
Abstract
Postmortem human brain tissue is an important resource for brain research. Spatial transcriptomics is a novel technology that utilizes formalin-fixed, paraffin-embedded (FFPE) or cryosections, with the former having good cytoarchitecture but poor RNA quality and vice versa for frozen tissue. Sheep brains (n=16) were used to test various protocols that simulate the conditions around the preparation and dissemination of human postmortem FFPE and frozen brain tissue to optimize for future spatial transcriptomic work. FFPE and frozen tissues were investigated for RNA quality, while hematoxylin and eosin-stained cryosections were analyzed by quantifying tissue voids as a proxy for cytoarchitectural integrity. Postmortem interval reduced the RNA integrity number equivalent (RINe) of frozen tissue from 7.2 (24 hr) to 4.8 (168 hr). In FFPE tissue, the percentage of RNA fragments greater than 200 nucleotides (DV200) values ranged from 18.9% to 69.01%, with the highest values observed in samples fixed for less than 24 hr. Pretreatment with liquid nitrogen before –80°C storage resulted in the lowest voids (11.6%) in cryosections, but cryoprotectants had little effect. These findings provide researchers with guidelines for tissue preparation in spatial transcriptomics. However, freezing protocols require further refinement to approach the cytoarchitecture of FFPE tissue:
Keywords
Introduction
The New South Wales Brain Tissue Resource Centre (BTRC) in Sydney, Australia, specializes in the preservation, storage, and dissemination of human postmortem brain tissue, primarily for research into alcohol use disorder. As an open-access biobank, it distributes these tissues to researchers worldwide. As research methods evolve, it is necessary to review and update how tissue is preserved. 1 Maintaining high-quality tissue is crucial in today’s “omics” era of research. In the present study, we aim to optimize brain tissue preservation for use with spatial resolved transcriptomic platforms.
There are approximately 139 brain banks globally, with four in Australia. 2 These repositories contain well-characterized human brain tissue and are invaluable for neurological disease and neuroscience research. Furthermore, brain banks such as the BTRC assist researchers in bridging the gap between animal models and human applications. Yet, with decreasing autopsy rates globally, primarily due to the rise in non-invasive imaging, it is crucial to maximize the potential applications of collected tissue.3,4 Postmortem tissue combined with emerging omics-age technology provides researchers with a unique opportunity to explore the unique human brain function and disease states from novel perspectives. 5
Brain banks typically hemisect a human brain, with one hemisphere prepared as fresh-frozen for molecular analysis and the other preserved as fixed tissue for histological studies. 6 Researchers often combine their findings from the differing hemispheres, assuming the pathology is the same in the contralateral region. However, given the known functional lateralization in the brain, this may be inappropriate. 7 Routinely, the hemisphere to be fixed is immersed in fixative, such as 15% neutral buffered formalin (NBF), for up to 3 weeks before slicing, blocking of high-interest regions, processing, and paraffin embedding. The hemisphere to be frozen is immediately sliced coronally, and areas of interest are dissected before all tissue is placed on a precooled metal tray and into a −80°C freezer until frozen. 8 This routine “slow freezing” method results in good molecular preservation but poor tissue cytoarchitecture due to ice crystal formation during the freezing process. 9 During subsequent thawing, cells rupture, disrupting structures and contents, often leaving pockets of space or voids, 10 and rendering the tissue unsuitable for immunohistochemical and other staining techniques. In addition, this freeze-thaw may result in the dissemination of RNA species away from their cell of origin. The opposite is true for formalin-fixed paraffin-embedded (FFPE) tissues where cytoarchitecture is excellent, but RNA quality is poor. 11 While RNA integrity number equivalent (RINe) is often used to assess RNA quality in fresh tissue, the distribution value percentage of RNA fragments of length >200 nucleotides (DV200) is more appropriate for FFPE samples. RINe is primarily designed for assessing intact RNA, relying heavily on the presence of ribosomal RNA peaks (e.g., 18S and 28S in eukaryotic samples). It works well for fresh or frozen RNA but tends to be misleading when dealing with fragmented or degraded RNA. 12 Moreover, banks may store residual fixed tissue in fixative (i.e., 10% NBF) and re-block from their long-term fixed tissue reserves as required, leading to further RNA degradation.
Recently, a platform called spatial transcriptomics has been developed that combines histochemistry and transcriptomics with kits available for frozen and FFPE tissue. 13 Excitingly, these techniques can explore cell-specific expressions adjacent to hallmark pathologies such as plaques in Alzheimer’s disease (AD) in a single section (from the one hemi-brain). 14 If the cytoarchitecture in cryosections could be improved toward that seen in FFPE sections, the RNA degradation and partial transcriptome issues seen in FFPE tissues could be avoided. Alternatively, if the RNA quality of FFPE tissue, as assessed by DV200, could be consistently improved, then FFPE may be a viable option for reproducible, high-definition spatial transcriptomic analyses. Beyond the technical challenges of tissue preservation, brain banks are under increasing pressure to focus on environmental sustainability. Current practices depend heavily on energy-intensive freezer storage and hazardous fixatives like formalin. While we explore methods to optimize tissue preservation, we must also consider how these protocols affect the environment more broadly and laboratory safety at a local level.
To facilitate this work and preserve human brain tissue, we collaborated with the School of Veterinary Science at The University of Sydney to gain access to ovine necropsy brain tissue. Sheep are an appropriate large mammal model and share many functional and neuroanatomical similarities with humans, including a large gyrencephalic brain, four cortical lobes, subcortical structures, and round skulls. 15 Microscopically, sheep also have distinct cellular cortical layers with similar protein expression patterns and neurochemicals. Sheep are naturally apolipoprotein ε4 homozygotes, and aged sheep develop amyloid plaques and tau tangles. 16 Sheep models have been used for various neurological diseases, including AD, traumatic brain injury, Batten’s disease, Huntington’s disease, and epilepsy.17,18 As such, improving the processing of sheep tissue is beneficial to both human brain banking and those using sheep as a large animal model.
Here, BTRC protocols for preparing either FFPE or cryosections for potential use in spatial transcriptomic workflows were replicated in sheep tissue to explore the effects on residual RNA quality. The RINe was measured to assess the differing combinations of freezing methods and impact on RNA quality. In addition, the effect of potential confounders, such as postmortem interval (PMI) on RINe, was assessed. We anticipated that PMI would have a negligible impact on RINe at low intervals <48 hr, with RINe decreasing gradually over longer time points. We also investigated the effect of fixation time and confounders on DV200 values in FFPE tissue, predicting that as fixation time increased, DV200 would decrease until it reached a stable minimum.
In addition, and associated with improving the cytoarchitecture of cryosections, three different freezing methods were investigated: standard −80°C freezing, powdered dry ice (PDI), and liquid nitrogen (LN2). Furthermore, different cryoprotectant agents (CPAs) were trialed with each freezing method, including alginate, which has been used to prevent ice crystal formation in the food industry. 19 These methods are relatively easy to implement and do not require specialized equipment. It was hypothesized that a combination of either PDI or LN2 freezing with a cryoprotectant agent would result in less ice crystal formation and, thus, provide better cytoarchitecture when compared with traditional freezing methods.
Materials and Methods
Ethics and Tissue Procurement
Sheep were paddock-housed and cared for by the School of Veterinary Science staff for their use in teaching clinical and necropsy skills under The University of Sydney animal ethics committee approval (AREC#2019/1587). Sheep (n=16) were euthanized via rapid intravenous injection of pentobarbital sodium solution (Valabarb; Zoetis; 300 mg/ml; 1 ml per 5 kg, Rutherford, NSW, Australia) by a registered veterinarian and necropsied as part of the Veterinary Science Student pathology curriculum. The brain was removed from the cranial cavity, placed in a plastic container, and stored at 4°C for transfer to the BTRC. While PMI (from death to brain removal) was less than 2 hr for all cases, the actual interval to process the tissue ranged from 18 to 168 hr (from here forth, PMI refers to the interval to processing). A total of 16 brains with sheep ranging in age from 1 to 8 years were collected over 3 months (Table 1) for use in various trials.
Summary of Sheep Used in Each Experimental Arm.
