
Abstract
Background
In humans, the main sources of reactive oxygen species (ROS), the molecules causing oxidative stress, are mitochondrial superoxide ions and neutrophil-derived oxidative radicals. Circulating antioxidants contribute to the protection against oxidative stress. Although the formation of ROS and secretion of antioxidants are independently regulated by exercise and diet, little is known about their combined effect. We hypothesized that relatively brief, intense exercise training may reduce systemic oxidation via an intrinsic mechanism, independent of changes in circulating antioxidants and of neutrophil-derived enzymes (as may be caused by concomitant caloric restriction).
Methods
Nineteen volunteers exercised for 7 days, 3 hours/day at 75% of oxygen uptake. Caloric intake was either 110% of caloric expenditure (high calorie, n = 10) or 75% of caloric expenditure (low calorie, n = 9). Blood samples for F2-isoprostanes, catalase, myeloperoxidase (MPO), interleukin-x (IL-x), white blood cells (WBCs), and other metabolic variables were taken at baseline, at the end of training, and 1 week after completion of the study.
Results
Serum F2-isoprostanes (μg/mL), markers of lipid peroxidation, were similarly reduced after 7 days of exercise in the high-calorie (from 35 ± 4 to 27 ± 2) and low-calorie (from 35 ± 3 to 24 ± 2) groups. Similar reductions were observed in IL-x concentrations. Conversely, no change was observed in circulating concentrations of the antioxidant catalase. Whereas total WBCs and neutrophil counts were significantly reduced in the low-calorie group only, no difference in neutrophil-derived MPO was measured between groups.
Conclusion
A significant reduction in systemic oxidation may occur relatively early during intense exercise training in healthy young men, independent of caloric intake. The potential contribution to these effects of circulating antioxidants and neutrophil-derived oxidative enzymes will require further investigation.
Reactive oxygen species (ROS) are highly reactive metabolic by-products causing cytotoxicity, tissue injury, and dysfunction. Although these negative effects are normally prevented by a wide range of antioxidant molecules, an imbalance between the production of ROS and antioxidant defenses is defined as oxidative stress, a pathogenetic mechanism common to numerous human diseases.1,2
Physical activity and nutrition are important naturally occurring modifiers of systemic oxidative stress. Both in animal and human models, intense exercise, even of brief duration, can acutely induce oxidative stress.3–5 Conversely, repeated exercise protects against oxidative stress by potentiating antioxidant mechanisms, as observed in untrained humans and in amateur and professional athletesx (with the degree of antioxidant protection proportional to the amount and intensity of exercise training). 7 Antioxidants, however, significantly increase after weeks to months of training.7,8 An early reduction in systemic oxidation during training, therefore, may result from a direct reduction in mitochondrial superoxide ion formation; to date, this has not been clearly documented.
Diet is also a major regulator of systemic oxidation: ingestion and intravenous infusion 9 of lipids induce oxidative stress, 10 whereas fibers, fruits and vegetables, and caloric restriction reduce the formation of ROS. 11 Quantitatively, ROS production derives from two main mechanisms: mitochondrial O2 flow (of which up to 4% can be diverted to form ROS) 12 and release of neutrophil-derived myeloperoxidase (MPO), elastase, and other enzymes.13,14 Although the former mechanism is closely coupled with work intensity, the latter depends on the number, activation status, and chemotactic responsiveness of circulating neutrophils, characteristics that may be increased by hyperlipidemia 15 and reduced during starvation. 16
To date, it is still unclear whether early during exercise training, that is, before circulating antioxidants are increased, systemic oxidation may already be significantly reduced and whether modulation of neutrophil-derived oxidative enzymes (as may be induced by the relative leukopenia of caloric restriction) contributes to this reduction.
To explore this issue, we measured lipid peroxidation, neutrophil-derived MPO, circulating catalase, and leukocytes in a group of healthy young men who underwent 7 days of intense physical training (3 hours/day at ≈ 75% of their predetermined maximal aerobic capacity), with or without simultaneous caloric restriction.
Methods
Study Design and Sample Population
The study was approved by the Institutional Review Board, University of California, Irvine (UCI). Nineteen healthy male subjects, age 18 to 25 years (Table 1), after giving written informed consent participated in a 7-day intense exercise program during which they followed either a calorie-supplemented (high calorie) or a calorie-restricted (low calorie) diet. Subjects had a peak oxygen uptake &OV0312;O2 of 35 to 45 mL O2/kg/min and normal body mass index, did not smoke, did not have a history of chronic disease (including asthma; cardiorespiratory, musculoskeletal, or metabolic conditions; or eating disorders), and were not taking any medications.
Subject Characteristics
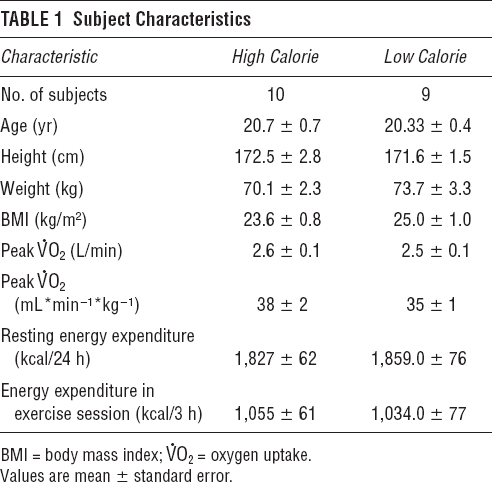
Preliminary Testing
All subjects underwent a preparticipation examination (medical history, physical examination, electrocardiography, and blood cell count) performed within a month prior to the beginning of the study by a physician who was not part of the study team.
