
Abstract
Background:
It has been demonstrated that hypertrophic scar fibroblasts (HSFs) overexpress collagen messenger ribonucleic acid (mRNA) and protein, especially α1 collagen. Antisense nucleic acids are effective in inhibiting harmful or uncontrolled gene expression, suggesting that antisense ribonucleic acid (RNA) can effectively downregulate the expression of α1 collagen gene and attenuate the scars.
Aims:
This study was conducted to observe the effect of recombinant plasmid pREP9-COL1 on α1 collagen expression in HSFs and clarify the prospect of antisense RNA on scar treatment.
Methods:
The α1 collagen gene fragment including the region of 5′ UTR to exon (229 bp) was cloned in the eukaryotic expression plasmid pREP9 in the antisense orientation relative to the RSV-LTR promoter to reconstruct the pREP9- COL1 plasmid. Then it was transferred into HSFs through lipofectamine. The expression of α1 collagen was examined by immunostaining, reverse-transcriptase polymerase chain reaction, and Western blots.
Results:
The recombinant plasmid pREP9-COL1 with a correct sequence was constructed successfully; pREP9-COL1 consistently inhibited human α1 collagen gene expression at both mRNA and protein levels.
Conclusions:
Antisense RNA was effective in downregulating α1 collagen expression of HSFs. Therefore, this approach offered a prospect of scar treatment by attenuation of α1 collagen production with antisense RNA.
THE ETIOLOGY OF HYPERTROPHIC SCAR is still unknown. It has been demonstrated that fibroblasts from hypertrophic scar overexpress collagen messenger ribonucleic acid (mRNA) and protein, especially type I collagen. Therefore, the overexpression of α1 collagen is the central link during hypertrophic scar formation,1,–3 which indicates that prevention of hypertrophic scar through interfering in a link or many links of α1 collagen synthesis can have some future therapeutic value.
The abnormality of α1 collagen is often caused by mutation or abnormal expression of one of its genes. Gene therapy has been a hot topic in recent years. Antisense nucleic acids are effective in inhibiting harmful or uncontrolled gene expression.4,–6 It has been demonstrated that the synthesis of α1 collagen can be specifically inhibited in vitro by antisense oligodeoxynucleotides. 7 The naturally occurring antisense transcription was present in chick embryo chondrocytes with the downregulation of the α1 collagen gene expression, suggesting that the antisense gene could effectively downregulate the expression of the collagen gene. 8
This study was conducted to construct a recombinant plasmid pREP9-COL1, observe the effect of antisense ribonucleic acid (RNA) on α1 collagen expression in hypertrophic scar fibroblasts (HSFs), and clarify the prospect of antisense RNA in scar treatment.
Method
Cell Culture
The skin biopsies obtained from six patients who had no dermatologic or systemic pathology were confirmed as hypertrophic scars by pathologic examination. Informed consent was obtained from each individual, and the study was approved by the medical and ethical committees of the First Affiliated Hospital of Sun Yat-sen University.
Primary fibroblasts were prepared as previously described, with minor modifications. Briefly, the epidermis and dermis were separated by floating skin with 0.1% dispase (w/v) in RPMI 1640 at 4°C overnight. Fibroblasts were isolated by mincing the dermis into small pieces, followed by digestion with 0.75% collagenase (Gibco-BRL, Carlsbad, CA) in RPMI 1640 for 1 hour. The digestion was then rinsed once with RPMI 1640, and the cells were cultured in RPMI 1640 supplemented with 1% fetal bovine serum, 100 U mL–1 penicillin, and 10 μg mL–1 streptomycin at 37°C in a humidified atmosphere of 5% CO2 and 95% air. The cells from the second to fifth passages were used in the present study. The medium of cells was changed twice a week. At 70 to 80% confluence, cells were trypsinized and seeded in culture tubes at cell density of 5 × 106 mL–1.
Construction and Identification of pREP9-COL1
Human α1 collagen gene sequence was obtained from the National Center for Biotechnology Information gene bank, and the polymerase chain reaction (PCR) primer was designed by using Premier 5.0 software (PREMIER Biosoft International, Palo Alto, CA). The upper primer sequence was 5′-ATAGAAGCTTCCGCCTGTCCCAGT-3′, and the lower primer was 5′-CTTTGG- TCTGCATTCACA-3′; the length of the fragment was 229 bp. The β-actin upper primer sequence was 5′-GTGGGGCGCCCCAGGCACCA-3′, and the lower primer was 5′-CTCCTTAATGTCACGCACGATTTC-3′; the fragment length was 548 bp.
