
Abstract
The functions of phosphatase and tensin homolog deleted on chromosome 10 (PTEN), a tumor suppressor, depend on its subcellular localization. At the plasma membrane, PTEN dephosphorylates phosphatidylinositol-3,4,5-triphosphate to inhibit AKT signaling, whereas nuclear PTEN contributes to the maintenance of genomic stability. Fluorescent proteins (FPs) are widely used to assess PTEN’s subcellular localization; however, both the intrinsic properties of FPs (e.g., molecular size) and the choice of FP can influence subcellular localization. This study aimed to determine whether FP fusion affects the subcellular localization of PTEN and its mutant forms under conditions involving DNA damage. mCherry typically promotes cytosolic localization of FP-fused PTEN, indicating that FP selection may affect the interpretation of localization data. Furthermore, FP fusion increases the molecular size of the truncated PTEN fragment, which may impede its nuclear import. In comparison, PTEN mutants such as PTENK13R or PTENA4, which predominantly localize to the cytoplasm or nucleus, respectively, show a minimal dependence on the type of FP. Similarly, DNA damage-induced nuclear accumulation of PTEN appears to be independent of the FP type. These findings underscore the importance of carefully considering the effects of FP fusion when investigating the mechanisms regulating the nuclear translocation of PTEN.
Introduction
Phosphatase and tensin homolog deleted on chromosome 10 (PTEN) is a critical tumor suppressor in multiple organs, including the endometrium and the brain. 1 PTEN exhibits a dual-specificity phosphatase activity toward both lipids and protein substrates. At the plasma membrane, its canonical substrate, phosphatidylinositol-3,4,5-triphosphate (PIP3), is dephosphorylated to phosphatidylinositol-4,5-bisphosphate (PIP2). Through this reaction, PTEN antagonizes phosphatidylinositol-3 kinase (PI3K) activity and negatively regulates the AKT signaling pathway, thereby suppressing cell proliferation, migration, and survival. 2 In addition to the cytoplasmic role, accumulating evidence highlights the importance of PTEN’s nuclear functions in tumor suppression. 3 Nuclear PTEN reduces MAPK phosphorylation and cyclin D1 expression, maintaining the G0-G1 phase of the cell cycle.4,5 Furthermore, nuclear PTEN contributes to chromosomal stability, in part through its direct interaction with centromere-specific binding protein C (CENP-C), which is essential for centromere integrity. 6 Given that PTEN’s function is closely associated with its subcellular localization, elucidating the molecular mechanism governing its nuclear transport is of significant importance.
PTEN fused to green fluorescent protein (GFP) or yellow fluorescent protein (YFP) has been used to investigate the mechanisms underlying its subcellular localization.7 –11 However, the potential impact of fluorescent protein (FP) tags on the subcellular localization of PTEN is unclear. Whether FP tagging affects the localization patterns of PTEN mutants typically found in the nuclei or cytoplasm is also unclear. Lysine13 is indispensable for PTEN’s nuclear transport, and substitution of this residue with arginine (K13R) prevents its nuclear entry. Mono-ubiquitination at K13 is actually required for the nuclear translocation of PTEN.7,10 The nuclear transport of PTEN is regulated by phosphorylation at Ser380/Thr382/Thr383/Ser385 residues.9,10 PTENA4, in which these four residues are replaced with alanine (A4), accumulates predominantly in the nucleus. PTEN also accumulates in the nucleus in response to DNA damage. We previously reported that oxidative stress induced nuclear translocation of PTEN-GFP. 7 Nevertheless, whether the properties of FP tags themselves affect the nuclear translocation of PTEN under conditions of DNA damage is unclear.
In this study, we aimed to determine whether FP fusion affects the subcellular localization of PTEN and its mutant forms under conditions of DNA damage. We compared the intracellular distribution of PTEN constructs fused to different FP tags to assess their effects on PTEN localization dynamics.
Materials and Methods
Plasmids
pmEGFP-N1, pmEGFP-C1, pmYFP-N1, pmCherry-N1, pmCherry-C1, and pmScarlet3-N1 plasmids were kindly provided by Dr. Nakamura (Kyoto University, Kyoto, Japan).12,13 pmApple-N1 was purchased from Addgene plasmid #54567 (Watertown, MA, USA). PCR-amplified PTEN, PTENK13R, PTENA4, PTENK13R,A4, and PTEN350 were cloned into the pmEGFP-N1, pmYFP-N1, and pmCherry-N1 plasmids to generate PTEN and its mutants fused with FPs. PCR-amplified mEGFP was cloned into the pPTEN350-mCherry plasmid to construct the PTEN350-GFP-mCherry plasmid.
Cell Culture and Transfection
HEK293T and HeLa cells were maintained in Dulbecco’s Modified Eagle Medium (DMEM) (043-30085; Fuji Film, Tokyo, Japan) supplemented with 10% fetal bovine serum (FBS) (A525671; Gibco, Waltham, MA, USA). Plasmid transfections were performed using Lipofectamine3000 (L3000015; Invitrogen, Waltham, MA, USA) to express each construct in HEK293T and HeLa cells following the manufacturer’s protocols.
Reagents
Camptothecin (038-18191) and Etoposide (055-08431) were purchased from Fujifilm.
Live-Cell Imaging
The day after plasmid transfection, samples were imaged using an Olympus FV1000 confocal laser-scanning microscope with the FV1000-ASW software (Olympus, Tokyo, Japan). Quantification of fluorescent signals was performed using NIH ImageJ, and statistical analyses were performed using GraphPad Prism 10 (GraphPad Software, Inc., San Diego, CA, USA). Nuclear and cytosolic fluorescence intensities were quantified as the mean fluorescence intensity within manually defined nuclear and cytosolic regions of interest, respectively, and background fluorescence was confirmed to be negligible (<0.1% of the measured signal).
