
Abstract
Pediatric sarcomas comprise approximately 10% of all childhood solid malignancies and are characterized by distinct genetic and proteomic alterations that have potential diagnostic, prognostic, and therapeutic significance. We have utilized digital spatial profiling (DSP) to identify protein expression in pediatric Ewing sarcoma (ES), Osteosarcoma (OS), Alveolar rhabdomyosarcoma (ARMS), and Embryonal rhabdomyosarcoma (ERMS), in association with clinical outcomes. Formalin-fixed, paraffin-embedded sections from a tissue microarray block containing eight ES, eight OS, five ARMS, and three ERMS cases were subjected to proteomic DSP on a GeoMx NanoString platform yielding information on expression of 580 proteins. Proteins related to epigenetic regulation, signaling pathways, and mesenchymal differentiation were broadly expressed across all tumor types. Tumor-specific protein profiles were defined based on highly expressed proteins. Differentially expressed proteins include Cyclin D1 in ES, S100A4 in OS and IKKi/IKKe in ARMS and ERMS. Immunohistochemical validation confirmed variable expression of H3K27me3 across the tumors, and elevated expression of Cyclin D1 in ES and S100 in OS. These findings underscore the utility of DSP as a high-resolution proteomic tool for the identification of clinically relevant biomarkers in pediatric sarcomas. The results provide a foundation for further investigation of candidate proteins with potential diagnostic, prognostic, and therapeutic applications:
Keywords
Introduction
Pediatric sarcomas comprise a heterogeneous group of mesenchymal malignancies arising in bone and/or soft tissue. 1 These aggressive tumors account for approximately 10% of childhood solid tumors. The most common bone sarcomas are osteosarcoma (OS) and Ewing (or Ewing’s) sarcoma (ES), whereas rhabdomyosarcoma (RMS) represents the most prevalent soft tissue sarcoma in the pediatric population. Despite distinct origins, these tumors frequently demonstrate overlapping clinical and radiologic features (e.g., OS and ES) or similar histopathologic characteristics, such as undifferentiated small round cell morphology, complicating accurate diagnosis in tissue specimens. Pediatric sarcomas exhibit distinct genetic and molecular profiles, leading to substantial heterogeneity in tumor biology, clinical behavior, and therapeutic response. Tumorigenesis is primarily driven by genetic and epigenetic alterations, including chromosomal translocations, gene fusions, and secondary oncogenic mutations, which lead to dysregulated expression of transcription factors and oncoproteins. These molecular alterations, influenced by both intrinsic and extrinsic factors, promote activation of receptor tyrosine kinases and downstream intracellular signaling cascades, notably the RAS–MAPK and PI3K–AKT pathways, which are commonly dysregulated across human malignancies and contribute to cellular transformation, proliferation, and tumor progression. 2
OS is characterized by aberrant expression of multiple proteins including growth factors, heat shock proteins, Ezrin, Runx2, SATB2, ATF4, and Rb1, which regulate key processes in carcinogenesis such as cell adhesion, migration, invasion, control of cell cycle, and apoptosis. 3 However, ES is defined by EWS-ETS chimeric gene fusion which drives widespread genomic and proteomic reprogramming resulting in the overexpression of 210 phosphopeptides such as IL6, STAT3, and STEAP1, many of which represent potential therapeutic targets.4,5 Pediatric RMS is broadly classified into embryonal (ERMS) or alveolar (ARMS) subtypes based on histomorphology and presence of FOXO1 gene rearrangements. Integrated genomic and proteomic analysis has identified frequent alterations in the RAS pathway in RMS resulting in activation of myogenic transcription factors and other oncogenic drivers.6,7 Although these tumors are generally aggressive, clinical outcomes vary considerably depending on factors such as tumor site, histologic subtype, genetic background, and metastatic status. 1
The overlapping morphologic and clinical features of these sarcomas often necessitate extensive immunohistochemical evaluation to establish an accurate diagnosis. Identification of additional protein biomarkers may further improve diagnostic precision, enable prognostic stratification, and inform therapeutic decision-making, including the development of targeted treatment strategies. 8 In this study, we employed digital spatial profiling (DSP) technology to characterize protein expression patterns in tumor tissues from ES, OS, ARMS, and ERMS. The GeoMx DSP platform is a recently validated, high-plex proteomic technology designed for immuno-oncology applications, enabling spatially resolved protein analysis in formalin-fixed, paraffin-embedded (FFPE) tissue sections.9,10 We have identified and analyzed highly expressed proteins in each tumor and validated selected findings with conventional immunohistochemistry (IHC).
Materials and Methods
Patients and Materials
A tissue microarray (TMA) block was constructed from archived pathology tissue materials of eight ES tumor cases, eight OS from seven patients, five ARMS from four patients and three ERMS cases collected between 2005 and 2019 from pediatric and young adult patients. Three-millimeter tissue cores extracted from the original blocks contained representative viable tissue of the selected cases. Tumor diagnoses were confirmed by re-review of the histology, characteristic immunophenotype and molecular tests (Table 1). All ARMS and most ES cases (7/8) were fusion positive with rearrangements of FOXO1 and EWSR1 genes, respectively, identified by fluorescent in situ hybridization (FISH) or RNA sequencing. Molecular sequencing results were obtained from some cases (Table 1). The OS samples were from high grade conventional tumors with two cases demonstrating moderate stromal lymphocytic inflammatory infiltrate. All cases were sampled from primary tumor locations except for one OS case that was from a lung metastasis. Two OS cases required decalcification because of included bone and cartilage. Two patients had two samples each, one from a primary diagnostic specimen and another from a post-chemotherapy resection or metastatic lung focus. Patients received standard treatment protocols with surgery and chemotherapy. Follow-up information was available on all patients enabling classification into two prognostic groups (good versus adverse outcome) according to the length of the overall survival (from the time of diagnosis until death or loss of follow-up) and status (whether alive or deceased) at end of follow-up.
Patients and Tumors Included in the Digital Spatial Protein Profiling Study.
Abbreviations: pt, patient; pts, patients.
GeoMx DSP
A TMA section was subjected to antigen retrieval and subsequently processed for GeoMx IO Proteome Atlas (IPA) collections according to NanoString protocols. The IPA panel contains probes for 580 protein targets. For this assay, the slide was stained with Syto13 nuclear DNA, pan-cytokeratin (AE1/AE3, Novus; Centennial, CO) for epithelium and CD45 (D9M8I, Cell Signaling Technology; Boston, MA) antibodies by immunofluorescence to identify tumor and stromal immune cells, respectively. The slide was scanned on the GeoMx instrument (NanoString; Seattle, WA) and two to three 500-micrometer-diameter circular regions of interest (ROIs) were placed on the tumor cores corresponding to areas with the highest viable tumor density (with more than 70% tumor nuclei) and least burden of stromal and inflammatory cells. After review of tumor tissue quality and in correlation with immunofluorescence staining, ROIs were discarded in one OS, one ES, and one ARMS cases. Remaining ROIs were further segmented into CD45-negative and CD45-positive compartments based on expression of each respective marker. DSP barcoded antibody probes were collected for each segment and sequenced on the Illumina NovaSeq X Plus platform as previously described. 10 Expression levels from each segment were recorded in the form of discrete measurements or counts that are normalized to house keeping proteins.
Selected targets screened included proteins involved in drug resistance, complement system, T-cell checkpoints, senescence, lipid metabolism, various signaling pathways, epithelial mesenchymal transition (EMT), lysosome, Myc, cytotoxicity, glutamine metabolism, T-helper (TH) differentiation, cellular differentiation, interferon response genes, Matrix remodeling and metastasis, lymphocyte regulation, epigenetic regulation, major histocompatibility complex (MHC) class I antigen presentation, immortality and stemness, cell cycle, myeloid inflammation, apoptosis, endocytosis, autophagy, lymphocyte trafficking, DNA damage repair, prostaglandins, MHC class II antigen presentation, T-cell exhaustion, mitochondrial metabolism/TCA, cell adhesion and motility, Treg differentiation, proteotoxic stress, oxidative stress and IDH1/2. Target antibodies have been previously validated by IHC with a single plex Abcam method as well as multiplex immunofluorescence-based TMA methods. Expression data was analyzed in the GeoMx Analysis Suite. 11 Statistical tests for significant differences were calculated with fold changes and p values that were adjusted with the Benjamini–Hochberg method.