Exercise Fitness and Basal Energy Expenditure
At least 1 week before starting the exercise program, subjects performed a ramp-type progressive exercise test on a cycle ergometer in which they exercised to exhaustion. Subjects were vigorously encouraged during the high-intensity phases of the exercise protocol. Gas exchange was measured breath by breath, and &OV0312;O2 peak was determined as previously described 17 using a commercially available metabolic cart (SensorMedics Corporation, Yorba Linda, CA).
Energy expenditure was determined by indirect calorimetry. Oxygen and carbon dioxide exchange were measured using a portable metabolic cart with an open-canopy system (Vmax calorimeter, SensorMedics Corporation). Measurements were performed at least 12 hours after meal consumption and after the patient had been recumbent for at least 30 minutes. Measurements were discarded if they failed to produce a steady state (defined as an average minute &OV0312;O2 and CO2 exchange with a variation < 10% during the testing period, typically 10 to 15 minutes). The flow, O2, and CO2 sensors were calibrated before each test.
Dietary Assessment
Prior to the start of the exercise program, participants completed a 3-day food record that was analyzed by the UCI General Clinical Research Center (GCRC) bionutrition manager using the Nutritionist Pro computerized database (First DataBank Inc., San Bruno, CA). This allowed us to assess each subject's eating patterns and exclude any significant difference in dietary habits among study participants while also allowing us to plan a palatable diet during the 7 days of intense exercise. Subjects were asked to avoid all dietary supplements and caffeine not only during the 7 days of intense exercise but also for the week preceding and the week following the exercise program.
7-Day Exercise Program
Starting on study day 1, subjects underwent an intense exercise training program for 7 consecutive days. Exercise was supervised by professional athletic trainers at the UCI student and faculty recreation center, which is a large, well-equipped sports and training facility located on the UCI main campus. Exercise prescriptions were generated individually according to resting energy expenditure and fitness level, as determined by preliminary testing. The goal of the training program was to increase total daily energy expenditure by approximately 25%.
Subjects exercised for 3 hours in the morning of each of the 7 exercise days, performing activities that included stepping, cycling/“spinning,” aqua-fitness, soccer, basketball, and jogging. Exercise procedures began at approximately 9:00 am following baseline blood draws, measurement of postvoiding weight, and breakfast provided by the GCRC bionutrition service. During the 3 hours of daily exercise, work intensity was individually adjusted for each subject to achieve an increase in energy expenditure of 20 mL O2/min/kg for 1 hour and of 10 mL O2/min/kg for the remaining 2 hours. To guarantee that exercise intensity at all times reflected predetermined target levels, heart rate was continuously monitored on Polar heart rate monitors (Polar Accurex Plus, Polar Electro, Woodbury, NY) and manually recorded every 10 to 20 minutes. All training regimens included brief warm-up and cool-down exercises.
Dietary Intervention
On the first day of the exercise program, subjects were randomly assigned to either a calorie-restricted or a calorie-supplemented group. Participants had not been previously informed of their group assignment to prevent individual changes in diet prior to the intervention.
In the high-calorie group, the total caloric intake for the exercise week was designed to exceed total caloric expenditure by ≈ 15%. Conversely, in the low-calorie group, the subjects’ diets were calculated to achieve a negative energy balance of about 33%. This figure was chosen so that data may be comparable to previously published calorie restriction studies, in particular with the work by Smith and colleagues. 18
During the 7 exercise days, breakfast and lunch were provided by the UCI GCRC Metabolic Kitchen. Meals were preportioned and individually provided with a known caloric content depending on which group the subject had been assigned to. Food items were weighed to the nearest gram before serving, and any uneaten items were evaluated after the meal. Subjects were given specific guidelines to follow for the caloric content of their evening meal. To document food and beverages consumed in the afternoons and evenings, subjects were asked to keep a log or diary of meals and any additional food or beverages consumed. Each morning, the previous day's records were reviewed by a dietitian to clarify any questions or problems that the subjects had regarding food portions, calorie content of foods, etc. The total caloric and nutrient consumption for the 7-day intervention was then evaluated, again using the Nutritionist Pro computerized database.
The subjects worked with the project GCRC dietitian to ensure that the diets (both low calorie and high calorie) were properly balanced and contained sufficient amounts of protein (> 1 g/kg/d), basic nutrients, and fluids. The GCRC dietitian was on call 24 hours a day to answer any dietary questions.
Blood Sample Analysis
Fasting blood samples were obtained by standard phlebotomy at approximately 8 am on days 1, 2, 8 (≈ 20 hours after the last exercise bout), and 14 (≈ 1 week after the first exercise bout). Part of each blood draw was preserved as whole blood with ethylenediaminetetraacetic acid and maintained at room temperature (23°C) for later flow cytometric analysis. The remaining blood volume was placed in an ice bath and immediately centrifuged. Aliquots of the resulting plasma were stored at -80°C for later analysis of F2-isoprostanes, MPO, and catalase. All specimens were analyzed in the same batch by technicians who were blinded to the order of the samples and grouping of subjects.