Immunofluorescence Assay
HEK293T and HeLa cells were cultured in 8-well chamber slides. The following day, cells were fixed with 4% paraformaldehyde (4% PFA) for 15 min at 20 ± 2C. After washing with PBS, cells were permeabilized with 1% TritonX-100 in PBS for 20 min and blocked with 5% normal goat serum in PBS for 30 min at 20 ± 2C. Cells were incubated with anti-PTEN antibody (#9559, Cell Signaling Technology, Danvers, MA, USA) overnight at 4C. After washing with PBS, cells were incubated with an Alexa488-conjugated secondary anti-rabbit IgG antibody (#111-545-045, Jackson ImmunoResearch, West Grove, PA, USA) for 1 hr at room temperature. Nuclear DNA was subsequently stained with DAPI (#D212, Dojindo, Kumamoto, Japan). Samples were observed using a confocal laser-scanning microscope and quantified as described for live cell imaging.
Results and Discussion
As an initial step, the subcellular localization of endogenous PTEN was characterized before evaluating the effects of FP tagging. Although we previously reported the localization of endogenous PTEN in primary hepatic cells and DU145 prostate cancer cells, 7 in this study, we examined endogenous PTEN localization in HEK293T cells by immunofluorescence analysis. Endogenous PTEN was detected throughout the cell; however, its concentration in the nucleus was approximately 2-fold higher than that in the cytoplasm in HEK293T cells (Fig. 1A and B). We next investigated the subcellular localizations of mGFP, mYFP, mApple, mScarlet3, and mCherry in HEK293T cells. Among the FPs examined, mCherry displayed a modest but consistent enrichment in the cytosol relative to the nucleus compared with the other FPs (Fig. 1C and D), indicating a preferential cytosolic localization. Based on these considerations, we next examined the subcellular localization of PTEN fused to each of the five FPs in HEK293T cells to determine whether FP fusion could modulate PTEN nuclear translocation. Compared with other PTEN-FPs, PTEN-mCherry predominantly accumulated in the cytosol compared with the nucleus (Fig. 1C). Quantitative analysis of nuclear PTEN-mCherry signal intensity relative to cytosolic signal revealed a significant reduction compared with the other constructs (Fig. 1E). Although mCherry also exhibited a cytosolic preference, PTEN-mCherry demonstrated a more pronounced cytosolic accumulation.
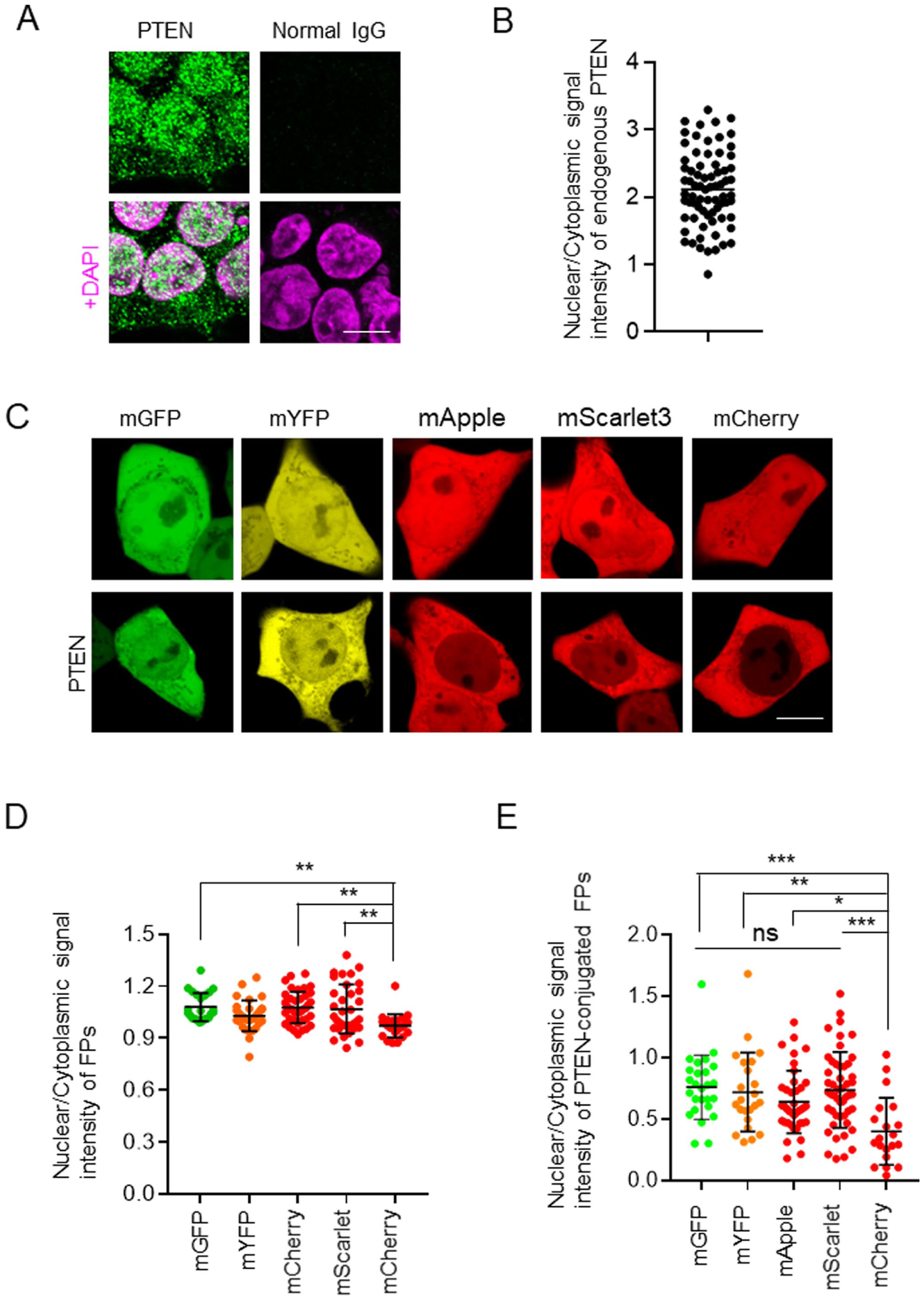
Effects of fluorescent proteins (FP) on the nuclear localization of PTEN in HEK293T cells. (A) Representative confocal images of immunofluorescence staining for PTEN in HEK293T cells. PTEN staining and the corresponding merged images with DAPI are shown. (B) Quantification of the signal intensity of PTEN in the nucleus versus cytosol of HEK293T cells (n=73). (C) Representative confocal images of HEK293T cells expressing mGFP, mYFP, mApple, mScarlet3, and mCherry (FPs) or PTEN fused with these FPs (PTEN-FPs). (D and E) Quantification of the signal intensity of each FP in the nucleus relative to the cytosol (D) and PTEN-FPs (E) in HEK293T cells. Data are presented as mean ± SD. (D: n=23–39, E: n=20–49). Scale bars, 10 µm (A and C). Statistical analysis was performed using one-way ANOVA followed by Tukey’s post hoc test. *p<0.05; **p<0.01; ***p<0.001; ns, not significant.
