
Abstract
Objective
Virtual surgery and virtual patients necessitate quantitative data on the area of interest. The study was conducted to exactly describe the embryonic and fetal uvular muscle (MU), relevant for clinical as well as virtual surgery and virtual patient generation.
Method
Serially sectioned viscerocrania of 10 aborted embryos and fetuses underwent three-dimensional reconstruction to obtain detailed anatomic data and perform finite element analyses.
Results
The MU was paired in 80% of cases, while 20% allowed no clear-cut distinction. The MU merged with the levator muscle beneath the palatal aponeurosis without a hard palate insertion. Superior longitudinal central fibers ran below the nasal mucosa, and few circular peripheral fibers crossed in the central third to the contralateral side. This was seen in 30% of the paired muscles and in all cases when no differentiation was possible; about 40% to 80% MU fibers crossed to the ipsi lateral and contralateral palatopharyngeus muscle behind the levator loop. MU fibers inserted 60% nasal and 40% oral to the basal membrane at the middle third of the macroscopic uvula, made of loose connective tissue and salivary glands. The results of the finite element simulation of the uvula showed no distinct patterns or distributions of local stress.
Conclusions
Detailed anatomical study supported the concept of mediocranial MU repositioning during corrective surgery, although the impact is minor to the levator muscle's action. Future mathematical models describing effects of such a maneuver should integrate surrounding structures.
Keywords
Background and Objective
The primary objective for surgical repair of cleft palate (CP) is the development of normal speech. The treatment outcomes have greatly improved with the evolution of today's palatoplasty techniques (Von Langenbeck, 1861; Ganzer, 1920; Ernst, 1925; Dorrance, 1930; Whillis, 1930; Veau, 1932; Browne, 1935; Wardill, 1937; Oldfield, 1941; Braithwaite, 1964; Ruding, 1964; Singer, 1964; Braithwaite and Maurice, 1968; Kriens, 1969; Delaire, 1972; Kriens 1997; Schendel et al., 1999). Analyzing late outcomes, several authors focus on the uvular muscle (MU, also musculus uvulae) in search of even better outcomes and lower rates of secondary surgery (Azzam and Kuehn, 1977; Lewin et al., 1980). For a long time the MU function was little understood, considered as being without a functional role and without surgical significance (Ruding, 1964; Dickson, 1975; Furlow, 1986; David et al., 1999). According to Dickson (1975) the MU and especially its main bulk lies superior to the levator at the dorsal velum, directly below the nasal mucosa (Dickson and Dickson, 1972; Dickson, 1975; Langdon and Klueber, 1978; Klueber and Langdon, 1979). Unpaired it traverses the velum in an anterioposterior direction, superior to all other muscles. Originating apparently from the anterior velar aponeurosis with occasional fibers passing to the posterior nasal spine, it ends inserting into the uvular basis. The muscle creates a hump in the dorsal velar third, which on velopharyngeal closure becomes displaced the most, short above its contact to the dorsal pharyngeal wall. Called the levator eminence, it appears reasonable to call the hump the uvular eminence (Dickson and Dickson, 1972; Dickson, 1975; Azzam and Kuehn, 1977; Millard, 1980).
Clinical nasoendoscopy confirms this eminence to occupy the central transversal velar third, fitting into the concavity at the posterior superior pharyngeal wall (Piggot, 1969; Piggot et al., 1969). The possible conclusion is a suitable convexity for velopharyngeal closure not in focus in traditional veloplasty. In contrast, Azzam and Kuehn (1977) found the muscle paired, originating lateral to the midline at the tendinous palatal aponeurosis posterior of the hard palate, anterior to the levator veli palatini. In their study on seven adult human noncleft cadavers, they describe both bundles to converge overlying the levator muscle sling and terminating as separate bundles between mucous glands of the uvula proper and the basal membrane. Because of localization and size, contraction is anticipated to elevate the soft palate. Huang et al. (1997) described a paired muscle in adult noncleft cadavers that extended between tensor aponeurosis anterior and the uvula posterior along the nasal aspect of the velum, exhibiting no attachment to the hard palate. Similar to Azzam and Kuehn (1977), they hypothesized a muscle contraction to increase to the midline bulk on the nasal aspect of the velum, contributing to levator eminence, extending the velum towards the posterior pharyngeal wall, and maximizing velopharyngeal contact. Electromyography shows a similar activity to that of the levator veli palatini, which implies an active role in velopharyngeal closure (Kuehn et al., 1988). The pharyngeal plexus does not innervate the muscle, but the lesser palatine nerve and the lesser palatine artery and veins maintain its perfusion.
