
Abstract
Background
Adrenomedullin (AM) is a vasodilator peptide produced by endothelial and smooth muscle cells in the systemic and pulmonary circulation. It promotes angiogenesis and alveolar growth and has protective effects in the cardiovascular and respiratory systems. Adrenomedullin's role in human pulmonary vascular and alveolar development is unknown.
Objective
To test the hypothesis that AM is expressed during normal human lung development and that its expression changes with advancing gestational age by investigating the messenger RNA and protein expression of AM and its receptor components, calcitonin-receptorlike receptor (CRLR), receptor activity–modifying protein (RAMP)2, and RAMP3 in human fetal lung from 10 to 24 weeks of gestation.
Methods
The gene expression of AM, CRLR, RAMP2, and RAMP3 was measured with real-time reverse-transcription polymerase chain reaction. Adrenomedullin protein expression was measured with Western blot. Immunohistochemical analyses of sections of lung tissue were performed. Statistical analysis was performed using linear regression and one-way analysis of variance followed by the Tukey range test.
Results
Adrenomedullin, CRLR, RAMP2, and RAMP3 transcripts were expressed in the midgestation human fetal lung. The gene expression of AM, CRLR, and RAMP2 increased with increasing gestational age, whereas the gene expression of RAMP3 decreased. Adrenomedullin protein expression increased with increasing gestational age.
Conclusion
Adrenomedullin is expressed in the midgestation human fetal lung and its gene and protein expression increased with increasing gestational age, suggesting a role for AM in human lung development. Supporting this conclusion, the AM1 receptor components CRLR and RAMP2 gene expression also increased with increasing gestational age. Conversely, the expression of RAMP3, a structural component of the AM2 receptor, decreased with increasing gestational age, suggesting different functions for the AM receptors in human fetal lung, as it has been demonstrated in animal models. This speculation requires further investigation.
Preterm birth is associated with dysregulation of pulmonary vasculature development. In infants born during the late canalicular stage of lung development, there is evidence of marked angiogenesis disproportionate to the growth of the air-exchanging lung parenchyma, 5 abnormal distribution of alveolar capillaries, 6 prominent corner vessels with variable capillary density in adjacent alveoli, 7 and increased distance between capillaries and the air surface.8,9 Pulmonary hypertension is also being increasingly recognized in survivors of extreme prematurity.10,11 Developmental abnormalities of the pulmonary circulation, including a decrease in the number of pulmonary vessels, altered lung architecture, and episodes of hypoxemia and hypercarbia, may contribute to the development of pulmonary hypertension.
Adrenomedullin (AM), a potent vasodilator, originally isolated from human pheochromocytoma, is a 52–amino acid peptide hormone with structural homology to calcitonin gene-related peptide (CGRP). 12 It promotes angiogenesis and has protective effects on the cardiovascular and respiratory system. 13 Adrenomedullin is produced by the endothelial and smooth muscle cells of both the systemic and pulmonary circulation and its peptide and messenger RNA (mRNA) are expressed in multiple organs including adrenal gland, heart, kidney, and lung. 14 The activity of AM is mediated mainly by 3 different receptor complexes expressed from the calcitonin-receptorlike receptor (CRLR) gene and chaperoned to the cell surface by 3 different members of the receptor activity–modifying proteins (RAMPs), known as RAMP1, RAMP2, and RAMP3. The coupling of CRLR with RAMP2 forms the AM1 receptor, with high AM specificity, whereas the coupling of CRLR with RAMP3 forms the AM2 receptor with a lower specificity for AM. The coupling of CRLR with RAMP1 forms the CGRP type 1 receptor, with weak AM specificity.15,16 In animal studies, inhibition of AM during the period of alveolar development caused arrested lung capillary and alveolar growth, associated with decreased lung vascular endothelial growth factor-A and endothelial nitric oxide synthase expression. 13 Conversely, in experimental oxygen-induced lung injury in newborn rats, AM administration attenuated the arrest in lung vascular and alveolar growth; and this was associated with decreased lung apoptosis. Adrenomedullin also prevented pulmonary hypertension in this model. 13
In a surfactant-depleted piglet model with pulmonary hypertension, the nebulization of AM (50 ng/kg per minute for 2 hours) resulted in a sustained reduction in mean pulmonary arterial pressure, whereas systemic blood pressure decreased minimally. No changes were seen in systemic vascular resistance, heart rate, or cardiac index. 17 Induction of pulmonary hypertension in adult rats (approximately 8 weeks old) by a hypobaric hypoxic environment resulted in markedly increased AM concentrations in both plasma and heart tissue. 18 These studies support the hypothesis that upregulation of AM plays a protective role in the pathogenesis of pulmonary hypertension and that exogenous AM administration could be a therapeutic approach. The role of AM in the developing human lung is not known; however, animal data suggest that AM contributes to lung angiogenesis and alveolar development. 13 We postulated that AM is expressed in the human lung during fetal development and that such expression would increase proportionally to fetal lung and vascular development. To explore this hypothesis, we measured the expression of AM and its receptor complexes, CRLR/RAMP2 (AM1 receptor) and CRLR/RAMP3 (AM2 receptor), in human fetal lung from 10 to 24 weeks of gestation. We determined protein expression of AM using Western blot analysis and performed immunohistochemistry staining of fetal lung tissue.