Furthermore, we examined the effects of FP on the subcellular localization of PTEN in HeLa cells. Consistent with the observations in HEK293T cells, endogenous PTEN exhibited nuclear accumulation in HeLa cells, with an approximately 1.5-fold higher signal intensity in the nucleus compared with the cytosol (Fig. 2A and B). Similarly, in HeLa cells, all fluorescent proteins examined were distributed throughout the cell (Fig. 2C), and no statistically significant differences were observed among them (Fig. 2D). However, PTEN-mCherry showed significant localization in the cytosol of HeLa cells (Fig. 2E). These observations support the modulation of the subcellular localization of PTEN by the localization characteristics of the fused FPs. Notably, all FPs-tagged PTEN constructs used in this study exhibited reduced nuclear accumulation compared with endogenous PTEN in both HEK293T and HeLa cells. These findings suggest that the molecular size of FP may influence nucleocytoplasmic transport. We further investigated the effects of FP size, focusing on its impact on transport across the nuclear membrane. PTEN comprises several domains, including the PIP2 binding, phosphatase, lipid-binding C2, C-tail, and the PDZ-binding domains. 14 In this study, the PDZ-binding domain was included in the C-tail domain. PTEN350, which spans from the N terminus to the C2 domain, represents the core of PTEN and is predominantly localized to the nucleus. 15 Notably, co-expression of the core and C-tail domains results in the cytosolic localization of PTEN, indicating that the C-tail domain inhibits nuclear transport. 15 In this study, PTEN and PTEN350 were expressed in fusion with mGFP and mCherry to investigate the effect of FP size on subcellular localization (Fig. 3A). PTEN350-mCherry was localized to the nucleus in HEK293T cells (Fig. 3B), and no significant differences were observed between PTEN350-mGFP and PTEN350-mCherry (data not shown). While PTENA4-mCherry also exhibited nuclear accumulation, the extent was much lower than that of PTEN350-mCherry (Fig. 3B and C). The effect of the C-tailA4 domain differs from that of the wild-type C-tail domain, as the wild-type C-tail domain completely inhibits the nuclear transport, according to the localization of PTEN-mCherry (Fig. 3B and C). Liu et al. 16 previously reported that the molecular sizes of PTEN and FP could inhibit nuclear transport. In this study, the fusion of two GFP proteins appeared to impede the PTEN diffusion in HeLa cells due to the increased molecular size. However, whether the presence of FPs interfered with the nuclear translocation of PTEN350 remained unclear. To examine this, we generated a PTEN350 construct fused to both mGFP and mCherry (PTEN350-mGFP-mCherry) (Fig. 3A). Notably, PTEN350-mGFP-mCherry exhibited nuclear accumulation; however, this extent was lower than that observed for PTEN350-mCherry (Fig. 3B and C). The increased molecular size of the mGFP-mCherry tag attenuated the nuclear transport of PTEN350. Therefore, when considering the subcellular localization of target proteins, the potential effect of the molecular size of the FP should be carefully considered. Further studies should quantitatively characterize nucleocytoplasmic transport kinetics, such as the rate of nuclear membrane translocation, using photobleaching experiments, thereby providing a more precise understanding of how FP size affects PTEN localization.
