
Abstract
The molecular mechanism of the interaction between pepsin and two typical ionic liquids (ILs), 1-butyl-3-methylimidazolium chloride ([C4mim]Cl) and 1-octyl-3-methylimidazolium chloride ([C8mim]Cl), was investigated with fluorescence spectroscopy, ultraviolet absorption, and circular dichroism spectroscopy at a pH value of 1.6. The results suggest that ILs could quench the intrinsic fluorescence of pepsin, probably via a dynamic quenching mechanism. The fluorescence quenching constants were determined by employing the classic Stern-Volmer equation. The constant values are very small, indicating that only a very weak interaction between ILs and pepsin exists. The Gibbs free-energy change, enthalpy change (ΔH), and entropy change (ΔS) during the interaction of pepsin and ILs were estimated. Positive values of ΔH and ΔS indicate that the interaction between ILs and pepsin is mainly driven by hydrophobic interaction. Synchronous and three-dimensional fluorescence spectra demonstrate that the addition of ILs (0–0.20 mol L−1 for each IL) does not bring apparent changes to the microenvironments of tyrosine and tryptophan residues. Activity experiments show that the activity of pepsin is concentration dependent; higher concentrations of ILs (>0.22 mol L−1 for [C8mim]Cl and >0.30 mol L−1 for [C4mim]Cl) cause the remarkable reduction of enzyme activity. The presence of ILs also does not improve the thermal stability of pepsin.
Keywords
INTRODUCTION
Ionic liquids (ILs) have attracted extensive attention mainly due to their negligible vapor pressure and low degree of flammability. Environmental and safety problems arising through the use of volatile organic solvents can be avoided by the use of these innovative materials. Therefore, ILs have been widely used as novel media for applications in organic synthesis, 1 electrochemistry, 2 and separation science. 3
Although the non-volatility of ILs likely means little atmospheric pollution if released, either intentionally after industrial waste processing or accidentally during a spill, they would accumulate in the environment because of their high solubility in water and high stability, thereby increasing the potential for contamination of aquatic ecosystems. 4 It is therefore important to obtain enough information on the environmental impact of ILs prior to intensive use in industrial settings. Recently, efforts have been made to understand the biological effects of each family of ILs, and a growing body of evidence suggests that they can be toxic to aquatic organisms.5–7
Molecular mechanisms of the interaction between ILs and biomacromolecules are fundamental to our understanding of the harmful effects of this kind of chemical on the environment and their more direct effects on human health. 8 Therefore, the effects of ILs on proteins and DNA have been reported during the past few years,9–19 mainly by investigation of their catalytic activity and stability in IL-containing systems. Baker's group 9 recently reported the extreme thermodynamic stabilization of monellin, offered by the IL 1-butyl-1-methylpyrrolidinium bis(trifluoromethane sulfonyl)imide ([C4mpy][Tf2N]); the unfolding temperature of monellin is about 105 °C in [C4mpy] [Tf2N] compared with 40 °C in bulk water. However, IL 1-butyl-3-methylimidazolium chloride ([C4mim]Cl) is a denaturant for green fluorescent protein, 10 human serum albumin, 11 and cytochrome c. 12 Akdogan's group 13 studied the effects of imidazolium-based ILs on the functional solution structure of human serum albumin (HSA) by using continuous-wave electron paramagnetic resonance spectroscopy and electron-electron resonance spectroscopy. Because competitive binding of imidazolium cations in HSA decreases the number of bound fatty acids (FAs), conformational changes of HSA after addition of ILs could be obtained by measurements of the spatial distribution of the binding sites of spin-labeled FAs to HSA. It is found that addition of imidazolium-based ILs to an aqueous solution of HAS-FA conjugates is accompanied by significant destabilization and unfolding of the protein's tertiary structure; however, the thermodynamics, which can provide information on the nature of the binding of ILs to HSA, are not mentioned in their work.
In this work, the interaction between imidazolium-based ILs and pepsin was investigated by spectroscopy including fluorescence spectroscopy, UV absorption, and circular dichroism (CD) spectroscopy. Great attempts were made to explore the interaction mechanism at a molecular level and the effect of imidazolium-based ILs on the enzymatic activity of pepsin. Fluorescence techniques are good aids in this study because of their high sensitivity, rapidity, and ease of implementation. From fluorescence quenching measurements, valuable information such as quenching mechanism, binding constants, and the thermodynamic parameters of the interaction process can be obtained. Two widely used ILs, 1-octyl-3-methylimidazolium chloride ([C8mim]Cl) and [C4mim]Cl, were selected, because this choice enabled us to investigate how the nature of the alkyl chain length of the ILs affects the interaction. Pepsin was chosen for study because of its well-characterized molecular structure and wide applications.
EXPERIMENTAL
Since pepsin is most active in acidic environments (pH 1.5–2.0), 21 all the pH values of aqueous media were fixed at 1.6. Therefore, stock solutions of pepsin (1.0 g L−1), NaCl (0.5 mol L−1), [Ctmim]Cl (2.0 mol L−1), [C8mim]Cl (2.0 mol L−1), and casein (0.5 g L−1) were prepared by dissolving a desired amount of that substance in HCl solution (pH 1.6). All stock solutions were stored in the dark at 0–4 °C.
Three-dimensional fluorescence spectra were obtained under the following conditions: The emission wavelengths were recorded from 240 to 500 nm; the excitation wavelengths ranged from 220 nm to 310 nm, with an increment of 5 nm, and the number of scanning curves was 19. The slit widths for both excitation and emission were also set at 5 nm. The concentration of pepsin was 0.4 g L−1, and the concentrations of [C4mim]Cl and [C8mim]Cl were both 5.0 × 10−3 mol L−1.