The total RNA was extracted from the human peripheral blood by using the Trizol total RNA extraction kit, reverse-transcripted, and amplified by PCR to prepare the α1 collagen complementary deoxyribonucleic acid (cDNA) fragment. Afterward, the target fragment was cloned into multiple cloning sites of the eukaryotic expression vector pREP9 in the antisense orientation relative to the RSV-LTR promoter, and received the new plasmid pREP9-COL1 highly purified, and kept at −20°C in an icebox for use. Then the plasmid was extracted using a small-scale plasmid DNA purification kit (Promega Corporation, Madison, WI) for identification by restriction enzyme and DNA sequence analysis.
Transfection
After being cultured in serum-free RPMI 1640 medium for 48 hours, cells were seeded in six-well plastic dishes at a density of 1.5 × 105 cell/cm2 for transfection of pREP9-COL1, and transfection was performed as recommended by the manufacturer of lipofectamine. The cells that underwent 72 hours' transfection were synchronized in serum-free medium for another 48 hours. These transfectants were selected in RPMI 1640 culture medium containing 800 μg/mL of G418. After about 3 weeks, small colonies of cells were scraped off from the flask and expanded as clones in culture medium containing 200 μg/mL of G418. The transfected cell lines were used for further analysis.
Immunostaining
Transfected or nontransfected fibroblasts were grown in the culture media. After reaching 90% confluency, the cells were washed with phosphate-buffered saline (PBS) and fixed with 100% methanol at 4°C for 2 minutes. The cells were washed with PBS again and incubated with a rabbit antihuman type I collagen antibody, followed by incubation with Texas redlabeled antirabbit antibody. The fluorescence label DAPI (4′,6-diamidino-2-phenylindole) was used for nuclear counterstaining. The images of fibroblasts with fluorescence-labeled proteins were acquired by fluorescence microscopy (Nikon, Melville, NY).
Quantitative Reverse-Transcriptase Polymerase Chain Reaction
Reverse-transcriptase polymerase chain reaction (RT-PCR) was performed as described previously. 9 The upstream and downstream primers were designed using Primer Express software 1.0 (PE Applied Biosystems, Foster City, CA). As described previously, total RNA from each sample was extracted from the cultured fibroblasts as described above using TRIzol reagent (Invitrogen, Carisbad, CA) and treated with DNase I (Ambion, Austin, TX). cDNA was synthesized using SuperScript II reverse transcriptase (Invitrogen). Synthesized cDNA was mixed with primers/probes in universal PCR buffer and then assayed on an ABI 7900 sequence detector (Applied Biosystems, Foster City, CA). The data obtained from the assays were analyzed with SDS 2.1 software (Applied Biosystems). The amount of total RNA in each sample was normalized with 18S recombinant RNA transcript levels.
Western Blot Analysis
Cells transfected with pREP9-COL1 and empty plasmid pREP9 were grown to 80% confluence in culture dishes. Cells were washed twice with PBS and covered with lysis buffer containing 20 mmol/L Tris Cl, pH 8.0, 20 mmol/L LiCl, 1% Triton X-100, and 1 mmol/L ethylenediaminetetraacetic acid. The liquids were centrifuged at 12,000 rpm for 10 minutes at 4°C, and the supernatant was transferred to a new tube. After protein concentration was measured, 200 mg of denatured protein was resolved on 15% SDS-PAGE and electroblotted onto nitrocellulose membranes (Amersham, UK). Membranes were incubated with mouse antihuman collagen α1 (Santa Cruz Biotechnology, Inc, Santa Cruz, CA) as the primary antibody (1:1,000), followed by incubation with horseradish peroxidase conjugated to goat antimouse IgG (Sigma-Aldrich, St. Louis, MO) as the second antibody (1: 5,000). Chemiluminesecence reaction was carried out for 1 minute using the ECL kit (Amersham, UK), followed by exposure to Kodak XAR film.
Statistical Analysis
All of the data were expressed as mean ± standard error and were processed by SPSS 11.0 for Windows (SPSS Inc, Chicago, IL). Two sets of isolated sample data were checked by t-test, and p < .05 served as significant difference.
Results
Identification by restriction enzyme and DNA sequence analysis showed that recombinant eukaryotic expression vector pREP9-COL1 with correct sequence and direction was constructed successfully (Figure 1).