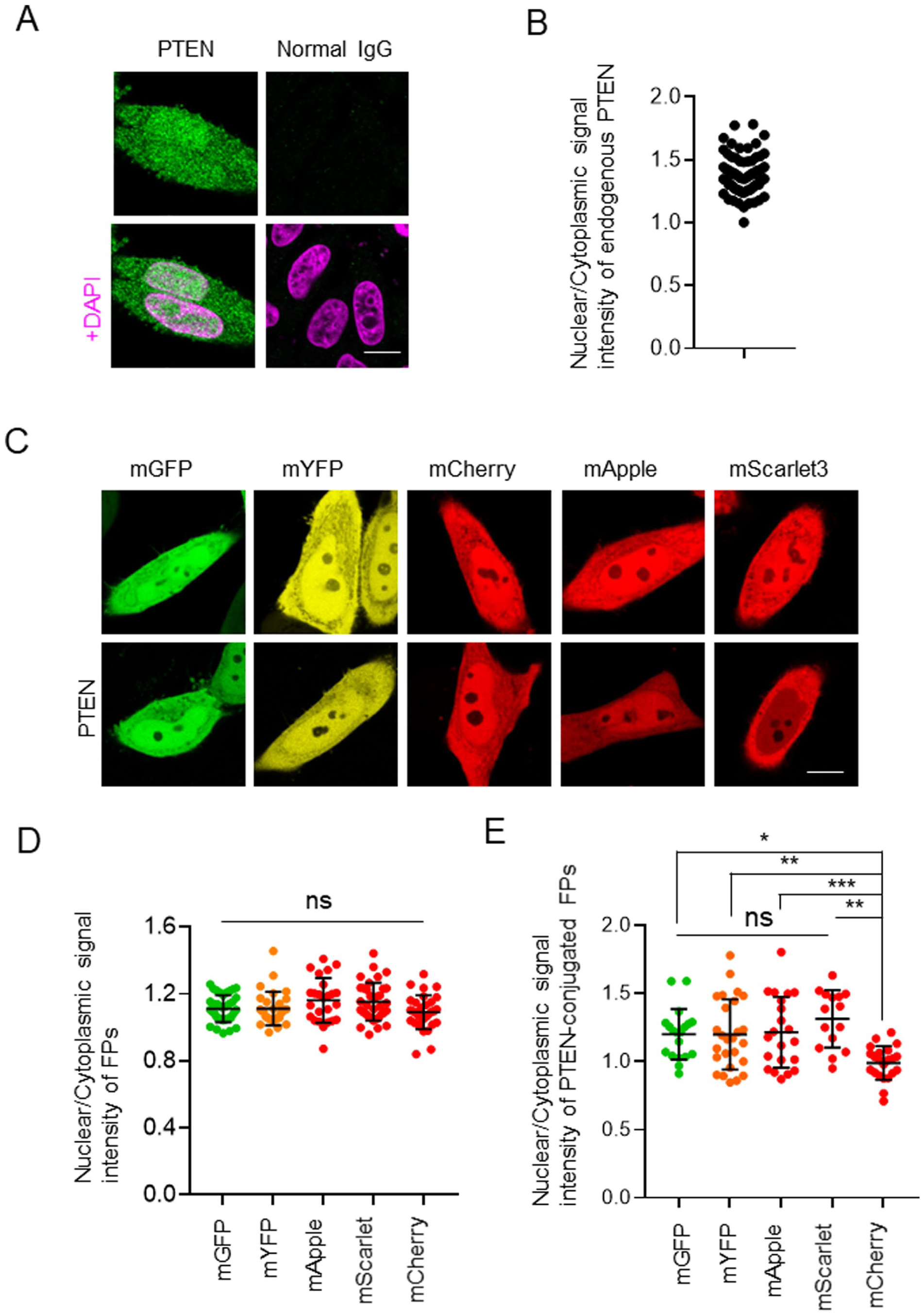
Effects of fluorescent proteins on the nuclear localization of PTEN in HeLa cells. (A) Representative confocal images of immunofluorescence staining for PTEN in HeLa cells. PTEN staining and the corresponding merged images with DAPI are shown. (B) Quantification of the signal intensity of PTEN in the nucleus relative to the cytosol of HeLa cells (n=71). (C) Representative confocal images of HeLa cells expressing mGFP, mYFP, mApple, mScarlet3, and mCherry (FPs) or PTEN fused with these FPs (PTEN-FPs). (D and E) Quantification of the signal intensity of each FP in the nucleus relative to the cytosol (D) and PTEN-FPs (E) in HeLa cells. Data are presented as mean ± SD (D: n=25–37, E: n=15–26). Scale bars, 10 µm (A and C). Statistical analysis was performed using one-way ANOVA followed by Tukey’s post hoc test. *p<0.05; **p<0.01; ***p<0.001; ns, not significant.