Uvular function is commonly hypothesized based on the results of anatomical dissections of inconspicuous adult samples without clefts, which are by no means a model close to the situation in an operated infant. The present trials were conducted to study selected histomorphological details of the prenatal anatomy of the MU and to obtain data of major relevance for future clinical procedures, mathematical models, and research activities on CP samples.
Materials and Methods
Samples
Serial sections of viscerocrania of 18 aborted embryos and fetuses beyond Carnegie Stage 20 (Hunter et al., 2003) were processed after parental informed consent and approval by the ethics board in charge (Landes et al., 2005, 2006). Samples beyond grade II maceration (Vogel 1997) were excluded. In brief, links between personal identifiers and a unique individual identifier (randomly assigned letters, A to R) were irretrievably destroyed, thus anonymizing the samples. The macroscopical area of interest was first preserved by fixation in 4% formaldehyde without further dissection. Two downstream approaches were chosen to allow comparative assessment of preparation artifacts. In randomized fashion 12 cases were chosen to be dehydrated and celloidin embedded, serially sectioned and stained with hematoxylin and eosin, mounted to glass slides followed by ≥3 months drying while six cases were sheet-plastinated in milled Biodurregd; (BiodurProductsregd;, Heidelberg, Germany) and stained using methylene blue and azure II (counter-staining with basic fuchsin). Eight midfaces were exhibiting various degrees of pathological clefting (Cohen et al., 1994) and therefore excluded from the present study. These samples will be analyzed in a subsequent article. After digitalizing the histological data, the MU of the remaining 10 cases (B, C, G, H, I, J, O, P, Q, and R) were interactively segmented using SeVise (a nonprofit software generated by the authors, free distribution for noncommercial professional use, see Landes et al., 2005, 2006), a computer program for segmentation, classification, and visualization. This resulted in a set of two-dimensional contours (segments) representing the MU along the x- and y-axis as circumstantiated by an average of 350 control points.
Vertical Alignment and Artifact Correction
Accuracy of the subsequent three-dimensional (3D) reconstruction of the MU was obtained by the simultaneous registration and rigid transformation of various control segments (muscles: buccinator, levator veli palatini, tensor veli palatini, palatopharyngeus, superior pharyngeal constrictor, and palatoglossus; bones like maxilla, palatine, sphenoidale, and vomer; arteries: pharyngeal ascending, major, minor palatine, ascending and descending palatine; veins: palatal, pharyngeal and venous plexus; other structures: palatal fascia, salivary glands, tonsils, tubal cartilage, membranous tube and others).
Every section's vertical thickness was assessed over neural, muscle, bone, cartilage, and connective tissue. A correction factor was derived from a vertical test piece calibration. Horizontal scaling used a reference object (Landes et al., 2005, 2006). An automatic matching algorithm (Arun et al., 1987; Umeyama, 1991) was used to align all polygonal contours that belonged to MU or the neighboring and control structures.