Materials and Methods
Tissue Collection
Human fetal lung tissue was obtained after elective termination between 10 and 24 weeks of gestation (courtesy of Dr. Curtis Boyd from Southwestern Women's Options, Albuquerque, NM). Gestational age was estimated based on fetal foot length. Sex could not be determined in most samples, and this information was not collected. The fetal lungs were isolated and immediately processed for RNA and protein extraction. The lung tissue was fixed, cryoprotected, snap frozen in liquid nitrogen, and stored at −80°C until further use.
Rna Extraction and Quantitative Real-Time Reverse-Transcription Polymerase Chain Reaction
The lung tissue was homogenized in phosphate-buffered saline (PBS), then processed using TriZol (Invitrogen, Carlsbad, CA) to extract total RNA and quantified by spectrophotometry. The RNA was reverse transcribed using the complementary DNA Archive kit (Applied Biosystems, Foster City, CA). Expression levels of a subset of genes were measured by quantitative real-time reverse-transcription polymerase chain reaction performed with TaqMan assay reagents according to manufacturer's instructions in a PRISM 7500 Fast Thermocycler (Applied Biosystems) using predesigned primers and probes (glyceraldehyde 3 phosphate dehydrogenase [GAPDH] Hs99999905 m1, AM Hs00181605 m1, RAMP Hs00195288 m1, RAMP2 Hs00237194 m1, RAMP3 Hs00389131 m1, and CRLR Hs00907738 m1). To normalize targets to total RNA loaded, human GAPDH (hGAPDH) (Applied Biosystems) was used as endogenous control.13,19 The ΔΔ Cycle threshold method was used to determine the relative levels of the selected genes and the mRNA expression of target genes in each sample was normalized to the values of hGAPDH mRNA in the same sample. Human GAPDH was the more stable of the housekeeping genes tested, and there is evidence of absence of zregulation of hGAPDH under hypoxic conditions. 20
Western Blot Analysis and Determination of AM Peptide
Lung tissue was dissected and homogenized in radioimmunoprecipitation assay buffer (1% nonidet P40, 0.25% sodium deoxycholate, 0.1% sodium dodecyl sulfate, and 1-mmol/L dithiothreitol), plus 1-mmol/L phenylmethylsulfonyl fluoride and phosphatase inhibitor cocktail (Sigma-Aldrich, St. Louis, MO), incubated in ice for 25 minutes and spun at 12,000g for 20 minutes. The total protein content in the supernatant was measured with bicinchoninic acid protein assay reagent (Thermo Fisher Scientific, Rockford, IL). Thirty micrograms of protein were resolved by a 12% sodium dodecyl sulfate polyacrylamide gel electrophoresis gel and electrophoretically transferred to Immun-blot polyvinylidene difluoride transfer membranes (Bio-Rad Laboratories, Hercules, CA). After blocking in 5% nonfat dry milk, membranes were reacted with rabbit anti-AM antibodies (Abcam, Cambridge, MA) overnight at 4°C and then were incubated with horseradish peroxidase–conjugated goat antirabbit IgG (Invitrogen, Grand Island, NY) for 1.5 hours at room temperature. The protein bands were detected using SuperSignal West Femto maximum sensitivity substrate (Thermo Scientific, Rockford, IL) and visualized on a Kodak image station 4000 mm (Eastman Kodak, Rochester, NY). A rabbit anti–β-actin antibody (Abcam) was used as a control for equal loading. The levels of protein were quantified by image analysis using the ImageJ software (National Institutes of Health, Bethesda, MD), and densitometric data were expressed as the ratio of protein to β-actin. 21
Immunohistochemistry Staining
Fresh lung tissue was fixed in 4% formaldehyde and embedded in Tissue-Tek optimal cutting temperature compound (Sakura Firetek USA Inc, Torrance, CA) using a cryomold. Sections of 10 μm were collected and permeabilized with 0.2% Triton X-100 in PBS for 10 minutes at room temperature. The sections were blocked with 3% donkey serum in PBS for 1 hour and incubated overnight with rabbit anti-AM antibodies (Santa Cruz Biotechnology Inc, Santa Cruz, CA) and mouse anti-SM22 alpha antibody (Abcam) at 4°C. Alexa Fluor 647–labeled antirabbit and Alexa Fluor 568–labeled antimouse secondary antibodies (Invitrogen) were used to detect the primary antibodies. 4’-6-Diamidino-2-phenylindole (Molecular Probes, Eugene, OR) was used to label cell nuclei. Samples were visualized with fluorescence microscopy. As negative controls, we omitted the primary antibodies and determined nonspecific binding of the secondary antibodies. The levels of fluorescence intensity were quantified by image analysis using ImageJ software (National Institutes of Health).
