
Abstract
Heme oxygenase-1 (HO-1) potently influences tumor growth and metastasis. To date, no study has been performed on HO-1 expression pattern and its clinicopathological significance in human gastric cancer (GC) cases. In this study, the expression of HO-1 in human GC tissues (n = 74) and matched non-tumoral adjacent parenchyma (n = 46) was investigated by immunohistochemistry. The correlation of HO-1 with the clinicopathological characteristics was analyzed. Results showed that HO-1 was expressed in 62 GC tissues from 74 cases (83.8%), which is significantly higher than non-tumoral adjacent parenchyma (20/46, 43.8%, P < 0.05). A high HO-1 expression rate showed a close association with well/moderate histological differentiation and negative lymph node metastasis (P < 0.05). The expression of matrix metallopeptidase 9 (MMP9) and vascular endothelial growth factor A (VEGF-A) as well as chemosensitivity to cisplatin of MKN-45 cell lines with genetically altered HO-1 status were then determined by realtime polymerase chain reaction and 3-(4,5 dimethyl thiazol-2-yl)-2,5-diphenyltetrazolium bromide (MTT), respectively. Whether the induction or inhibition of HO-1 by cobalt-protoporphyrin-IX (CoPP) or zinc-protoporphyrin-IX (ZnPP) could affect the sensitivity of MKN-45 cells to cisplatin was also studied. Results showed that the expression of MMP9 and VEGF-A were up-regulated in MKN-45 cells overexpressing HO-1, and down-regulated in HO-1 interfered cells. HO-1 overexpression could lead to an increased resistance to cisplatin, whereas down-regulation of HO-1 expression by siRNA or chemical inhibition of HO-1 could lead to increased chemosensitivity to cisplatin in MKN-45 cells. HO-1 may have multiple effects on protection against carcinogenesis and progression in GC.
Keywords
Introduction
Gastric cancer (GC) is one of the most common cancers and a major cause of cancer-related death worldwide, especially in East Asia, South America, Eastern Europe and the Middle East. 1 Clinical outcomes of GC still remain unsatisfactory despite recent progress in diagnosis and medical treatments. 2 Surgery and combination chemotherapies have been shown to confer only modest survival benefits in advanced GC, and result in a five-year survival rate of <7% in patients with metastases to distant organs. 3 Recent advances in molecular biology have resulted in the discovery of a wide range of alterations in gene expression during the course of gastric carcinogenesis. 4 However, the molecular mechanisms by which disease development, progression and resistance to chemotherapies occur in GC remain largely unknown.
Heme oxygenase (HO) is a rate-limiting enzyme of heme catabolism, which degrades heme to produce biliverdin, carbon monoxide and ferrous iron. 5 HO-1, also termed heat-shock protein 32 (Hsp32), is a stress responsive protein, which can be induced by a wide range of stimuli and activated signaling molecules, such as heat, ultraviolet irradiation, the HO-1 substrate heme, reactive oxygen species, hypoxia and nitric oxide signaling elements. 6 HO-1 mitigates cellular injury by exerting antiapoptotic, antioxidant and anti-inflammatory effects; these benefits of HO-1 have been widely demonstrated in a variety of pathological states including vascular injury, hypoxic lung disease and cardiac transplant rejection. 7–9 Recently, accumulating evidence has suggested that HO-1 activation may play a role in carcinogenesis and can potently influence the growth and metastasis of tumors. 10 Elevated HO-1 was observed in various tumor tissues, including lymphosarcoma, hepatoma, glioblastoma, melanoma, pancreatic cancer, brain tumors and esophageal cancer, among others. 11–17 Reports indicate that HO-1 seems to facilitate tumor growth and metastasis. However, the exact effects depend on the type of tumor. 10 Until now, few reports have focused on the relationship between HO-1 and GC. 18 Liu et al. 19 demonstrated that GC cells with elevated HO-1 induced by hemin showed higher resistance to apoptosis. A recent study indicated that the HO-1 gene promoter polymorphism was associated with a risk of gastric adenocarcinoma. 20 However, to date, no study has been performed on the HO-1 expression pattern and its clinicopathological significance in human GC cases. In addition, the biological behavior and function of HO-1 in GC have not been fully elucidated.
In the present study, we assessed the expression profile of HO-1 protein in human GC tissue by immunochemistry staining, including GC and non-tumoral adjacent parenchyma. The correlations between HO-1 expression and clinicopathological characteristics were investigated. In addition, we explored the function of HO-1 in GC cells by determining the functional consequences of genetically altered HO-1 status in MKN-45 cell lines, in which HO-1 was over-expressed or knocked down by transduction with lentiviral vectors.
Materials and methods
Patients and tissue specimens
Summary of patients characteristics involved in the study (n = 74)
TNM, tumor node metastasis
Immunohistochemical staining and evaluation
Samples were fixed in 4% formaldehyde in phosphate-buffered saline (PBS) at pH 7.2 and, after dehydration, embedded in paraffin wax and processed for immunohistochemical analysis of HO-1. Serial sections, 4 μm thick, were deparaffinized in xylene and re-hydrated through a graduated alcohol series to distilled water. Sections were then placed in citrate buffer (10% citrate buffer stock in distilled water, pH 6.0) and microwaved for 15 min for antigen retrieval. After blocking endogenous peroxidase for 10 min and protein for five minutes, diluted mouse anti-human HO-1 monoclonal antibody (1:800; Santa Cruz Biotechnology, Santa Cruz, CA, USA) was applied, respectively, for one hour. Serial sections were also stained by mouse anti-human CD8 monoclonal antibody (1:500; Abcam, Cambridge, MA, USA) for detecting the HO-1-positive T lymphocytes. Then the slides were incubated with biotin-labeled goat anti-mouse secondary antibody and detected with diaminobenzidine (Sigma, St Louis, MO, USA). Nuclei were counterstained with Mayer hematoxylin. Washing steps using PBS were conducted between each step, and all incubations were conducted at room temperature unless otherwise specified. Positive and negative immunohistochemical controls were routinely used. Blind examination was performed by two pathologists. The HO-1-positive grade was determined based on the proportion of stained cells on a scale of negative to strong: negative: <5% of stained cells with the grade of 0; weak: 5–25% of stained cells with grade 1; moderate: 25–50% of stained cells with grade 2; and strong: >50% of stained cells with grade 3.
