (ACI) is widely used for the repair of cartilage defects. However, due to the lack of chondrocyte growth factor and dedifferentiation of the cultured primary chondrocytes, cell source has limited the clinical potential of ACI. Auricular cartilage is an attractive potential source of cells for cartilage tissue engineering. Here we demonstrated that recombinant human midkine (rhMK) significantly promoted proliferation of rat primary auricular chondrocytes cultured and passaged in monolayer, which was mediated by the activation of mitogen-activated protein kinase and phosphoinositide 3-kinase pathways. Furthermore, rhMK attenuated the dedifferentiation of cultured chondrocytes by maintaining the expression of chondrocyte-specific matrix proteins during culture expansion and passage. Importantly, rhMK-expanded chondrocytes reserved their full chondrogenic potential and redifferentiated into elastic chondrocytes after being cultured in high density. The results suggest that rhMK may be used for the preparation of chondrocytes in cartilage tissue engineering.
Introduction
Adult human cartilage shows a poor capacity for repair and regeneration. Autologous chondrocyte implantation (ACI) is widely used for the treatment of patients with cartilage damage. 1 This approach involves harvesting biopsies of cartilage, isolating chondrocytes from the donor tissue and expanding the cells in vitro. 1 However, chondrocytes are limited in number, comprising only 5–10% of cartilage tissue and dedifferentiation of chondrocytes is the main problem accompanying prolonged cultivation in monolayer. 1,2 Thus, obtaining sufficient cartilage biopsies and getting enough chondrocytes multiplied in vitro is crucial for the success of ACI.
Auricular cartilage is an elastic cartilage found in the ear and epiglottis, which, in contrast to hyaline cartilage, expresses elastin in addition to type II, IX, X and XI collagen and chondroitin sulfate proteoglycan. 2–4 Auricular cartilage is an attractive potential source of cells for many tissue engineering applications because the auricular chondrocytes are easily and safely obtained. 5 Auricular chondrocytes cultured in vitro have the potential not only for regeneration of cartilage in the auricle, nose and trachea, 4 but there are also indications for other tissue engineering applications, such as treatment of vesicoureteral reflux, 6 damaged articular cartilage 7 and prevention of further degeneration of intervertebral disc after removal of disc hernia. 5,8
Midkine (MK) is a member of the pleiotrophin (PTN)/MK family and was originally described as a retinoic acid inducible, developmentally regulated, heparin-binding, secreted neurotrophic factor. 9,10 Previous studies have mostly focused on MK effects in neurons and neurite outgrowth, 11 bone fracture healing, 12 neutrophil activation and osteoclast differentiation in rheumatoid arthritis (RA), 13 tumorigenesis and tumor progression, 14 cerebral infarction, 15 cardiac remodeling and live regeneration. 16,17 In the present study, we demonstrate that recombinant human midkine (rhMK) promotes proliferation of primary auricular chondrocytes and maintains the characteristics of chondrocytes through inhibiting dedifferentiation of the chondrocytes cultured in monolayer.
Materials and methods
rhMK protein expression and purification
A DNA sequence (GenBank NM_001012334) encoding the mature human MK (hMK) protein was expressed in Escherichia coli BL21 (DE3) using pET30a (+) vector (Novagen, Beijing, China) as described previously.
18
Briefly, the expression of rhMK was induced by isopropylthio-β-
Isolation, passage and high-density culture of auricular chondrocytes in monolayer
The animal experiments were approved by the Animal Care and Use Committee of Shanghai Jiaotong University (Shanghai, China). Rat auricular chondrocytes were isolated from auricular cartilage of eight-week-old male Sprague-Dawley rats as previously described, with minor modifications. 19 Briefly, eight rats were sacrificed and auricular cartilage was dissected from the perichondrium, and diced into small pieces. The cartilage specimens were treated with 0.25% trypsin solution containing 0.02% ethylenediaminetetraacetic acid (EDTA). The chondrocytes were released by digestion with 0.2% type II collagenase (Invitrogen, Shanghai, China) in Dulbecco's modified Eagle's medium (DMEM [high glucose]; Invitrogen) for 16 h and filtered through a 100-μm nylon filter (Falcon, Franklin Lakes, NJ, USA). Rat primary auricular chondrocytes were plated at a density of 1 × 104 cells/cm2 into six-well plates and cultured in DMEM containing 10% fetal bovine serum (FBS) with or without rhMK (3.0 μg/mL) at 37°C. The media were changed every two days. After seven days, primary chondrocytes (passage 0, P0) which reached subconfluence were recovered by incubation with 0.25% trypsin solution containing 0.02% EDTA. The cells were cultured for two additional passages (P1 and P2) with three days' culture duration for each passage under the same culture conditions as the primary culture (P0). The viable cell number of initial plated (N i) and final recovered (N f) in each passage were determined in a blood cell counting chamber by trypan blue dye exclusion and the cell expansion fold of each passage was calculated by N f/N i. The cumulative cell expansion fold was calculated as the product of the cell expansion folds at successive passages. For high-density culture, the dedifferentiated auricular chondrocytes cultured with or without rhMK in monolayer after three passages were recovered, plated at 4 × 105 cells per well in 24-well plates and cultured in DMEM containing 10% FBS. After the cells reached confluence, they were cultured for an additional two weeks without passage. The medium was changed every two days.