Flow Cytometry
Whole blood samples for flow cytometry (FACSCalibur, Becton, Dickinson, and Company, San Jose, CA) were tested using CellQuest software (Becton, Dickinson and Company, Franklin Lakes, NJ) to quantify leukocytes and lymphocyte subsets. A complete blood count (CBC) analysis was performed by using a Coulter STKS CBC Counter (Coulter Pharmaceutical, Inc., South San Francisco, CA). Whole blood was stained with monoclonal antibodies conjugated to various fluorochromes (Becton, Dickinson, and Company and Pharmingen, San Diego, CA). The lysing reagent was FACS Brand Lysing Solution (Becton, Dickinson, and Company), which caused a simultaneous lysis of red blood cells and partial fixation of leukocytes. Fluorescence compensation was performed using CaliBRITE beads (Becton, Dickinson, and Company) and FACSComp software. Optimal amounts of antibodies were used, and 8,000 to 15,000 events were analyzed per tube. Isotypic controls were used for each assay to determine nonspecific staining. For density determinations, flow cytometric estimation of antibodies bound/cell (ABC) was performed using Quantibrite PE beads (Becton, Dickinson, and Company). ABC, being the number of antibodies that bind to the specific cell or microbead population, provides a good approximation of antigen density expressed on the cell. The Quantibrite PE beads were run at the same instrument settings as the assay, and the fluorescent label 2 (phycoerythrin [PE]) axis was converted into the number of PE molecules bound/cell.
Markers of Oxidative Stress
F2-isoprostanes were quantified in plasma by a highly precise and accurate protocol using stable isotope dilution methodology that employs gas chromatography and negative ion chemical ionization mass spectrometry. 19
MPO levels in serum were determined by enzyme-linked immunosorbent assay (ELISA) with the use of BIOXYTECH MPO-EIA (Oxis Health Products, Portland, OR). Interassay coefficient of variation (CV) was 11.17%, intra-assay CV was 2.32%, and sensitivity was 1.5 ng/mL.
Catalase levels in serum were determined by ELISA with the use of BIOXYTECH Catalase-520 (Oxis Health Products); serum was pretreated using centrifugal ultrafiltration tubes with a 30,000 molecular weight cutoff (Millipore Corp., Bedford, MA). Filtered serum was assayed for catalase activity as per Catalase-520 protocol. Interassay CV was 2.38%, intra-assay CV was 4.03%, and sensitivity was 2 U/mL.
Cytokine and Leptin Assays
Tumor necrosis factor (TNF)-α, interleukin (IL)-x, IL-1β, IL-1Ra, and leptin serum levels were determined by ELISA with the use of R&D System Quantikine High Sensitivity kits (R&D System, Minneapolis, MN). Intra- and interassay CVs and sensitivity were as follows:
TNF-α: intra-assay CV 8.7 to 14.8%, interassay CV 16.1 to 22.6%, sensitivity 0.18 pg•mL−1 IL-x: intra-assay CV 3.8 to 11.1%, interassay CV 7.1 to 29.5%, sensitivity 0.0094 pg•mL−1 IL-1β: intra-assay CV 1.6 to 4.0%, interassay CV 5.3 to 9.0%, sensitivity 0.059 pg•mL−1 IL-1Ra: intra-assay CV 3.1 to 6.2%, interassay CV 4.4 to 6.7%, sensitivity 22 pg•mL−1 Leptin: interassay CV 3.5 to 5.4%, intra-assay CV 3.0 to 3.3%, sensitivity < 7.8 pg/mL
Statistical Analysis
Statistical analysis was performed by the UCI GCRC Biostatistics Core using the SAS statistics software package (SAS Institute, Cary, NC). Both the original sets of data and residuals were tested for normal distribution using the Shapiro-Wilk test. Two-way (time x group) analysis of variance (ANOVA) for repeated measures was then used to determine the effects of exercise on all variables. Repeated measures over time and groups (ie, both low and high calorie) were accounted for in the covariance structure of the ANOVA models.
Pairwise t-test comparisons of interest (two-tailed) were made for each outcome variable when main effects were found to be significant using the Tukey test. Data are presented as mean ± standard error of measurement. Statistical significance was set at p < .05.
Results
The exercise protocol was well tolerated by all participants, and adherence to the assigned caloric regimens was confirmed by changes in body weight, which at the end of the 7 days of exercise training reflected the expected changes based on calorie balance calculations (there was an average significant 1.45 ± 0.25 kg body weight loss in the low-calorie group and a significant 0.89 ± 0.27 kg weight gain in the high-calorie group).
Dietary Intake
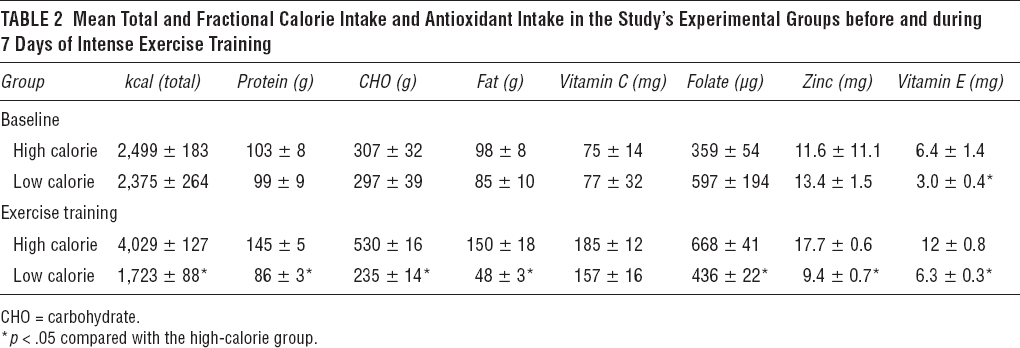
The mean caloric intake recorded during the 3 days preceding the exercise intervention was similar in the two experimental groups, in terms of both total calories and the percentage of calories per major nutrient group (Table 2). Similarly, no differences were observed in the mean intake of antioxidants, with the exception of a lower vitamin E intake in the low-calorie group. During the exercise intervention, on the other hand, both total and fractional caloric intake was significantly lower in the low-calorie group; despite a larger proportional consumption in fruits and vegetables, in this group, the intake of several major antioxidants (folate, zinc, vitamin E) was also significantly lower (see Table 2).