Statistical analysis
All statistical analyses were carried out using SPSS software, version 17.0 (SPSS, Chicago, IL, USA). The differential frequency of HO-1 expression between GC and adjacent normal gastric tissue was analyzed by using Mann-Whitney U test. The association between HO-1 expression and clinicopathologic variables of GC was assessed by Mann-Whitney U test or Kruskal–Wallis H test. A two-sided P < 0.05 was considered significant.
Construction of lentiviral vectors
These vectors were based on the lentiviral vector LL3.7. The cassette of SV40 promotor and blasticidin selection gene from plenti6/V5-TOPO plasmid (Invitrogen, Carlsbad, CA, USA) was cloned into LL3.7. pCMV-HO-1, a plasmid overexpressing hHO-1, was constructed by inserting the entire protein-coding region (867 bp) of the human HO-1 cDNA behind the LL3.7-BSD-CMV promotor. The pLL3.7-BSD-CMV was used as a control. pS-shRNAHO-1, a plasmid carrying shRNA for HO-1, was engineered by annealing the single-strand oligonucleotides: 21 5′ TGGCAGAGGGTGATAGAAGATTCAAGAGATCTTCTATCACCC TCTGCCTTTTTTC, 3′ ACCGTCTCCCACTATCTTCTAAGTTCTCTAGAAGATAGTGGGAGACGGAAAAAAGAGCT, and inserting the double-stranded oligonucleotide behind the LL3.7-BSD-U6 promotor. Correspondingly, the non-silencing control duplex was inserted behind the LL3.7-BSD-U6 promotor as a control.
Cell culture and construction of transgenic cell lines
GC cell line MKN-45 was obtained from the American Type Culture Collection (ATCC, Manassas, VA, USA) and was maintained at 37°C in a humidified atmosphere of 5% CO2 and 95% air in RPMI 1640 containing 10% fetal bovine serum (Hyclone, Logan, UT, USA). Lentivirus production was prepared according to the protocol which Karolewski et al. described previously. 22 For each collection, viral supernatants were filtered through a 0.45-μm polyvinylidene fluoride (PVDF) syringe filter (Millipore, Billerica, MA, USA). The MKN-45 cells were incubated overnight with 1 mL diluted solution (100 μL viral supernatant and 900 μL RPMI 1640 containing 10% fetal bovine serum) containing 8 μg/mL of polybrene (Sigma) for a 24-well plate. The next day, the media containing virus were removed and replaced with 2 mL complete culture medium. A measure of 10 μg/mL blasticidin was used for stable cell line selection at 24 h post-transduction for 15 d. As controls, we also isolated MKN-45 cells transduced with pLL3.7-BSD-CMV vector (CON-OX) and CON-RNAi vector (CON-RNAi), respectively, using the protocol described above. Stable cell lines were tested by realtime quantitative polymerase chain reaction (PCR), Western blotting and HO enzyme activity measurement.
Realtime quantitative PCR
Total RNA was isolated from cultured cells using TRIzol reagent (Invitrogen) according to the manufacturer's instructions. Reverse transcription was carried out using RevertAid First Strand cDNA Synthesis Kit (Fermentas, Burlington, Canada). Realtime quantitative PCR was performed using SYBR Green Realtime PCR Master Mix (Toyobo, Osaka, Japan) and Realtime PCR Detection System Bio-Rad IQ5. The specific primers for hHO-1, matrix metallopeptidase 9 (MMP9), vascular endothelial growth factor A (VEGF-A) and glyceraldehyde-3-phosphate dehydrogenase (GAPDH) were designed as following: hHO-1, forward primer ATGACACCAAGGACCAGAGC, reverse primer GTGTAAGGACCCATCGGAGA; MMP9, forward primer GGGAAGATGCTGCTGTTCA, reverse primer GAACTCACGCGCCAGTAGA; VEGF-A, forward primer GGGCAGAATCATCACGAAGT, reverse primer TGGTGATGTTGGACTCCTCA; and GAPDH, forward primer CCCATCACCATCTTCCAGGAG, reverse primer GTTGTCATGGATGACCTTGGC. To confirm amplification specificity, PCR products were subjected to a melting curve analysis and gel electrophoresis. The experimental results were analyzed by the 2−ΔΔCt method.
Western blotting
Cells were harvested and lysed in appropriate buffer containing 1% Triton-100, 1 × PBS, protease inhibitors cocktail (Roche, Indianapolis, IN, USA). Equal amounts of proteins, which were determined using the bicinchoninic acid method, were separated on a 12% sodium dodecyl sulfate-polyacrylamide gel, transferred to a PVDF membrane and subjected to immunoblot using specific antibodies against HO-1 (1:800; Santa Cruz Biotechnology) and α-tubulin (1:1000; Santa Cruz Biotechnology). α-Tubulin was used to normalize the amounts of total protein present in the samples. Membranes were incubated with an appropriate peroxidase-conjugated secondary antibody (Santa Cruz Biotechnology), and protein bands were visualized by enhanced chemiluminescence (Millipore) and X-ray film imaging.
HO enzyme activity assay
Cells were washed twice with PBS, scraped with a rubber policeman and pelleted at 2500
3-(4,5 Dimethyl thiazol-2-yl)-2,5-diphenyltetrazolium bromide assay
In total, 100 μL cells were seeded in 96-well culture plates (5000 cells/well). After an overnight preincubation, cells were exposed for 24 hours to indicated concentrations of cisplatin, in the absence or presence of zinc-protoporphyrin-IX (ZnPP) or cobalt-protoporphyrin-IX (CoPP) with the concentration of 10μmol/L, respectively. 3-(4,5 Dimethyl thiazol-2-yl)-2,5-diphenyltetrazolium bromide (MTT) solution was added into each well. After four hours, the supernatant was carefully removed. One hundred and fifty microliters of dimethyl sulfoxide was used to dissolve the crystals by agitation for 10 min. The absorbance at 490 nm (A490) of each well was read on a microplate luminometer (Applied Biosystems, Foster City, CA, USA). Each experiment was done in triplicate. The toxicity of cisplatin was quantified as the fraction of cells surviving relative to untreated controls. The data were expressed as mean ± SD, and Student's t-test was used to determine the significance of differences in multiple comparisons. P < 0.05 was considered to be statistically significant.