Bromodeoxyuridine incorporation assay of primary auricular chondrocyte proliferation
Rat primary auricular chondrocytes were plated at a cell density of 1 × 104 cells/well in 96-well plates and cultured in DMEM containing 10% FBS in the presence of different concentrations of rhMK (0, 0.1, 0.3, 1.0, 3.0, 9.0 μg/mL) for 48 h, then bromodeoxyuridine (BrdU) reagent was added and incubated for four hours. Following partial denaturation of double-stranded DNA, BrdU was detected immunochemically, allowing the assessment of the population of chondrocytes in DNA replication (Millipore, Billerica, MA, USA). The absorbance was measured at 450 nm using the ELX-800 ELISA plate reader (Bio-Tek Instruments, Winooski, VT, USA) according to the manufacturer's instructions. For observation of the effect of the mitogen-activated protein kinase kinase 1/2 (MEK1/2) and phosphoinositide 3-kinase (PI3K) pathway on the rhMK-stimulated proliferation of primary auricular chondrocytes, primary auricular chondrocytes were cultured in DMEM containing 10% FBS and rhMK (3.0 μg/mL). The inhibitor of MEK1/2, PD98059 (2.8, 8.3 and 25 μmol/L), or the PI3K inhibitor, LY294002 (1.1, 3.3, and 10 μmol/L), was added. After culture for 48 h, the cells were labeled with BrdU for an additional four hours followed by the quantification of BrdU-incorporated cells.
Analysis of auricular chondrocyte dedifferentiation and redifferentiation by reverse transcription-polymerase chain reaction
Total RNA was extracted from cultured chondrocytes (P0, P1, P2, high-density culture) using TRIzol reagent (Invitrogen). Isolated RNA (1 μg) was reverse transcribed and then amplified using the reagents from a commercial kit (Takara, Dalian, China) according to the manufacturer's instructions. Following an initial denaturation step of two minutes at 95°C, amplification consisted of 24–30 cycles of 30 s at 95°C, 30 s at optimal annealing temperature (AT) and 30 s at 72°C. The primers and conditions for reverse transcription-polymerase chain reaction analyses were as follows: Agc (AT 54°C, 30 cycles), forward CTACGACGCCATCTGCTACA and reverse GCTTTGCAGTGAGGATCACA; Col2a1 (AT 54°C, 24 cycles), forward GAATGGCTGACCTGACCTGATA and reverse GGCGTCTGACTCACACCAGATA; Col1a1 (AT 50°C, 24 cycles), forward GGGCAAGACAGTCATCGAATA and reverse ATGTCCATTCCGAATTCCT; elastin (AT 54°C, 30 cycles), forward TCCGACGACTTCTCACACTG and reverse GGTTGATAGTGGGTGTGCCT; Col10a1 (AT 54°C, 30 cycles), forward TCTAAAGTGCACTCCGGGAC and reverse TAAGGATGGGACGACAGGAG; β-actin (AT 54°C, 24 cycles), forward GAGGCATCCTGACCCTGAAG and reverse CATCACAATGCCAGTGGTACG. Polymerase chain reaction (PCR) products were separated on 1.5% (w/v) agarose gels and quantified after staining with ethidium bromide. The density of each band was measured using the Fluor-S MultiImager (Bio-Rad, Hercules, CA, USA) with attached software program to generate the semi-quantitative expression data. The data were normalized to the internal control β-actin signals for comparison among the experiments.
Western blot analysis of mitogen-activated protein kinases, Akt, type II collagen and aggrecan
To detect signal pathways during the proliferation of rhMK-treated auricular chondrocytes, rat primary auricular chondrocytes stimulated with or without rhMK (3.0 μg/mL) were lysed in RIPA buffer containing proteinase inhibitors (Beyotime, Wuhan, China) at predeterminate time points (0, 30, 60, 120 min). Total protein extracts (15 μg) were resolved by SDS-PAGE, electroblotted onto polyvinylidene difluoride membranes. The membranes were blocked by 5% non-fat milk, and probed with 1:1000 dilution of primary antibodies (extracellular signal-regulated kinase [ERK]1/2, phospho-ERK1/2, p38, phospho-p38, c-Jun N-terminal kinase [JNK], phospho-JNK, Akt and phospho-Akt; all from Cell Signaling Technology, Boston, MA, USA) overnight at 4°C. Antibodies were detected using the reagents from the enhanced chemiluminescence kit as described by the manufacturer (Pierce, Rockford, IL, USA). For detecting the effect of rhMK on the dedifferentiation and redifferentiation of auricular chondrocytes cultured in monolayer, P1 and high-density culture auricular chondrocytes were lysed in RIPA buffer; type II collagen and aggrecan expression were detected using Western blotting as described above. Protein extracts (30 μg) was resolved by SDS-PAGE. Antibodies for detecting type II collagen, aggrecan and internal control β-actin (Santa Cruz Biotechnology, Santa Cruz, CA, USA) were diluted as 1:500, 1:300 and 1:1000, respectively. The density of each band was measured using the Fluor-S MultiImager (Bio-Rad) with attached software program to generate the semi-quantitative expression data. The data were normalized to the internal control β-actin signals.