For the fluorescence lifetime measurements, samples were excited at 280 nm; the emission wavelength was set at 342 nm. The slit widths were both 4 nm for excitation and emission. Concentration of pepsin was 0.4 g L−1, and the concentrations of [C4mim]Cl and [C8mim]Cl were both 0.04 mol L−1.
The blank solution was prepared in the same manner: 0.2 mL of pepsin (0.5 g L−1, pH 1.6) and 3.0 mL of trichloroacetic acid (0.8 mol L−1) were added to a glass-stoppered tube and incubated at 40 °C for 10 min. Then, 12.0 mL of casein (0.5 g L−1, pH 1.6) was added to the mixture, and the final volume of the mixture was adjusted with water to 20 mL. After centrifuging, the UV absorbance of the supernatant was also measured at 275 nm.
To study the IL effects, a specified amount of each IL was added after the addition of enzyme solution to reach a final concentration of 0–1.0 mol L−1. The results are reported as the ratio between the activity of the enzyme with and without the presence of each IL (A:A0).
RESULTS AND DISCUSSION
Fluorescence Spectra Analysis. Fluorescence quenching processes in solution fall into two general types,25,26 (i) static quenching through the formation of a ground state complex and (ii) dynamic quenching (collisional quenching) due to diffusive collisions between the fluorophore and the quencher. They can be differentiated by their dissimilar dependence on temperature and viscosity, or preferably, by lifetime measurements. 