Results
HO-1 protein expression pattern in gastric cancer
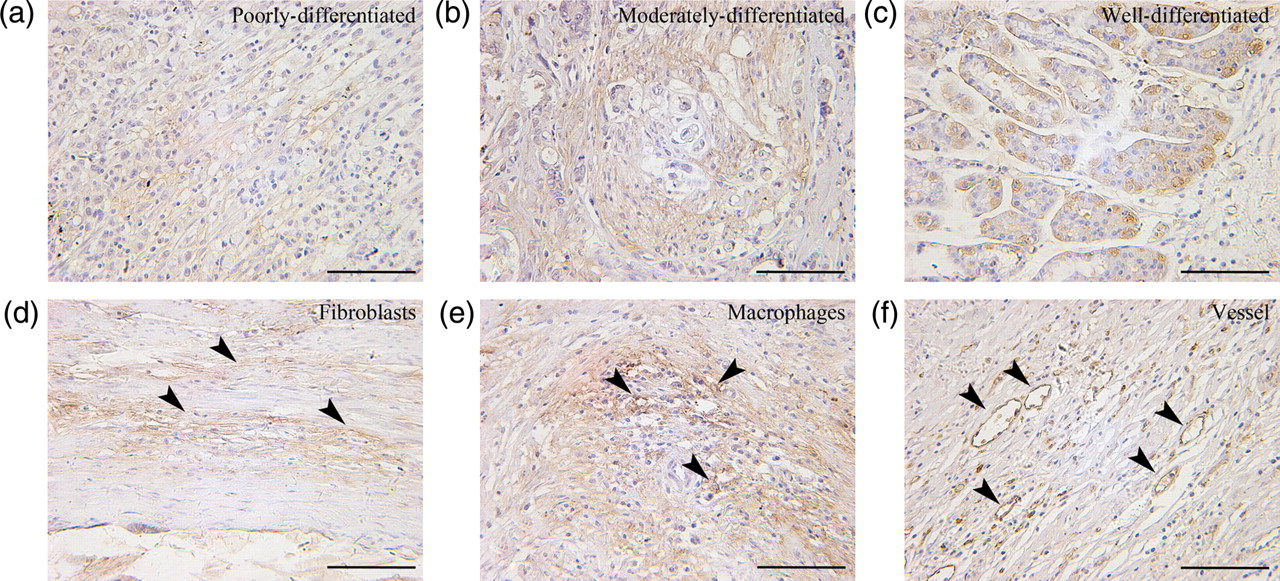
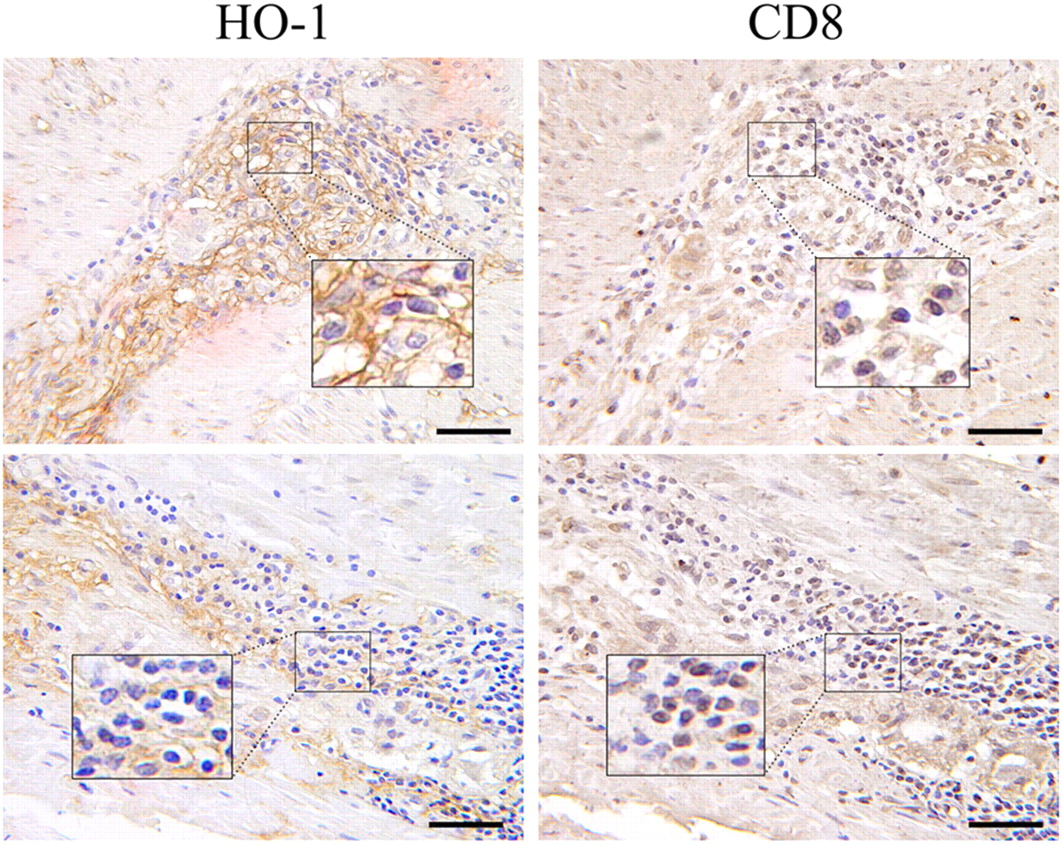
Immunohistochemical analysis of surgical resections of GC tissue and non-tumoral adjacent parenchyma showed that HO-1 protein was expressed in tumor tissues in 62 of 74 (83.8%) cases, and with a relatively lower expression in adjacent normal gastric tissues (20/46, 43.8%) taken near the excision site from the same tumor patients (P < 0.01; Figure 1). In the GC tissues, some cases showed negative or weak staining (Figures 1c and d), whereas other cases revealed moderate to strong staining (Figures 1e and f). In addition, HO-1 protein was expressed diffusely in well/moderately differentiated adenocarcinoma and poorly differentiated adenocarcinoma (Figures 2a–c). Stromal cells including fibroblasts and macrophages were also positive for HO-1 (Figures 2d and e). Moreover, the intratumoral vessels within the cancer nests exhibited positive staining for HO-1 (Figure 2f). To further evaluate whether the HO-1-positive cells contained T lymphocytes, serial sections were stained for HO-1 and CD8, a maker of cytotoxic T lymphocytes. It was shown that a number of HO-1-positive cells were also CD8-positive in the sections of human GC (Figure 3).