3D Reconstruction, Rendering, Volumetry, and Simulation
A resulting image dataset of approximately 2200 by 1700 by 80 voxels was visualized with direct volume rendering at real-time frame-rates using the volume graphics library VGL (Volume Graphics GmbH, Heidelberg, Germany). All visualization was performed by combined volume and polygon rendering, offered by the computer program DeformModeler (a nonprofit software generated by the authors, free distribution for noncommercial professional use, see Landes et al., 2005, 2006). 3D voxel models of each specimen with superimposed clipping box revealed views into the relevant structures. An interactive parameterization of the 3D model, transparency, smoothing, and histological textures were assigned individually. Objects could be seen as wire frames, transected at will, visualized in stereoscopy, and individually blinded out.
Three-dimensional mesh representations of whole segments were required to enable volumetry and mathematical modeling. Volumetric assessment of the MU was calculated from the generated 3D mesh. Muscular activity of the MU was calculated by finite element simulation using the library Sofar (developed at the Chair for Scientific Computing at the Technische Universität Dortmund and the Institute for Mathematics at the Humboldt-Universität zu Berlin; see Weichert et al. 2008, 2009a, 2009b, 2010; Walczak et al. 2009); resulting stress distributions to judge mechanical load were visualized with DeformModeler (for further details concerning technical aspects of the mesh generation process, the simulation with finite elements and visualization we refer to Weichen et al. 2008, 2009a, 2009b, 2010; Walczak et al. 2009).
The embryonic and fetal specimens were supplemented with morphological findings by archival histological slides of the uvula of two routinely autopsied and subsequently anonymized adults. These were triple-marked for their relevant structures (Gomori silver stain and polarized light microscopy). To generate the images, a histochemical ammoniacal silver impregnation procedure (conventional Gomori stain) was carried out to detect collagen (especially reticulin) fibers. Polarized light microscopy was performed to visualize their maturation. “Juvenile” fibers are known to have positive form and negative intrinsic birefringence, the latter of which was constructed to change to positive during the course of maturation. This conventional setting was supplemented by an immunohistochemical double-labeling approach using antibodies to myoglobin to flag striated muscle fibers (avidin biotin complex method) and anti-cytokeratin 7 to mark glands (alkaline phosphatase antialkaline phosphatase technique).
Results
Preparatory problems rendered the region of interest incompletely covered in one of the 10 cases. Only five cases provided complete data applicable to volumetry.
A paired midline MU was discerned in eight specimens (80%); two specimens (20%) did not allow light microscopic diagnosis of bilateral muscle bodies, based on data of the x- and y-axis only (Fig. 1). Bilateralism could be discerned only by adding the z-axis by conducting the 3D reconstruction (Fig. 2).
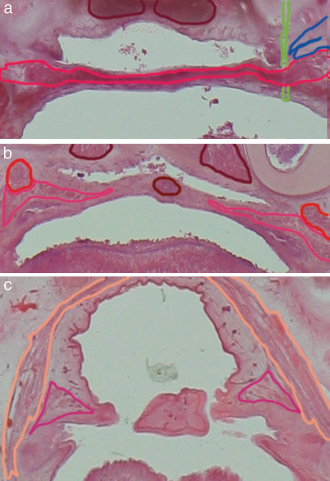
Case R (axial plane), a: Transection through the anterior (top is cranial) soft palate with dense connection of uvular to levator fibers with outlined anatomical structures of interest including palatine levator in red, segmented disruption artifact in green, dark red outlined longus capitis muscle and the Eustachian tube onset in blue, b: Transection more dorsal. The MU segmented in dark red is transected within its dorsal third lying median to the palatopharyngeus, the bilateral muscle bellies are separated by a connective tissue sheath, but segmented together in contrast to the cleft specimen in the subsequent report; outlined anatomical structures also include the palatopharyngeus muscle in pink, c: Transection at the tip of the uvula, sparse fiber connections merging to dorsal uvular basal membrane and minimal residual muscle; no connections to constrictor and tensor were seen. The outlined structures include the superior pharyngeal constrictor in light brown.