Immunohistochemical staining for type II collagen and aggrecan
The immunohistochemical staining was carried out in the cell culture plate wells. Cell cultures (P0, P1 and high-density culture) were fixed with 4% paraformaldehyde and sequentially incubated in phosphate-buffered saline, hydrogen peroxide and methanol solution (50:1), and 5% bovine serum albumin solution. Subsequently, the cells were incubated with primary rabbit polyclonal antibodies to type II collagen (1:100) (Santa Cruz Biotechnology) and aggrecan (1:100) (Santa Cruz Biotechnology) for one hour, followed by goat anti-rabbit IgG antibody conjugated with biotin (1:100) (Boster, Wuhan, China), detected by avidin-conjugated horseradish peroxidase (Boster) and its substrate diaminobenzidine (DAB; Boster), and counterstained by hematoxylin. The stained cells were kept wet and photographed using an inverted microscope (Nikon, Shanghai, China).
Statistical analysis
Results are expressed as the mean ± standard deviation. Statistically significant differences were determined by two-tailed Student's t-test and one-way analysis of variance with SPSS 18.0 software (SPSS Inc, Chicago, IL, USA). Statistical significance was assumed for P values < 0.05.
Results
rhMK promotes the proliferation of auricular chondrocytes in monolayer cultures
To investigate the biological effects of rhMK on primary auricular chondrocytes, we expressed and purified the recombinant protein as described previously. 18 After successfully purifying the protein, we examined whether rhMK enhances the proliferation of rat auricular chondrocytes in a primary monolayer culture. The results showed that rhMK increased DNA synthesis of the chondrocytes in a dose-dependent manner as measured by the BrdU incorporation assay (Figure 1a). rhMK increased DNA synthesis by 18%, 29%, 37% and 45% over the negative control in the 0.1, 0.3, 1.0 and 3.0 μg/mL rhMK dose groups, respectively. The maximal response was observed at the 3.0 μg/mL rhMK dose group, and the response was maintained even at 9.0 μg/mL rhMK dose group. Using rhMK at 3.0 μg/mL, we further determined the proliferation rate of the chondrocytes by sequential passaging of the cultured cells. The chondrocytes were expanded for three passages (P0, P1 and P2) in culture medium DMEM containing 10% FBS. The fold of cell expansion was calculated and compared between the cultures supplemented with or without rhMK (3.0 μg/mL). rhMK promoted expansion of the chondrocytes in each passage (P0–P2). The percentages of proliferation increase by rhMK (3.0 μg/mL) over the control were 65%, 65% and 55% in passage 0, 1 and 2, respectively (Figure 1b). Cumulative expansion showed that during the entire culture period (P0–P2), rhMK stimulated the expansion of auricular chondrocytes in the monolayer by 1042-fold compared with the control cultures of 247-fold (Figure 1c). Taking the above results together, rhMK stimulates the proliferation of auricular chondrocytes in vitro.
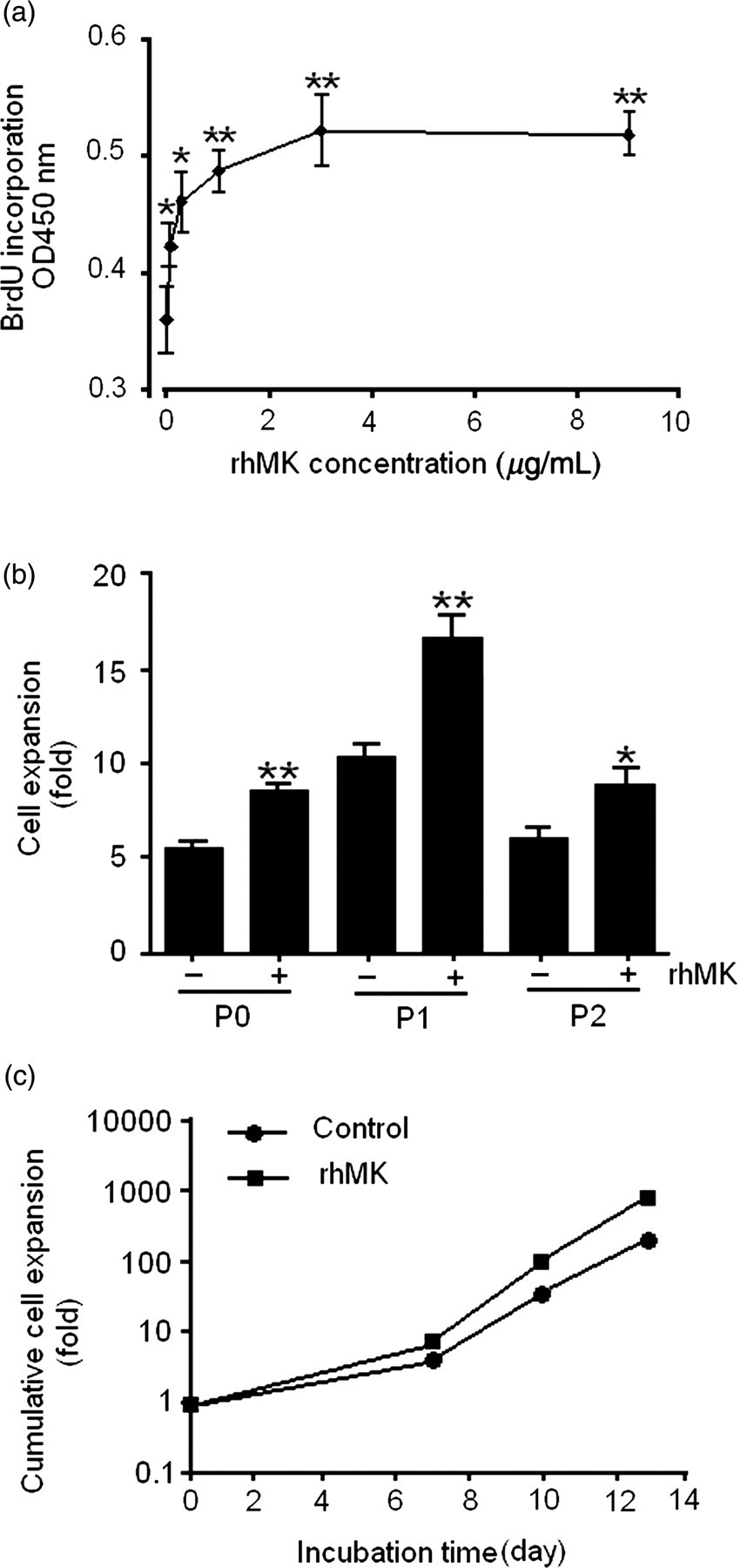