27 Dynamic quenching is diffusion controlled, because the quencher must diffuse to the fluorophore during the lifetime of the excited state. Since high temperature results in faster diffusion, a larger extent of collisional quenching is expected with increasing temperature, but a reversed effect is observed for the static quenching, because high temperatures will also result in the dissociation of weakly bound complexes.
Fluorescence emission spectra of pepsin with the addition of different levels of [C4mim]Cl and [C8mim]Cl are shown in Fig. 1. Pepsin shows a strong fluorescence emission, with a peak at 342 nm at Λex = 280 nm, while ILs were almost nonfluorescent under the present experiment conditions. Obviously, the fluorescence of pepsin is effectively quenched by the addition of ILs, which indicates that interaction between ILs and pepsin occurred. However, the shape of the fluorescence spectra and peak position do not alter, suggesting that the interaction of pepsin with ILs is weak and there might be no complexes formation between ILs and pepsin.27–29 However, additional information is required to distinguish between dynamic or static processes, for example, the temperature dependence of the quenching rate constants, UV absorption spectra, and fluorescence lifetime measurements.
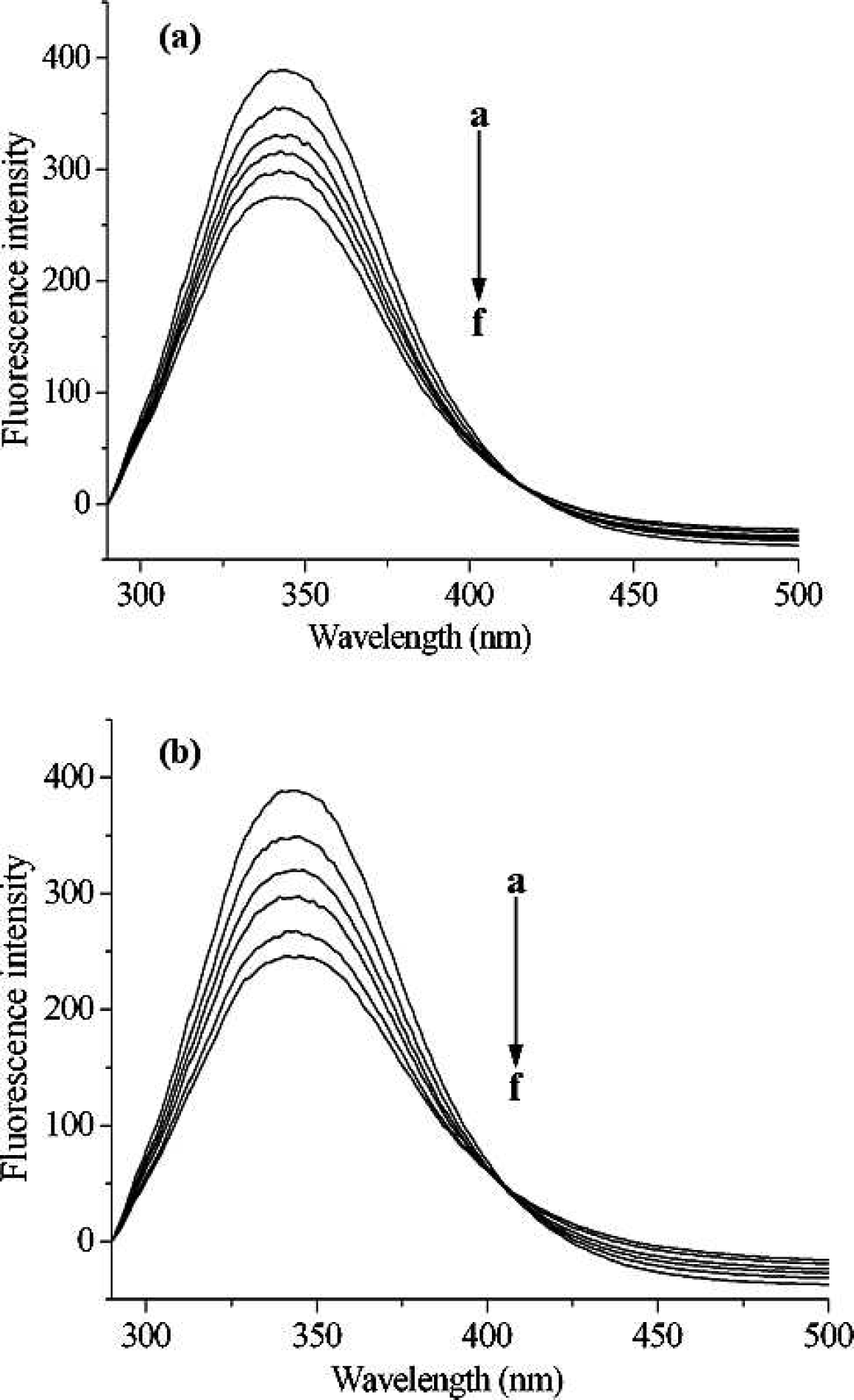
Fluorescence emission spectra of pepsin in the presence of various concentrations of [C4mim]Cl (