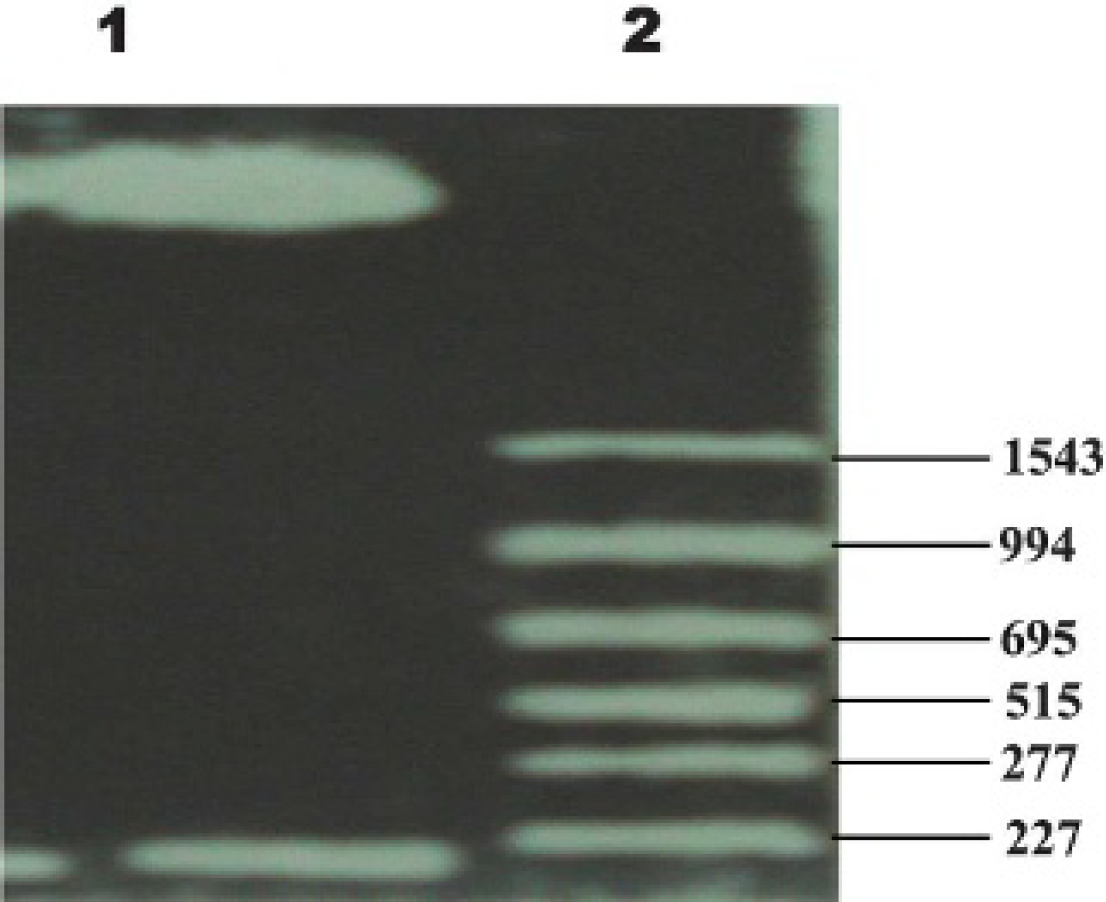
The pREP9-COL1 was treated with restriction enzymes, Xhol and BamHL (20 μL), at 37°C for 2 hours. Gelatin was used to withdraw the target fragment. Then the final product was taken and sent to identify its sequence.
Immunostaining revealed a relatively large amount of α1 collagen, which increased as early as 24 hours and reached higher levels at 72 hours in pREP9-transfected HSFs or parental HSFs (Figure 2). Conversely, the expression of collagen α1 in pREP9-COL1-transfected HSFs began to decline significantly at 24 hours, reaching the lowest peak at 72 hours and remaining at a very low level (Figure 3).