rhMK stimulates the proliferation of rat auricular chondrocytes. (a) rhMK stimulated the proliferation of rat primary auricular chondrocytes in a dose-dependent manner. Freshly isolated chondrocytes were treated with rhMK at indicated concentrations for 48 h. The cells were labeled with BrdU followed by the quantification of BrdU incorporated cells. (b) The proliferative role of rhMK on different passages. Rat auricular chondrocytes was cultured and passaged as described in the Materials and methods section. The cell expansion fold of each passage was calculated by N f/N i. (c) Cumulative cell expansion of rat auricular chondrocytes during the entire culture period (P0–P2) was calculated as the product of the cell expansion folds at successive passages. Data in (a) and (b) are expressed as the mean ± SD from triplicate cultures. *,**Values were compared with their respective control without rhMK using one-way analysis of variance (a) and Student's t-test (b). *P < 0.05, **P < 0.01. N i, viable cell number of initial planted in each passage; N f, viable cell number of final recovered in each passage; rhMK, recombinant human midkine; BrdU, bromodeoxyuridine
ERK and Akt activation mediates the proliferation of primary auricular chondrocytes by rhMK
To investigate the molecular mechanisms underlying the effects of rhMK on primary auricular chondrocytes, the phosphorylation status of mitogen-activated protein kinases (MAPKs) (ERK1/2, p38, JNK) and Akt, which are involved in chondrocyte proliferation, 20,21 was examined by Western blot analysis. rhMK stimulated phosphorylation of ERK1/2 after 30 min following addition of rhMK to the culture of freshly isolated auricular chondrocytes, and the levels of phosphorylated ERK1/2 were maintained for 120 min (Figure 2a), while in contrast, phosphorylated p38 and JNK were not detected (data not shown). There were no significant changes in the levels of total ERK in the same samples as demonstrated using a polyclonal anti-ERK1/2 antibody which recognizes both activated and non-activated ERK1/2 (Figure 2a). Furthermore, Akt in the PI3K pathway was also phosphorylated after 30 min following the addition of rhMK (Figure 2a).
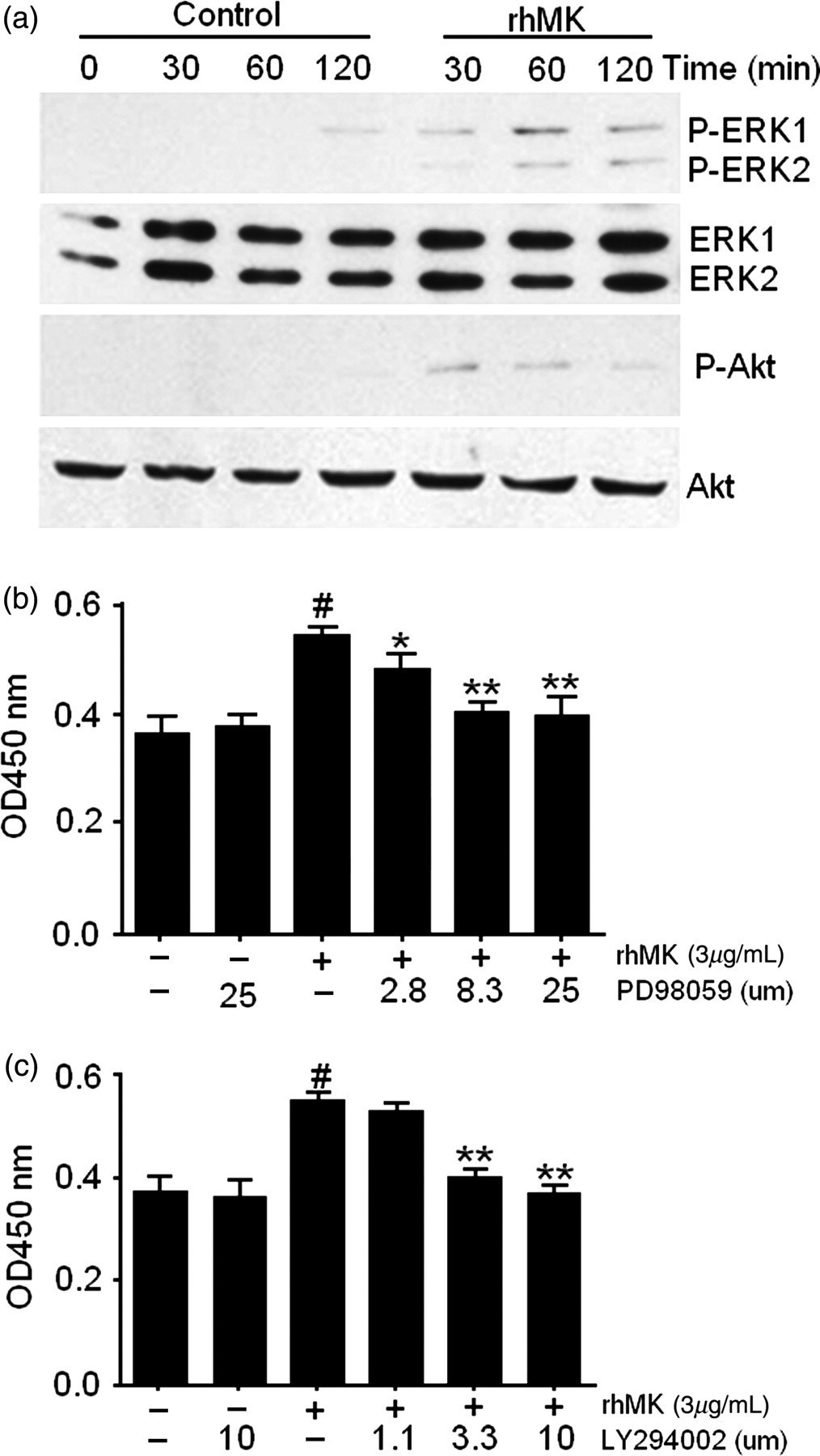
ERK and PI3K signaling pathways mediate the proliferation of primary rat auricular chondrocytes stimulated by rhMK. (a) The phosphorylation states of the indicated kinases in the auricular chondrocytes cultured in the presence of rhMK were examined by Western blot. Freshly isolated chondrocytes were cultured with and without rhMK (3.0 μg/mL) for the indicated time periods (30–120 min) and lysed. Immunoblottings with antibodies against phospho-ERK1/2, ERK1/2, phospho-Akt and Akt were performed. The experiments were repeated three times. (b, c) The MEK1/2 inhibitor PD98059 (b) and PI3K inhibitor LY294002 (c) inhibited the proliferative role of rhMK in the culture of primary auricular chondrocytes. Primary auricular chondrocytes were cultured with rhMK (3.0 μg/mL), PD98059 or LY294002 as indicated at various concentrations. After culture for 48 h, the BrdU assay was used for detecting the proliferation. All data are expressed as the mean ± SD from triplicate cultures. #Values were compared with their respective rhMK (−)/inhibitor (−) group using Student's t-test, *,**values were compared with their respective rhMK (+)/inhibitor (−) group using one-way analysis of variance. *P < 0.05, **P < 0.01, # P < 0.01. ERK, extracellular signal-regulated kinase; PI3K, phosphoinositide 3-kinase; rhMK, recombinant human midkine