Control cases in 3D reconstruction. Case B from a: anterior, b: superior, c: inferior, d: dorsal. The anatomical structures are partially transparent for better viewing and not all structures are always displayed for better viewing. Bright red is the levator muscle; pink, the palatopharyngeus; violet, the tensor veli palatini running around the gray hamulus of the pterygoid process bilaterally; blue, the Eustachian tube; gray-blue, the palatal fascia; brown-red, the glossopharyngeal muscle; orange-brown, the upper pharyngeal constrictor. From posterior the close interdigitation of MU with the levator veli palatini (bright red) and the close MU vicinity and interdigitation to the palatopharyngeus can be observed. The palatopharyngeus itself clasps the levator muscle with two thirds of its fibers lying anterior to the levator and one third lying posterior to the levator. Even when differentiation on a light microscopic level allowed analysis of fiber distribution and interdigitation, the software's grade of resolution did not permit segmentation and subsequent 3D reconstruction. This is also due to mathematical error when volumes are attributed to two structures at the same time.
All cases showed dense muscle fiber interdigitation between the MU and the anterior levator veli palatini muscle. No direct insertion to the hard palate could be circumstantiated, one specimen showed single fibers extending to the palatal aponeurosis. The MU laid most superior in the velum below the nasal mucosa. Rather rostral parts of the samples, however, showed interdigitation with the levator below the palatal aponeurosis. The bilateral muscle bundles had mostly longitudinal central fibers and few circular peripheral fibers. No intrinsic or extrinsic bundle could be discerned even using additional techniques (e.g., fluorescence microscopy). The central and moreover the peripheral fibers terminated by densely interdigitating with the levator muscle. The muscle bellies spread anterior in all cases. Fibers crossed from 30% in paired muscles to 100% when no clear-cut differentiation was possible on two-dimensional light microscopical evaluation. In the anterioposterior plane, this occurred mostly in the central third of the velum.
Forty percent to 80% (mean: 50%) of the MU fibers crossed to the ipsilateral and contralateral palatopharyngeus muscle in the anterioposterior central to dorsal one third behind the levator loop. All cases showed MU fiber insertion to the basal membrane at the central third of the uvula. The latter itself consisted of loose connective tissue and salivary glands. Amongst these, 60% of the single muscle fibers revealed a rather nasalward direction while 40% went to the oral cavity. No connections to the upper constrictor were found. The tensor veli palatini was not connected to the MU.
This anatomical examination was carried out to analyze the quality of the data used for subsequent finite element simulation. At the current stage of the project, the muscles were described by a transversely, isotropic hyperelastic material model. It was assumed that a portion of the upper and lower cap of the mesh representation of the uvular muscle was fixed, while a volume force was applied to induce deformation. No interaction with surrounding structures was modeled. Varying the size of the caps, principal stress values were computed in due consideration of the quality of the underlying tetrahedral mesh and plotted as a color-coded representation of the stress values (Figs. 3 and 4). Surprisingly, some regions were coded red and blue, which represent stress peaks. These areas were situated in vicinity to the outer boundaries of the area where the muscle was fixed arbitrarily. However, this pattern was interpreted as a (possibly too) high local strain because it showed relative inhomogeneity (average values with higher variance). In contrast, stress levels were rather homogeneous (average values with lower variance) towards the center and off the caps of the virtual representation of the MU. These findings were consistent in all cases and cap sizes.
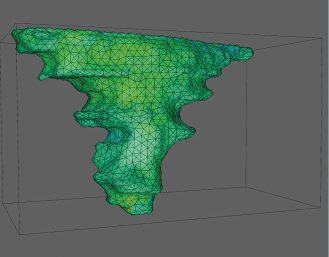
Stress value simulation. Case B anterior-lateral view arbitrarily chosen to exemplify plots of color-coded representations of the stress values associated with the respective uvular muscles. Except for the areas of simulated fixation, the muscles were lacking distinct stress peaks, a condition considerably suggestive of a rather homogeneous distribution of stress.