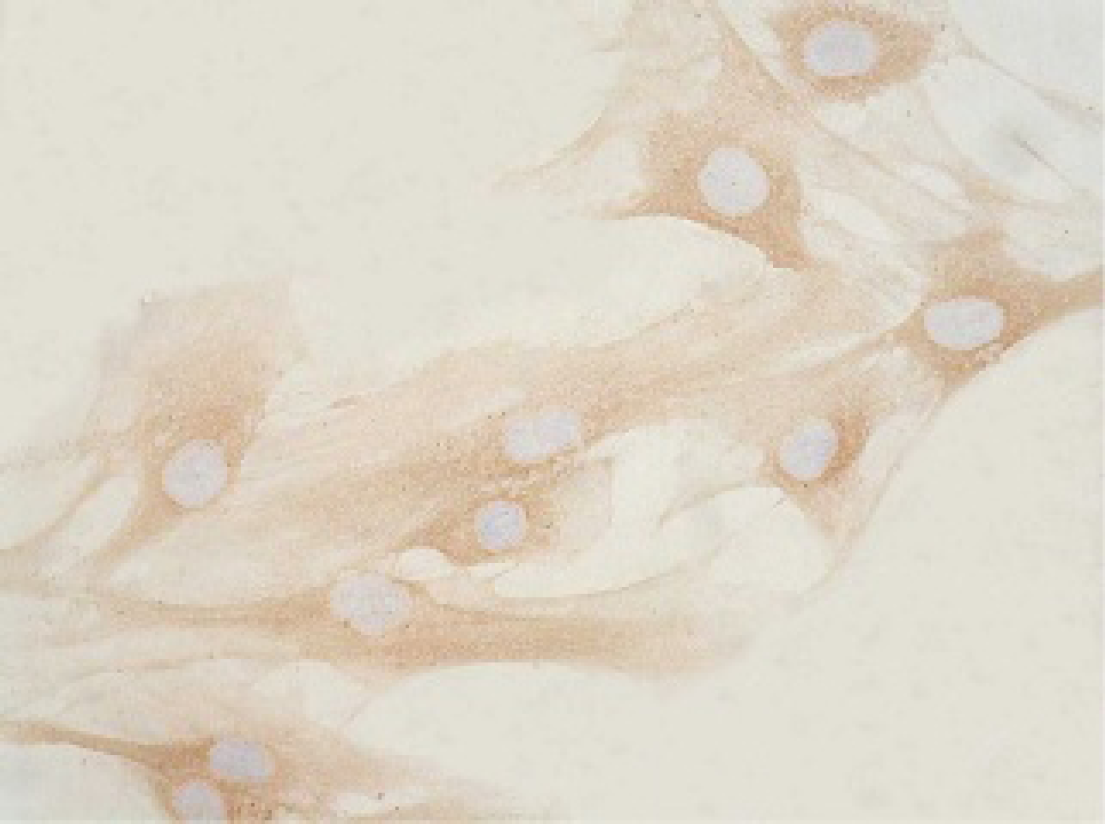
A relatively large amount of α1 collagen was detected; it increased as early as 24 hours and reached higher levels at 72 hours in pREP9-transfected hypertrophic scar fibroblasts (HSFs) or parental HSFs (see Figure 1). The positive cells expressing collagen α1 protein stained yellow (×200 original magnification).
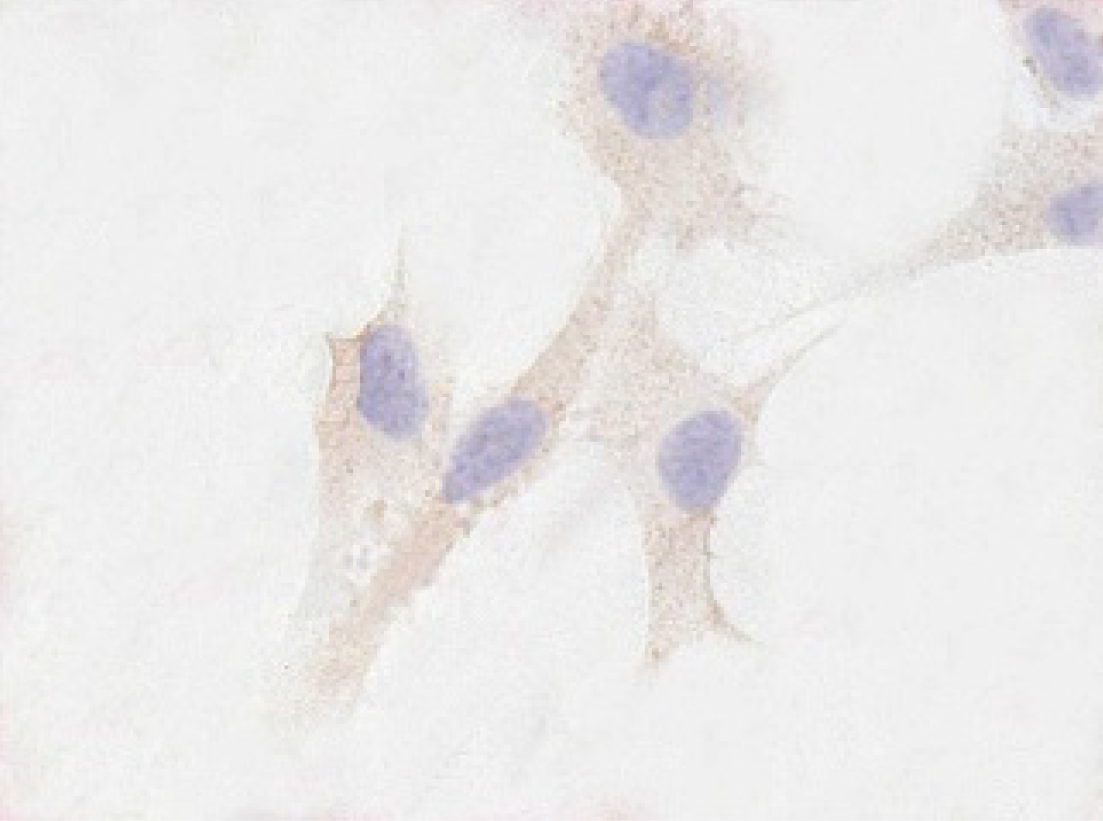
Conversely, the expression of collagen a1 in pREP9-COL1-transfected hypertrophic scar fibroblasts began to decline significantly at 24 hours, reaching the lowest peak at 72 hours, and remaining at a very low level. The positive cells expressing collagen α1 protein stained yellow (×200 original magnification).
Western blots showed that the pREP9-COL1-transfected HSFs showed a reduction in the α1 collagen protein by 68.46% compared with the parental HSFs and by 63.52% compared with the the HSFs transfected with empty pREP9 72 hours after transfection (Figure 4). The expression of α1 collagen protein in pREP9-COL1-transfected HSFs was significantly lower than that in pREP9-transfected or parental HSFs.
RT-PCR showed that the levels of collagen α1 mRNA in HSFs transfected with pREP9-COL1 were markedly downregulated by 46.87% (compared with the HSFs transfected with empty pREP9) and 42.68% (compared with the parental HSFs) (Figure 5).
pREP9-COL1 can consistently inhibit human α1 collagen gene expression at both mRNA and protein levels.

Effect of pREP9-COL1 on collagen α1 protein expression. The pREP9-COL1-transfected hypertrophic fibroblasts (HSFs) showed a reduction in the α1 collagen protein by 68.46% compared with the parental HSFs and by 63.52% compared with the HSFs transfected with empty pREP9 72 hours after transfection. The expression of α1 collagen protein in pREP9-COL1-transfected HSFs was significantly lower than that in pREP9-transfected HSFs or parental HSFs. Levels of α1 collagen are presented relative to β-actin messenger ribonucleic acid levels. Each column represents the mean ± standard error in each group. *p < .05 when compared with the corresponding controls.