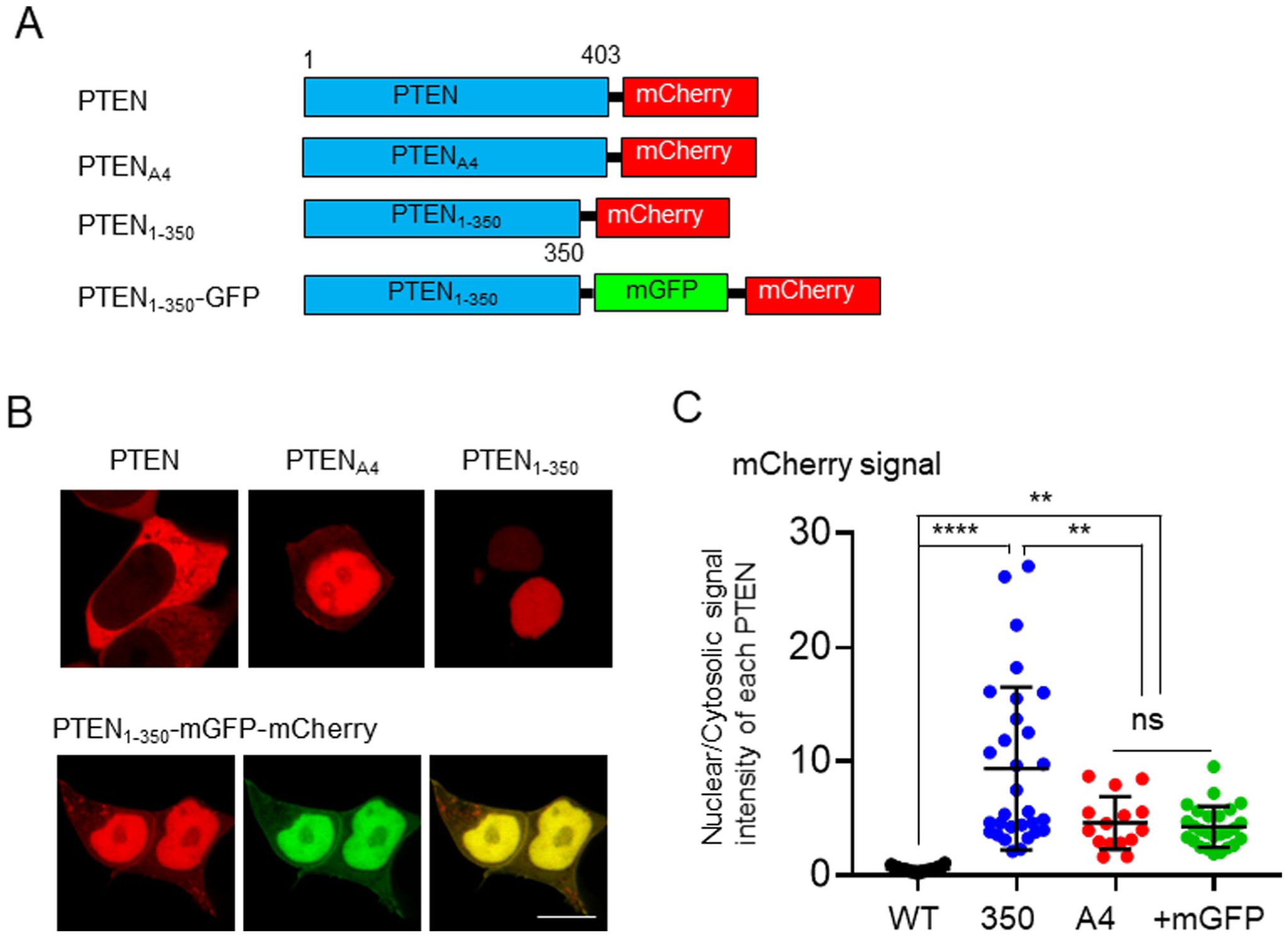
Effect of molecular size of fluorescent proteins on the subcellular localization of PTEN constructs. (A) Schematic diagrams of PTEN constructs. (B) Representative confocal images of HEK293T cells expressing mCherry-fused PTEN, PTENA4, PTEN350, and PTEN350-mGFP. Scale bars, 10 µm. (C) Quantification of the signal intensity of mCherry fused with PTEN, PTENA4, PTEN350, and PTEN350-mGFP in the nucleus relative to the cytosol of HEK293T cells (n=15–38). Statistical analysis was performed using one-way ANOVA followed by Tukey’s post hoc test. **p<0.01; ***p<0.001; ****p<0.0001; ns, not significant.
Notably, we mainly utilized the N1 fluorescent vectors in this study, including PTEN-mGFP and PTEN-mCherry, but there was a significant difference between mGFP-PTEN and mCherry-PTEN, which were generated from the C1 fluorescent vectors (Fig. 4). The effects of the location of the FP tag on the PTEN subcellular localization were not determined.
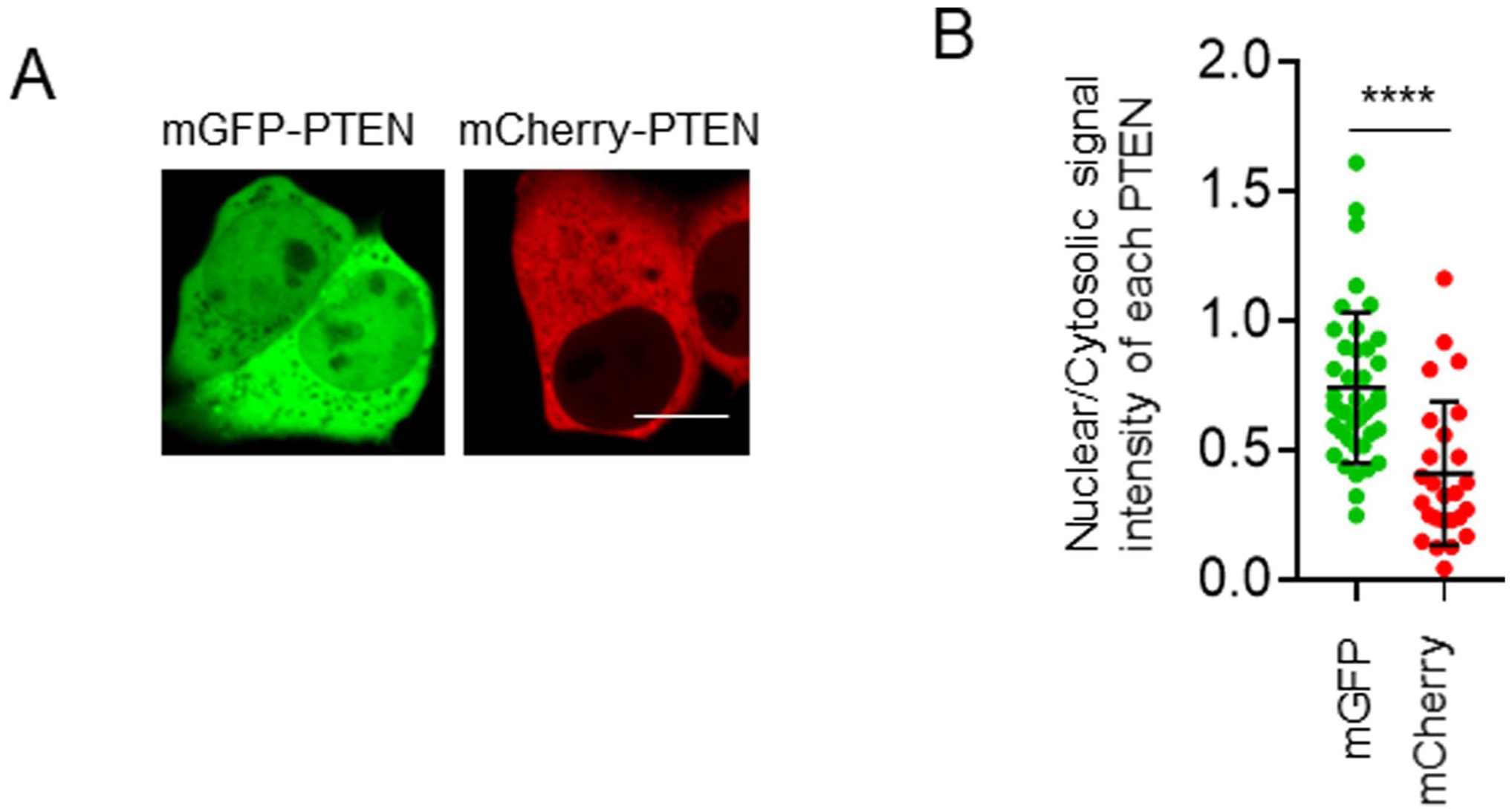
Subcellular localization of PTEN with C-terminal fluorescent protein tags. (A) Representative confocal images of HEK293T cells expressing mGFP-PTEN and mCherry-PTEN (hereafter referred to as FPs-PTEN). Scale bars, 10 µm. (B) Quantification of the signal intensity of FP-PTEN in the nucleus relative to the cytosol (n=26–41). Statistical analysis was performed using the Student’s t-test. ****p<0.0001.