Phosphorylation of ERK and Akt requires their upstream kinases of MEK1/2 and PI3K, respectively. 22,23 To correlate the rhMK-mediated ERK and Akt activation to the rhMK-proliferative role on auricular chondrocytes, we analyzed the effects of MEK1/2 and PI3K inhibition on the proliferation of primary auricular chondrocytes. The MEK1/2 inhibitor, PD98059 at a dose of 25 μmol/L, did not alter the proliferation of auricular chondrocytes without rhMK, but significantly attenuated the effect of rhMK on the proliferation of auricular chondrocytes. PD98059 inhibited the stimulation of BrdU incorporation by rhMK in a dose-dependent manner (Figure 2b). Likewise, LY294002, a PI3K inhibitor at a concentration of 10 μmol/L, inhibited the rhMK-induced chondrocyte proliferation, but not the cells without rhMK treatment. The inhibitory role of LY294002 was also dose-dependent (Figure 2c). Thus, we demonstrated that activation of ERK and Akt phosphorylation correlates to the rhMK-stimulated chondrocyte growth. Our data strongly suggest that ERK and Akt activation mediates the proliferative role of rhMK in auricular chondrocytes.
rhMK attenuates the dedifferentiation of auricular chondrocytes in monolayer culture
Chondrocytes are known for their phenotype changes from chondrocytic to fibroblastic when cultured in monolayer. 24,25 After defining the role of rhMK in promoting proliferation of chondrocytes in monolayer culture, we examined the phenotypes of culture-expanded auricular chondrocytes by semi-quantitative PCR. The mRNA expression levels of Col2a1 encoding collagen type II α1, Agc encoding aggrecan, Col1a1 encoding collagen type I α1, Col10a1 encoding collagen type X α1 and elastin, relative to the house-keeping gene β-actin, were determined for the auricular chondrocytes cultured for three passages (P0, P1 and P2) with or without rhMK. The results showed that the mRNA expression of Col2a1, Agc and elastin decreased dramatically over two passages without addition of rhMK, while the mRNA expression of Col2a1, Agc and elastin decreased slowly over two passages with addition of rhMK (Figures 3a–c). The percent increases of Col2a1 by rhMK (3.0 μg/mL) over control were 22%, 254% and 558% in passage 0, 1 and 2, respectively (Figure 3b). rhMK increased the elastin expression by 3.6- and 3.3-fold compared with the control in passage 0 and 1, respectively (Figure 3c). The expression of Col10a1, the major marker of chondrocyte hypertrophy, was slightly enhanced by rhMK in the primary chondrocytes (P0) (Figure 3d). The elevated expression of Col2a1 and Agc by rhMK (3.0 μg/mL) over control in P1 auricular cartilage was correlated well with their protein expression demonstrated by Western blot (Figures 3e and f) and immunohistochemical analysis (Figures 3g and h). Taken together, the results demonstrate that rhMK slows down the dedifferentiation process of auricular chondrocytes in monolayer cultures.