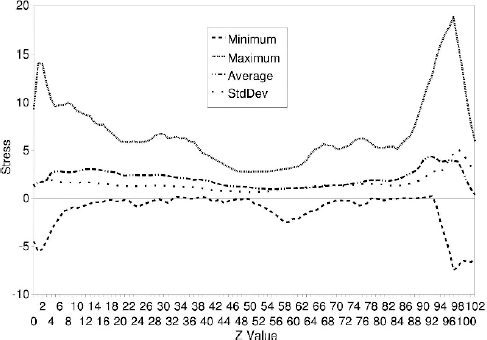
Stress diagrams. The minimum, maximum, and average stress values as well as the standard deviation along the z-axis (height) plotted for case B. Except for the areas of simulated fixation, the muscles lacked distinct stress peaks, a condition considerably suggestive of a rather homogeneous distribution of stress.
Results of triple marking of relevant structures suggest that the glanchmuscle ratio may be considerably higher in the adult uvula than that in the analyzed embryonic and fetal samples (Fig. 5).
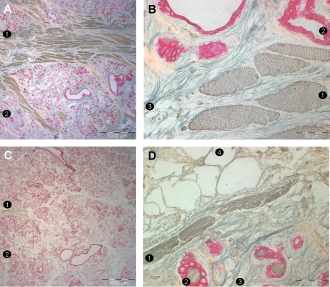
Adult uvula. The results of the present study should not be transferred to adult settings because considerable atrophy may be apparent later in life. A and B: A region near the base of the uvula of an adult. This individual has a considerable amount of muscle fibers. C and D: A region near the base of the uvula of another adult. This individual had only a few muscle fibers. 1, uvular muscle fibers; 2, glands; 3, collagen fibers; 4, adipocytes.
Discussion
As a preparatory step to virtual surgery and virtual patients (Moschos et al., 2004) as well as to also enhance clinical patient care, the anatomy of the MU was analyzed. Prospectively, larger anatomical entities will be included: described paired in normal anatomy (Azzam and Kuehn, 1977; Kuehn and Kahane, 1990; Huang et al., 1997), a single bundle is also reported (Dickson and Dickson, 1972) but was not found within this evaluation, yet partial merging of bilateral muscle bodies was present in 67%. Fluorescence microscopic sections supported the fact no intrinsic transverse fibers were seen (Azzam and Kuehn, 1977), yet internal longitudinal and external circular fibers were present. Anterior and posterior muscle fibers were found divergent from the median-sagittal. The MU evinced no hard palate origin or origin at the posterior nasal spine in contradiction to Schendel et al. (1999) but dense interdigitation to the anterior levator veli palatini, less so to the palatine aponeurosis contrary to Huang et al. (1997).
A wide consensus says that the MU acts as an intrinsic velar muscle contracting to change the shape of the velum (Calnan, 1954; Piggot, 1969; Piggot et al., 1969; Dickson and Dickson, 1972; Azzam and Kuehn, 1977; Croft et al., 1978; Lewin et al., 1980). On nasoendoscopy the MU bulges beneath the mucosa and maximizes velopharyngeal contact adding thickness to the elevated velum's dorsal third on lateral radiography and videofluoroscopy as well as anatomical dissection. The heaped up mound of mucosa is found in hemimaxillectomy patients on the nasal surface of the center of the posterior palate, medial of the levator. It overlies the levator sling, whose pull is primarily responsible for flexion and dorsocranial displacement of the velum (Boorman and Sommerlad, 1985). Differentiation of the proportionate impact of the levator and MU is impossible (Huang et al., 1997). Attenuation of such a bulge has been repeatedly described in velopharyngeal incompetence after CP repair and submucous CP (Piggot, 1969; Piggot et al., 1969; Croft et al., 1978; Lewin et al., 1980).
Electromyographic study showed MU activity to be present in all speech maneuvers, correlating with the levator's activity, and the MU was hypothesized to have additional active and passive space-occupying function in support of the levator's dorsocranial velar displacement (Kuehn et al., 1988). Increase in MU and velar diameter by active contraction, enhancing contact between the velum and the pharyngeal walls, was proposed, but this has so far not been substantiated (Huang et al. 1997).