In order to judge the possible fluorescence quenching mechanism, the process of fluorescence quenching was first assumed to be dynamic. For dynamic quenching, the mechanism can be described by the Stern-Volmer equation: 28
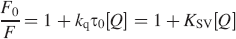
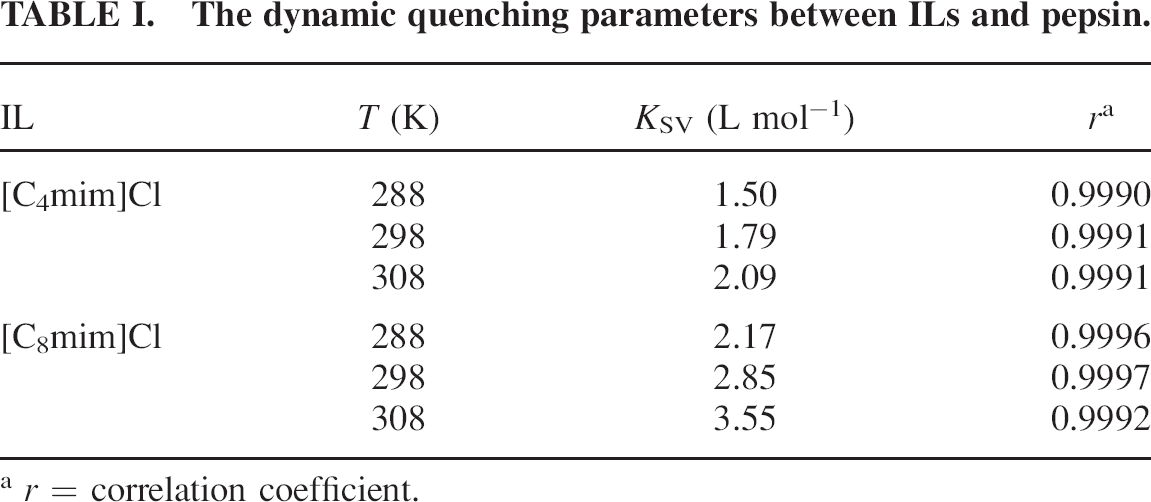
where F0 and F are the fluorescence intensities in the absence and presence of quencher, respectively; kq, KSV, τ0, and [Q] are the bimolecular quenching rate constant, the Stern-Volmer quenching constant, the average lifetime of molecule without quencher, and concentration of quencher, respectively. The Stern-Volmer plots of quenching of pepsin by different ILs and at different temperatures are displayed in Fig. 2. Based on the experimental data in Fig. 2, the dynamic quenching constants at different temperatures are shown in Table I.
The dynamic quenching parameters between ILs and pepsin.
r = correlation coefficient.
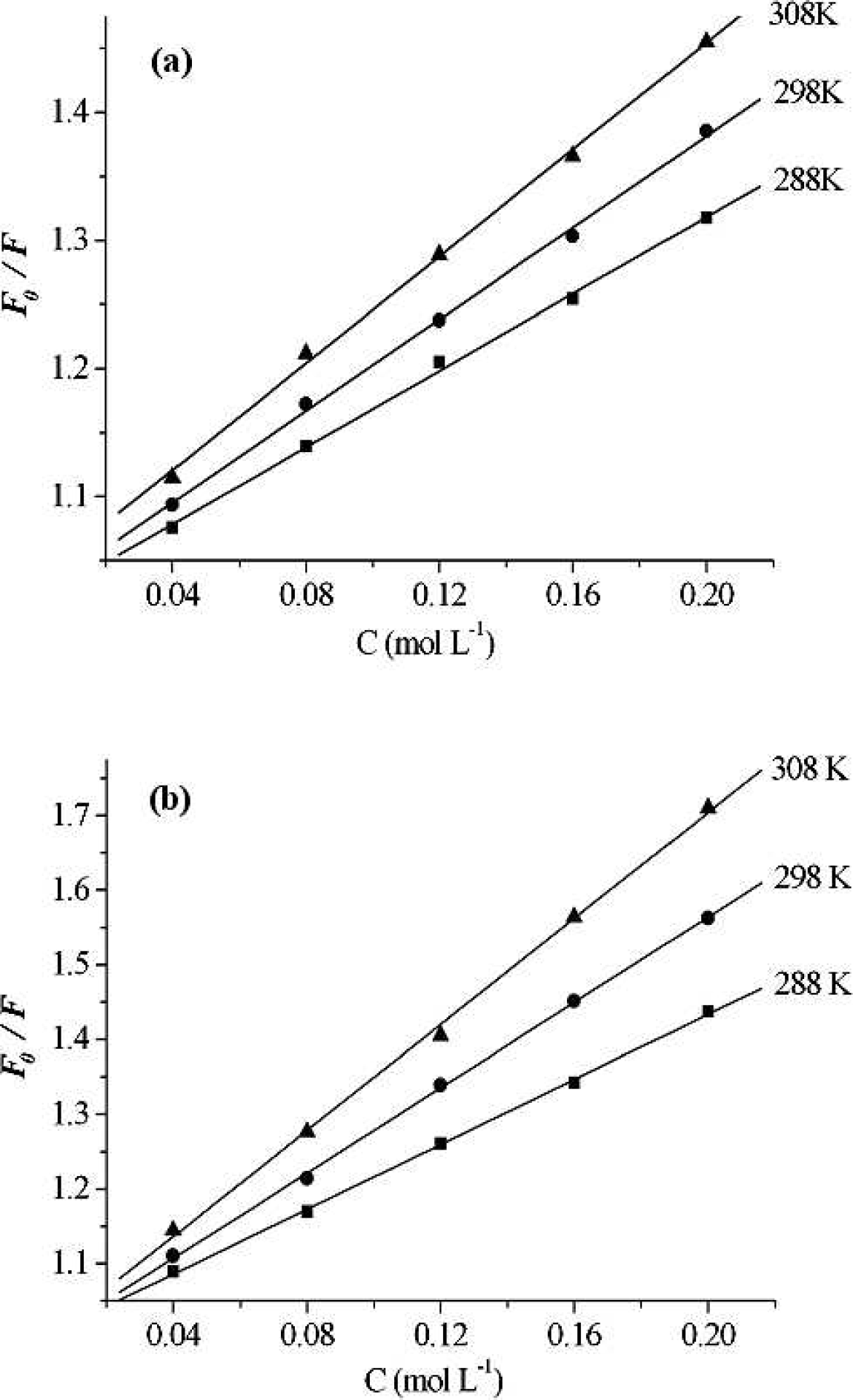
Stern-Volmer plots of pepsin (0.4 g L−1). (
The results shown in Fig. 2 and Table I suggest that the Stern-Volmer plots are linear, and the slope increases with increasing temperature, indicating that the fluorescence quenching is initiated by dynamic collision.
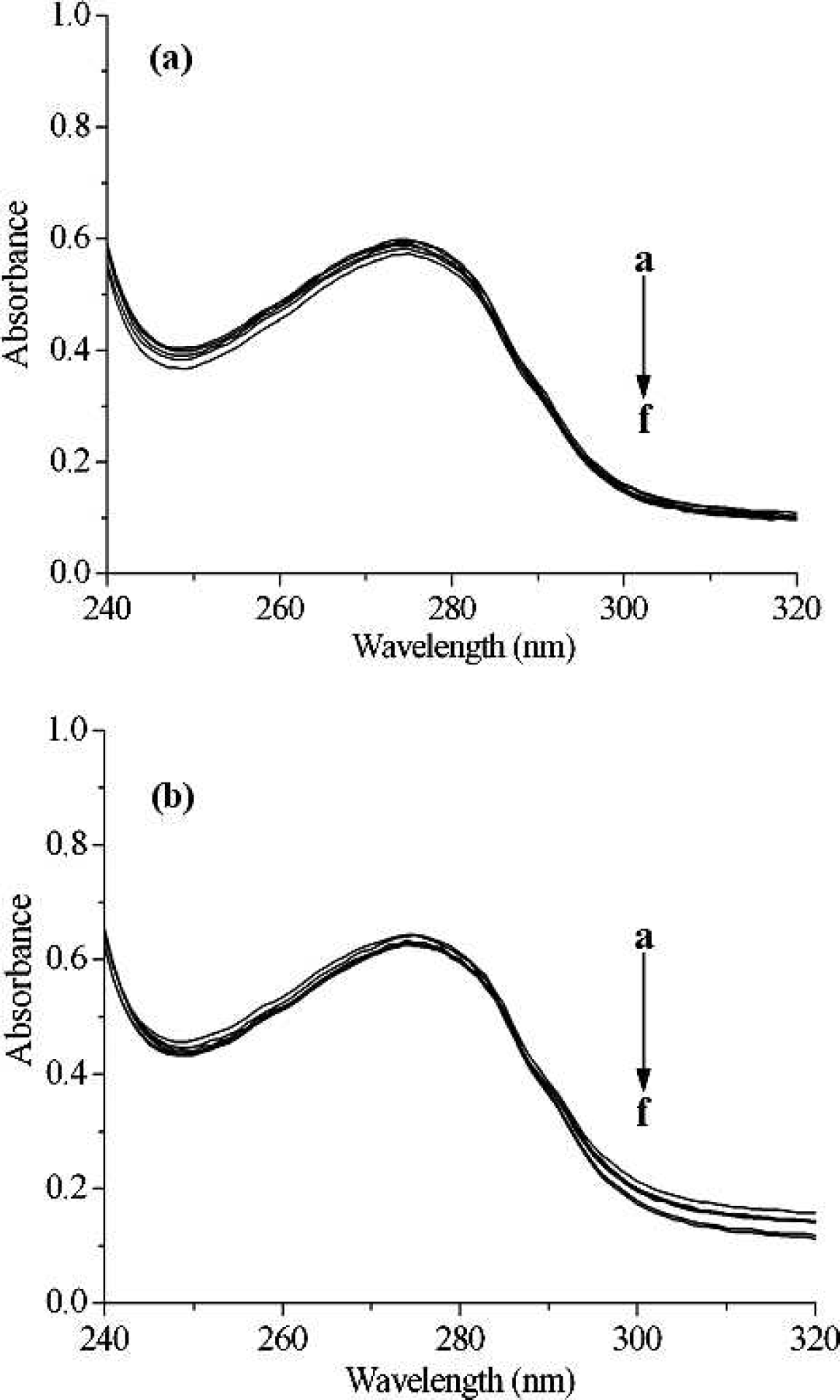
The UV absorption spectra of pepsin in the presence of different concentrations of [C4mim]Cl (
Fluorescence lifetimes (τ) of pepsin, without and with the addition of ILs.