Immunohistochemical Validation
Based on the DSP results, TMA slides were stained by IHC for S100, Cyclin D1 and the histone trimethylated protein H3K27me3. After antigen retrieval, automated IHC was performed on Ventana Benchmark Ultra instrument (Indianapolis, IN), in a CLIA-certified clinical histology lab workflow. Standard protocols were followed including the use of positive controls.
Results
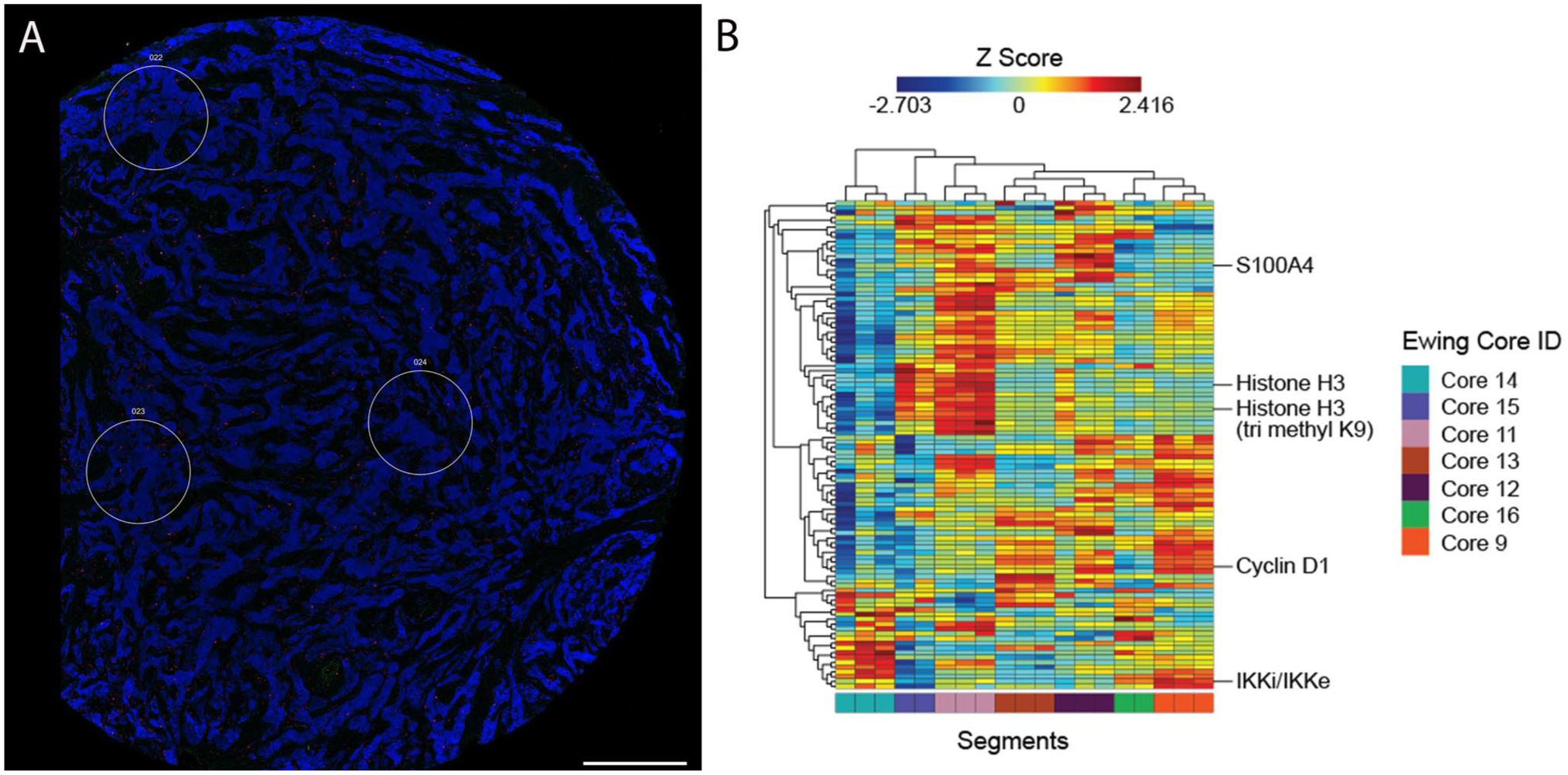
Quality analysis of the samples revealed adequate hybridization with no quality control flags or abnormalities detected including the OS samples previously treated with decalcification. Read counts were normalized to the geometric mean expression of several housekeeping genes, thereby mitigating the effect of variability of analyte abundances or quality across our regions of interest (Fig. 1). A distinct protein expression profile was elucidated in each tumor, and the most highly expressed proteins were characterized (Supplementary Tables 1–4). Mesenchymal differentiation markers, epigenetic modifiers, and signaling pathways were consistently identified among the top 20 expressed proteins. These included vimentin, alpha smooth muscle actin, the histone proteins H3, H2A, and histone deacetylase (HDAC) isoforms, the signaling pathway proteins, MEK2 and NF-κB, the apoptotic protein Smac/Diablo, and the apoptosis-related proteins, calreticulin and GAPDH (Fig. 2). Furthermore, certain proteins exhibited unique expression patterns specific to particular tumor types. High expression of Caveolin and IGF2BP1 was markedly associated with ES. HLA-A expression was observed in OS and ES while HMGB1 was identified in ERMS and ARMS. Both ES and ARMS demonstrated significant expression of the mitochondrial protein Tomm20 (Fig. 2A–D).
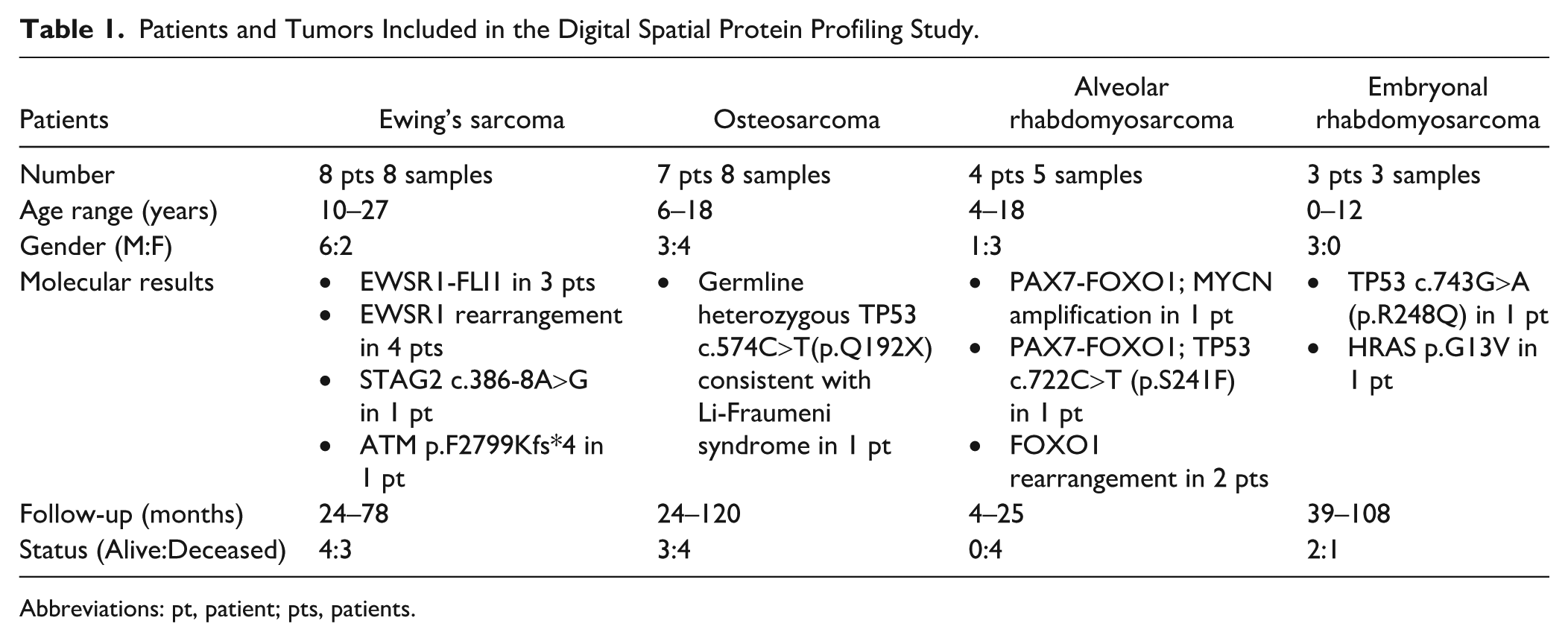
Immunofluorescence-based selection of ROI and DSP analysis of Ewing sarcoma tumors. Section reveals abundant tumor cells (labeled blue) with scant stroma (red). Target protein probes bound to the tissue were collected from each ROI (A). Probes were sequenced and read counts were normalized to generate target expression profiles. Scale bar = 0.5 mm. (B) Heat map of the top 100 proteins expressed in Ewing sarcoma demonstrates heterogeneity between segments and cores. Values within the figure are log-transformed and z-scored; color represents z-score. Unsupervised clustering on the x and y axes shows clustering of segments from the same cores. Select proteins are included within the top 100 proteins and highlighted.