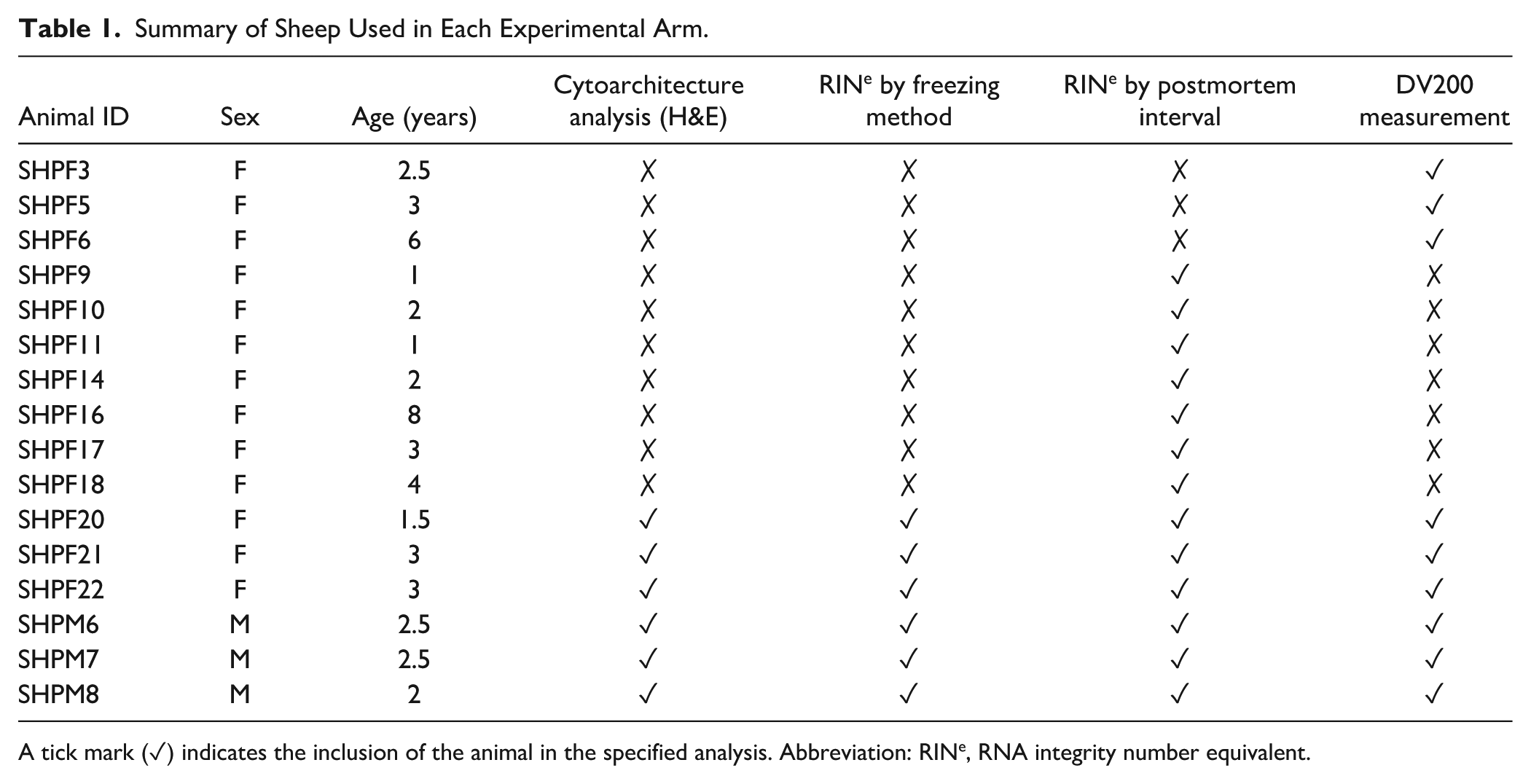
A tick mark (✓) indicates the inclusion of the animal in the specified analysis. Abbreviation: RINe, RNA integrity number equivalent.
Brain Processing
Ovine brains were held at 4°C before dissection. Brains were transferred to ice-cold 0.9% (w/v) saline and rinsed before being weighed and photographed. Any significant damage or abnormalities were noted. Hemispheres were alternatively selected to be fixed or frozen; the brains were hemisected, and then both sides were hand-sliced coronally at 5 mm.
Freezing Methods and Cryoprotectants
The slices to be frozen were laid on an acetate-lined, precooled (–80°C) aluminum sheet tray and photographed before being placed into a –80°C freezer for standard freezing or subjected to a trial freezing method. For frozen protocol experiments, 48 regions containing cortical gray matter and underlying white matter were sampled from six sheep (3M, 3F) as summarized in Table 2. Samples were placed in small plastic weigh boats on a precooled, acetate-lined aluminum sheet tray and treated with one of three cryoprotective agent (CPA) regimes: no CPA(0.9% (w/v) saline), 3% (w/v) sodium alginate (3% A) (#SL117-500G; ChemSupply, SA, Australia) in 0.01 M PBS, or OCT (#4583; Tissue-Tek, Sakura, Torrance, CA) and held at 4°C for 15 min while the CPA penetrated the tissue. Following CPA treatment, trays were frozen using one of three methods: standard “slow freezing” directly in a −80°C freezer, pretreatment by covering with PDI, or submerging in LN2. Once samples were frozen solid, they were individually stored in resealable polyethylene bags at −80°C until sectioning.
Allocation of Cortical Samples to CPA and Freezing Method Combinations.

Each row represents one of the nine CPA-freezing method combinations applied to cortical tissue. A tick mark (✓) indicates the inclusion of the animal in the specified preparation. Columns 3 to 8 indicate which of the six sheep contributed tissue to each combination; the numbers following the hyphen, 24, 96, or 144, refer to hours PMI. LN2-treated samples were not available for SHPM8 and SHPF22 due to experimental constraints. Each animal contributed one tissue sample per combination shown unless indicated otherwise. Abbreviations: CPA, cryoprotective agent; PMI, postmortem interval; LN2, liquid nitrogen.
Sectioning and H&E Staining
Each of the 48 samples described in Table 2 were mounted in OCT and sectioned on a Cryostar NX50 cryostat (#957240; Epredia, Kalamazoo, MI, USA) at −12°C; five 10-µm sections were collected from each block. A single section from each case and each preparation combination was selected at random and fixed in 10% neutral buffered formalin (NBF; #NFB5; POCD, NSW, Australia) for 15 min before staining. H&E staining was performed using Harris hematoxylin, and slides were mounted using DPX (#06522; Sigma-Aldrich, VIC, Australia).
Microscopy and Analysis
Whole slide images of the resultant H&E slides were captured with the Leica Aperio AT DX (Leica Biosystems Nussloch GmbH, Nussloch, Germany), using a 20×/0.75 NA Plan Apo objective for a scanning resolution of 0.503 um / pixel, 8-bit RGB. Images were analyzed using Orbit Image Analysis (Idorsia Pharmaceuticals, Allschwil, Switzerland), a machine learning–based tissue quantification tool. 20 A model was trained to discern tissue from voids, and tissue cytoarchitecture was evaluated by calculating the percentage of voids averaged over three sites for each slide (total area approximately 5 mm2).
CPA-Treated Cortical Samples for RINe Analysis
Samples for RINe analysis were collected from each of the differing freezing preparation combinations before embedding in OCT for sectioning. Tissue samples (20–35 mg) were collected and held at −80°C for subsequent RNA extraction and RINe analysis.
Untreated Cerebellar Samples for RINe Analysis
The cerebellum was detached from six sheep brains and held at 4°C before sample collection at defined PMI time points: 24, 96, 120, 144, and 168 hr. An additional 18 cerebellar samples from seven other sheep, which were used in other trials, also had samples taken for RINe analysis at 24 and 144 hr PMI. Of these, three also contributed samples at 96 hr PMI, and two of these three cases had an additional sample taken at 120 hr PMI. A total of 48 tissue samples, each weighing 20–35 mg, were taken across the varying PMI time points previously listed and stored at −80°C for subsequent RNA extraction and RINe analysis. These sample distributions are detailed in Table 3, which outlines the individual animals and the time points at which they were sampled.
Summary of Samples Used for PMI Versus RINe Analysis.