FP-conjugated PTEN mutants displayed marked differences in their subcellular distribution. We first focused on PTENK13R exhibiting cytoplasmic accumulation and utilized mGFP, mYFP, and mCherry as FP tags. PTENK13R-mGFP, PTENK13R-mYFP, and PTENK13R-mCherry were localized predominantly in the cytosol of HEK293T cells (Fig. 5A). However, no significant differences were observed among the three constructs (Fig. 5B). To determine the effects of FPs on the nuclear transport of PTENA4, we also examined the nuclear accumulation of PTENA4-mGFP, PTENA4-mYFP, and PTENA4-mCherry in HEK293T cells (Fig. 5A). Quantitative analysis revealed no significant differences in the nuclear signal intensity relative to that in the cytosol among the three groups (Fig. 5C). Thus, FPs did not alter the subcellular localization of PTENK13R and PTENA4, suggesting that the effects of these mutations override the intrinsic localization tendencies of the FPs. We further investigated whether conjugation with an FP affected the plasma membrane localization of PTEN. PTENK13R,A4, a double mutant harboring K13R and A4 substitution, is known to accumulate at the plasma membrane.8,11 In HEK293T cells, all three FP-conjugated PTENK13R,A4 were predominantly localized to the plasma membrane (Fig. 5D), and no significant differences were detected among the constructs.