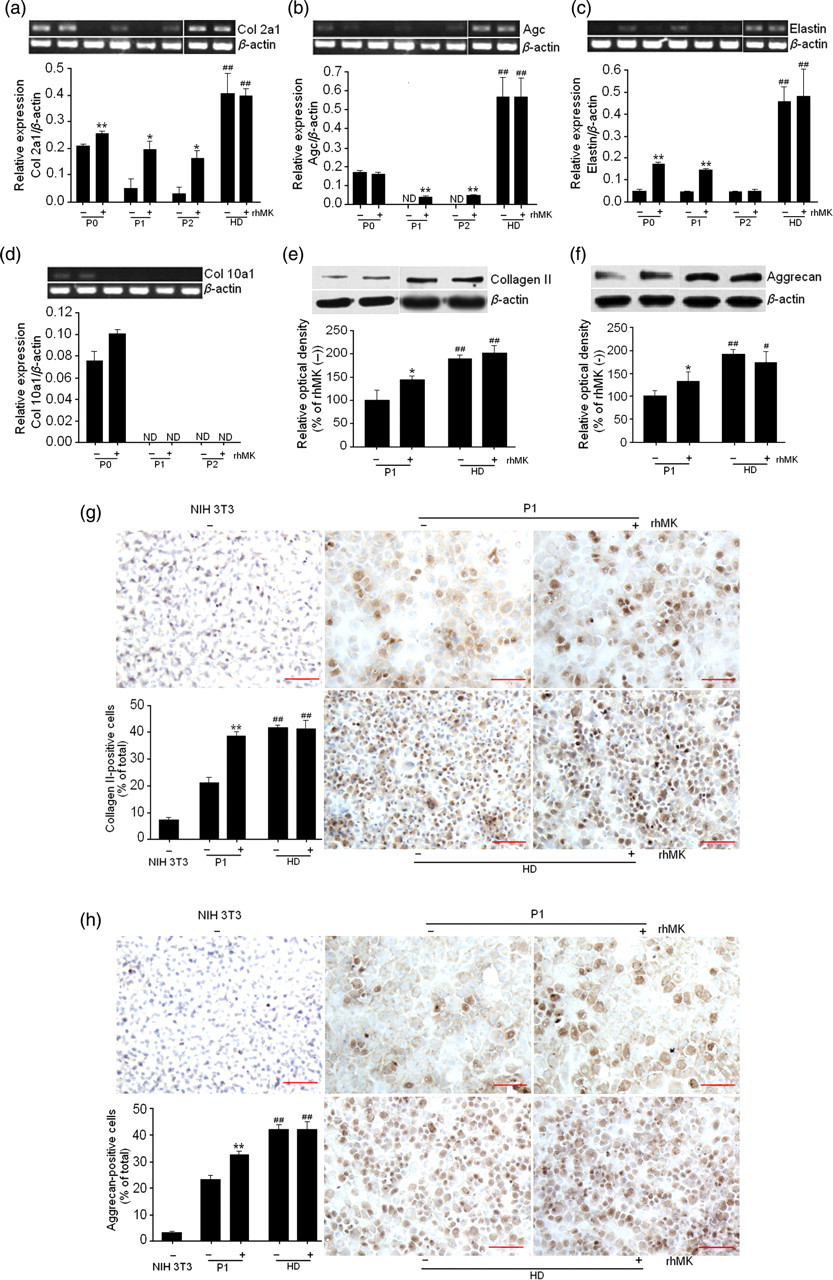
rhMK attenuates the dedifferentiation of auricular chondrocytes during passages and rhMK-expanded chondrocytes redifferentiate in high-density culture. (a–d) The expression of Agc (a), Col2a1 (b) and elastin (c) were maintained in the rhMK (+) group compared with their respective rhMK (−) group during passages (but not for Col10a1 [d]) and regained in the HD group. Rat auricular chondrocytes were cultured with or without rhMK (3.0 μg/mL) for three passages then subjected to high-density culture. The target gene mRNA was measured by semi-quantitative reverse transcription-polymerase chain reaction. The quantitative data are shown (n = 3). (e–h) Western blotting (e, f) and immunohistochemical analysis (g, h) of type II collagen and aggrecan in P1 and HD group. The quantitative data of Western blot and immunohistochemical analysis are shown (n = 3). Murine NIH 3T3 cell line was used as negative control for immunohistochemical analysis with the same culture conditions as the auricular chondrocytes. All data are expressed as the mean ± SD. *,**Values were compared with their respective control group (rhMK (−)) and #values were compared with P1 rhMK (−) group using two-tailed Student's t-test. *P < 0.05, **P < 0.01, # P < 0.05, ## P < 0.01. Scale bar 50 μm. HD, high-density culture of rat auricular chondrocytes expanded with or without rhMK (3.0 μg/mL) for three passages; rhMK, recombinant human midkine; ND, not detected. (A color version of this figure is available in the online journal)
Redifferentiation of rhMK-expanded chondrocytes in high-density culture
It is well known that expanded chondrocytes may re-express the cartilage-specific molecules during high-density culture in vitro. 26 As rhMK did not fully prevent dedifferentiation of the auricular chondrocytes in monolayer culture, we proposed that the rhMK-expanded cells may still retain their redifferentiation potential like the fully dedifferentiatiated chondrocytes cultured without rhMK. After three passages of the primary chondrocytes cultured with and without rhMK, they were subjected to high-density cultures as described in the Materials and methods section and the expression of cartilage-specific markers was analysed. Comparing with the dedifferentiated P1 auricular chondrocytes cultured without rhMK, auricular chondrocytes cultured in monolayer with and without rhMK regained higher expression of Col2a1, Agc and elastin genes after they were cultured in high density (Figures 3a–c). The expression of type II collagen and aggrecan in the high-density cultured auricular chondrocytes was also detected by Western blot (Figures 3e and f) and immunohistochemical staining (Figures 3g and h) and showed similar results to gene expression. There were no differences in the expression levels of the cartilage-specific markers between the cells pretreated with or without rhMK. We also performed the culture-expanded auricular chondrocytes in 3D culture, which was close to the condition of chondrocytes in vivo. The results showed that rhMK-expanded chondrocytes also re-expressed chondrogenic markers after 3D culture, and has no differences with or without rhMK, consistent with the results in high-density culture (data not shown). Thus, the rhMK-expanded auricular chondrocytes in monolayer culture retain their redifferentiation ability in subsequent high-density cultures.