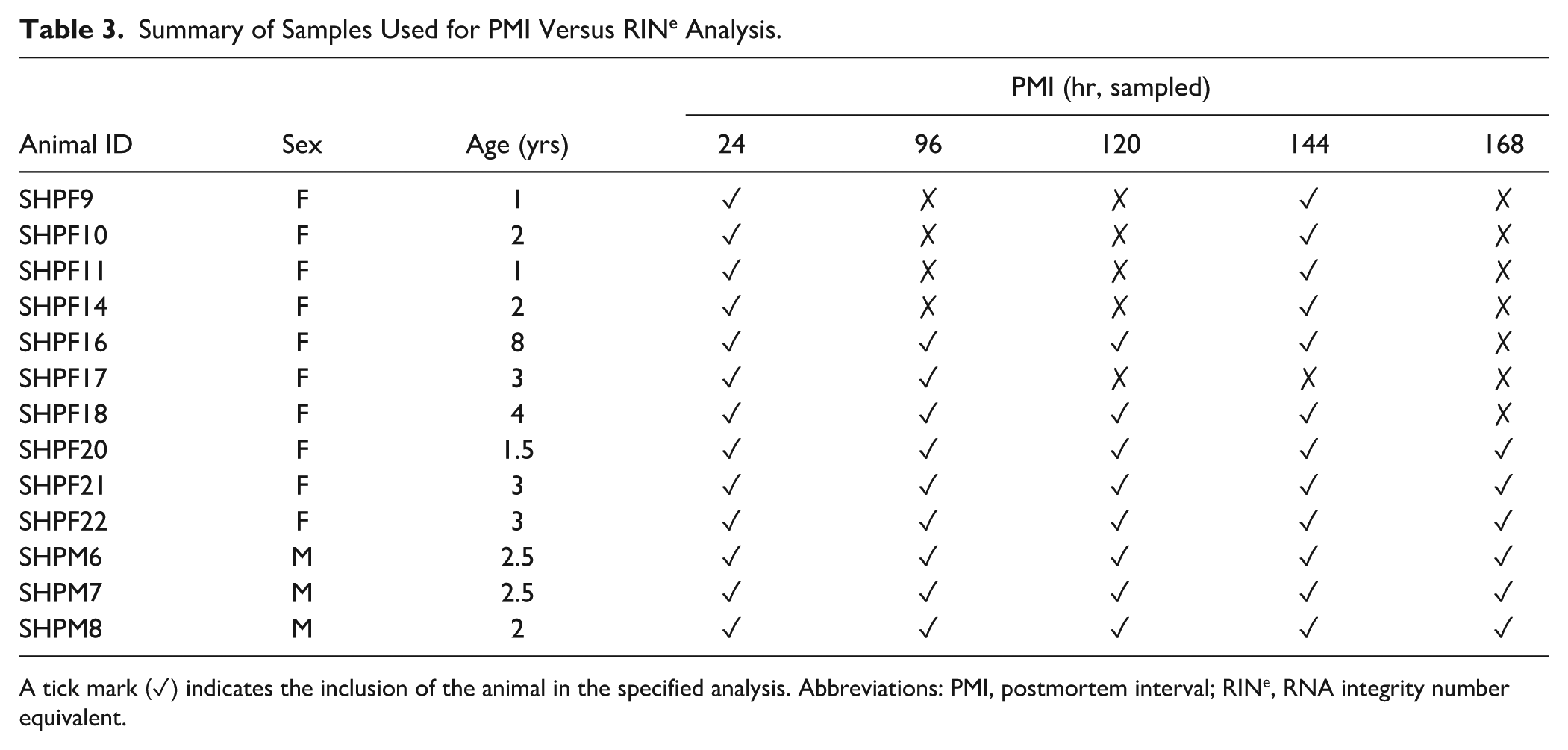
A tick mark (✓) indicates the inclusion of the animal in the specified analysis. Abbreviations: PMI, postmortem interval; RINe, RNA integrity number equivalent.
Fresh Tissue RNA Extraction and RINe Analysis
RNA extractions were paired to reduce batch effects, and all frozen samples from the same animal (n=13) were extracted simultaneously. RNA was extracted using an Isolate II RNA Mini kit (cat. #BIO-52072; Bioline, London, United Kingdom), and the single-step Tri-reagent (Trizol) BCP method was followed per the manufacturer’s instruction. Cerebellar samples were homogenized using a rotor-stator homogenizer, and RNA was eluted in 30 µl of RNase-free water. RNA quality was evaluated first using a Nanodrop 2000 microvolume spectrophotometer (cat. #ND-2000; ThermoFisher Scientific, Wilmington, DE, USA) to ensure adequate RNA concentration and purity. An Agilent TapeStation 4200 automated electrophoresis unit (cat. #G2991BA) was used to evaluate the RINe value 1–10 of each sample, using an R6K ScreenTape as per manufacturer specification (cat #5067-5576, Agilent, Santa Clara, CA, USA).
Formalin-Fixed, Paraffin Embedding
The 5 mm slices to be fixed were placed on gauze in a large plastic container and submerged in 10–20 volumes of 10% NBF for periods ranging from 1 to 180 days. At various intervals over the 180 days, blocks of interest were cut for paraffin embedding as per our human tissue protocol. 21 Blocks were held in 10% NBF before processing as per BTRC protocol 8 and embedded using Paraplast Plus Paraffin (125387-89-5; Sigma-Aldrich). The subsequent FFPE blocks were sectioned at 10 µm (HM 325 Manual Microtome; ThermoFisher Scientific). Three 10-µm FFPE sections per block were collected in a 1 ml tube for RNA extraction and analysis.
Fixed Tissue RNA Extraction and DV200 Analysis
FFPE tissue was collected for DV200 analysis at differing fixation durations and PMI (n=24) from nine different sheep and was extracted using an RNeasy FFPE Kit (#73504; Qiagen, Hilden, Germany) as per the manufacturer’s instructions. An Agilent TapeStation 4200 was used to measure the DV200 of each sample, using an R6K ScreenTape as per manufacturer specification (#5067-5576; Agilent, Santa Clara, CA, USA).
Data Analysis
Data were collated, and statistical analyses were conducted in RStudio (v2024.04.2+764). Levene’s test for homogeneity of variances was performed first, followed by a one-way analysis of variance (ANOVA) to determine group differences. Where the ANOVA was statistically significant (p<0.05), a post hoc Tukey HSD test was conducted to determine pairwise differences in freezing methods and CPAs. The effect of PMI on RINe was investigated similarly. DV200 data were visualized using heatmaps created with ggplot2 (Rstudio), and subsequent surface interpolation was performed using the MBA package to visualize the effect on DV200 at short <7 days and long, >1 month, time points. Data were visualized using RStudio (2024.04.2+764) and GraphPad Prism 10 (v10.2.3).