Human Subjects
The Human Research Review Committee at the University of New Mexico reviewed and approved the research protocol Human Research Review Committee No. 10-509. This study involved the use of discarded medical samples. There were no identifying aspects of the tissue samples beyond the date of collection, and those who read the study results will not have the capacity to identify a particular study subject. We did not have contact with the women undergoing the termination procedure, did not have bearing on the decision concerning pregnancy termination, and were not involved in performing the procedure. The samples collected would otherwise be disposed of as medical waste. There were no risks to human subjects in this study.
Statistical Analysis
The relationship between protein and mRNA expression concentrations and gestational age were analyzed using linear regression; P < 0.05 was considered significant. In addition, the samples used for gene expression of AM and its receptors were divided by weeks of gestation in 5 groups (10–12, 13–14, 15–16, 17–18, and >19 weeks of gestation), and one-way analysis of variance (ANOVA) was used to analyze the relationship between gestational age and the mRNA expression; P < 0.05 was considered significant. For post hoc analysis, the Tukey range test was used. Similarly, one-way ANOVA followed by the Tukey range test was used to analyze the Western blot and the immunohistochemistry results.
Results
Gene Expression of AM and the AM Receptors in Fetal Lungs
The mRNA encoding for AM, CRLR, RAMP2, and RAMP3 were detected in fetal lung tissue from 10 to 24 weeks’ gestational age. The AM mRNA expression in the fetal lungs increased with increasing gestational age (n, 65 samples; R2, 0.2020; SE, 0.0052; P < 0.0002; Fig. 1A). One-way ANOVA showed increased expression of AM mRNA with increasing gestational age (n, 5 groups; R2, 0.7329; P < 0.0001), and post hoc analysis showed a significant increase (P < 0.05) between early gestation (10–12 weeks) and later gestation (17–18 weeks and more). The CRLR mRNA expression increased with increasing gestational age (n, 65 samples; R2, 0.3720; SE, 0.01179; P < 0.0001; Fig. 1B). One-way ANOVA showed increased expression of CRLR mRNA with increasing gestational age (n, 5 groups; R2, 0.4870; P < 0.0032) with post hoc analysis showing a significant increase between the samples at 10 to 12 weeks of gestation and the samples at 17 to 18 weeks and more (P < 0.05). The RAMP2 mRNA expression also increased (n, 67 samples; R2, 0.1931; SE, 0.01048; P = 0.0002; Fig. 1C). One-way ANOVA showed increased expression of RAMP2 mRNA with increasing gestational age (n, 5 groups; R2, 0.6198; P < 0.0061), post hoc analysis showed a significant increase when comparing samples at 10 to 12 weeks of gestation with those at 17 to18 weeks and more (P < 0.05). The RAMP3 mRNA expression decreased with advancing gestational age (n, 67 samples; R2, 0.3696; SE, 0.08023; P < 0.0001; Fig. 1D). One-way ANOVA showed decreased expression of RAMP3 mRNA with increasing gestational age (n, 5 groups; R2, 0.5404; P < 0.0001), with post hoc analysis also showing a significant decrease when comparing samples at 10 to 12 weeks of gestation with those at 17 to 18 weeks and more (P < 0.05).