______ = not detectable.
Free-energy change can be estimated from the following equation, based on the quenching constants at different temperatures: 33
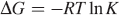
where K is the Stern–Volmer quenching constant at corresponding temperature, T is the experimental temperature, and R is the gas constant. If ΔH and ΔS values do not vary significantly over the temperature range studied, then they can be determined by plotting the binding constants according to the van't Hoff equation: 34
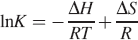
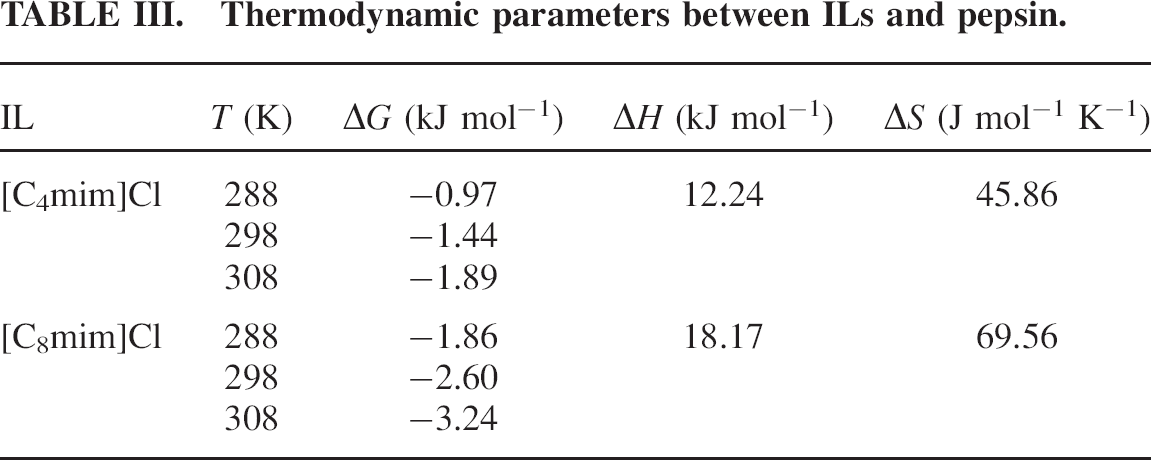
The van't Hoff plots are shown in Fig. 4, and the results are presented in Table III.
Thermodynamic parameters between ILs and pepsin.
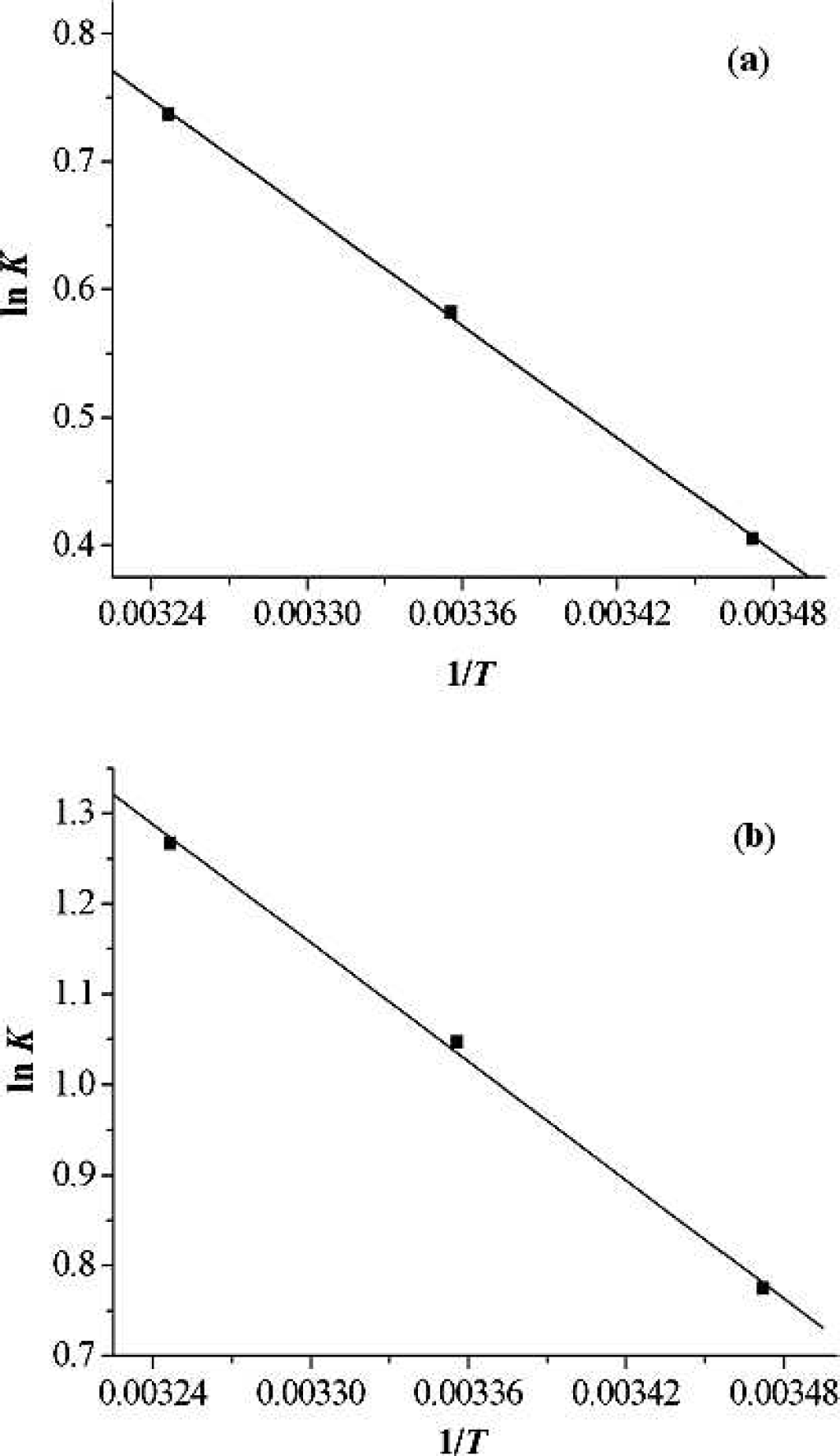
The van't Hoff plots for [C4mim]Cl (
As described in Table III, the negative change in free energy means that the binding process between ILs and pepsin is spontaneous; both ΔH and ΔS have positive values, indicating that hydrophobic interaction could play a major role in the interaction process.34,35 Recently, several research groups reported that the hydrophobicity of ILs increases with alkyl chain length increase.36–38 Since the alkyl chain length of the two ILs increases in the order [C8mim]Cl > [C4mim]Cl, it would be easy to understand why the interaction of pepsin with [C8mim]Cl is stronger than that with [C4mim]Cl.