Results
Freezing Preparation and Voids
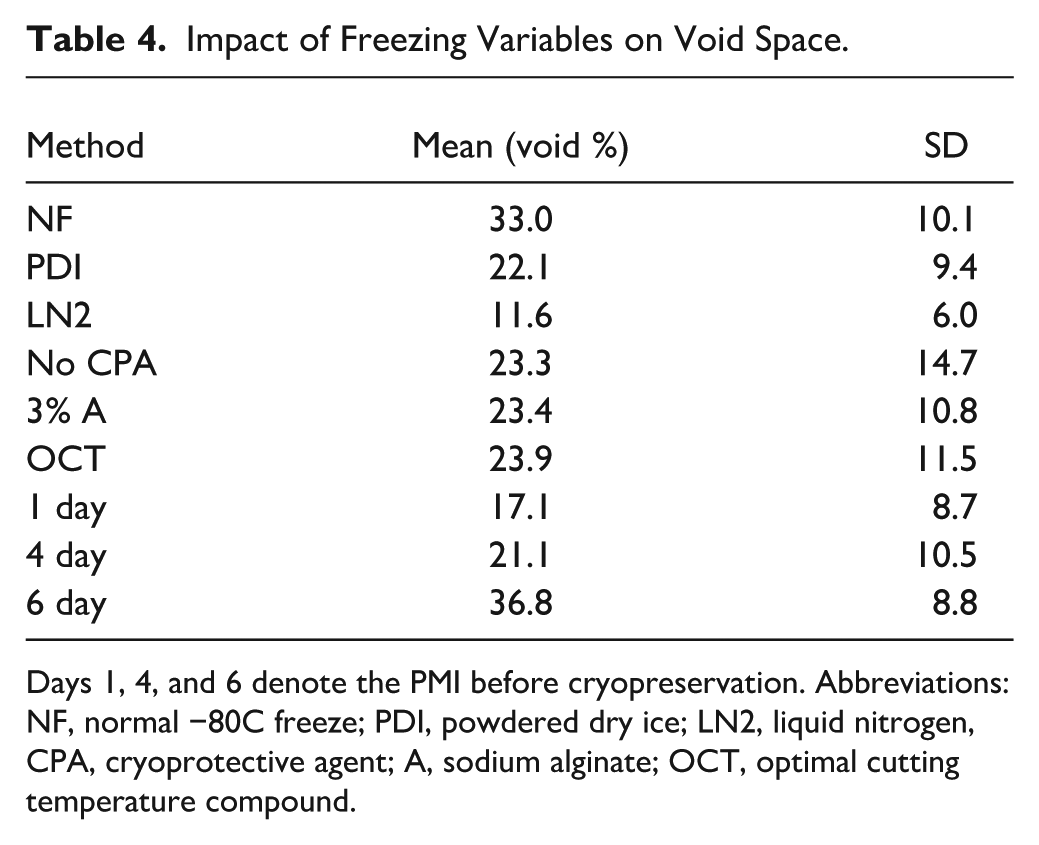
The full allocation of CPA and freezing method combinations applied to each sample is shown in Table 2. This design enabled a systematic comparison of structural preservation across all nine treatment conditions. The three freezing methods were significantly different (p<0.0001) in terms of the voids seen in H&E-stained cryosections, but CPAs were not (p=0.99) (Fig. 1). The average area occupied by voids across all cryosections was 23.5 ± 12.7%, with liquid nitrogen (LN2) freezing the least damaging method, yielding a mean void area of 11.6 ± 6.0% (see Table 4 for “LN2” and other subgroup summary statistics and Appendix Table A1 for all data).
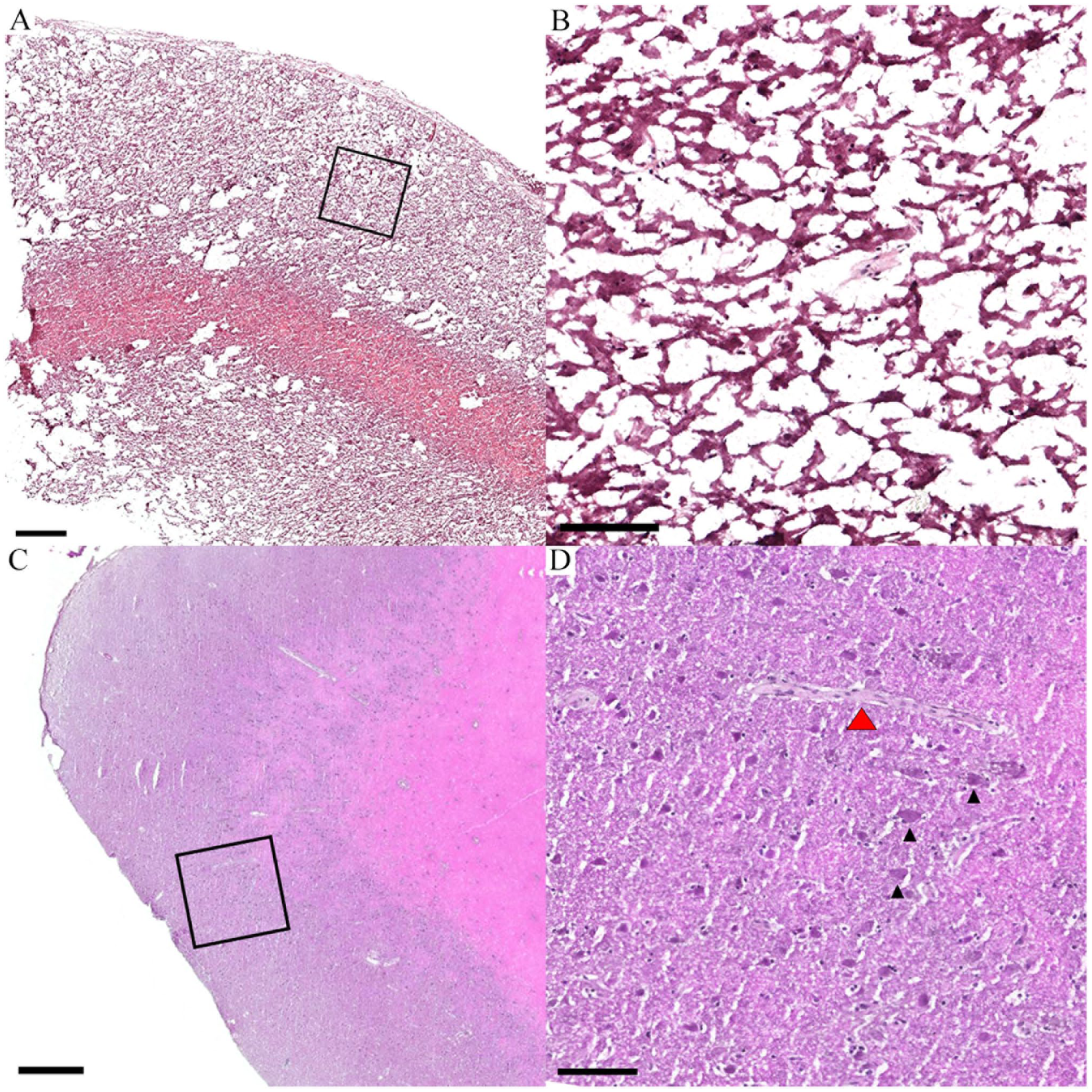
The effects of freezing method on brain cytoarchitecture. A composite image of low- and high-resolution photomicrographs from haematoxylin and eosin-stained sheep necropsy brain cortex cryosections shows: A. A low-resolution view of tissue prepared as per current human brain freezing protocols (−80°C or normal freeze (NF), no CPA) B. A higher resolution view of the inset (black box) from ‘A’ demonstrating the loss of cytoarchitecture in the cortical grey matter. C. A low-resolution view of tissue frozen with liquid nitrogen + OCT D. A higher resolution view of the inset (black box) from ‘C’ . Here, pyramidal neurons (black arrowheads) and a blood vessel (red arrowhead) are clearly visible in the cortical grey matter. Scales bar A, C = 500 μm; B, D = 100 μm. CPA = Cryoprotective agent, OCT = Optimal cutting temperature compound.
Impact of Freezing Variables on Void Space.
Days 1, 4, and 6 denote the PMI before cryopreservation. Abbreviations: NF, normal −80C freeze; PDI, powdered dry ice; LN2, liquid nitrogen, CPA, cryoprotective agent; A, sodium alginate; OCT, optimal cutting temperature compound.
The standard −80°C normal freezing method (NF) caused the most void area with poor feature resolution and an irregular border (Fig. 1A). Upon closer inspection, there was a loss of structural integrity and distinctive cytoarchitecture (Fig. 1B). Conversely, tissue frozen with LN2 (Fig. 1C) had few voids, regular borders, and relatively high structural integrity. For example, layer V pyramidal neurons were an obvious cytoarchitectural feature (Fig. 1D). Subsequent pairwise comparisons revealed statistical differences between NF (33.0 ± 10.1%) versus powdered dry ice (PDI; 22.1 ± 9.4%; p=0.002), NF versus LN2 (p<0.0001), and PDI versus LN2 (p=0.008; Fig. 2A). A longer postmortem interval (PMI) resulted in greater tissue voids at 6 days (36.7 ± 8.8%) compared with 4 days (21.1 ± 10.5%; p<0.0001) and 1 day (17.1 ± 8.7%; p=0.0002) albeit there was no significant difference between 1 and 4 days (p=0.41) (Fig. 2B).