Mean Total and Fractional Calorie Intake and Antioxidant Intake in the Study's Experimental Groups before and during 7 Days of Intense Exercise Training
Markers of Oxidative Stress
F2-isoprostane levels were similar in the two groups before the start of the 7-day exercise program (35 ± 4 μg/mL in the high-calorie group, 35 ± 3 μg/mL in the low-calorie group) (Figure 1). No difference became apparent on the morning following the first day of exercise training (33 ± 4 μg/mL in the high-calorie group, 34 ± 3 μg/mL in the low-calorie group), but on the morning following the last day of exercise, F2-isoprostanes had become significantly and similarly reduced in both groups (27 ± 2 μg/mL in the high-calorie group, 24 ± 2 μg/mL in the low-calorie group; p < .05 vs baseline). Interestingly, this significant reduction below pre-exercise basal levels was still present 1 week after the end of the exercise program (28 ± 2 μg/mL in the high-calorie group, 27 ± 1 μg/mL in the low-calorie group; p < .05 vs baseline).
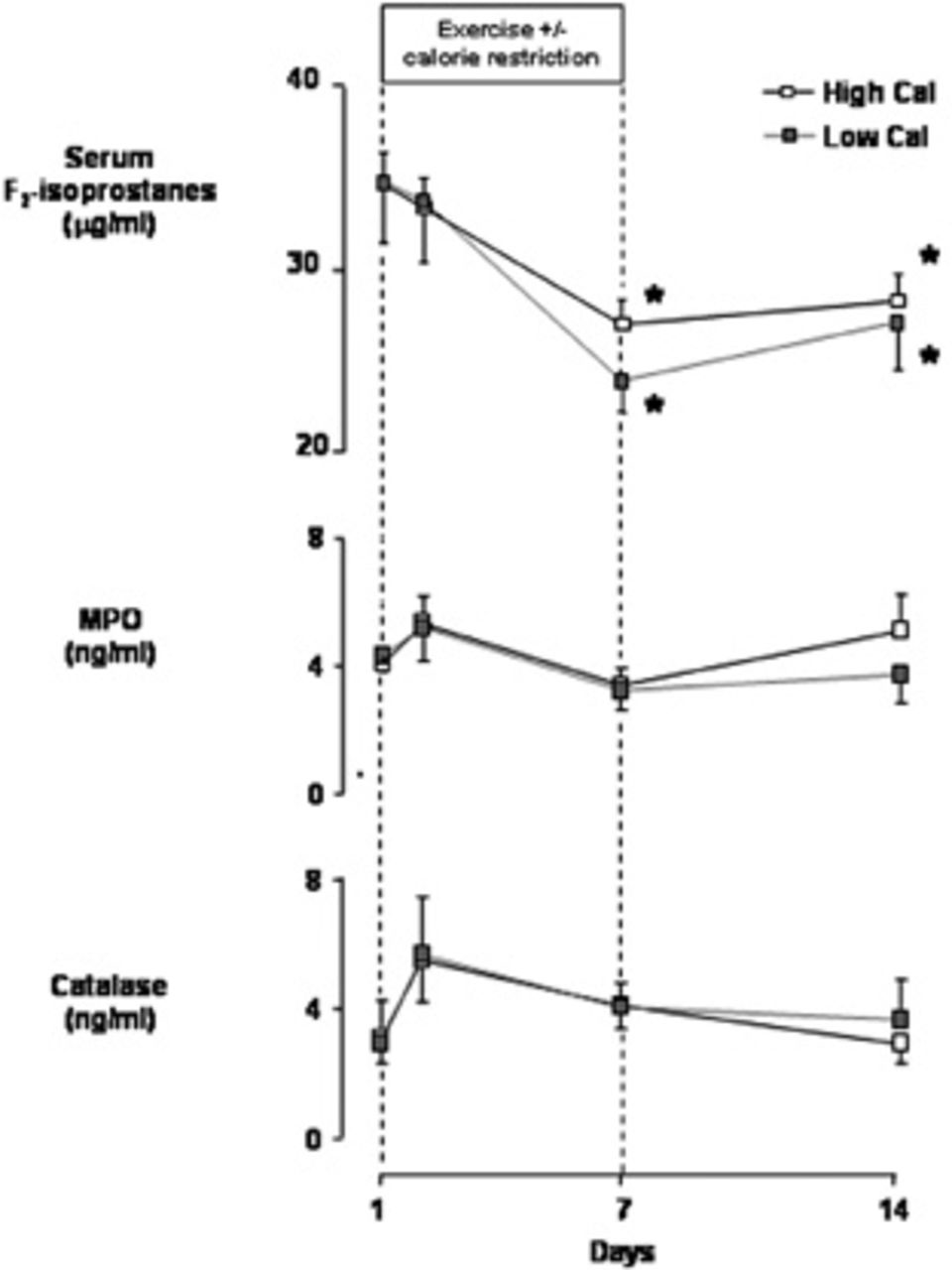
Serum levels of F2-isoprostanes, myeloperoxidase (MPO), and catalase in 19 healthy young male subjects at baseline and during and 1 week after completing a 7-day intense exercise training program (3 hours/day at ≈ 75% maximal work capacity). Subjects were separated into a weight maintenance group (High Cal, n = 10, dietary caloric intake 110% of estimated caloric expenditure) and a calorie-restricted group (Low Cal, n = 9, dietary caloric intake 75% of estimated caloric expenditure). Data are group mean ± standard error.