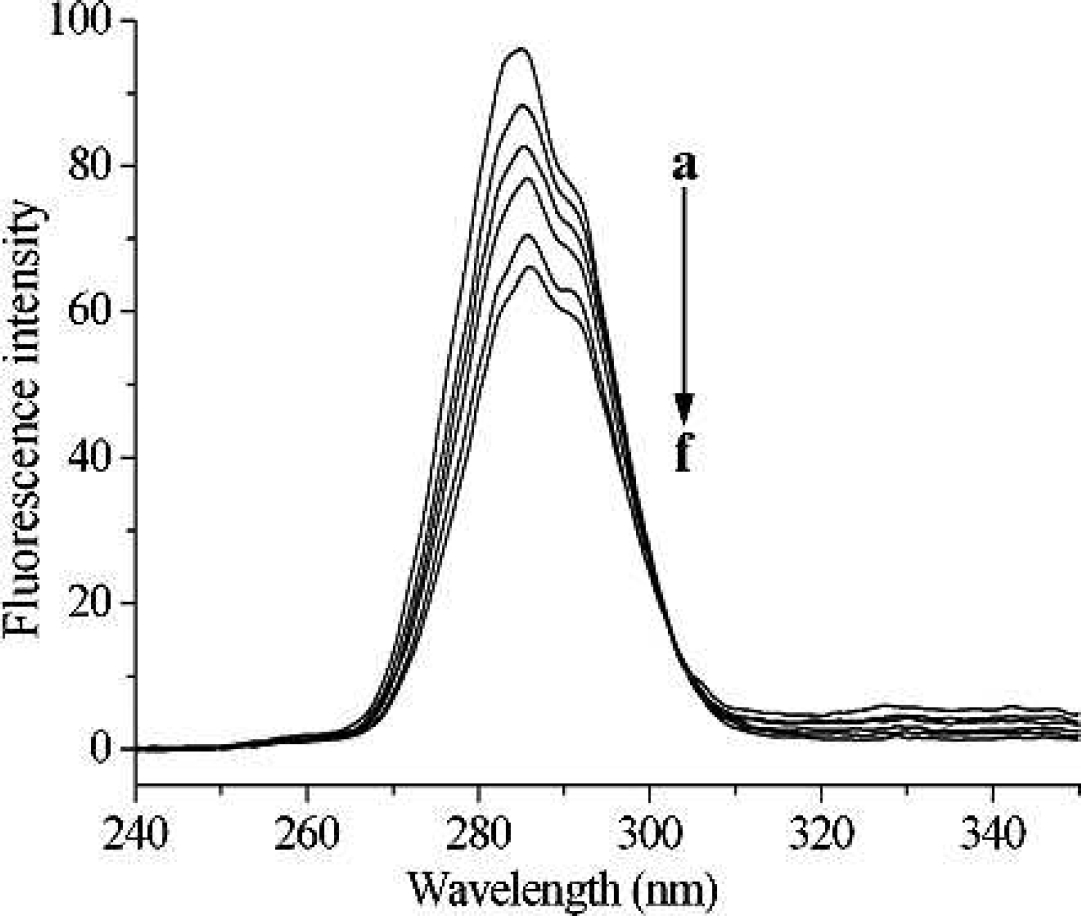
Synchronous fluorescence spectra of pepsin (0.4 g L−1) in the presence of different concentrations of [C8mim]Cl, with ΔΛ = 15 nm. From (
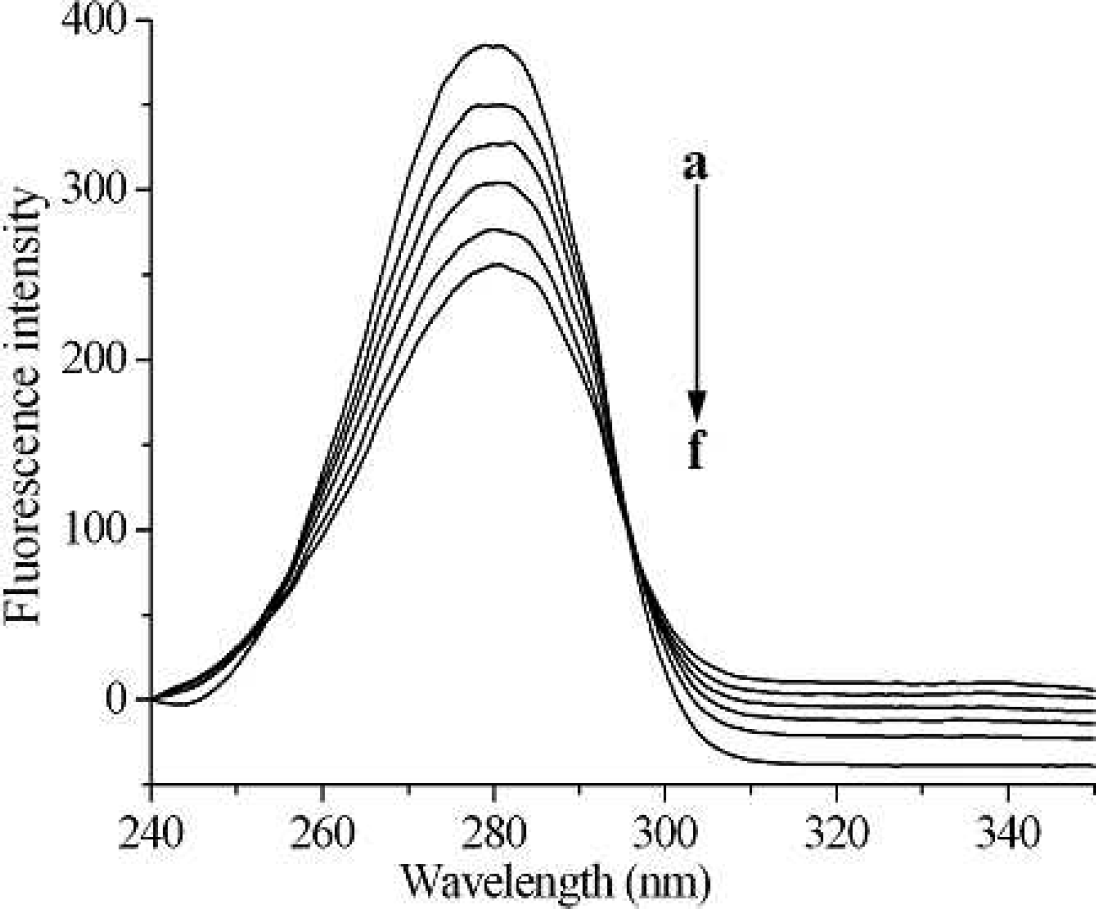
Synchronous fluorescence spectra of pepsin (0.4 g L−1) in the presence of different concentrations of [C8mim]Cl, with ΔΛ = 60 nm. From (