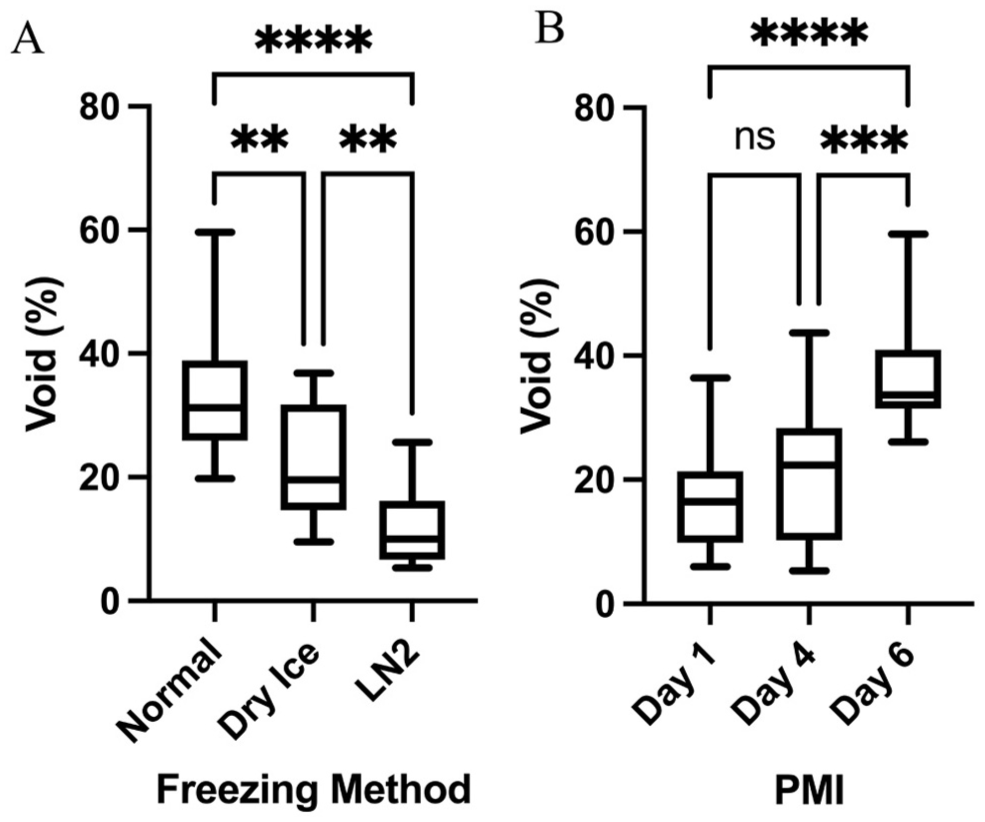
The determinants of tissue voids in cryosections. A. A box plot shows the relationship between the freezing method and percentage of voids (space) seen in hematoxylin and eosin-stained cryosections. Voids are an artifact of the freeze-thaw cycle. B. A box plot shows the relationship between postmortem interval (PMI) and void space. ns = not significant; **p<0.01; ***p<0.001; ****p<0.0001.
RNA Analysis
Impact of Different Freezing Methods on RNA Quality
The RINe did not differ across the various freezing techniques (p=0.6620) or CPA (p=0.2684).
Impact of PMI
The impact of PMI on RINe, independent of the freezing method, was explored using cerebellar samples (Table 5). RINe was different across the PMIs (p<0.0001) and highest on day 1 (7.2±0.7) and lower at later time points, with the lowest mean values observed on days 5 and 7 (both 4.8), although the difference between these two time points was not statistically significant. The RINe values of fresh-frozen samples were related with their respective DV200 values (for full details, see Appendix Table A2). Pairwise comparisons showed differences between 24 hr and other time points but not between any of the time points beyond 24-hr PMI (Fig. 3).
Impact of PMI on RNA Quality.
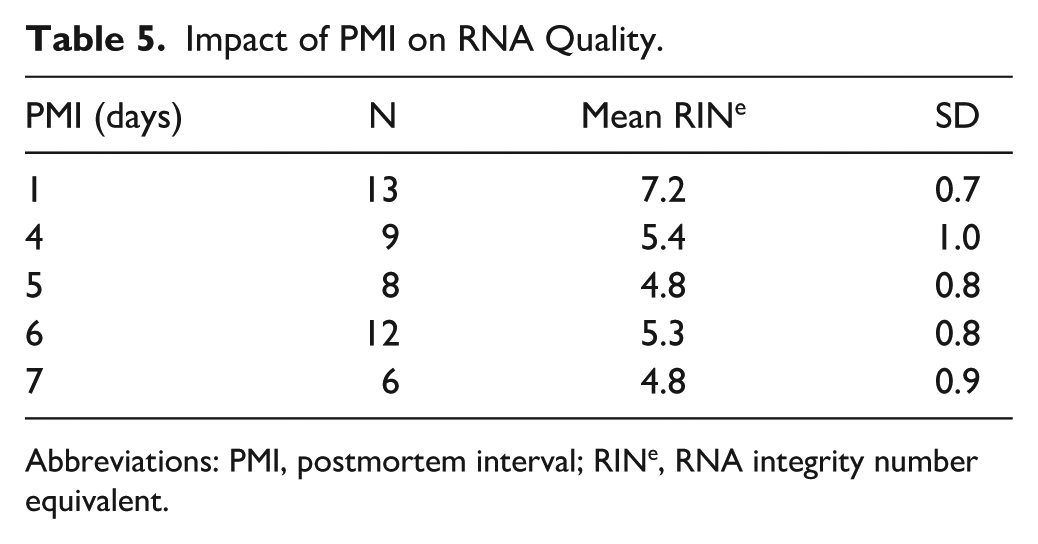
Abbreviations: PMI, postmortem interval; RINe, RNA integrity number equivalent.
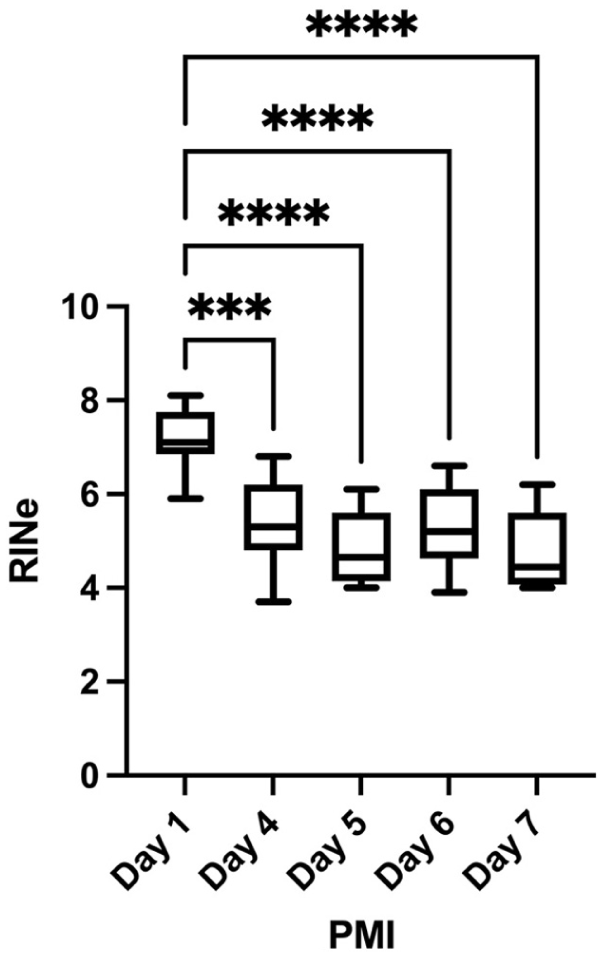
Postmortem interval and RNA quality. A histogram shows the relationship between the time (days) from death to tissue homogenization for RNA quality estimation (RNA integrity number equivalent, RINe). ***p<0.001; ****p<0.0001.
Impact of Fixation Time on DV200
FFPE samples (n=24) had expectedly low RINe (mean 2.1). The alternative metric, DV200, is known to be more accurate than RINe for assessing degraded RNA, 12 so it was used here to assess FFPE RNA quality. FFPE sample RNA quality was primarily influenced by fixation duration. DV200 values ranged from 18.90% to 69.01%, with the highest values (>65%) observed in samples fixed for 24 hr (F3, F5), although some variability was evident between the short-fixed samples (F6 = 44.43%). Unlike fresh-frozen tissue, there was no apparent relationship between RINe and DV200 (for full details, see Appendix Table A3). A marked decline in DV200 values occurred within the first week of fixation, followed by a more gradual decrease, resulting in values between 20% and 25% after 6 months of fixation. While samples with shorter PMI ~24 hr typically showed higher DV200 values, some samples maintained relatively high RNA quality despite extended PMI (>90 hr), suggesting that fixation time is more critical than PMI in preserving RNA integrity in FFPE tissue (Fig. 4).