Serum MPO levels were also similar in the two groups before the start of exercise training (4.0 ± 0.5 ng/mL in the high-calorie group, 4.3 ± 0.5 ng/mL in the low-calorie group; see Figure 1). No statistically significant change in this variable was observed in either group during or after the exercise intervention; however, an identical response pattern occurred in both groups, with an initial ≈ 25% increase after the first day (5.3 ± 0.9 ng/mL in the high-calorie group, 5.2 ± 1.0 ng/mL in the low-calorie group) and a ≈ 40% drop by the end of the exercise intervention (3.4 ± 0.5 ng/mL in the high-calorie group, 3.2 ± 0.6 ng/mL in the low-calorie group; see Figure 1). One week after the end of the intervention, MPO levels were 5.0 ± 1.2 ng/mL in the high-calorie group and 3.7 ± 0.8 ng/mL in the low-calorie group.
Serum catalase levels were similar in the two groups at baseline (3.1 ± 0.8 ng/mL in the high-calorie group, 2.9 ± 1.3 ng/mL in the low-calorie group; see Figure 1) and, with the exception of small, nonsignificant increases observed on the morning following the first day of exercise (5.5 ± 1.3 ng/mL in the high-calorie group, 5.7 ± 1.8 ng/mL in the low-calorie group), remained close to baseline at both the end of exercise (4.1 ± 0.7 ng/mL in the high-calorie group, 4.1 ± 0.8 ng/mL in the low-calorie group) and 1 week postexercise (2.9 ± 0.6 ng/mL in the high-calorie group, 3.7 ± 1.3 ng/mL in the low-calorie group).
Cytokines and Leptin
Circulating concentrations of IL-x (Table 3) were not significantly different between groups at baseline (3.2 ± 1.0 pg/mL in the high-calorie group, 2.4 ± 0.5 pg/mL in the low-calorie group); at the end of training, IL-x was significantly reduced in both groups, although more markedly so in the low-calorie subjects (2.5 ± 0.8 pg/mL in the high-calorie group, p < .05 vs baseline; 1.3 ± 0.2 pg/mL in the low-calorie group, p < .01 vs baseline). By 1 week after the end of training, IL-x concentration returned (and indeed slightly surpassed) to baseline levels in both groups. Concentrations of TNF-a and IL-1b, as well as the anti-inflammatory cytokine IL-1Ra, were not affected by the exercise program and did not differ between groups at any time during the study (see Table 3).
Circulating Concentration of Inflammatory Cytokines and Leptin in the High-Calorie (n= 10) and Low-Calorie (n = 9) Groups
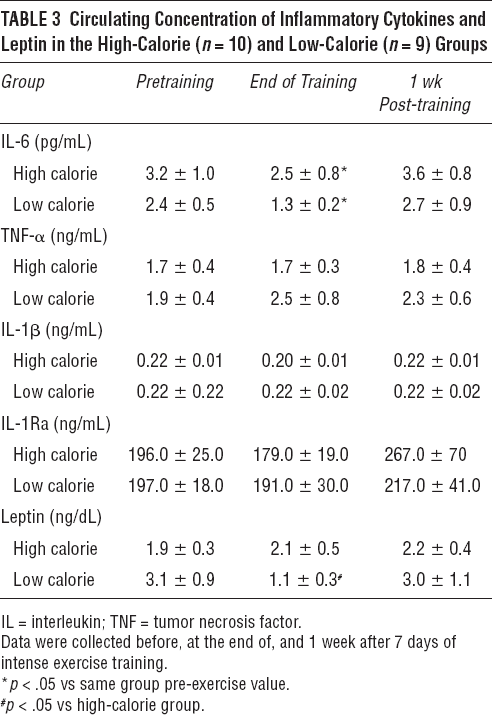
Circulating leptin was unaffected by experimental procedure in the high-calorie group; as expected, on the other hand, it was significantly reduced in the low-calorie subjects at the end of training but returned to baseline levels by 1 week after training.
Peripheral WBC Counts
The total WBC count (Figure 2) was similar in the two groups before the exercise program (5,700 ± 392 cells/mm3 in the high-calorie group, 5,933 ± 391 cells/mm3 in the low-calorie group); at the end of 7 days of exercise, the total number of WBCs remained unchanged in the high-calorie group (x, 150 ± 442 cells/mm3) but was reduced in the low-calorie group (4,244 ± 260 cells/mm3; p < .05). Interestingly, the decrease in WBCs in the low-calorie group was not homogeneous across cell subpopulations but was mostly accounted for by significant reductions in neutrophils (from 3,422 ± 418 to 2,178 ± 175 cells/mm3; p < .05) and monocytes (from 456 ± 34 to 322 ± 36 cells/mm3; p < .05) (see Figure 2). Although the total lymphocyte count was not reduced, a small but statistically significant reduction in B cells was observed (from 160 ± 16 to 127 ± 11 cells/mm3; p < .05) (Figure 3); T cells (total, CD4+, and CD8+) and natural killer cells were unchanged before and after the exercise program. In the high-calorie group, all WBC subpopulations were not affected by the 7-day exercise program.