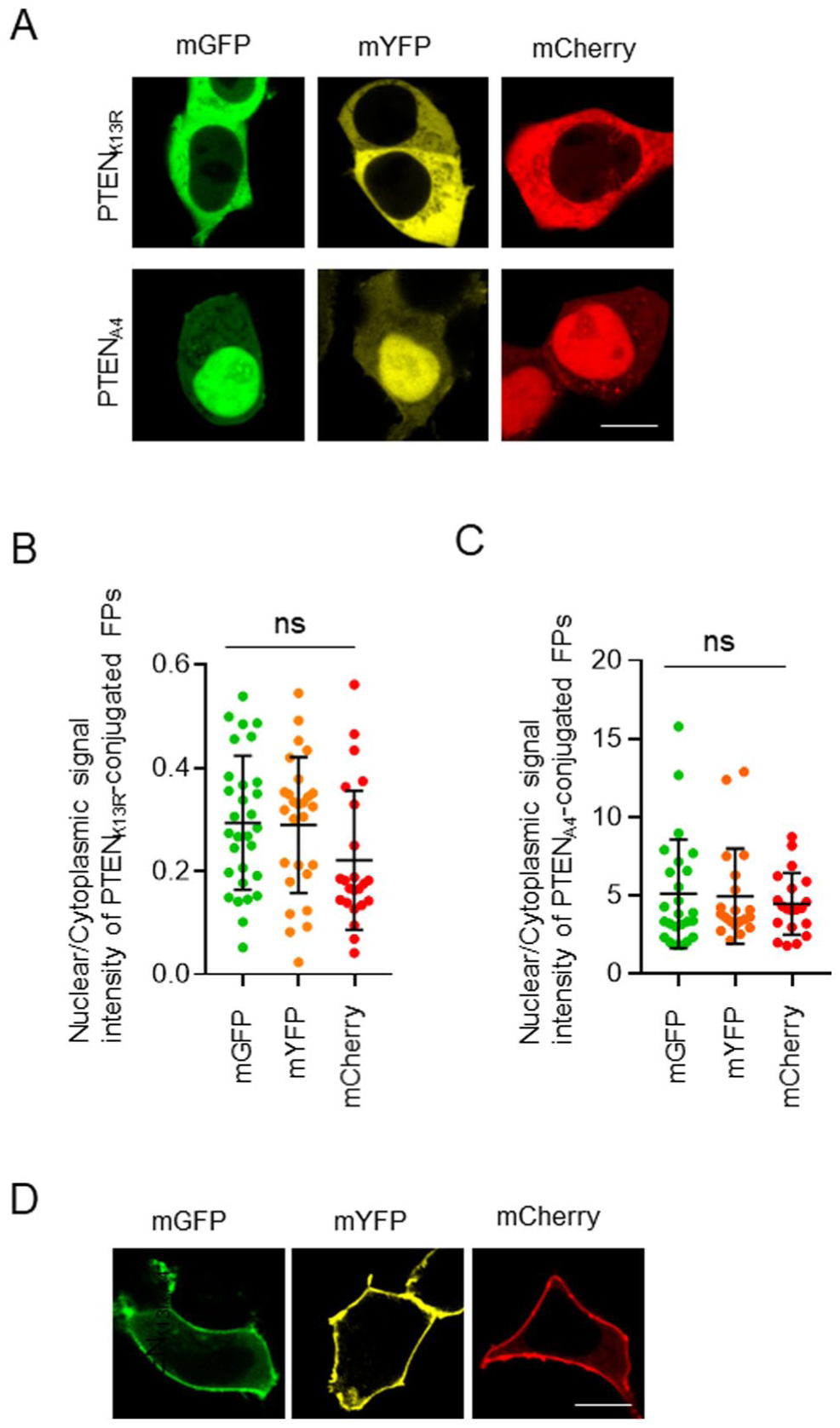
Effects of the fluorescent proteins on the nuclear translocation of PTEN mutants. (A) Representative confocal images of HEK293T cells expressing FP-fused PTENK13R and PTENA4. (B and C) Quantification of the signal intensity of PTENK13R-FPs (B) and PTENA4-FPs (C) in the nucleus relative to the cytosol of HEK293T cells (PTENK13R, n=23–41, PTENA4, n=20–25). (D) Representative confocal images of HEK293T cells expressing PTENK13R,A4 with FPs. Scale bars, 10 µm (A and D). Statistical analysis was performed using one-way ANOVA followed by Tukey’s post hoc test; ns, not significant.
Nuclear PTEN exerts a DNA-protective effect under oxidative stress conditions.17,18 Consistently, H2O2, spermine NONOate (SPNO), and camptothecin (CPT) induce the nuclear accumulation of PTEN-GFP. 7 In this study, CPT and etoposide (ETO), which are inhibitors of topoisomerase I and II, respectively, were used as DNA-damaging agents.19 –22 Exposure to these DNA-damaging agents caused nuclear translocation of PTEN-mGFP (Fig. 6A), with the extent of translocation being significantly greater than that observed in DMSO-treated cells (Fig. 6B). Similarly, PTEN-mCherry exhibited nuclear translocation under DNA damage conditions (Fig. 6C). Notably, significant differences were also observed between DNA-damaged cells and DMSO-treated controls (Fig. 6D). These results suggest that mCherry can serve as a reliable marker for analyzing the nuclear translocation of PTEN induced by DNA damage.
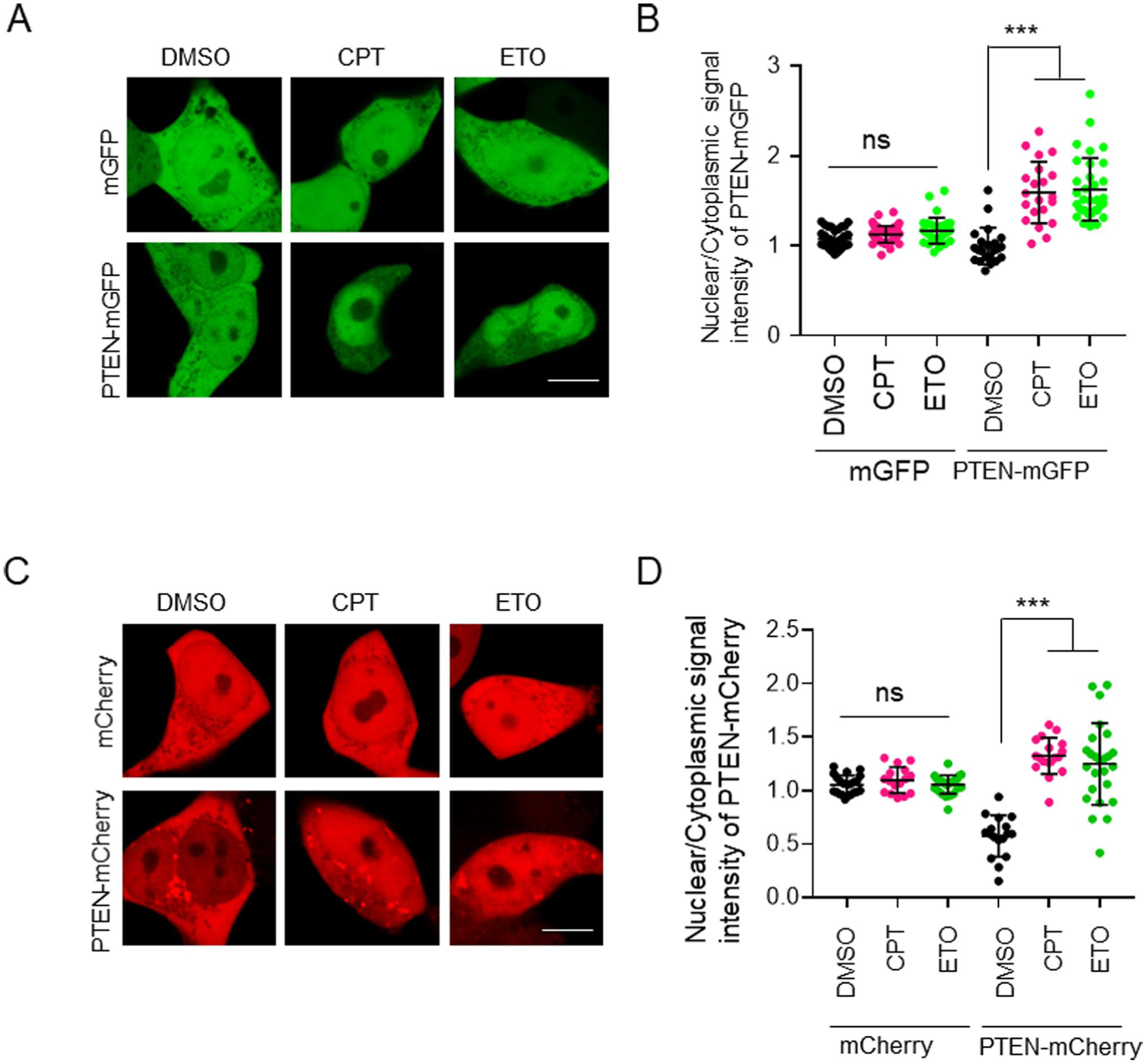
The nuclear translocation of PTEN in response to DNA damage was not affected by the type of FP used as a tag. (A) HEK293T cells expressing mGFP and PTEN-mGFP were treated with two topoisomerase inhibitors, camptothecin (CPT; 50 µM) and etoposide (ETO; 1 mM) for 8 hr. (B) Quantification of the nuclear-to-cytosolic signal intensity ratio of mGFP and PTEN-mGFP in HEK293T cells (n=21–53). (C) HEK293T cells expressing mCherry and PTEN-mCherry were treated with CPT (50 µM) and ETO (1 mM) for 8 hr. (D) Quantification of the nuclear-to-cytosolic signal intensity ratio of mCherry and PTEN-mCherry in HEK293T cells (n=16–25). Scale bars, 10 µm. Statistical analysis was performed using one-way ANOVA followed by Tukey’s post hoc test. ***p<0.001; ns, not significant.
Dynamic changes in subcellular localization are critically important for the execution of diverse cellular functions. For example, under oxidative stress conditions, such as H2O2 treatment, PTEN is rapidly redistributed from the cytoplasm to the nucleus, where it exerts its DNA-protective function. This redistribution occurs within approximately 1 hr and is not accompanied by any detectable change in the total