Discussion
We demonstrated in this study that rhMK stimulates the proliferation of auricular chondrocytes, and attenuates the dedifferentiation of primary auricular chondrocytes during serial passages in monolayer cultures. rhMK stimulated the proliferation of articular chondrocytes through enhancing their DNA synthesis, as measured by BrdU incorporation. The cellular proliferative role of rhMK was apparently mediated by the activation of MEK1/2 and PI3K signaling pathways, which were strongly suggested by the rhMK-dependent ERK and Akt phosphorylation and ERK/Akt-dependent activity of rhMK. Furthermore, rhMK attenuated the dedifferentiation of auricular chondrocytes, as demonstrated by the retaining of their expression of chondrocyte-specific markers of Agc, Col2a1 and elastin. Importantly, addition of rhMK in monolayer cultures did not affect the redifferentiation ability of dedifferentiated chondrocytes in subsequent high-density cultures (Figure 4).
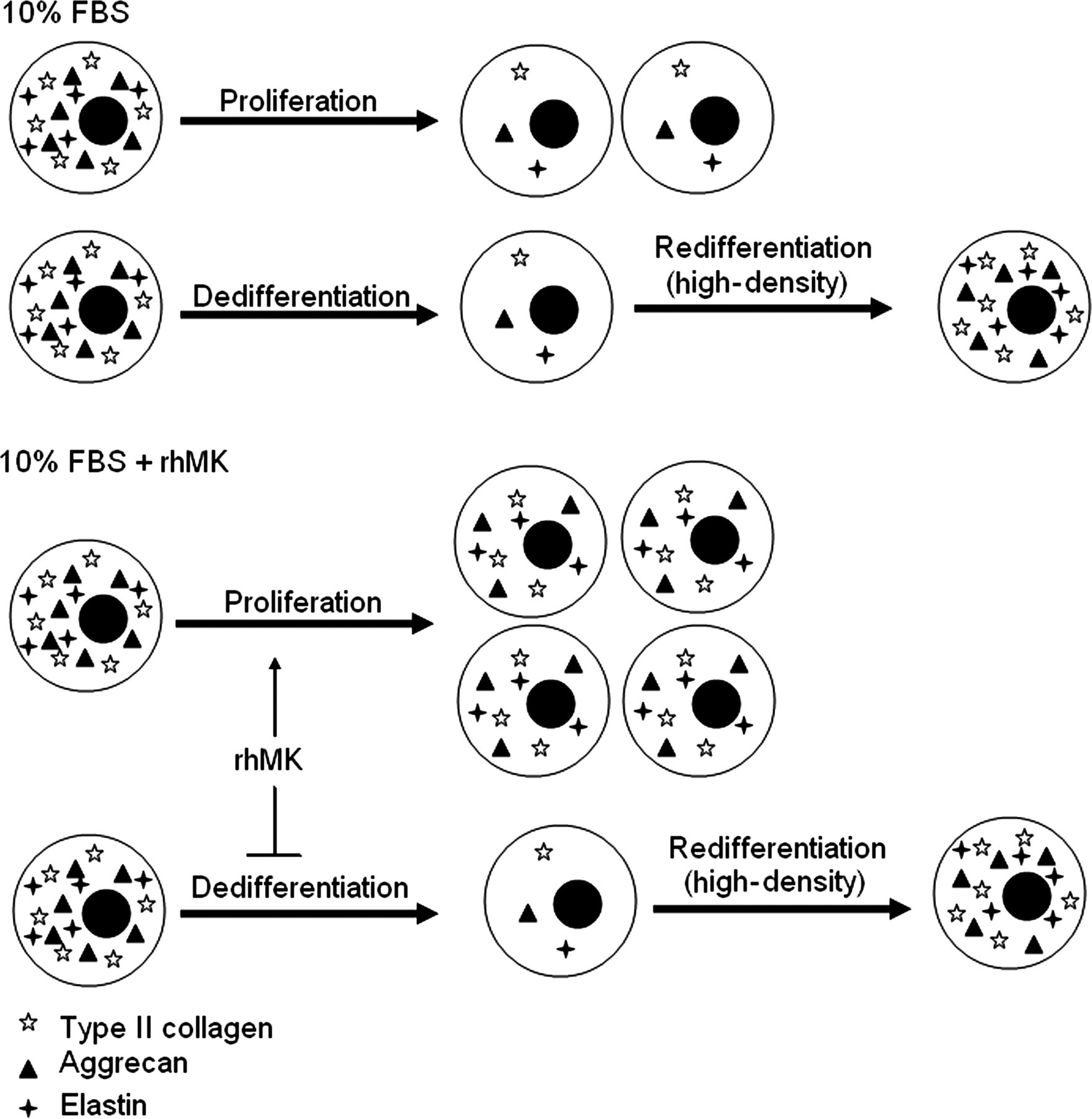
A schematic view of the roles of rhMK in auricular chondrocytes. The primary auricular chondrocytes cultured in monolayer is characterized by their proliferation accompanied by dedifferentiation quickly losing their cellular marker of collagen II and elastin. rhMK enhances the proliferation of the cells while it slows down their dedifferentiation. The rhMK-expanded cells in monolayer culture retain their redifferentiation potential when they are cultured in high density. rhMK, recombinant human midkine; FBS, fetal bovine serum
MK is known to activate several cellular signaling pathways in a number of different cell types. MK induced phosphorylation of MAPKs and PI3K, which was responsible for its mitogenic function and inhibition of apoptosis, in the culture of tumor cell lines, mouse neurons, ventricular myocytes and human primary endothelial cells. 27,28 Furthermore, phosphorylation of ERK contributed to the enhanced proliferation and inhibition of differentiation of primary chondrocytes cultured with fibroblast growth factor (FGF)-2 and FGF-18. 20 In this study, we provide new evidence that rhMK activates the phosphorylation of ERK and Akt, and utilizes the MEK1/2 and PI3K pathways to mediate its mitogenic function in primary auricular chondrocytes like that of other cell types.