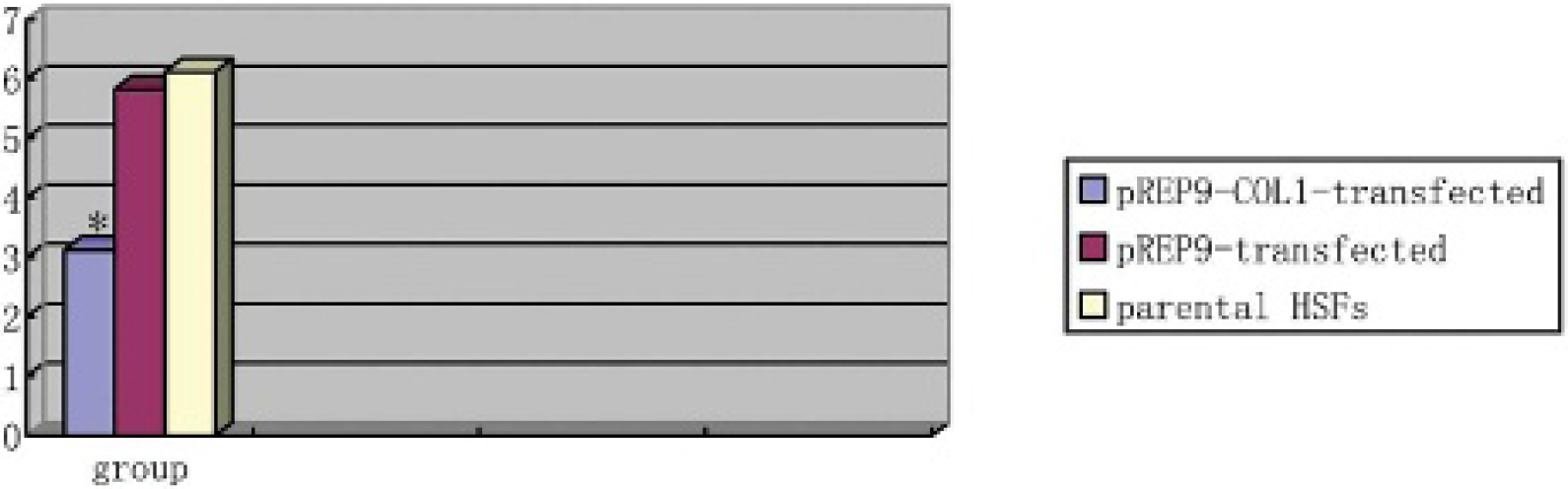
Effect of pREP9-COL1 on collagen α1 protein messenger ribonucleic acid (mRNA) expression. The levels of collagen α1 mRNA in hypertrophic fibroblasts (HSFs) transfected with pREP9-COL1 were markedly downregulated by 46.87% (compared with the HSFs transfected with empty pREP9) and 42.68% (compared with the parental HSFs). Levels of collagen α1 are presented relative to glyceraldehyde-3-phosphate-dehydrogenase mRNA levels. Each column represents the mean ± standard error in each group. *p < .05 when compared with the corresponding controls.
Discussion
Hypertrophic scar is characterized by excessive deposition of fibrosis and extracellular matrix (ECM) and is often cosmetically and functionally unappealing to the affected individual. 1 The mechanism of hypertrophic scar remains unclear. Currently, widely used conservative treatments of hypertrophic scar are limited to steroid injections, radiation therapy, pressure, and topical silicone gel application.2,10 Although effective, all of these treatments are inconvenient in some way to the patient and have no satisfactory effect. Therefore, hypertrophic scar presents a formidable therapeutic challenge.
In recent years, gene therapy has become the most attractive therapy, although the potential use of antisense RNA in gene therapy is an additional new frontier of molecular biology on the basis of existing molecular cloning and genetic recombinant techniques.9,11
An artificial expression vector (plasmid or bacterical vector) of antisense RNA can be constructed by positioning a DNA fragment coding for a portion of a specific mRNA between a suitable promoter and a transcription terminator in the antisense orientation through a genetic recombination technique. Then the expression vector will be transfected to the cell. The artificial antisense RNA transcripts from such a construction will be complementary to the target mRNA and specific inhibition of the expression of the gene targeted at the level of translation, transcription, and replication. So this approach is used as a powerful tool to dissect the expression