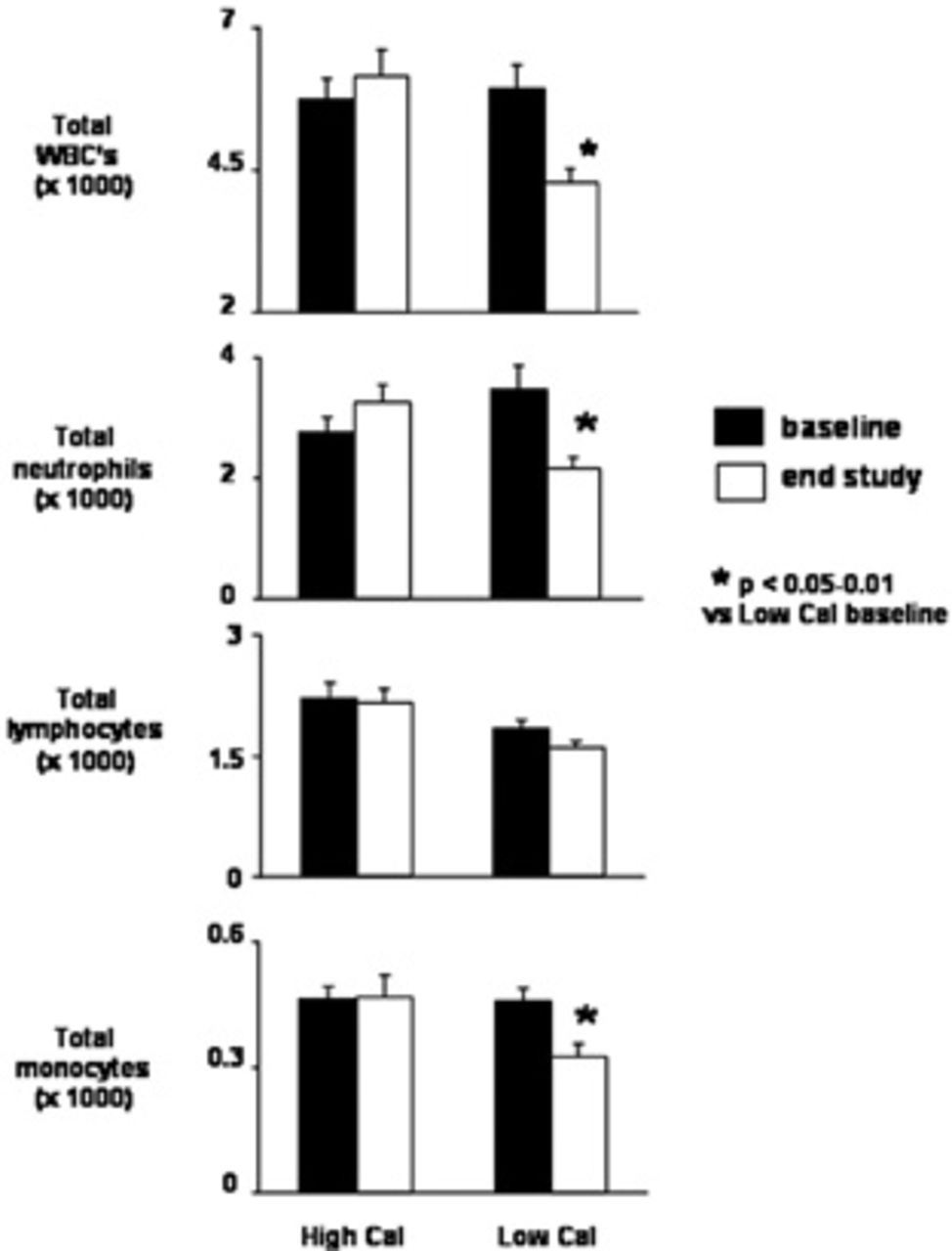
Circulating levels of total white blood cells (WBC's), neutrophils, lymphocytes, and monocytes in 19 healthy young male subjects at baseline and at the end of 1 week after completing a 7-day intense exercise training program (3 hours/day at ≈ 75% maximal work capacity). Subjects were separated into a weight maintenance group (High Cal, n = 10, dietary caloric intake 110% of estimated caloric expenditure) and a calorie-restricted group (Low Cal, n = 9, dietary caloric intake 75% of estimated caloric expenditure). Data are group mean ± standard error. *p < .05-.01 vs corresponding baseline value.