Immunohistochemical staining for heme oxygenase-1 (HO-1) in representative specimens of gastric cancer (GC) tissue and non-tumoral adjacent parenchyma. (a) An example case of negative expression in non-tumoral adjacent parenchyma. (b) An example case of positive expression in adjacent normal gastric tissue. HO-1 was stained in glandular epithelial cells but not in foveolar epithelial cells. (c) An example case of negative expression in GC tissue. (d–f) Weak, moderate and strong expression in GC. Arrowheads indicate the representative positive sites. Scale bar = 50 μm. (A color version of this figure is available in the online journal) Heme oxygenase-1 (HO-1) localization in gastric cancer. (a) HO-1 was stained in poorly differentiated adenocarcinoma cells. (b) HO-1 was stained in moderately differentiated adenocarcinoma cells. (c) HO-1 was stained in well-differentiated adenocarcinoma cells. (d) HO-1 was stained in fibroblast cells. (e) HO-1 was stained in macrophages. (f) HO-1 was stained in the intratumoral vessels within the cancer nests. Arrowheads indicate the representative positive sites. Scale bar = 50 μm. (A color version of this figure is available in the online journal) Representative immunohistochemistry of serial sections of gastric cancer (GC) by using anti-human heme oxygenase-1 (HO-1) antibody and anti-human CD8 antibody. It was shown that a number of HO-1-positive cells were also CD8-positive in the sections of human gastric cancer. Inset shows the corresponding boxed area magnified threefold. Scale bar = 30 μm. (A color version of this figure is available in the online journal)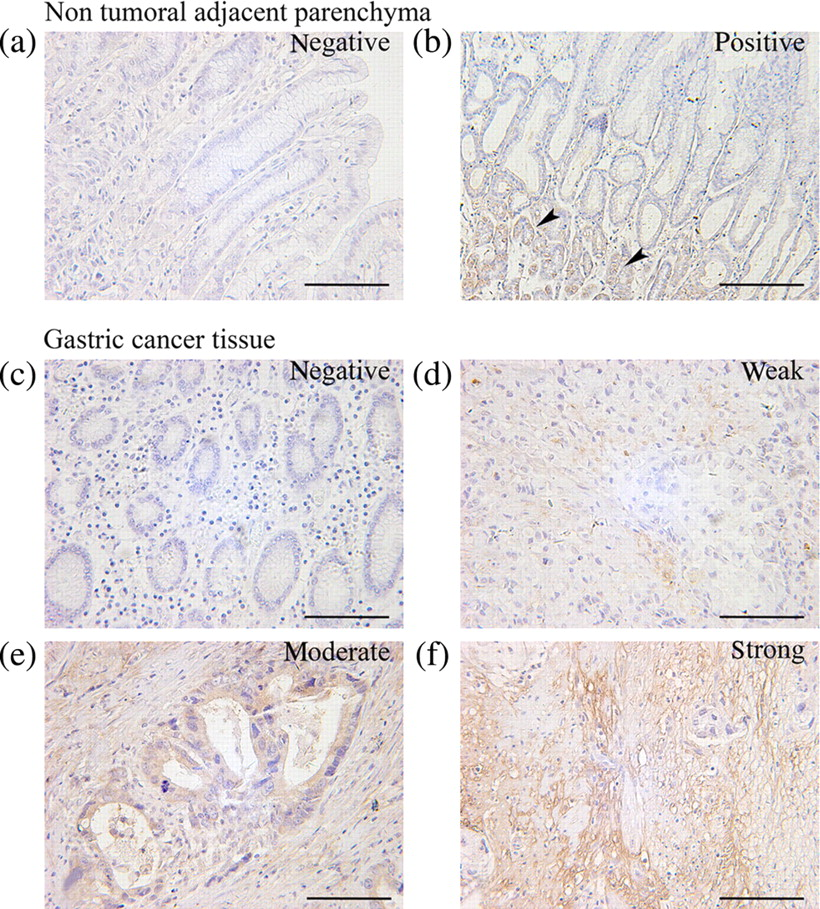
Relationship between HO-1 expression in GC and clinicopathological factors
HO-1 expression in gastric cancer and its correlation with clinicopathological characteristics
TNM, tumor node metastasis; HO-1, heme oxygenase-1
Characterization of transgenic MKN-45 cell lines
To elucidate the functional consequences of altered HO-1 status in human GC cells, we established a HO-1 over-expressed GC cell line (MKN-45/HO-1) in which HO-1 was over-expressed by transduction with lentiviral vectors and an HO-1-silenced GC cell line (MKN-45/RNAi-HO-1) in which endogenous HO-1 expression was knocked down by transduction with RNAi lentiviral vectors. Realtime quantitative PCR results showed that the mRNA level of HO-1 was dramatically increased by ∼6-fold in the MKN-45/HO-1 cell line compared with in the MKN-45/CON-OX cell line. In the case of the MKN-45/RNAi-HO-1 cell line, the mRNA level was significantly decreased by ∼80% compared with the MKN-45/CON-RNAi cell line (Figure 4a). Western blotting confirmed that the MKN-45/HO-1 cell line had significantly increased HO-1 protein levels, and that the MKN-45/RNAi-HO-1 cell line had significantly decreased HO-1 protein levels compared with controls, respectively (Figure 4b). Biochemical quantification of HO enzymatic activity further confirmed that the MKN-45/HO-1 cell line had significantly increased HO activity, whereas the MKN-45/RNAi-HO-1 cell line had significantly decreased HO activity compared with controls, respectively (P < 0.05) (Figure 4c). We then analyzed the MKN-45 cell lines with modified HO-1 status for their MMP9 and VEGF-A mRNA expression levels and resistance to chemotherapeutic treatment.