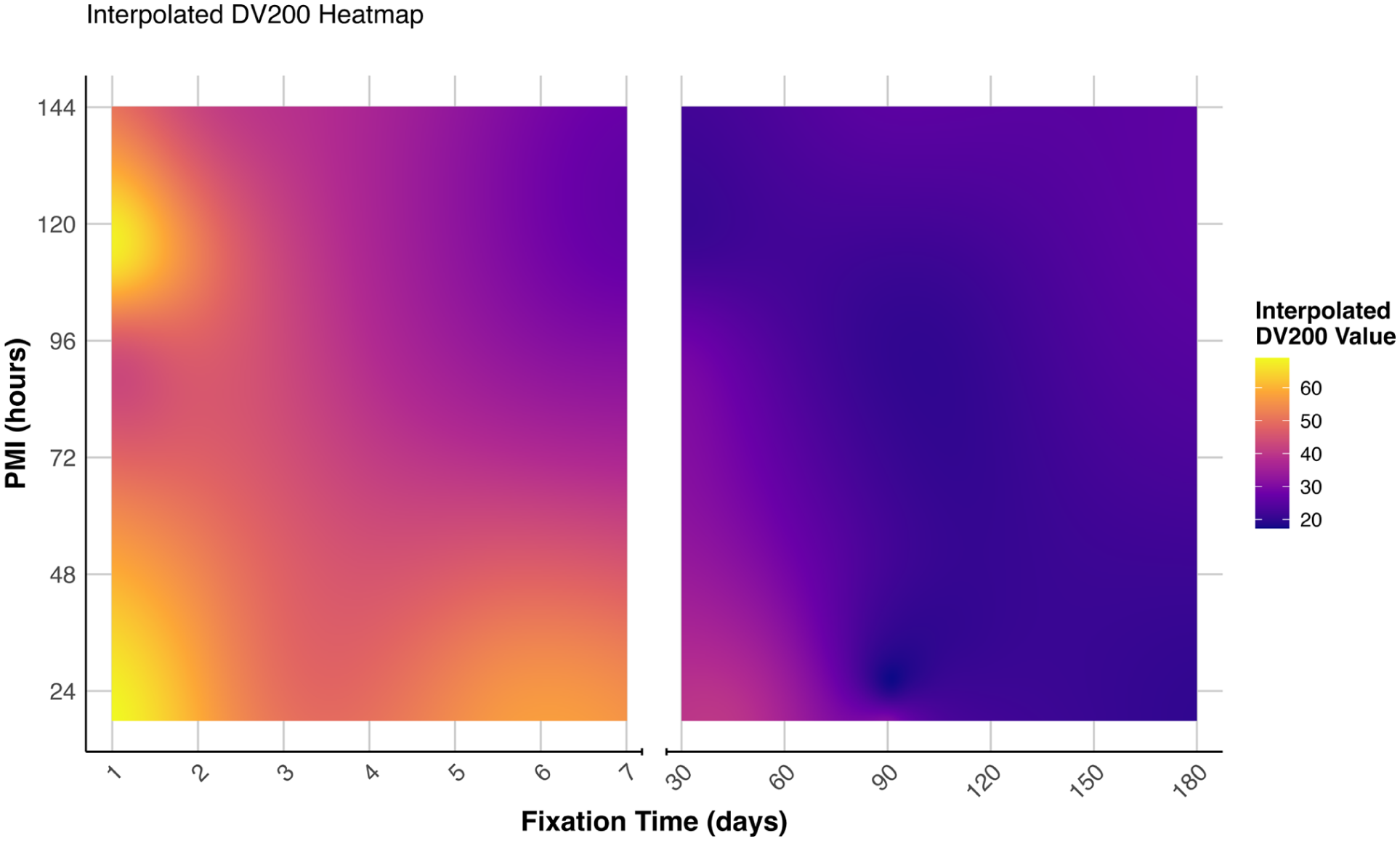
Interpolated DV200 values across fixation time and PMI. A heatmap showing the relationship between tissue fixation duration (days), PMI (hours), and RNA quality (DV200%). The color gradient represents DV200 values, with yellow indicating higher values (better RNA quality) and purple indicating lower values. The plot is split into two time ranges: 1–7 days and 25–180 days and was generated in R (2024.04.2+764).
Discussion
Brain banks serve as crucial resources for advancing our understanding of neurological disorders, facilitating the development of treatments, and providing invaluable insights into human brain function. 22 Organizations like the BTRC enable researchers to bridge the gap between animal models and human tissue, validating their findings in clinically relevant samples or providing primary data in diseases unique to the human brain. However, the traditional brain banking approach of hemisecting the brain—fixing one half for cytoarchitectural studies while freezing the other for molecular analyses—presents inherent limitations. This practice assumes pathological symmetry across contralateral regions, potentially overlooking the known functional lateralization of the human brain. 7 Furthermore, the size of the brain has meant that pragmatic but relatively slow approaches to freezing and fixation of the two hemi-brains of each donor have been typically employed. 23
The emergence of spatial transcriptomics has transformed this landscape by enabling simultaneous analysis of cytoarchitecture and molecular profiles within the same tissue section. This technological advancement allows unprecedented insights into cell-specific expression patterns and spatial relationships, such as enabling direct comparison of microglia within, adjacent to, or distant from amyloid plaques in AD tissue. 14 However, the success of these analyses heavily depends on tissue preparation methods that can maximally preserve both structural integrity and molecular quality. 24
Currently, several spatial transcriptomic platforms are available for various validated species, accommodating both FFPE and frozen tissue, each presenting distinct trade-offs. While FFPE protocols excel in maintaining cytoarchitecture but yield relatively poor RNA quality, standard freezing methods preserve RNA integrity but often compromise tissue architecture. 25 This dichotomy becomes particularly critical with the advent of high-definition (HD) spatial transcriptomic technologies, where precise maintenance of cellular relationships is paramount. Furthermore, the potential migration of RNA species associated with plasma membrane rupture during freeze-thaw cycles raises concerns about the spatial fidelity of molecular signals.
In this study, we objectively evaluated various freezing methods and cryoprotective agents (CPAs), employing quantitative analysis to assess their impact on improving tissue cytoarchitecture. We also investigated how these preservation methods might affect RNA quality while examining fixation duration on DV200 values for FFPE samples and the influence of postmortem interval (PMI) on both. Our goal was to optimize both fresh-frozen and fixed tissue preparation protocols for future spatial transcriptomic applications.
Cytoarchitecture
Cytoarchitecture plays a fundamental role in spatial biology platforms, as structural integrity is crucial for accurate cell segmentation and cell-to-cell and cell–pathology interactions. The presence of freezing artifacts, primarily due to ice crystal formation, can distort tissue morphology and potentially alter the spatial distribution of biomolecules such as RNA. 10 Our findings demonstrate that the freezing method significantly affects tissue preservation, with LN2 yielding superior results (11.6 ± 6.0% void space) compared with both normal (−80°C) freezing (33.0 ± 10.1%) and powdered dry ice methods (22.1 ± 9.4%).
The effectiveness of LN2 freezing can be attributed to its ultra-rapid cooling rate, which minimizes the time available for ice crystal formation. When tissue freezes slowly, as in conventional –80°C freezing, water molecules have time to aggregate into large ice crystals that physically disrupt cellular structures. 26 LN2 enables near-instantaneous freezing, resulting in the formation of vitreous ice rather than crystalline ice, thereby better preserving cellular architecture. 27 This is reflected in our observations of the regular border and minimal structural damage observed in LN2-frozen tissue sections compared with the irregular borders and poor feature resolution of conventionally frozen samples. We did, however, encounter experimental difficulties with LN2 freezing, with some larger tissue blocks cracking during the freezing process. This is likely due to the thermal-mechanical stress experienced as the large tissue sections freeze. 28
Interestingly, CPAs, which are commonly used to mitigate ice-induced damage, did not significantly affect voids in this study (p=0.99). This unexpected finding suggests that CPA concentration or diffusion within the tissue may not have been sufficient to prevent freezing damage under the tested conditions. CPAs work primarily by replacing intracellular water and reducing ice nucleation; however, their effectiveness is highly dependent on proper penetration and equilibration times. 29 Several factors might explain this result: Insufficient penetration depth, suboptimal timing of CPA application, inadequate concentration, or the specific type of CPA used might not have been optimal for use with brain tissue. These variables warrant further investigation to optimize CPA protocols for better tissue preservation.