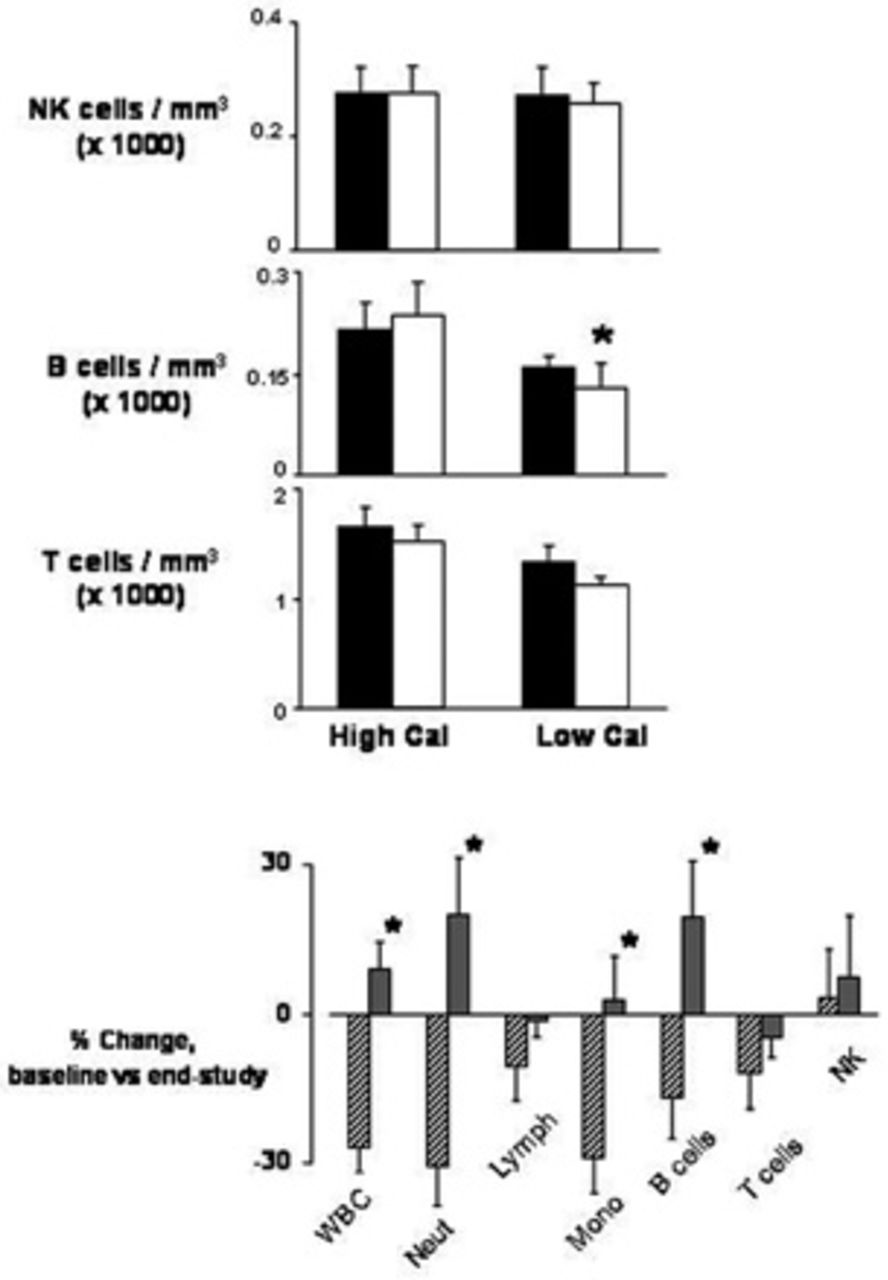
Circulating levels of total natural killer (NK), T, and B lymphocytes (Lymph) and percent changes in white blood cell (WBC) subpopulations in 19 healthy young male subjects at baseline and at the end of 1 week after completing a 7-day intense exercise training program (3 hours/day at ≈ 75% maximal work capacity). Subjects were separated into a weight maintenance group (High Cal, n = 10, dietary caloric intake 110% of estimated caloric expenditure) and a calorie-restricted group (Low Cal, n = 9, dietary caloric intake 75% of estimated caloric expenditure). Mono = monocytes; Neut = neutrophils. Data are group mean ± standard error.
Discussion
In our study, strenuous exercise for 7 consecutive days resulted in a significant reduction in basal lipid peroxidation, the quantitatively most important index of systemic oxidation. This occurred without apparent changes in concentrations of neutrophil-derived oxidative enzymes (MPO) or circulating antioxidants (catalase). Consistent with previous, longer training studies, the reduced systemic oxidative status that we observed confirms the ability of the human body to limit the damage of recurrent free radical formation. One of the novel findings of our study is that such a protective effect could be achieved in the relatively short time span of 1 week.