Characterization of transgenic cell lines. (a) Realtime quantitative polymerase chain reaction results showed that the mRNA level of heme oxygenase-1 (HO-1) was dramatically increased by ∼6-fold in the MKN-45/HO-1 cell line compared with in the MKN-45/CON-OX cell line. In the case of the MKN-45/RNAi-HO-1 cell line, the mRNA level was significantly decreased by ∼80% compared with the MKN-45/CON-RNAi cell line. (b) Western blotting confirmed that the MKN-45/HO-1 cell line had significantly increased HO-1 protein levels, and that the MKN-45/RNAi-HO-1 cell line had significantly decreased HO-1 protein levels compared with controls, respectively. (c) Biochemical quantification of HO enzymatic activity further confirmed that the MKN-45/HO-1 cell line had significantly increased HO activity, whereas the MKN-45/RNAi-HO-1 cell line had significantly decreased HO activity compared with controls, respectively. Values are the average of three measurements with the SD indicated by error bars. *P < 0.05. (A color version of this figure is available in the online journal)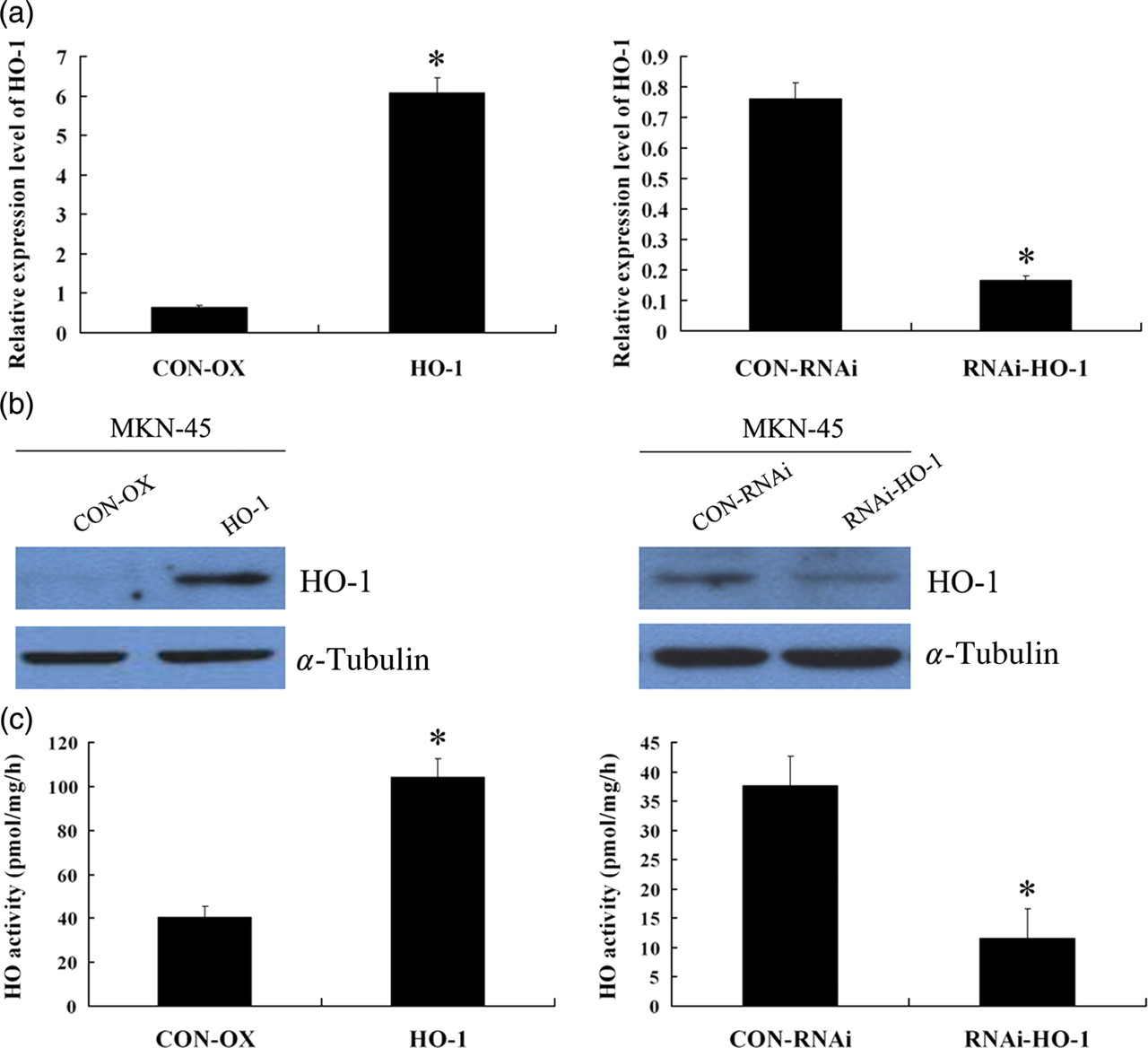
MMP9 and VEGF-A mRNA expression levels in transgenic MKN-45 cell lines
We analyzed MMP9 mRNA expression level in the human MKN-45 cell lines with modified HO-1 status by realtime quantitative PCR. Results demonstrated that the mRNA level of MMP9 was dramatically increased by ∼3-fold in the MKN-45/HO-1 cell line compared with in the MKN-45/CON-OX cell line. In the case of the MKN-45/RNAi-HO-1 cell line, the mRNA level was significantly decreased by ∼80% compared with the MKN-45/CON-RNAi cell line (Figure 5a).