After normalization of housekeeping proteins, target protein expression in pediatric sarcoma samples is presented with box plot graphs. Each dot represents the normalized count from each individual segment. The top 20 expressed proteins reveal enrichment in epigenetic modifiers, signaling pathways, apoptosis molecules and mesenchymal differentiation markers in ES (A) OS (B), ARMS (C), and ERMS (D). The y-axis represents signal counts.
Differential Expression
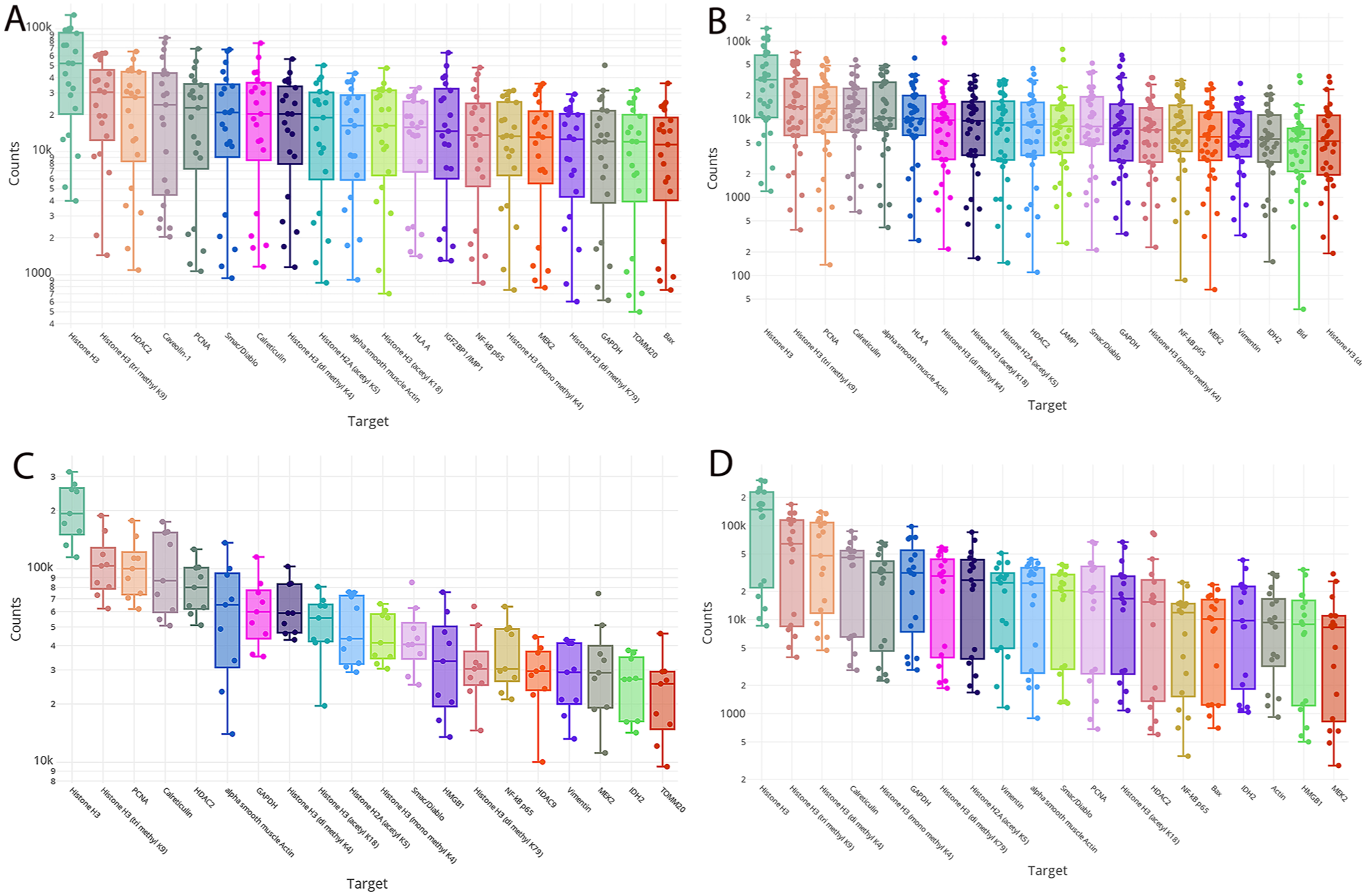
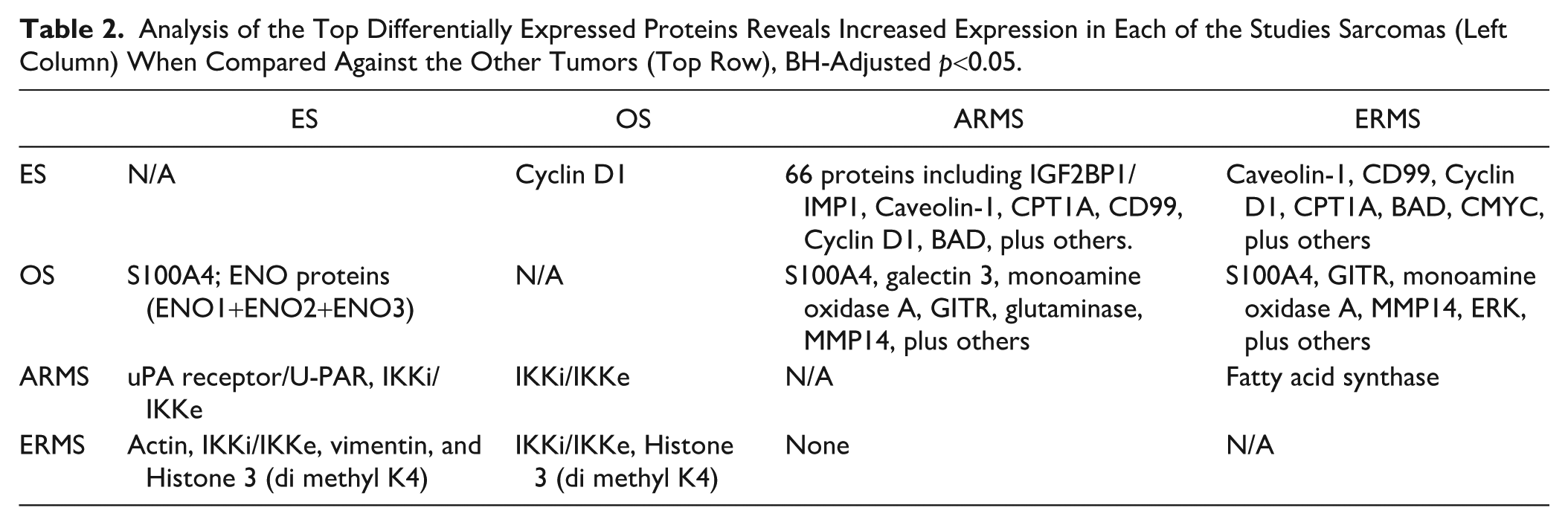
By performing a linear mixed model to calculate the differential protein expression between each of the studied tumors, we identified increased expression of Cyclin D1 in ES, S100A4 in OS and IKKi/IKKe (or IKKi/IKKє) in ERMS and ARMS. Normalized counts for each of these proteins grouped by tumor type highlighted their differential expression (Fig. 3). In comparison to ERMS and ARMS, ES and OS exhibited distinct protein profiles characterized by significant differential expression of more than 50 proteins including cyclin D1, CD99, IGF2BP1/IMP1, Caveolin-1, CPT1A, ERG, and BAD in ES; and S100A4, monoamine oxidase A, GITR, MMP14, and glutaminase in OS (Figs. 4 and 5). Both ERMS and ARMS had increased expression of IKKi/IKKe compared with the other tumors. In contrast, less than five proteins were differentially expressed between OS and ES and between ARMS and ERMS (Fig. 6). Differential expression of fatty acid synthase enzyme was observed in ARMS, compared with ERMS (Table 2).