It has been shown how physical training enhances the activity of antioxidant enzymes, reducing the susceptibility of skeletal muscle to lipid peroxidation, with circulating antioxidant increasing proportionally to the amount and intensity of exercise performed.7,8,20 Increased antioxidant activity, however, does not gauge the subjects’ basal level of systemic oxidation. Our results clearly indicate that the actual basal level of systemic lipid peroxidation was lowered. Further, the fact that F2-isoprostanes remained significantly low for at least 1 week after training indicates that a residual memory of these oxidant stimuli may constitute a prolonged defense mechanism against oxidative stress. In recent years, F2-isoprostanes emerged as potentially ideal markers of oxidative stress in vivo. These compounds derive from the nonenzymatic free radical-induced peroxidation of arachidonic acid, 19 ubiquitously present in the phospholipid bilayer of cells and tissues throughout the organism. F2-isoprostanes display remarkable molecular stability, are measurable in all normal biologic fluids and tissues, and increase with known oxidant injuries. 21 Paralleling data obtained with other markers of lipid peroxidation, recent studies reported increased F2-isoprostanes following various formats of intense physical activity. 22 Oxidative stress has also been proposed to parallel inflammatory processes in numerous pathologic conditions.2,23 Interestingly, IL-x, the cytokine displaying the more robust and consistent changes in response to exercise, 24 decreased significantly in both experimental groups at the end of exercise training, paralleling F2-isoprostanes. This finding is consistent with previous observations of reduced IL-x in physically active versus sedentary women 25 and of reduced inflammatory markers by endurance training in subjects at risk of coronary events. 26
In contrast to previous, longer training studies, our experiments did not elicit significant changes in the circulating antioxidant catalase. Although we cannot exclude that alterations may have occurred in other antioxidants, which could not be measured owing to limited sample volume, this observation raises the possibility that exercise-induced adaptation to oxidative insults includes a rapid reduction in the rate of systemic oxidation followed by potentiation of antioxidant defenses, the latter requiring a more prolonged exercise intervention. As the other major pathway of ROS production, neutrophil-derived oxidative enzymes, was not altered by our exercise intervention, our data suggest that this early reduction in systemic oxidation may have been induced by a direct effect on superoxide ion formation in the mitochondrion. Although this could not be measured directly in our study, this possibility is consistent with previous reports indicating that the percentage of O2 molecules flowing through the mitochondrion can be modulated in differing metabolic conditions. 12
In our study, the protective effect of exercise against oxidative stress was independent of caloric balance. Although detecting the effect of caloric restriction per se was not one of our aims, it may be interesting to speculate what this effect may have been as diet is a known modulator of oxidative stress. 27 Available published data indicate that caloric restriction results in reduced production of reactive oxygen species, 28 increased antioxidants, 29 and reduced target tissue damage, including lipids, proteins, and deoxyribonucleic acid (DNA).10,30,31 This may explain the slightly greater, albeit nonsignificant, reduction in F2-isoprostanes in the low-calorie group at the end of training. Calorie restriction could also have an independent effect on WBC counts, but this normally occurs only after prolonged and intense nutrient restriction, as observed in starvation or chronic malnutrition states. Given the short duration and moderate level of calorie restriction used in our study (only ≈ 25% less calorie intake than prestudy levels in the low-calorie group), virtually no effect should be measurable on the circulating WBC populations owing to diet alone. Several previous studies, in fact, showed no change in WBC levels following caloric restriction of similar and greater duration and intensity than used in our experiments.32,33
An interesting finding from our study is the fact that at the end of training, serum MPO, the quantitatively most important neutrophil-derived oxidative enzyme, was similar in the two groups, despite different circulating neutrophil counts. Indeed, MPO similarly decreased in both groups at the end of training, paralleling the systemic rates of lipid peroxidation and suggesting that reduced MPO may have contributed to the exercise-induced reduction in systemic oxidation. The discrepancy between serum MPO and neutrophil counts is consistent with recent evidence that neutrophilic function may be highly susceptible to modulation by stress. Neutrophilic levels of gene expression and intracellular protein content have been recently observed to be acutely regulated by external stimuli, including physical exercise. 34 Fewer neutrophils could therefore conceptually sustain high circulating MPO levels if sufficiently activated.
A potential modulator of neutrophil concentrations or function is circulating cortisol, which is likely to increase significantly during the type of stress induced in our study.25,35 Although sample volume limitation did not allow cortisol measurements in our subjects, we cannot exclude that different increases in this hormone may have contributed to the observed effects on neutrophils. Finally, reduced circulating neutrophils may not necessarily reflect a net loss of cells but potentially just a redistribution, with marginated cells not contributing to the circulating pool but still secreting oxidative enzymes into the bloodstream.
Our calorie-restricted group received necessary amounts of all essential nutrients, including >1 g/kg/d of proteins, to ensure that metabolic disequilibrium, other than a negative caloric balance, did not occur and that protein breakdown did not contribute significantly to weight loss. Most of the 1.4 kg average weight loss that occurred in the calorie-restricted group, therefore, was accounted for by net loss of adipose tissue. As mobilized fat was rapidly metabolized to match the increased energy needs during training, however, circulating lipids were likely not increased, and, indeed, lipid peroxidation was similar between groups. It may also be noted that vitamin E ingestion was lower in the low-calorie group. This difference, however, is likely to have had negligible, if any, effects on our results. Prestudy vitamin E ingestion was very low in both groups, in fact, when compared with the recommended daily intake of 15 mg/d, and most of the between-group difference was due to high intake by a single subject. Further, ingestion of other antioxidants and circulating catalase were similar or greater in the low-calorie group. Finally, although training vitamin E ingestion increased more in the high-calorie group owing to the greater food intake, the vitamin E to insaturated fat ratio, a crucial index of vitamin E antioxidant capacity, was greater in the low-calorie group owing to much lower fat ingestion.
It should be noted that several of the mechanistic interpretations and conclusions presented in this article are based on the assumption that measured serum concentrations of oxidative markers closely parallel corresponding intracellular milieus. Although these assumptions are supported by the existing literature (such as the accuracy of F2-isoprostanes as indicators of systemic oxidation after exercise 36 and of MPO as an indicator of neutrophil-derived ROS production 37 ), direct confirmation of these assumptions will require further and more complex investigations.
In conclusion, basal levels of systemic oxidation, as reflected by circulating F2-isoprostanes, were significantly reduced by 1 week of intense exercise in a group of healthy young men, independent of caloric intake. The addition of caloric restriction to the exercise regimen, however, induced significant reductions in total WBC and neutrophil counts. Although the role of circulating antioxidants or neutrophil-derived oxidative enzymes will require further investigation, the reported reduction in systemic oxidative status suggests a direct inhibitory effect of exercise on mitochondrial free radical formation.
Footnotes
Acknowledgments
We would like to acknowledge the excellent support provided by the staff and nurses of the UCI General Clinical Research Center and by the staff of the UCI Anteater Recreation Center. We also acknowledge Dr. Jason Morrow and Erin Terry, from the Vanderbilt University Medical Center, for performing the F2-isoprostane assays.