Postmortem interval (PMI) emerged as a critical factor in tissue cryopreservation, with samples collected after 6 days showing significantly more voids (36.7 ± 8.8%) compared with both 4-day (21.1 ± 10.5%) and 1-day (17.1 ± 8.7%) intervals. The lack of significant difference between 1 and 4 days suggests a threshold effect, where tissue degradation accelerates after approximately 4 days postmortem. This observation aligns with previous research showing that prolonged PMI leads to tissue degradation, enzymatic breakdown, and RNA degradation, all of which compromise sample integrity. 30
These findings have important practical implications for downstream spatial biology applications. While LN2 freezing provides superior preservation, it requires more specialized handling and safety considerations compared with conventional freezing methods (Table 6). Researchers must, therefore, weigh the trade-off between optimal tissue preservation and practical constraints. Overall, preserving cytoarchitecture in frozen tissue samples requires an integrated approach that considers freezing speed, storage duration, and handling conditions. LN2 appears to be the superior method for minimizing ice artifacts, but optimizing CPA protocols, reducing block fragmentation, and mitigating PMI-related degradation are also critical factors in maintaining tissue integrity for downstream analyses.
Summary of Freezing Methods.
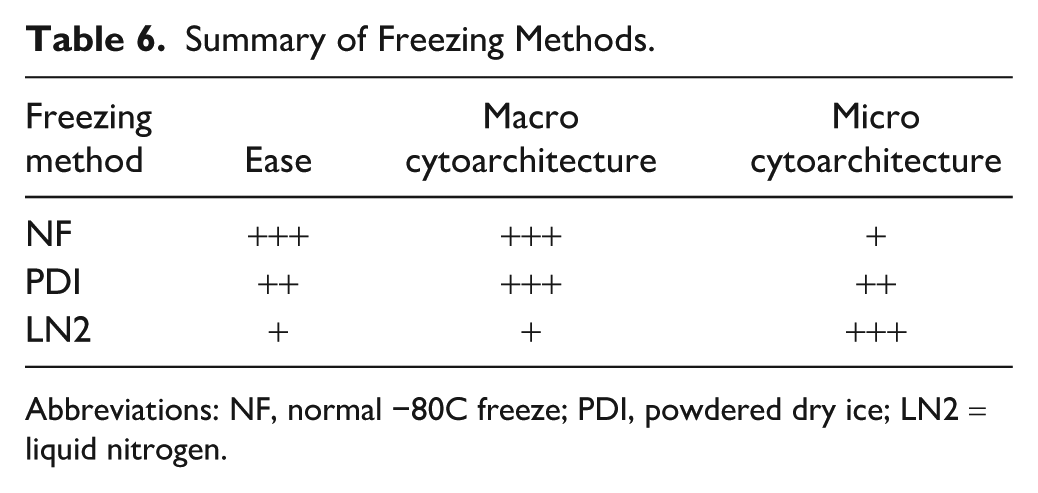
Abbreviations: NF, normal −80C freeze; PDI, powdered dry ice; LN2 = liquid nitrogen.
RNA Quality (RINe)
Molecular integrity is crucial when preserving frozen brain tissue for downstream research applications. The RINe provides a standardized measure (scale 1–10) of RNA quality by evaluating degradation and fragmentation. 31 Higher RINe values indicate better RNA quality, which is essential for applications such as gene expression analysis, microarray studies, and RNA sequencing. In our study, we examined how different preservation methods and postmortem intervals affect RINe values to optimize tissue processing protocols for future spatial transcriptomics.
Interestingly, our analysis revealed that neither the freezing method (p=0.66) nor the use of cryoprotective agents (p=0.27) significantly impacted RINe values. This finding contrasts with our observations regarding cytoarchitecture, where the freezing method substantially influenced tissue integrity. This suggests that while rapid freezing is crucial for preserving cellular architecture, RNA stability may be more resilient to varying freezing rates, at least within the parameters tested in our study.
Much like cytoarchitecture, several factors are thought to impact RNA integrity; of primary concern are sample handling and time intervals/PMI, as well as storage conditions and extraction methods.30,32 PMI emerged as the primary determinant of RNA quality, but this may be due to sampling and extraction limitations. We observed the highest RINe values at 24 hr postmortem (7.2 ± 0.7), followed by a significant decline by day 4 (5.4 ± 1.0). After this initial drop, RINe values appeared to stabilize, with minimal further degradation through day 7 (4.8 ± 0.9). Statistical analysis confirmed significant differences between 24-hr PMI and all later time points (p<0.0001), though comparisons between time points beyond 24 hr showed no significant differences. This pattern suggests a critical window for tissue collection within the first 24 hr postmortem to obtain optimal RNA quality.
Our findings highlight the stark contrast between frozen and FFPE preservation methods. While our frozen samples maintained relatively high RINe values (~6–8), FFPE samples typically yield much lower values (~2.4) due to the extensive processing required, including ambient temperature handling and high-temperature paraffin embedding (>60°C). This difference in RNA quality between frozen and FFPE samples has important implications for spatial transcriptomics applications, where the more complete transcriptome is available in frozen tissue.
Our results emphasize that optimal RNA preservation requires careful attention to PMI and other factors rather than specific freezing protocols. This finding may simplify tissue processing workflows, as teams can focus on rapid tissue collection and basic freezing protocols without requiring complex CPA treatments or specialized freezing methods to maintain RNA integrity. However, when considering spatial transcriptomics applications, the dual requirements of RNA quality and cytoarchitectural preservation necessitate a more nuanced approach to tissue processing. A caveat here is that the impact of agonal effects on RNA quality, which often exceed PMI effects in human studies, was not relevant to our sheep necropsy tissue protocol.
RNA Quality (DV200)
Our analysis of RNA quality in FFPE samples (n=24) revealed that fixation duration is the primary determinant of RNA preservation, with DV200 values ranging from 69.01% in minimally fixed tissue to 18.9% in long-term fixed samples. The relationship between fixation time and RNA quality follows a distinct biphasic pattern: an initial sharp decline in DV200 values during the first week of fixation, likely reflecting the immediate chemical modifications induced by formalin cross-linking with nucleic acids, followed by a more gradual deterioration, reaching a plateau of 20%–25% after 6 months. This secondary phase may represent ongoing degradation processes distinct from the initial cross-linking events. 33 The consistent plateau suggests a baseline level of RNA stability in long-term archived samples, potentially representing molecules resistant to further degradation.
Samples fixed for 24 hr generally had DV200 values above 65%, suggesting that shorter fixation time, while still ensuring complete fixation, is crucial for optimal RNA preservation. It is well documented that formalin penetrates tissue at a rate much faster than it binds tissue and eventually cross-links.11,34 The need to balance RNA preservation and quality, and morphological detail and stability is paramount in FFPE tissue if it is to be used in downstream spatial applications. To allow sufficient fixation while maintaining high-quality RNA, fixation time should be minimized to less than 48 hr for tissue <5 mm thick.
The influence of PMI proved more complex than expected. While shorter PMIs generally corresponded with higher DV200 values, some samples with extended PMI (>90 hr) maintained relatively high RNA quality when fixation time was minimal. This suggests that postmortem degradative processes may be distinct from those induced by formalin fixation and that tissue-specific or environmental factors might influence RNA degradation during the postmortem interval.
These findings have important implications for biobanking protocols, indicating that while minimizing fixation time is crucial for preserving RNA quality, there may be more flexibility in acceptable PMIs than previously thought. However, protocols must still balance RNA preservation with the maintenance of tissue architecture for successful spatial transcriptomic applications.