Three-dimensional fluorescence spectral characteristic parameters of pepsin in the absence and presence of the two ILs.
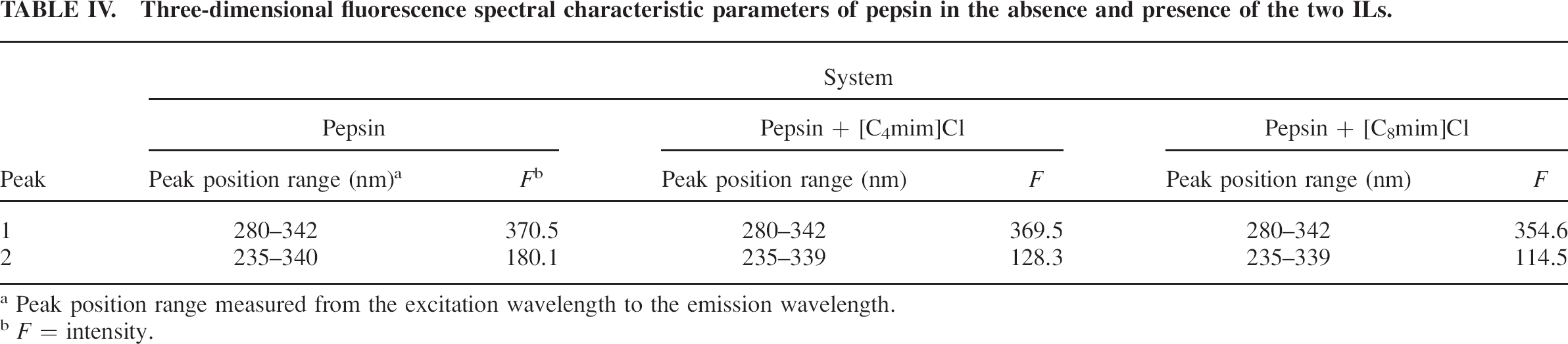
Peak position range measured from the excitation wavelength to the emission wavelength.
F = intensity.
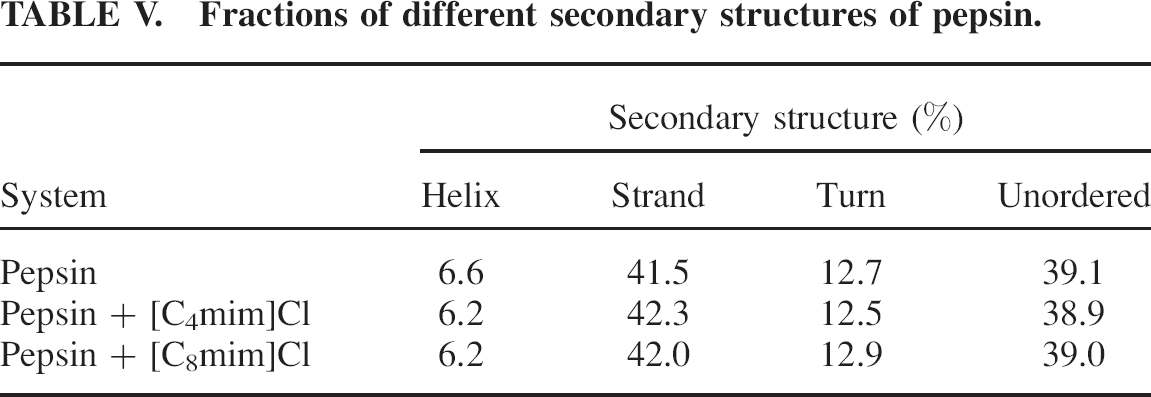
Fractions of different secondary structures of pepsin.