The deformations calculated suggest that the levator not only lifts the nasal layer of a flaccid velum, but the MU's transmitted longitudinal bracing also enforces higher pressure of the dorsal connective tissue of the uvula to the dorsal pharyngeal wall. The levator, which alone enforces a much higher velar movement, basically transverses the middle part of the velum, but the dorsal third stays flaccid if not stiffened by the MU. Providing additional evidence, Simpson and Austin (1972), Mourino and Weinberg (1975), and Simpson and Colton (1980) described the anterior half of the velum as contributing little to the overall degree of velar stretch, while the greatest degree occurred in the posterior part. This may also be due to a fibrous inelastic palatal fascia that was found within the anterior velar half (Fig. 2).
The results of examining the origin and insertion of the MU at light microscopic scale suggest the MU functions as a “backbone” in prenatal individuals. Soft tissue deformation upon contraction of the MU could be cogitated to put considerable pressure on the dorsal pharyngeal wall.
Future finite element simulations using the virtual 3D objects that were generated in the present study may show stress patterns or peaks different from those currently found, once interaction with surrounding structures is implemented. Muscle fibers need to be prospectively segmented in direction, and the material model has to be extended to include fiber anisotropics.
Present findings from prenatal individuals, however, cannot be directly transferred into adults because the prenatal and adult MU might differ in relevance regarding function. Uvular ablation in snorers can be carried out without considerable detriment to speech or swallowing (Furlow, 1986; David et al., 1999). The increase of the gland:muscle ratio in older specimens appeared to be due to postnatal atrophy of the muscle. Striated muscle fibers were replaced by salivary glands and adipose tissue. Considerable inter-individual variability, however, can be observed even in samples from people of the same age group (note, for example, the differences in muscle fiber diameter in Fig. 5). These findings might have an impact on finite element studies on this anatomical region (which are considered for future craniomaxillofacial surgery simulations).
The impact of the minute muscle is minor compared to the influence the levator exerts and will possibly become quantifiable as much as mathematical deformation analyses may be applied in giving an idea for elastic muscle deformation using standard values. In normally occurring cleft uvulae, only minimal effect is present.
An adequate material model for muscle tissue has to take into account that muscle tissue is viscoelastic, inhomogeneous, and undergoes large deformations. Furthermore, since fiber families are embedded, usually within collagen or elastin, muscle tissues have to be considered as anisotropic. General time- or rate-dependent viscoelastic material models are described in the body of literature (Wriggers, 2001). However, those models do not include the particular structure of muscle tissues. Sources of error were general tangential compression and distension during the processing of serial sections. Although the vertical is constant with each transection, global deformation makes the exact fitting of multiple independent structures restricted to maximal congruence, which was calculated by a matching algorithm. This should not be over-interpreted though because multiple structures do equalize minor displacements through their required common fitting. Furthermore, the rendering process smoothes the rough edges resulting from super-positioning of each individual transection.
The number of cases in this study was limited because legal elective or spontaneous abortion at appropriate gestational ages is not frequent in Western Europe. The process of preparation is time consuming and laborious (Landes et al., 2005, 2006). Systems biological approaches providing data on deformation calculation may be one of the future standards in functional anatomy. A study of the cleft uvular microanatomy in a subsequent article will further corroborate the findings of this study.
Footnotes
Acknowledgments.
The authors wish to thank Karl Meller, M.D., PhD., Department of Anatomy and Cytology, Ruhr-University, Bochum, Germany, for his help in generating the celloidin-HE sections as well as Heinrich Mueller, Com. Sci., PhD., Department of Computer Science VII, Technical University, Dortmund, Germany, for his support in generating software. Moreover, the authors would like to express their gratitude to A. Schofer, M. Ertz, and G. Walter for their excellent technical assistance.