and regulation of a specific gene and has been to applied to manipulate various bacterial and oncogenetic diseases. In addition to facilitating to dissect or identify the function of specific gene in the antisense orientation, artificial antisense RNA has provided a potential new solution to block or inhibit the expression of a harmful or uncontrolled gene. Moreover, development of antisense mRNA has provided insight and direction to the therapy of hypertrophic scar. Naturally occurring antisense transcription was present in chick embryo chondrocytes with the downregulation of α1 collagen gene expression, suggesting the possibility that antisense RNA can effectively down regulate the expression of the collagen gene.
In our study, an α1 collagen gene fragment including the region of 59 UTR to exon (229 bp) was cloned in the eukaryotic expression plasmid pREP9 in the antisense orientation relative to the RSV-LTR promoter to reconstruct the pREP9-COL1 plasmid so they could generate antisense RNA of α1 collagen. After the pREP9-COL1 plasmids were transfected into HSFs by lipofectamine, the antisense RNA had a complementary sequence against the special site of the α1 collagen gene. They could bind to the target mRNA sequence and prevent translation, transcription, and replication of the sense mRNA of collagen α1. The results showed that pREP9-COL1 could consistently inhibit human α1 collagen gene expression at both mRNA and protein levels.
The results of this study put forward a new thought on the treatment of hypertrophic scar. The antisense RNA of α1 collagen can further provide a new weapon against excessive synthesis and deposition of type I collagen. However, many problems of gene therapy remain unresolved. The transfection efficiency, stability, security, and immunoreaction that are adjustable to exogenous gene expression and carrier choice need to be thorougly studied.
Footnotes
Acknowledgments
We thank Dr. Shan Yuexing (Department of Biochemistry, Sun Yat-sen University) and Dr. Chen Jianding (Department of Forensic Medicine, Sun Yat-sen University) for their assistance.