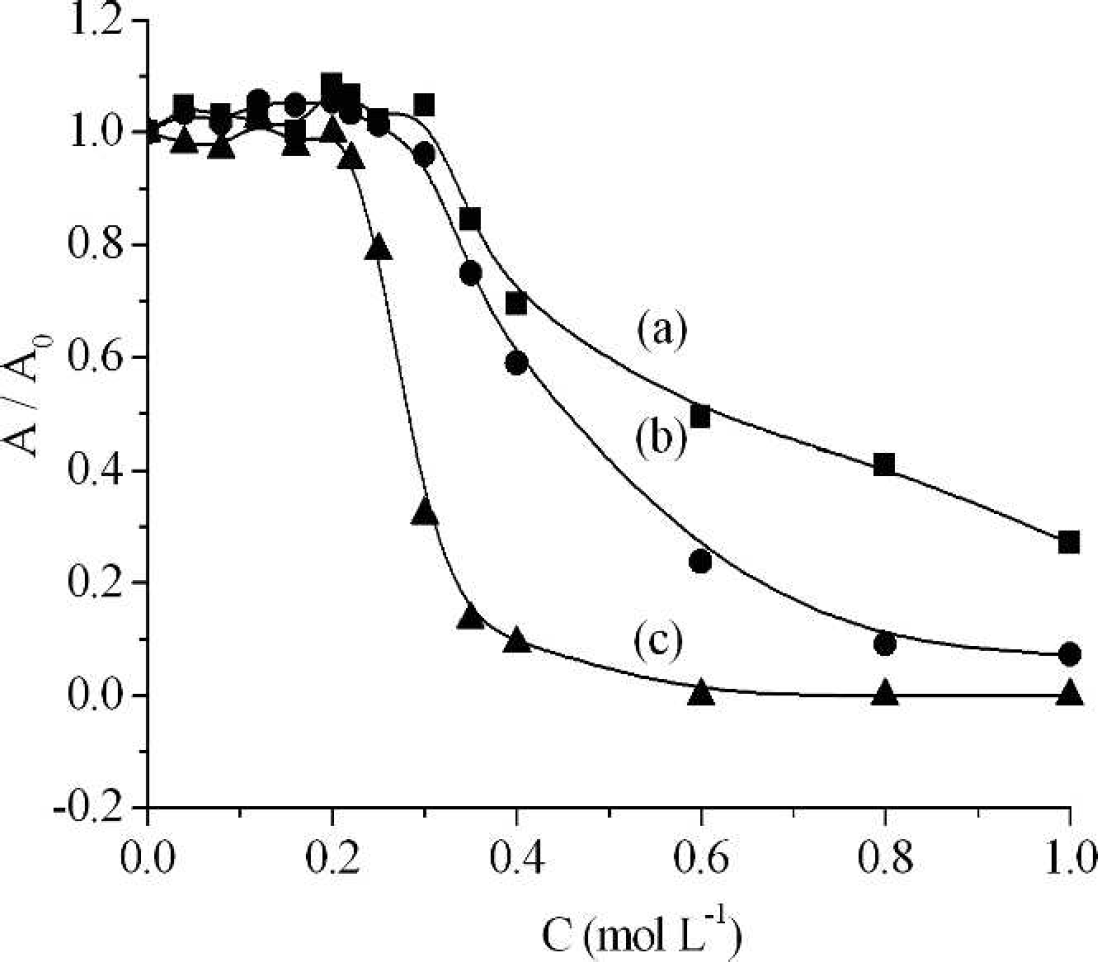
Activity of pepsin in the presence of different concentrations of NaCl (
CONCLUSION
This work demonstrates the interactive nature of ILs and pepsin by using fluorescence, CD, and UV spectroscopic techniques. The results show that the Stern-Volmer quenching constants, KSV, in all cases are directly proportional to temperature, which indicates that the possible quenching mechanism is initiated by dynamic quenching. Low KSV values and no changes in the shapes of fluorescence and UV spectra demonstrate that the interaction between pepsin and ILs is weak, and no pepsin-IL complexes are formed within the studied concentration range (0–0.2 mol L−1 for each IL). Thermodynamic parameters (ΔH > 0 and ΔS > 0) suggest that hydrophobic interaction is the main driving force in the interaction of pepsin with ILs. The IL [C8mim]Cl has stronger quenching ability with pepsin than that of [C4mim]Cl due to the former having stronger hydrophobicity. Synchronous and three-dimensional fluorescence spectra indicate that the microenvironments of tryptophan and tyrosine residues do not change; enzyme activity measurements suggest that at lower concentration levels (<0.22 mol L−1 for [C8mim]Cl and <0.30 mol L−1 for [C4mim]Cl), the activity of pepsin remains constant; however, the enzymatic activity decreases remarkably with further increasing the IL concentrations. Finally, the presence of [C8mim]Cl or [C4mim]Cl does not improve the thermal stability of pepsin, indicating that dialkylimidazolium chloride-water mixtures are poor solvents for biocatalytic reactions. However, it remains possible that other water-miscible ILs can provide better enzyme activity for use in biocatalytic processes, and continued research is required.
Footnotes
ACKNOWLEDGMENTS
This work was financially supported by the Natural Science Foundation of the Education Department of Henan Province (nos. 12B150017 and 12A150010) and the Foundation of Henan Polytechnic University (No. B2010–14).