Matrix metallopeptidase 9 (MMP9) and vascular endothelial growth factor A (VEGF-A) mRNA expression levels in transgenic MKN-45 cell lines. (a) mRNA level of MMP9 was dramatically increased by ∼3-fold in the MKN-45/HO-1 cell line compared with in the control MKN-45/CON-OX cell line. In the case of the MKN-45/RNAi-HO-1 cell line, the mRNA level was significantly decreased by ∼80% compared with the control MKN-45/CON-RNAi cell line. (b) mRNA level of VEGF-A was increased by ∼1.5-fold in the MKN-45/HO-1 cell line compared with in the control cell line, whereas the mRNA level was significantly decreased by ∼80% in the MKN-45/RNAi-HO-1 cell line, compared with control. Values are the average of three measurements with the SD indicated by error bars. *P < 0.05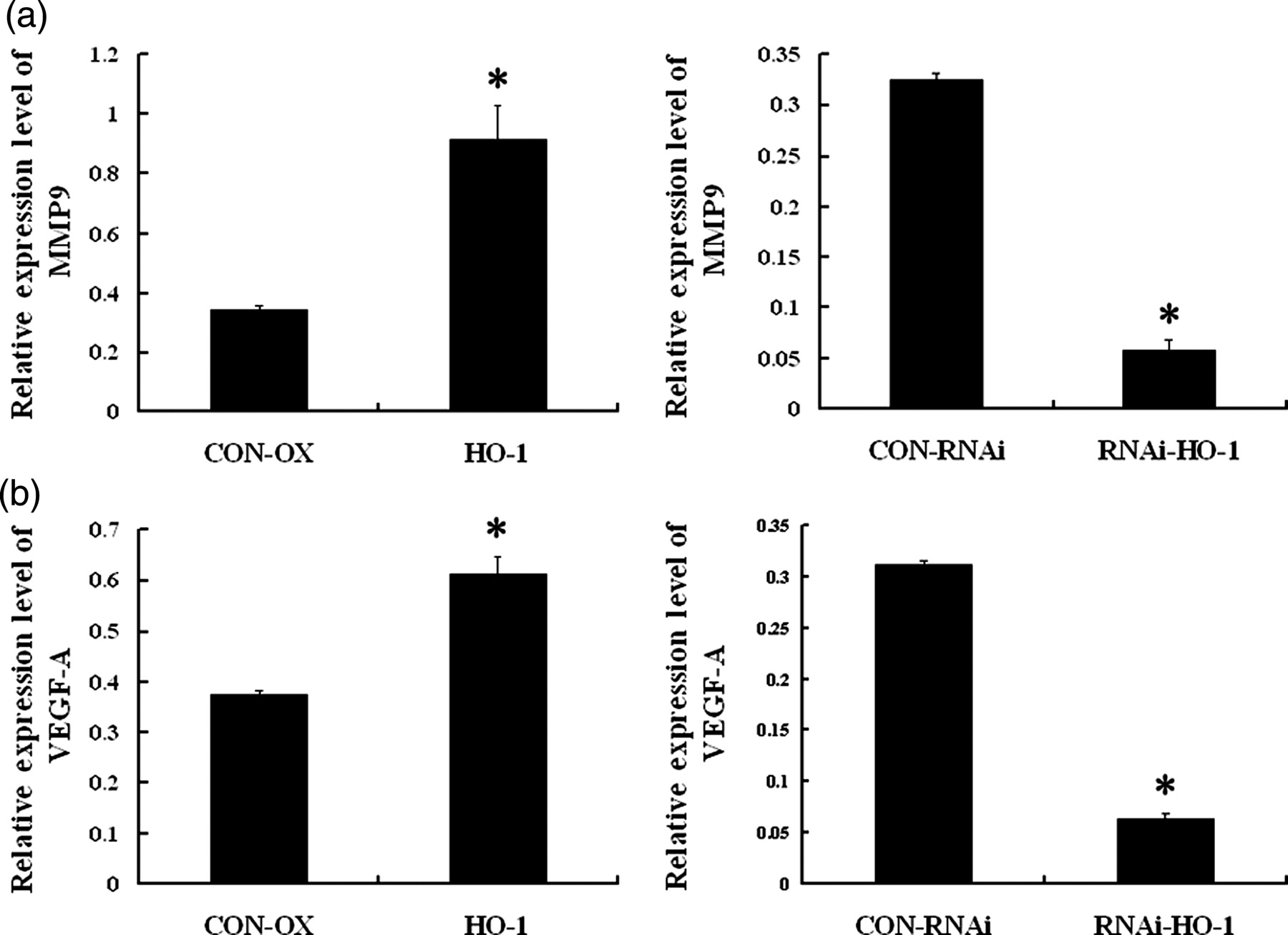
In order to understand the relationship between the HO-1 and VEGF-A expression in human GC cells, we then analyzed VEGF-A mRNA expression level in these transgenic cell lines by realtime quantitative PCR. Results demonstrated that the mRNA level of VEGF-A was increased by ∼1.5-fold in the MKN-45/HO-1 cell line compared with in the control cell line, whereas the mRNA level was significantly decreased by ∼80% in the MKN-45/RNAi-HO-1 cell line, compared with the MKN-45/CON-RNAi cell line (Figure 5b). Theses results indicated that increased HO-1 expression was associated with up-regulated expression of MMP9 and VEGF-A, and decreased HO-1 expression was in accordance with down-regulated expression of MMP9 and VEGF-A in human GC cells.
Effect of HO-1 on the cisplatin sensitivity of MKN-45 cells
To further evaluate the functional aspects regarding HO-1 expression in GC cells, we next investigated whether the over-expression of HO-1 or the inhibition of HO-1 by genetic manipulation affected the sensitivity of MKN-45 cells to the antitumor drug cisplatin. The survival rate in MKN-45/HO-1 culture was markedly enhanced compared with the respective MKN-45/CON-OX control culture for each cisplatin concentration (P < 0.05) (Figure 6a), whereas the survival rate in MKN-45/RNAi-HO-1 culture was markedly decreased compared with the respective MKN-45/CON-RNAi control culture for each cisplatin concentration (P < 0.05) (Figure 6b). Since CoPP is a known inducer, while ZnPP is an inhibitor, of HO-1, we then studied whether the over-expression or the inhibition of HO-1 by CoPP or ZnPP could affect the sensitivity of MKN-45 cells to cisplatin, too. The results showed that the survival rate in CoPP-treated MKN-45 culture was significantly enhanced compared with the respective MKN-45 control culture for each cisplatin concentration, whereas the survival rate in ZnPP-treated MKN-45 culture was markedly decreased compared with the respective MKN-45 control culture for each cisplatin concentration (P < 0.05) (Figure 6c). These results indicated that increased HO-1 expression could lead to a resistance to chemotherapy in MKN-45 cells. Targeted knockdown gene expression of HO-1 by siRNA or chemical inhibition of HO-1 enzymatic activity by ZnPP increased the chemosensitivity to cisplatin in MKN-45 cells.