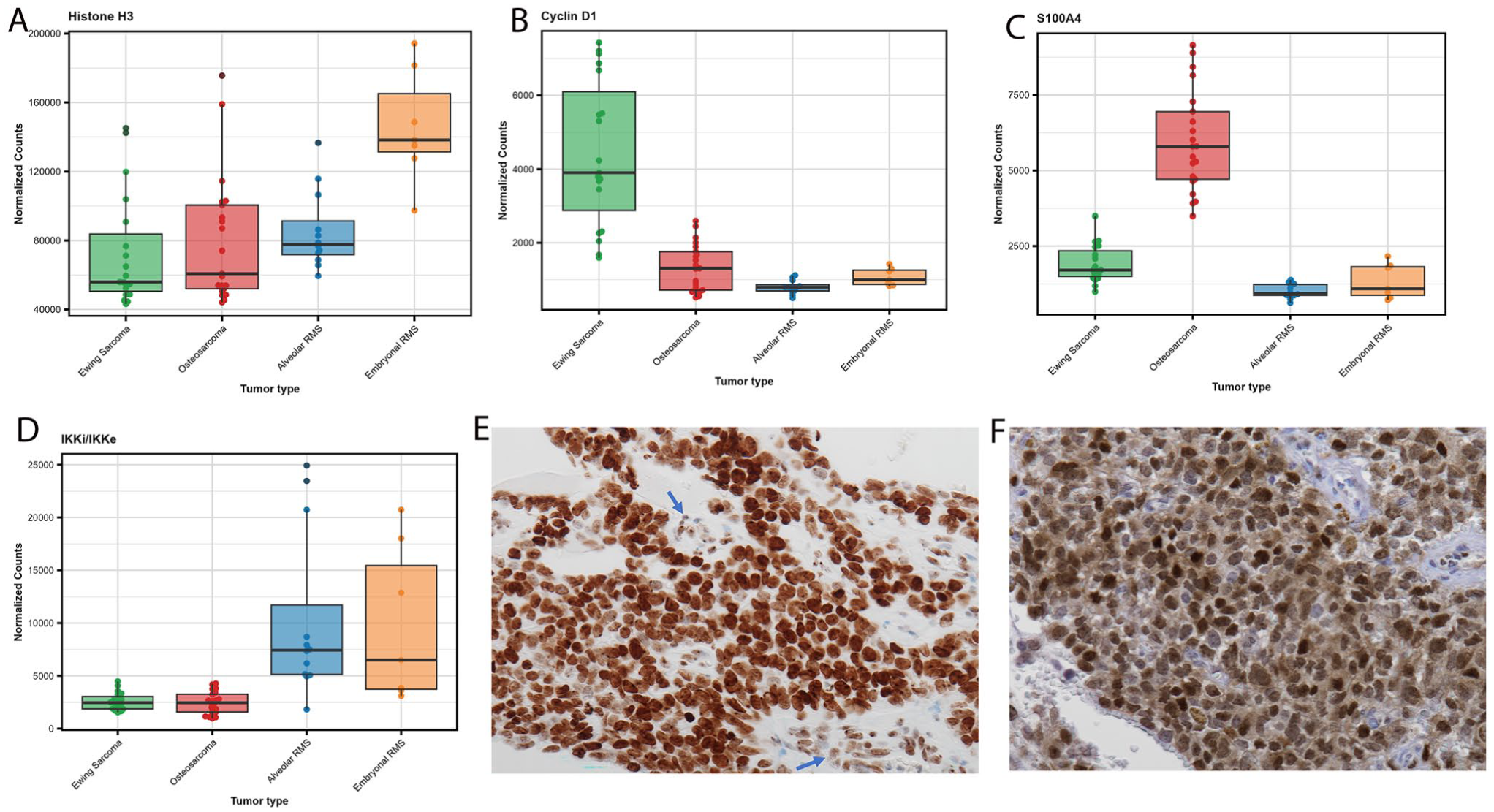
Differentially expressed proteins in the studied samples. A) Box plots reveal universal expression of Histone 3 in all tumors that was slightly higher in ERMS (A). Higher expression of Cyclin D in Ewing sarcoma (B), S100A4 in osteosarcoma (C) and IKKi/IKKe in rhabdomyosarcomas (D) were also illustrated in supporting volcano plots results. The y-axis demonstrates normalized counts as reflection of the degree of expression. (E) Immunohistochemistry for H3K27me3 reveals increased expression in Ewing sarcoma cells with stronger staining intensity compared with the weak staining in the vascular endothelium (arrows). (F) Cyclin D1 immunohistochemical staining was mostly confined to Ewing sarcoma.
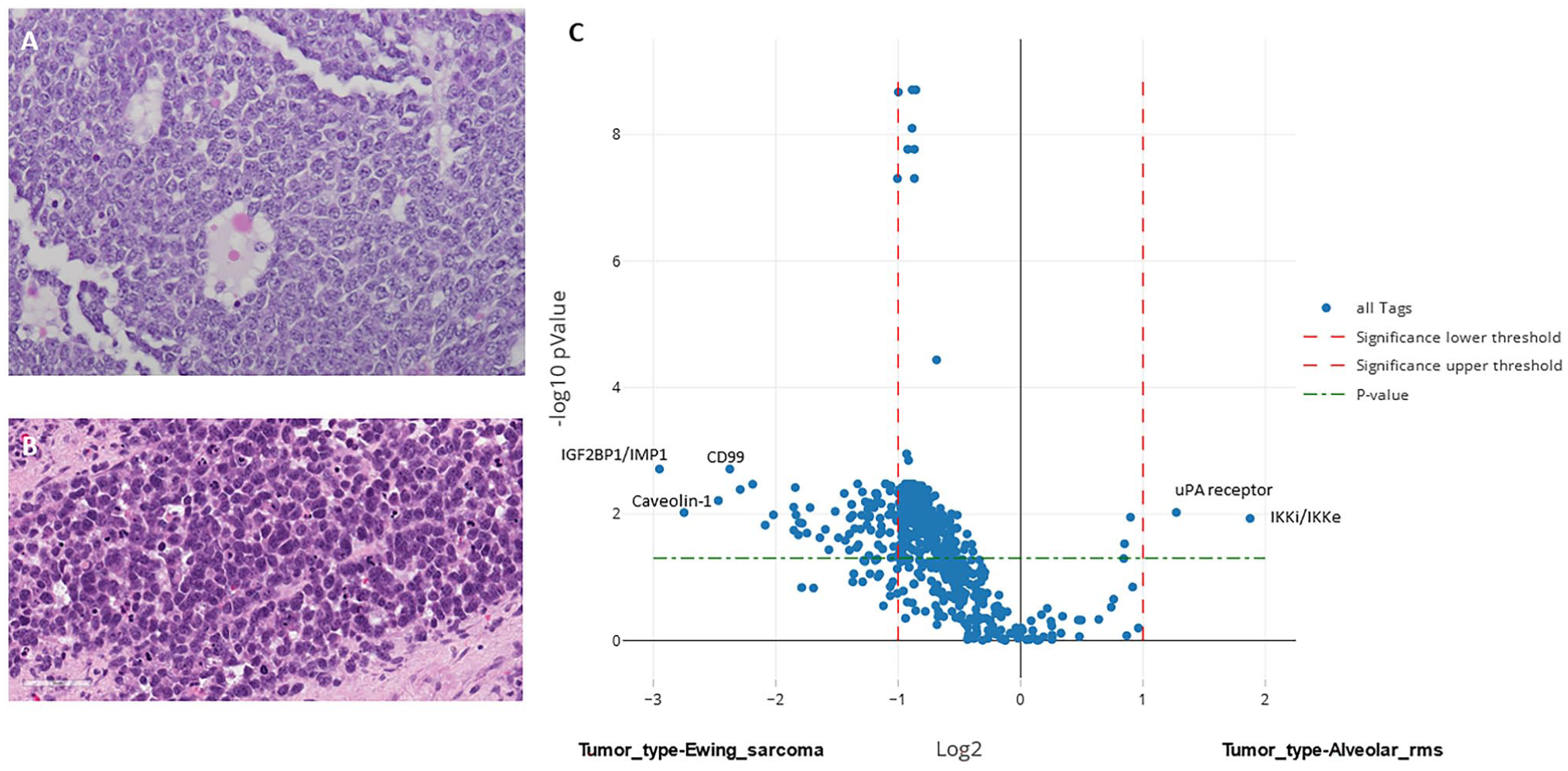
Proteomic differences between ES and ARMS. Despite similar histologic microscopic appearances, ES (A) and ARMS (B), tumors yield distinct proteomic profiles with differential expression of more than 60 proteins (C). The y-axis represents log2 of fold change and the x-axis is −log10 p value.