Comparing RNA Quality Metrics
While the present study did not attempt to directly compare RNA quality metrics across differing (frozen vs. FFPE) tissue preparations, it remains an essential consideration for downstream sample application. The RINe is derived from a combination of several features describing the curve of the electropherogram and is scaled 1–10, 31 with a value of 10 indicating good RNA quality. RINe has been shown to have a strong correlation with downstream sequencing in fresh tissue samples. The DV200 index is the percentage of RNA fragments greater than 200 nucleotides in length. It has been shown to have a stronger correlation with the amount of NGS library product than the RINe in FFPE samples. 12 While both DV200 and RINe can be calculated in frozen and FFPE tissue, the direct comparison of the values from tissues that have undergone different preparation is not recommended.11,12,35
Indeed, in our own hands, we saw little correlation between RINe and DV200 in our FFPE samples (n=24); five of the samples failed to return a RINe at all. The sample with the highest DV200 values, 69.01% returned a moderately low RINe value of 1.5, while samples with RINe values ~2.5 had consistently lower DV200 values ~39%. If the five samples that were below instrument RINe detection were estimated to the minimum detected (1.2), then any relationship between RINe and DV200 disappears (also Appendix Table A3).
Conversely, in our frozen samples (n=48), we did see some correlation between RINe and DV200, with samples with RINe values greater than 7 returning an average DV200 value of ~ 90% (see Appendix Table A2). The differing degradation pattern of RNA from different tissue preparations could also be due to several other factors, including PMI, storage conditions, and extraction protocol. 35 Given the variance between sample preparation and the complexity of the RNA degradation process, it may be worth using alternative metrics such as the paraffin-embedded RNA metric. This method employs a weighted area under the curve analysis to assess RNA quality in FFPE tissue and correlates well with mRNA transcripts and reverse transcription–polymerase chain reaction data, which could be more informative than just the RINe or DV200. 36
Strengths and Weaknesses
Our use of sheep brain tissue as a model system offered controlled experimental conditions but introduced important considerations for translation to human studies. While this approach allowed precise control of postmortem conditions and systematic evaluation of preservation methods, species-specific differences may influence tissue preservation dynamics. 17 The young age of our sheep cohort (1.5–3 years) and the absence of neurodegenerative pathology differ substantially from typical human brain bank cases, where aging and disease processes may affect tissue preservation. 16
Experimental conditions also differed from typical human brain banking procedures. Our samples were immediately refrigerated postmortem and extracted from the cranial vault, whereas human brains often remain in situ until autopsy. This difference may be significant, as the skull and surrounding tissue could buffer temperature changes, alter degradation patterns, and protect from contamination. In addition, our experimental design created an unavoidable sampling gap at 48 hr PMI due to workflow constraints, and we did not assess the impact of agonal factors—both important considerations for human tissue processing. 30
While our quantitative analysis of tissue voids provided an objective measure of freezing-induced damage, it represents only one aspect of tissue preservation. Furthermore, the controlled conditions of our study may not fully capture the variability encountered in human brain banking. These limitations should be considered when applying our findings to human tissue preservation protocols, particularly in the context of neurodegenerative disease research.
In addition, we did not perform spatial transcriptomics in the study due to platform limitations for analyzing sheep tissue and financial constraints. However, our results provide the necessary groundwork for such applications by identifying preservation strategies that maintain key quality metrics. Our preliminary data with human postmortem tissue suggest that a 2-fold difference in DV200 for unstained FFPE sections results in a near 6-fold difference in the genes subsequently detected by the Visium HD platform (Aladyeva et al. unpublished data; also see Appendix Table A4). Spatial transcriptomics offers unprecedented opportunities for brain research by enabling simultaneous molecular and structural analyses. However, current preservation methods present a critical trade-off: FFPE tissue maintains excellent cytoarchitecture but compromises RNA integrity, while frozen tissue preserves RNA quality but often disrupts tissue architecture. This dichotomy is particularly concerning as RNA degradation may selectively affect specific transcripts, potentially masking disease-related molecular signatures. 37
This work is intended as a reference for labs working up spatial biology protocols for human clinical samples and as is often the case with postmortem brain tissue choosing between FFPE and frozen tissue. Our findings using sheep brain tissue demonstrate that liquid nitrogen freezing offers superior structural preservation compared with conventional methods but also introduces the risk of block fragmentation. Surprisingly, CPA treatments did not significantly improve outcomes. Postmortem interval emerged as a critical factor affecting both tissue architecture and RNA quality, with optimal preservation achieved within 24 hr. In fixed tissues, shorter fixation time resulted in higher DV200 values, suggesting better RNA quality. These insights will inform protocol refinements at brain tissue repositories to better serve downstream spatial transcriptomic applications.
Looking ahead, the field of brain banking faces broader challenges beyond technical optimization. Climate change considerations necessitate a balance between tissue preservation requirements and environmental impact. While transitioning to all-frozen storage could eliminate formalin use and its associated hazards, the increased energy demands of expanded freezer capacity raise concerns about sustainability. Future developments must, therefore, address both the practical and technical demands of spatial transcriptomics and the environmental responsibility of long-term tissue storage.
Footnotes
Appendix
The relationship between DV200 and UMI in human postmortem brain tissue samples (Aladyeva et al. unpublished data).
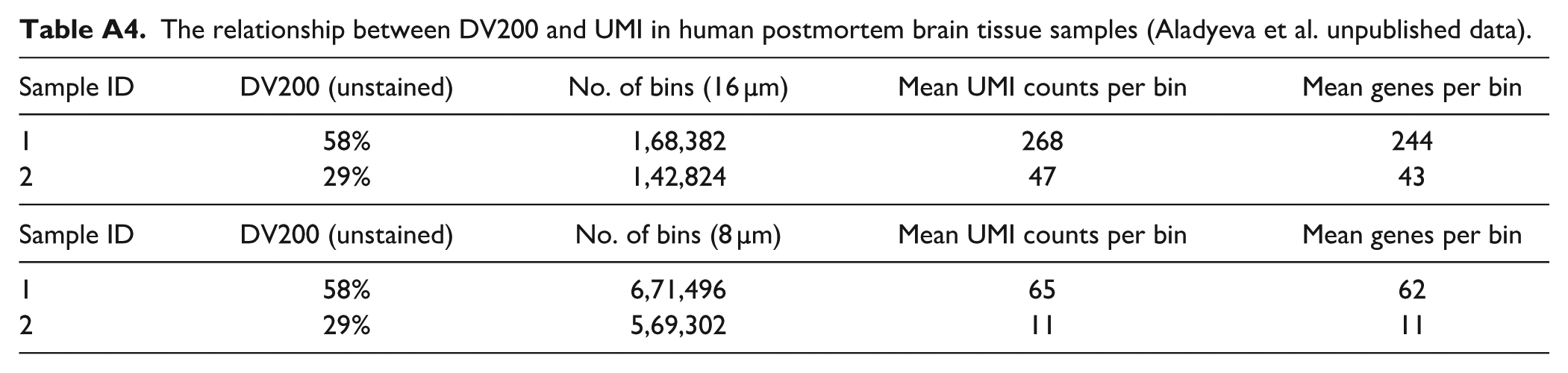
| Sample ID | DV200 (unstained) | No. of bins (16 µm) | Mean UMI counts per bin | Mean genes per bin |
|---|---|---|---|---|
| 1 | 58% | 1,68,382 | 268 | 244 |
| 2 | 29% | 1,42,824 | 47 | 43 |
| Sample ID | DV200 (unstained) | No. of bins (8 µm) | Mean UMI counts per bin | Mean genes per bin |
| 1 | 58% | 6,71,496 | 65 | 62 |
| 2 | 29% | 5,69,302 | 11 | 11 |
Acknowledgements
We acknowledge and thank Sydney Microscopy & Microanalysis, the University of Sydney node of Microscopy Australia, for its technical assistance.
Competing Interests
The authors declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Author Contributions
M.N.: Conceptualization; methodology; writing (original draft); investigation, data curation, formal analysis, visualization. C.C.S.: Conceptualization; methodology; data curation; writing (review and editing). D.M.: Methodology; investigation; writing (review and editing). J.S.: Formal analysis; validation; writing (review and editing); project administration. M.G. and N.H.: Resources. G.T.S.: Supervision; writing (review and editing); resources; funding acquisition; project administration. All Authors contributed to editing and reviewing the final manuscript.
Funding
The authors disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: Research reported in this publication was supported by the National Institute of Alcohol Abuse and Alcoholism of the National Institutes of Health under Award Number R28AA012725. The content is solely the responsibility of the authors and does not represent the official views of the National Institutes of Health.
ORCID iDs
Data
All data are included in the main manuscript or Supplementary data.