A major problem in cartilage tissue engineering is to obtain sufficient cells and reduce chondrocyte dedifferentiation during monolayer culture expansion. Primary auricular chondrocytes cultured in a monolayer are known to lose their expression of extracellular matrix molecules and change in morphology. 24,29 To address this issue, growth factors were applied to the culture system. The application of growth factors (e.g. epidermal growth factor, FGF-2 and transforming growth factor β) not only accelerated the dedifferentiation of chondrocytes during passages, but also attenuated their ability of redifferentiation in subsequent pellet culture. 30 Martin et al. 31 found that FGF-2 greatly stimulated the proliferation of chondrocytes, fully maintained the redifferentiation ability in 3D culture, but accelerated the dedifferentiation in the monolayer. One possible explanation for the difference of redifferentiation ability is related to the subsequent 3D or pellet culture. In comparison to these growth factors, rhMK enhanced the expansion of auricular chondrocytes without affecting their redifferentiation potential; it slowed down their dedifferentiation while accelerating their proliferation. Other important problems of tissue engineering of elastic cartilage are undesired hypertrophy and mineralization of the repair tissue. In this study, we observed that the expression of type X collagen decreased over two passages, and rhMK did not affect the expression of type X collagen, consistent with the previous reports. 32 Therefore, rhMK may provide a powerful solution in the preparation of auricular chondrocytes for the clinical applications of cartilage tissue engineering.
The optimal cell source for cartilage tissue engineering is still being identified. 1 Chondrocytes and various stem cells have been explored for their potential as a viable cell source for cartilage repair. 33–35 Articular chondrocytes as a potential cell source for cartilage repair have been extensively investigated. 36 However, several impact factors, including the complicated procedure of harvesting articular cartilage with potential surgery risk, low cell yields, low mitotic rates and low bioactivity limit the use of articular chondrocytes. 1 Other potential autologous chondrocyte sources, such as auricular, nasoseptal and costal cartilage, are considered viable alternatives. 33–35 Among them, auricular cartilage is an attractive cell source since they have higher cell yield and faster cell proliferation rate in comparison to articular chondrocytes. 1 In addition, auricular chondrocytes produced neocartilage with better biochemical and histological similarity to that of native cartilage than articular counterparts when implanted in vivo. 37 Taken together, the auricular chondrocytes are favored over articular chondrocytes with the availability of the rhMK chondrocyte expansion system in the preparation of seed cells for cartilage tissue engineering.
The unique properties of rhMK in supporting proliferation while attenuating the dedifferentiation of auricular chondrocytes should aid in the preparation of a large quantity of chondrocytes for cartilage tissue engineering treating unmet cartilage defects.
Footnotes
ACKNOWLEDGEMENTS
This work was supported by the National Natural Science Foundation of China (81001388/H3004), Medicine-Engineering Collaboration Foundation of Shanghai Jiao Tong University (YG2010MS87), and National Comprehensive Technology Platforms for Innovative Drug R&D (2009ZX09301-007).