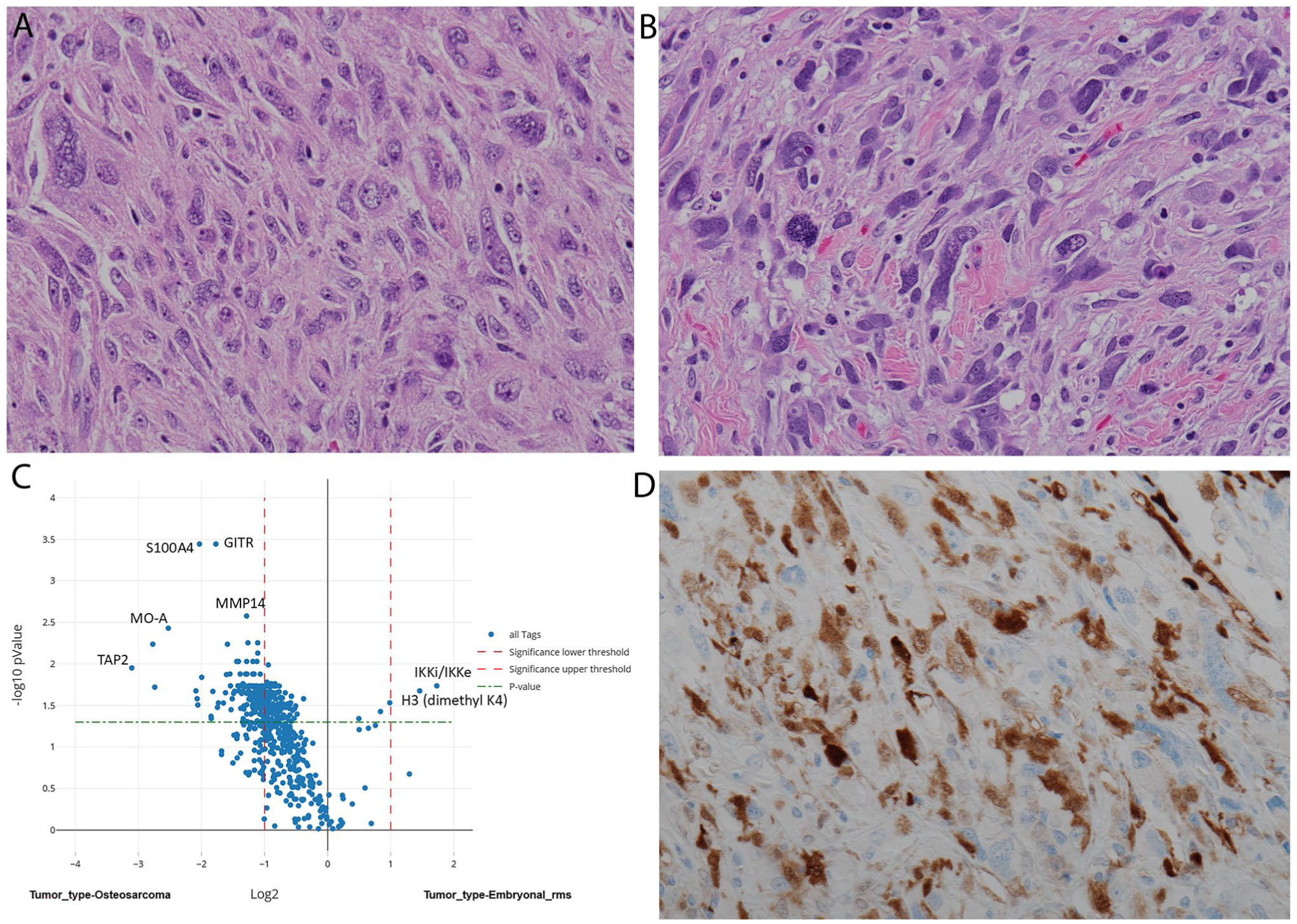
Metastatic osteosarcoma (C; ×400) without significant osteoid differentiation reveal histologic similarities to embryonal rhabdomyosarcoma (D; ×400). Digital spatial profiling has unveiled S100A4 differential expression in osteosarcoma with high statistical significance that was validated by traditional immunohistochemistry for S100 in osteosarcoma.