cellular expression level of PTEN. 7 These observations indicate that nucleocytoplasmic transport of PTEN is pivotal in regulating its function, underscoring the necessity for careful and quantitative evaluation of PTEN subcellular localization. Accordingly, when analyzing PTEN localization using FP tags, rigorous assessment of the potential impact of FPs on PTEN subcellular distribution is merited. This issue is not unique to PTEN but is likely applicable to a broad range of proteins; therefore, we consider it important to document these findings. We previously demonstrated that loss of nuclear PTEN enhanced DNA damage in hepatocytes and promoted the development of liver cancer. 7 Consistently, Nguyen et al. 10 reported that hydroxyurea (HU) treatment induced DNA damage in PTEN-null HCT116 cells. Importantly, exogenous expression of PTEN suppresses HU-induced DNA damage, and nuclear-specific PTENA4 confers particularly robust protection against HU-mediated genotoxic stress. The DNA-protective function of nuclear PTEN is determined by the absolute amount of PTEN in the nucleus relative to its cytoplasmic abundance. Therefore, future studies should focus on elucidating how functional outcomes correlate with absolute protein levels in distinct subcellular compartments.
Since the discovery of GFP, several FP tags have been developed and widely utilized as essential tools for studying subcellular localization. Thus, a thorough understanding of the unique properties of these FPs is critical. In wild-type PTEN under DNA damage conditions, as well as in PTEN mutants, the subcellular localization was largely unaffected, and no significant differences attributable to FP type were observed. Nevertheless, in this study, FPs were found to influence the nuclear translocation of PTEN and its fragments. In conclusion, potential biases arising from the inherent characteristics and molecular sizes of the FP tags should be considered when interpreting experimental data.
Footnotes
Competing Interests
The authors declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Author Contributions
TK had overall responsibility for the study, including conception and design, experimental performance, data analysis and interpretation, and manuscript preparation. YK participated in the study design, experimental performance, and data analysis and interpretation.
Funding
The authors disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This work was supported by Yasuda Women’s University, Scientific Research Aid.
Ethics Approval and Consent to Participate
Not applicable.
Patient Consent for Publication
Not applicable.
Availability of Data and Materials
The data generated in the present study may be requested from the author.