Effect of heme oxygenase-1 (HO-1) on the cisplatin sensitivity of MKN-45 cells. (a) MKN-45/CON-OX and MKN-45/HO-1 cell lines were treated with various concentrations of cisplatin. (b) MKN-45/CON-RNAi and MKN-45/RNAi-HO-1 cell lines were treated with various concentrations of cisplatin. Cell viability was determined by MTT (3-(4,5 dimethyl thiazol-2-yl)-2,5-diphenyltetrazolium bromide) assay. *P < 0.05 compared with the respective control culture for each cisplatin concentration. (c) MKN-45 cell lines were treated with cobalt-protoporphyrin-IX (CoPP) or zinc-protoporphyrin-IX (ZnPP) for six hours before they were treated with cisplatin. CoPP-induced HO-1 inhibits the cisplatin sensitivity of MKN-45 cells. HO-1 enzyme activity inhibition with ZnPP led to a significant increase of cisplatin sensitivity in MKN-45 cells. *P < 0.05 compared with the respective control culture for each cisplatin concentration. Values are the average of three measurements with the SD indicated by error bars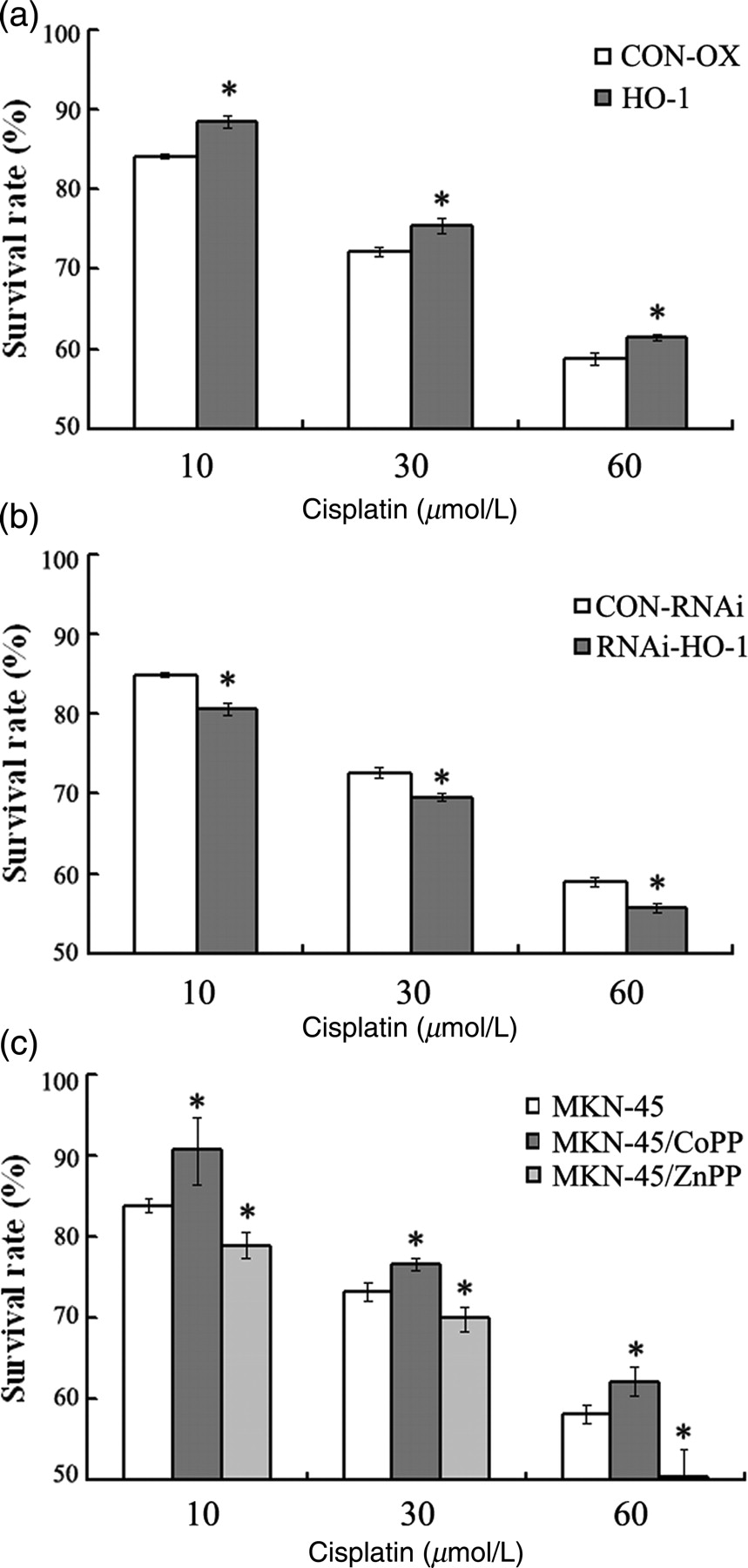
Discussion
HO-1 is involved in many physiological and pathophysiological processes. A great number of researches have investigated the roles of HO-1 in the formation, growth and metastasis of tumors. 10,18 The role of HO-1 in GC is only just beginning to be studied in detail, and few papers on HO-1 have touched upon this subject.
This is the first clinical report concerning HO-1 expression pattern and its clinicopathological significance in human GC cases. Expression of HO-1 is usually up-regulated in tumors compared with adjacent non-tumoral tissues, which was shown in lymphosarcoma, hepatoma, pancreatic cancer and brain tumors, etc. 11–16 In gastrointestinal tumors, Becker et al. 24 showed that focal HO-1 expression in colorectal cancer was higher that in colonic adenoma. In the present study, we assessed the expression profile of HO-1 protein in human GC tissue by immunochemistry staining. Results showed that the HO-1 protein was expressed in tumor tissues by 62 of 74 (83.8%) human GC cases, with significantly lower expressed in adjacent non-tumoral gastric tissues (20/46, 43.8%, P<0.05). This result indicated that the percentage of cells that express HO-1 is higher in tumor tissues than that in adjacent non-tumoral gastric tissues of human GC. This result was in agreement with the findings obtained from the research mentioned above to some extent.
The exact cell types that express HO-1 are not very clear and may depend on the type of tumor. In human prostate, esophageal, tongue and renal cancer, only the tumor cells revealed strong HO-1 signals. 25–28 In rat hepatoma, HO-1 was found only in tumor cells. 12 In human melanomas or gliomas, HO-1 was specifically expressed in macrophages, but only slightly in tumor cells, 13 whereas in human pancreatic cancer, cancer tissues revealed marked HO-1 immunoreactivity in tumor cells and in tumor-associated immunocytes. 15 In rat experimental esophageal cancer, the main target for HO-1 induction was the inflammatory cells inside the tumor or in subepithelial areas. 17 In human colorectal cancer, HO-1 expression was detected mainly in cancer cells and occasionally in macrophages close to the cancerous front, but was hardly observed in immunocytes. 24 In our study, HO-1 expression in GC specimens was detected not only in tumor cells but also in tumor-associated macrophages and CD8+ T lymphocytes (Figure 3). Moreover, the intratumoral vessels and the fibroblasts within the cancer nests exhibited positive staining for HO-1 (Figure 2). The results suggesting that the effects of HO-1 in human GC may be complex.