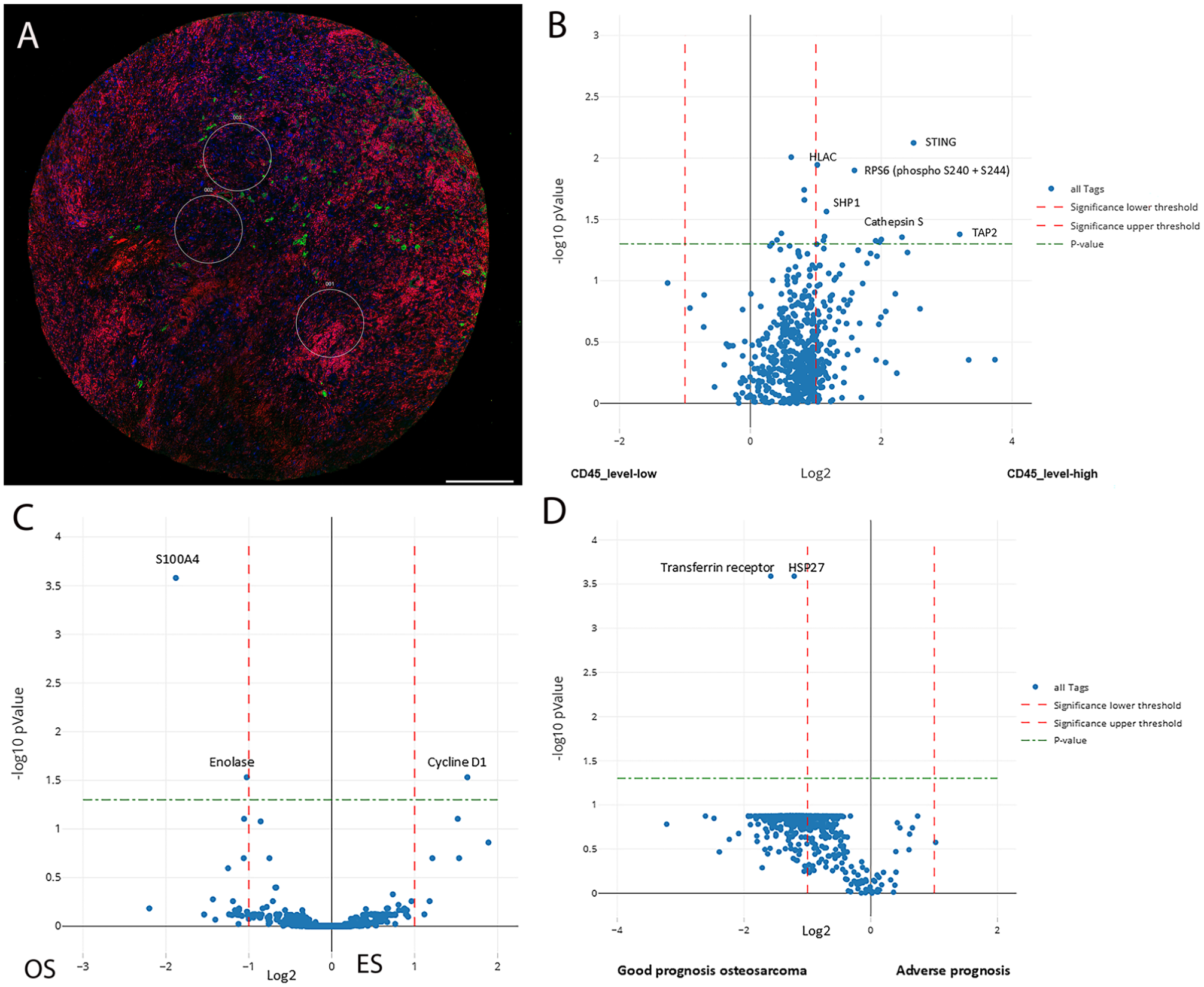
Immunofluorescence-based selection of ROI and DSP analysis in osteosarcoma. (A) Three ROIs were selected in OS (A) based on immunofluorescence with CD45. Tumor nuclei are labeled blue, stromal background as red and CD45-positive cells as green. Scale bar = 0.5 mm. Volcano plots of differentially expressed proteins in OS with respect to the microenvironment display upregulation of several immune-related proteins in OS associated with high CD45-positive stromal cells (B). Similarly, OS versus ES (C), and OS with good prognosis versus worse prognosis tumors (D) reveal differential expression of several proteins of interest. The y-axis represents log2 of fold change and the x-axis is −log10 p value.
Analysis of the Top Differentially Expressed Proteins Reveals Increased Expression in Each of the Studies Sarcomas (Left Column) When Compared Against the Other Tumors (Top Row), BH-Adjusted p<0.05.
OS cases exhibiting stromal inflammatory infiltrate (n=2) presented a unique protein profile characterized by expression of STING, RPS6, SHP1, Cathepsin S, and TAP2, compared with other OS cases with no inflammatory infiltrate (n=6) (Fig. 6). Protein expression within the stromal cells was not assessed. A change in the protein profile was also noted in the single OS case with sequential sampling. The paired OS tumor samples were from an 11-year-old female with a stage IIB OS of the right proximal femur. She was treated with standard chemotherapy followed by surgical resection and remained disease-free for 2 years. Subsequently, a localized recurrence (specimen OS1) was treated with ifosfamide and etoposide. Three months later, the patient underwent limb salvage surgery with hip disarticulation but ultimately succumbed to the disease. The disarticulation specimen (OS2) had viable tumor with less than 50% necrosis. Proteomic differences were identified between OS1 and OS2 with significant expression of SLC7A5/LAT1, uPA receptor, FAK, Dihydrofolate reductase, topoisomerase, C-reactive protein, MEK1, and PP2A in OS2 accompanied by loss of the mesenchymal markers vimentin and alpha-smooth muscle actin (Fig. 7A). There was also loss of expression of collagen I, fibronectin and PDGFR in OS2 with less significant differences.
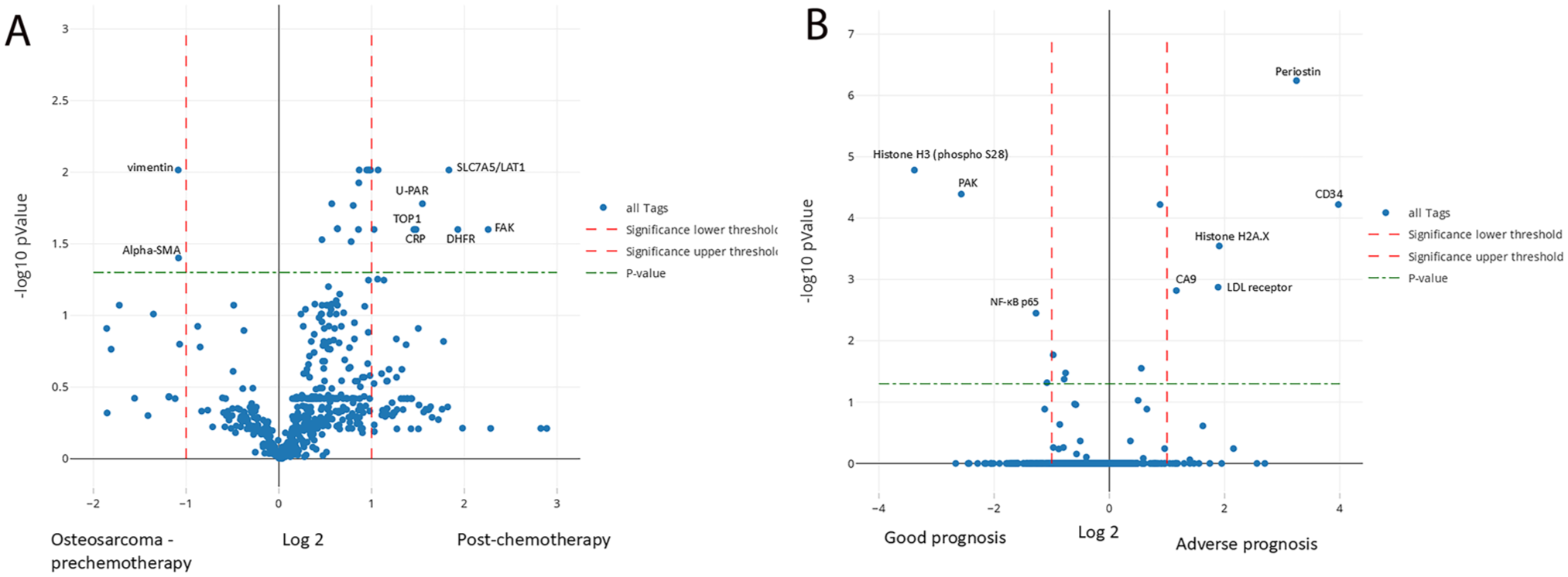
Aggressive tumors are associated with different protein profiles. (A) OS chemotherapy-resistant specimen revealed expression of specific proteins and loss of expression of mesenchymal markers compared with the prechemotherapy specimen. (B) Similarly, ERMS with adverse outcome had a different protein profile compared with good prognosis ERMS. The y-axis represents log2 of fold change and the x-axis is −log10 p value.
IHC
Immunohistochemical staining findings mirrored DSP results. Percentage of cells with positive staining was calculated and classified as diffuse or non-diffuse. Diffuse staining, defined as staining of 80–100% of cells, of H3K27me3 was noted in 8/8 ES, 6/8 OS, and 3/5 ARMS (Table 3). The nuclear staining was localized to less than 50% of the cells in two ARMS and two ERMS and was completely negative in two OS and one ERMS. In positive cases, the staining intensity was stronger than normal stromal cells and endothelium of blood vessels (Fig. 3E). Diffuse staining, nuclear and cytoplasmic, of Cyclin D1 was present in 7/8 ES with absent or scant staining (<20%) in all the other tumors. Similarly, diffuse staining for S100, nuclear and cytoplasmic, was limited to OS (4/8) with the other OS cases showing only focal staining (20%–30%). The staining was also focal in one EWS and one ERMS and was absent in ARMS.
Immunohistochemical Methods and Staining Results of H3K27me3, S100 and Cyclin D1 in the Studied Sarcomas.
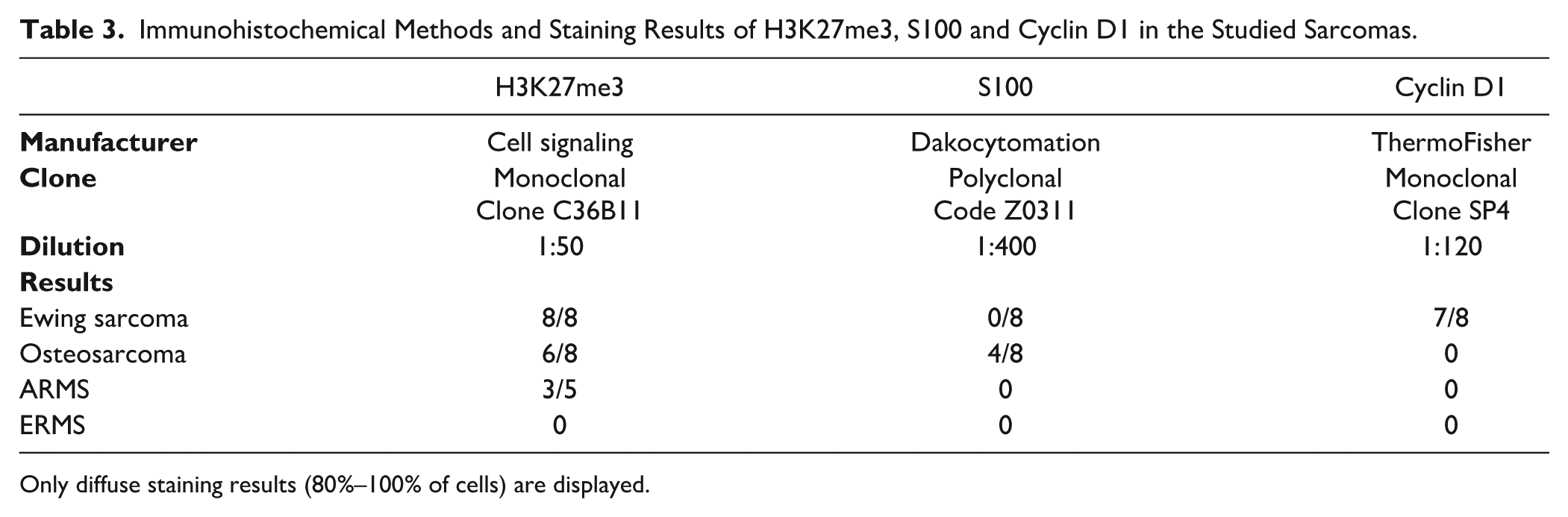
Only diffuse staining results (80%–100% of cells) are displayed.
Prognostic Classification
Few proteomic differences were identified in tumors from patients with good versus adverse prognosis. OS patients with adverse prognosis (n=4, deceased after a follow-up of <48 months) displayed loss of expression of several proteins including transferrin receptor and HSP27 (Phospho S78) (adjusted p=0.00026) in their tumors compared with patients with good prognosis (n=3, alive and well after a follow-up of 48–120 months) (Fig. 6D). ARMS patients (n=4) in this study were deceased after a short follow-up of less than 30 months. In contrast, ERMS patients (n=3) had variable outcome with two patients being alive and well after a follow-up of 94 and 108 months. A third 4-year-old male patient with ERMS of the soft tissues of the pelvis was identified to have HRAS (p.G13V) and BRAF (p.D594G) pathogenic alterations and subsequently died of disease 39 months after diagnosis. This patient’s tumor revealed significant expression of periostin, CD34, Histone H2A.X, CA9 and LDL receptor (adjusted p<0.00002) compared with the good prognosis tumors (n=2) that had increased expression of Histone H3, the P21 activated kinase (PAK) and NF-κB p65 (Fig. 7B). ES patients were also categorized into two prognostic groups. The adverse prognosis group (n=3, deceased after a follow-up of 36–54 months) displayed increased expression of several proteins including HLA-C, but there were no significant differences on adjusted p values.
Discussion
Our DSP profiling experiments have identified distinct tumor-specific protein signatures across the three most common pediatric sarcomas, characterized by the expression of multiple proteins. These expression patterns likely reflect underlying genomic alterations, including characteristic gene fusions and copy number variations, thereby providing insight into tumor-specific biology. DSP-based differential protein expression may enhance diagnostic accuracy in clinical specimens, inform risk stratification, and support the identification of patients who may benefit from more intensive therapeutic approaches. Furthermore, the highlighted proteins represent potential therapeutic targets, and their modulation using small-molecule inhibitors may offer novel avenues for targeted treatment in pediatric sarcomas.
DSP profiling of the top 20 expressed proteins (Fig. 2) revealed consistent enrichment of signaling pathways, epigenetic regulators and histone proteins, all of which play various critical roles in gene expression control and cell proliferation. Epigenetic regulation mediated by histone modifications and HDAC complexes has been recognized as a key driver of tumor growth, genomic stability, and transcriptional control in both ES and OS.12 –14 Emerging evidence also supports a contributory role of epigenetic alterations in RMS tumorigenesis and tumor classification.15,16 Elevated expression of histone proteins in these tumors was also confirmed by conventional IHC for H3K27me3, a key epigenetic marker associated with transcriptional regulation. EZH2 catalyzes trimethylation of histone H3 at lysine 27 leading to the formation of H3K27me. Accordingly, H3K27me3 expression serves as a surrogate indicator of histone-mediated regulatory activity in multiple malignancies. Its role is complex and context-dependent, with both increased and decreased expression implicated at various stages of tumor initiation, progression, and metastasis.17,18 DSP has also highlighted the expression of NF-κB and MEK2, key components of the Ras/Raf/MEK/ERK mitogen-activated protein kinase (MAPK) signaling cascade, a pathway frequently dysregulated in oncogenesis. 19 In addition, the expression of Bid, Bax, and Smac/DIABLO, alongside phagocytosis-associated molecules such as calreticulin and glyceraldehyde-3-phosphate dehydrogenase (GAPDH), suggests activation of apoptotic and immunogenic cell death pathways. These processes are integral to tumor biology and have significant implications for immune modulation and the development of targeted therapeutic strategies.19 –22
Among the highly expressed proteins in ES, Caveolin-1, IGF2BP1, and Cyclin D1 have been implicated in its oncogenic transformation, which is primarily driven by the EWSR1–FLI1 fusion. Caveolin-1 is a direct transcriptional target of the EWSR1–FLI1 fusion protein and contributes to downstream activation of oncogenic signaling pathways, including MAPK/ERK and insulin-like growth factor (IGF) signaling, and upregulation of proteins such as IGF2BP1, CD99, and CDK4/CCND1, which collectively promote tumorigenesis, proliferation, and metastatic potential in ES.23,24 –26 Expression of Caveolin-1 and CD99, has been utilized in the immunohistochemical identification and therapeutic targeting of ES.27,28 In our study, immunohistochemical analysis confirmed high Cyclin D1 expression in ES, supporting its potential utility as an adjunct diagnostic marker in distinguishing ES from other small round cell tumors. 28 Additionally, expression of the MHC class I antigen HLA-C, was observed in association with adverse clinical outcomes, suggesting a potential role in prognostic classification. 29
DSP has unveiled multiple differentially expressed proteins in various subcategories of OS. A notable finding is the consistent differential expression of S100A4 in OS. S100A4 belongs to the calcium-binding S100 protein family which has been implicated in promoting OS tumor growth and metastatic progression. Functional studies have demonstrated that S100A4 activation enhances cellular migration, invasion, and metastatic potential, whereas its knockdown suppresses these oncogenic properties. 30 Selective expression of S100A4 in OS, with minimal or absent expression in other sarcoma subtypes, reflects its potential utility as a discriminatory biomarker in differentiating primary OS from metastatic tumors involving bone. This observation was supported by IHC for the parent family member, S100, which demonstrated predominant expression in OS specimens. This study has also provided insight into OS and microenvironment interaction, particularly highlighting the upregulation of specific proteins in tumor cells adjacent to stromal inflammatory components. Such localized protein expression may contribute to tumor progression by promoting immune evasion, enhancing tumor growth, and conferring resistance to therapeutic interventions. 31 Furthermore, DSP has unveiled loss of expression of transferrin receptor and the heat shock protein 27 (HSP27) in OS cases associated with unfavorable clinical outcome (Fig. 6D). Although these proteins are often oncogenic, they may play a tumor suppressive role in OS. Loss of HSP27 is associated with dysregulation of intracellular iron metabolism leading to increased tumorigenesis and adverse patient’s prognosis. 32 Additionally, reduced expression of mesenchymal differentiation markers, including vimentin and alpha-smooth muscle actin was observed in the aggressive chemotherapy-resistant recurrence specimen (OS2) of the patient who succumbed to progressive disease. During its development, OS differentiates along multiple mesenchymal lineages with heterogeneous expression of related markers such collagens, vimentin, fibronectin, PDGFRA, and smooth muscle actin alpha 2. The degree of this differentiation appears to influence tumor behavior, with loss of differentiation associated with increased aggressiveness, therapeutic resistance, and poorer overall survival. 33