Furthermore, in this study, clinicopathological correlation analysis indicated that the higher HO-1 expression rate in human GC was significantly associated with a more favorable histological differentiation and a lower frequency of lymph node metastasis (Table 2). These findings seem to be in contrast to the findings that HO-1 inhibition could suppress tumor growth, induce apoptosis and inhibit the occurrence of metastasis in solid tumor in human and experimental animal models. 10 However, the results were in keeping with the findings in human tongue squamous cell carcinoma and human colorectal cancer. 24,28 In human colorectal cancer, HO-1 expression was associated with a significantly lower rate of lymphatic invasion and a significantly better long-term survival. 24 Furthermore, Yanagawa et al. 28 showed that low levels of HO-1 expression were associated with lymph node metastasis and unfavorable histological differentiation in human tongue cancer.
Information about the role of HO-1 expression on tumors is still controversial. Many studies convincingly indicated that HO-1 was a protumoral molecule because of its antiapoptotic and proangiogenic effects. Melanoma cells overexpressing HO-1 formed many more nodules in the lungs after the intravenous injection than did their wild-type counterparts. 29 Targeted knockdown of HO-1 expression led to pronounced growth inhibition of the pancreatic cancer cells and made tumor cells significantly more sensitive to radiotherapy and chemotherapy. 15 In the lung carcinoma, VEGF synthesis decreased after treatment with HO-1 inhibitor. 30 In addition, HO-1 inhibition and silencing significantly suppressed MMPs and invasion of lung cancer cells. 31 However, HO-1 has also been shown to inhibit the proliferation and induce apoptosis in breast carcinoma cells. 32 Furthermore, HO-1 inhibited breast cancer invasion via suppressing the expression of MMP9. 33 In prostate cancer cells, MMP9 production and activity was down regulated by HO-1 overexpression. 34 Most recently, Ferrando et al. 35 convincingly demonstrated that the pro-angiogenic genes were down-regulated in response to HO-1 overexpression in prostate cancer cell lines and HO-1 overexpression in prostate cancer had a strong in vivo antiangiogenic effect challenging tumor development. These conflicting results imply that HO-1 may have both pro and antagonist effects on tumor progression and its role in cancer is far from being completely understood.
In order to understand the effect of HO-1 on the metastasis in human GC, we then analyzed MMP9 and VEGF-A mRNA expression levels in MKN-45 cell lines with altered HO-1 expression levels by realtime quantitative PCR. Unexpectedly, the results showed that decreased HO-1 expression was in accordance with down-regulated expression of MMP9 and VEGF-A, and HO-1 over-expression was associated with up-regulated expression of MMP9 and VEGF-A, indicating an increased ability of metastasis. This finding was not in accordance with our previous immunochemistry study that the higher HO-1 expression was related to a lower frequency of lymph node metastasis. Although unexpected, we believe our results to be reasonable. One possible reason is that, as HO-1 is a sensitive marker of cellular stress, its expression in GC tissue might represent some recognition by the host. If the human immune system recognizes the tumor cell as a non-self-antigen, tumor cells might be attacked by the defense mechanisms of the host, making metastasis to the peripheral regions difficult. 28 In addition, besides the cancer cells, HO-1 was also expressed in other cell types, including macrophages, T lymphocytes, vessel cells and fibroblasts, as shown in our immunochemistry study. It was suggested that the anti-inflammatory effects of HO-1 might be related to the pathogenesis of GCs. 36 It was also possible that the vascular status modulated by HO-1 plays a role in the lymph node metastasis of GC. 20 Moreover, most of the data regarding the role of HO-1 in cancer are derived from studies in vitro or from animal models. Cells grown on tissue culture substrates or animal models can differ considerably in their morphology, cell–cell and cell–matrix interactions, and differentiation from those growing in human physiological environments. It should be noticed that the incongruence between the in vivo and in vitro studies also existed in other research of the relationship between HO-1 and tumors. In colorectal cancer, HO-1 expression was associated with a significantly lower rate of lymphatic invasion, a tendency towards fewer lymph node metastases and a significantly better long-term survival, 24 whereas in human colon cancer Caco-2 cells, the induction of HO-1 inhibited apoptosis via activation of the Akt pathway. 37 So, whether HO-1 may have a multiple effects on protection against carcinogenesis and progression in GC needs further study. Additional studies from bench to bedside are needed to further elucidate the role of HO-1 in GC, which seems to be more complex than expected. Anyway, although HO-1 may act differently in human GC tissue in vivo and GC cells in vitro, the in vitro analysis of MMP9 and VEGF-A in GC cells indicated that a therapy targeting the HO-1 expression in GC cells could be effective for GC.
Defects in apoptosis are important in tumorigenesis and in the treatment of cancer. A number of chemotherapeutic agents have been shown to induce apoptosis. However, most GCs are not well responsive to chemotherapy, suggesting that these tumors are intrinsically resistant to apoptosis-inducing agents. 38 We therefore examined the effect of altered HO-1 expression to see whether it could enhance or reduce the sensitivity of MKN45 towards chemotherapeutic agent cisplatin. Results showed that increased HO-1 expression could lead to a resistance, and inhibition of HO-1 expression could increase the chemosensitivity to cisplatin in MKN-45 cells, suggesting HO-1 is an adjuvant chemotherapy target for GC. This finding was in agreement with many reports that HO-1 was a cytoprotective and antiapoptotic enzyme in tumor cells exposed to diverse stimuli. 10 It has been demonstrated that GC cells with elevated HO-1 induced by hemin showed higher resistance to apoptosis. 19 Our results demonstrate for the first time that, in GC, targeted knockdown gene expression of HO-1 by siRNA or chemical inhibition of HO-1 enzymatic activity by ZnPP could both enhance cisplatin-induced apoptosis.
In conclusion, this is the first report concerning HO-1 expression pattern and its clinicopathological significance in human GC cases. The occasion of HO-1 protein expressed in tumor tissues was significantly higher than that in adjacent non-tumoral gastric tissues. HO-1 expression was significantly associated with the histological differentiation and lymph node metastasis. A higher HO-1 expression rate might possess a protective effect against GC with a more favorable histological differentiation and a lower frequency of lymph node metastasis. However, HO-1 acts differently in human MKN-45 GC cells, in which increased HO-1 expression was associated with an increased ability of metastasis. Finally, down-regulation of HO-1 expression by siRNA or chemical inhibition of HO-1 enzymatic activity could lead to increased chemosensitivity to cisplatin.
Footnotes
ACKNOWLEDGEMENTS
This work was supported by the National Basic Research Program of China (No. 2009CB521702).