RMS tumors represent a heterogeneous group characterized by upregulation of myogenic transcription factors. According to DSP, they expressed the high mobility group protein HMGB1 which is known to stimulate myogenic differentiation. 34 In contrast to ES and OS, they also demonstrated expression of IKKi/IKKє, a component of the IκB kinase enzyme complex which regulates NF-κB signaling and is involved in the inflammatory immune response and oncognesis. 35 ARMS shares morphologic similarities with ES as a small round cell tumor but is distinguished by distinct clinical and molecular features. Not surprisingly, ARMS lacked expression of markers commonly associated with ES including CD99, caveolin, cyclin D1 and BAD. ARMS tends to be clinically more aggressive than ERMS and is morphologically different with small round cells that may contain cytoplasmic vacuoles. The DSP finding of increased expression of fatty acid synthase in ARMS explains differences in lipid metabolism which may affect tumor morphology, cell proliferation and tumor growth. 36 Although ERMS generally has a more favorable prognosis than ARMS, it carries a wider behavioral spectrum. In the present study, the ERMS case associated with adverse outcome exhibited increased expression of several biomarkers, such as periostin and LDL receptor which are probably related to HRAS and BRAF alterations present in the tumor leading to subsequent activation of MAPK signaling. 37 Expression of CD34 and carbonic anhydrase CA9 likely reflects modulation of angiogenesis and development of tumor microenvironment, respectively. 38
In conclusion, this study underscores the utility of spatial protein profiling methods as a robust approach for characterizing protein expression in pediatric sarcoma FFPE tissue. Relevant literature review and IHC validation support the biological relevance of the identified proteins in tumor pathogenesis and diagnostic pathology. Although limited by a relatively small cohort, this study establishes the feasibility of applying DSP to clinical specimens and highlights its potential as a high-throughput screening tool in actionable biomarker discovery, comparable to other sequencing or multiplex immune-labeling methods. The identified proteins may also serve as candidates for the future development of diagnostic and prognostic assays. These findings provide a foundation for subsequent investigations involving larger patient cohorts and functional in vitro studies to further elucidate the role of these biomarkers in tumor biology. DSP-based proteomic profiling addresses key challenges associated with limited tissue availability and quality, offering a scalable solution for comprehensive biomarker analysis in small samples. Integration of DSP into clinical trials may enhance biomarker-driven stratification and ultimately expand precision medicine approaches for pediatric sarcoma patients.
Supplemental Material
sj-xlsx-1-jhc-10.1369_00221554261455962 – Supplemental material for Spatial Proteomic Profiling, a Novel Method for Detecting Diagnostic and Prognostic Proteins in Pediatric Sarcoma
Supplemental material, sj-xlsx-1-jhc-10.1369_00221554261455962 for Spatial Proteomic Profiling, a Novel Method for Detecting Diagnostic and Prognostic Proteins in Pediatric Sarcoma by Abderrahman Ouban, Amani Alkofide, Antonello Podda, Awatif Eid Alanazi, Nader Ashraf, Tarek Ziad Arabi, Belal Nedal Sabbah, Katherine Elston, Vijay Baichwal, Erin Rudzinski, Eleanor Y. Chen, Nya D. Nelson, Catherine Albert and Atif A. Ahmed in Journal of Histochemistry & Cytochemistry
Supplemental Material
sj-xlsx-2-jhc-10.1369_00221554261455962 – Supplemental material for Spatial Proteomic Profiling, a Novel Method for Detecting Diagnostic and Prognostic Proteins in Pediatric Sarcoma
Supplemental material, sj-xlsx-2-jhc-10.1369_00221554261455962 for Spatial Proteomic Profiling, a Novel Method for Detecting Diagnostic and Prognostic Proteins in Pediatric Sarcoma by Abderrahman Ouban, Amani Alkofide, Antonello Podda, Awatif Eid Alanazi, Nader Ashraf, Tarek Ziad Arabi, Belal Nedal Sabbah, Katherine Elston, Vijay Baichwal, Erin Rudzinski, Eleanor Y. Chen, Nya D. Nelson, Catherine Albert and Atif A. Ahmed in Journal of Histochemistry & Cytochemistry
Supplemental Material
sj-xlsx-3-jhc-10.1369_00221554261455962 – Supplemental material for Spatial Proteomic Profiling, a Novel Method for Detecting Diagnostic and Prognostic Proteins in Pediatric Sarcoma
Supplemental material, sj-xlsx-3-jhc-10.1369_00221554261455962 for Spatial Proteomic Profiling, a Novel Method for Detecting Diagnostic and Prognostic Proteins in Pediatric Sarcoma by Abderrahman Ouban, Amani Alkofide, Antonello Podda, Awatif Eid Alanazi, Nader Ashraf, Tarek Ziad Arabi, Belal Nedal Sabbah, Katherine Elston, Vijay Baichwal, Erin Rudzinski, Eleanor Y. Chen, Nya D. Nelson, Catherine Albert and Atif A. Ahmed in Journal of Histochemistry & Cytochemistry
Supplemental Material
sj-xlsx-4-jhc-10.1369_00221554261455962 – Supplemental material for Spatial Proteomic Profiling, a Novel Method for Detecting Diagnostic and Prognostic Proteins in Pediatric Sarcoma
Supplemental material, sj-xlsx-4-jhc-10.1369_00221554261455962 for Spatial Proteomic Profiling, a Novel Method for Detecting Diagnostic and Prognostic Proteins in Pediatric Sarcoma by Abderrahman Ouban, Amani Alkofide, Antonello Podda, Awatif Eid Alanazi, Nader Ashraf, Tarek Ziad Arabi, Belal Nedal Sabbah, Katherine Elston, Vijay Baichwal, Erin Rudzinski, Eleanor Y. Chen, Nya D. Nelson, Catherine Albert and Atif A. Ahmed in Journal of Histochemistry & Cytochemistry
Footnotes
Acknowledgements
The authors acknowledge with gratitude the tremendous work of the histology staff of Seattle Children’s Hospital for preparing study materials. This manuscript was presented in abstract form at the 26th Annual Meeting of the Congress of Clinical Chemistry and Laboratory Medicine, Brussels, Belgium, 2025.
Competing Interests
The authors declared the following potential conflicts of interest with respect to the research, authorship, and/or publication of this article: K.E. and V.B. are employees of Bruker Spatial Biology, whose technology is the focus of this research. Other authors report no conflicts of interest.
Author Contributions
AO, AA, AP, NA, TA, AEA, BNS, AAA: Substantial contributions to conception and design, and acquisition of data. AAA: supervised clinical data and allocation of experiments. AO, VB, KE, NDN, CA, AAA: Substantial contributions to analysis and interpretation of data. AO, ER, EC, CA, AAA: Drafted the article and revised it critically for important intellectual content; and AO, ER, EC, CA, AAA: Final approval of the version to be published. VB is deceased.
Funding
The authors disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: Sanad Children’s Cancer Support Association, Riyadh, Saudi Arabia (grant #RGP-2020-21).
Supplemental Material
Supplemental material for this article is available online.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.