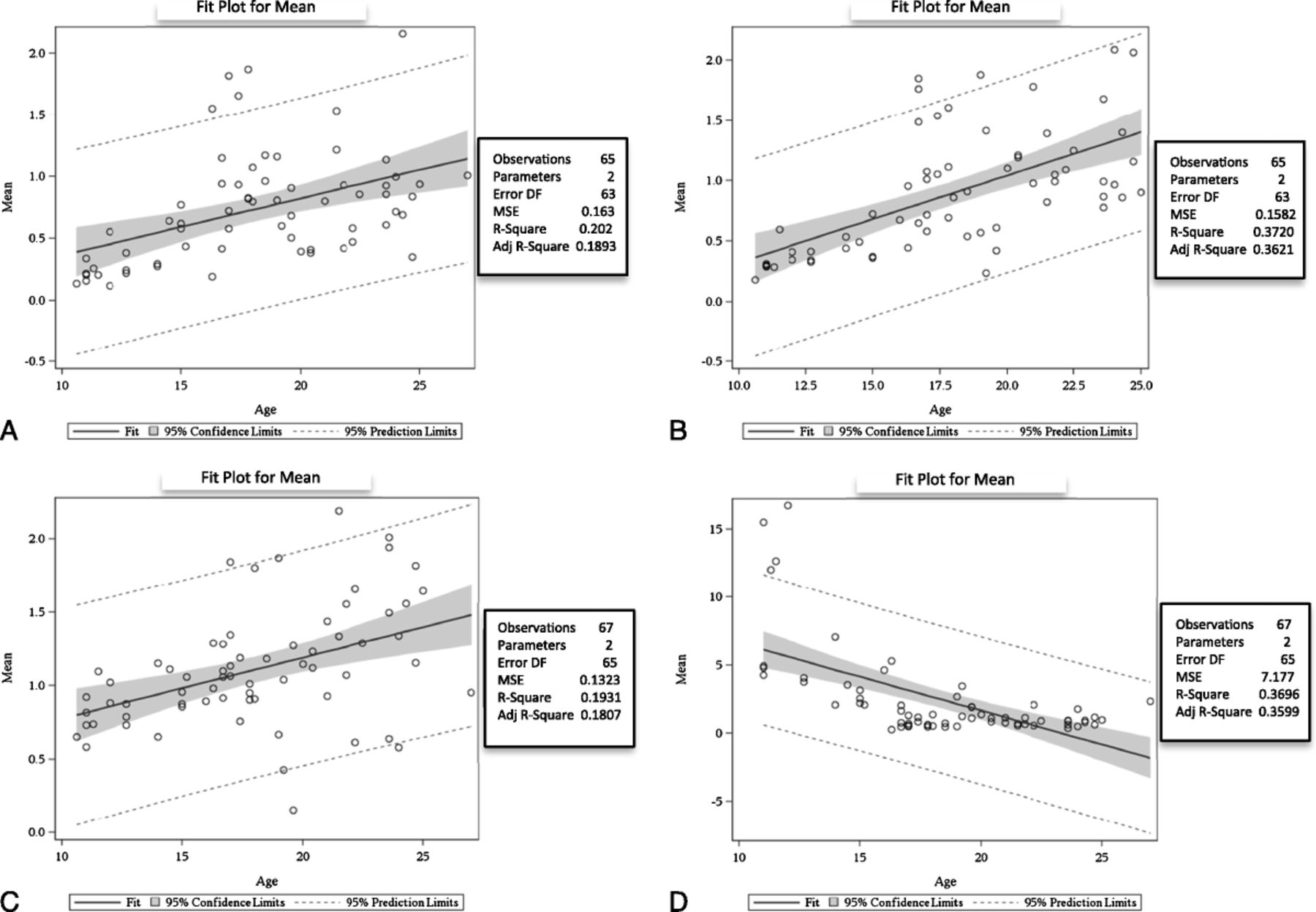
Gene expression of AM, CRLR, RAMP2, and RAMP3 measured by reverse-transcription polymerase chain reaction (RT-PCR). A, Adrenomedullin mRNA expression measured by RT-PCR: fit plot for a mean of 65 human fetal lung samples showing increased expression with increasing gestational age (n, 65 samples; R2, 0.2020; SE, 0.0052; P < 0.0002). B, Expression of CRLR mRNA measured by RT-PCR: fit plot for a mean of 65 human fetal lung samples showing increased expression with increasing gestational age (n, 65 samples; R2, 0.3720; SE, 0.01179; P < 0.0001). C, Expression of RAMP2 mRNA measured by RT-PCR: fit plot for a mean of 67 human fetal lung samples showing increased expression with increasing gestational age (n, 67 samples; R2, 0.1931; SE, 0.01048; P < 0.0002). D, Expression of RAMP3 mRNA measured by RT-PCR: fit plot for a mean of 67 human fetal lung samples showing decreased expression with advancing gestational age (n, 67 samples; R2, 0.3696; SE, 0.08023; P < 0.0001).
Adrenomedullin Protein Expression in Fetal Lungs
Fetal lung AM protein expression increased with increasing gestational age. We measured AM expression in fetal lung tissue at 10.6, 11.5, 13, 14, 16.7, 17.8, 19.2, and 21 weeks of gestation using Western blot. The relative expression of AM increased with advancing gestational age (R2, 0.9063; P < 0.01; Fig. 2). One-way ANOVA showed increased protein expression with increasing gestational age (n, 8; R2, 0.9852; P < 0.0001), with post hoc analysis showing significant increase between 10.6 weeks of gestation and 21 weeks (P < 0.05).
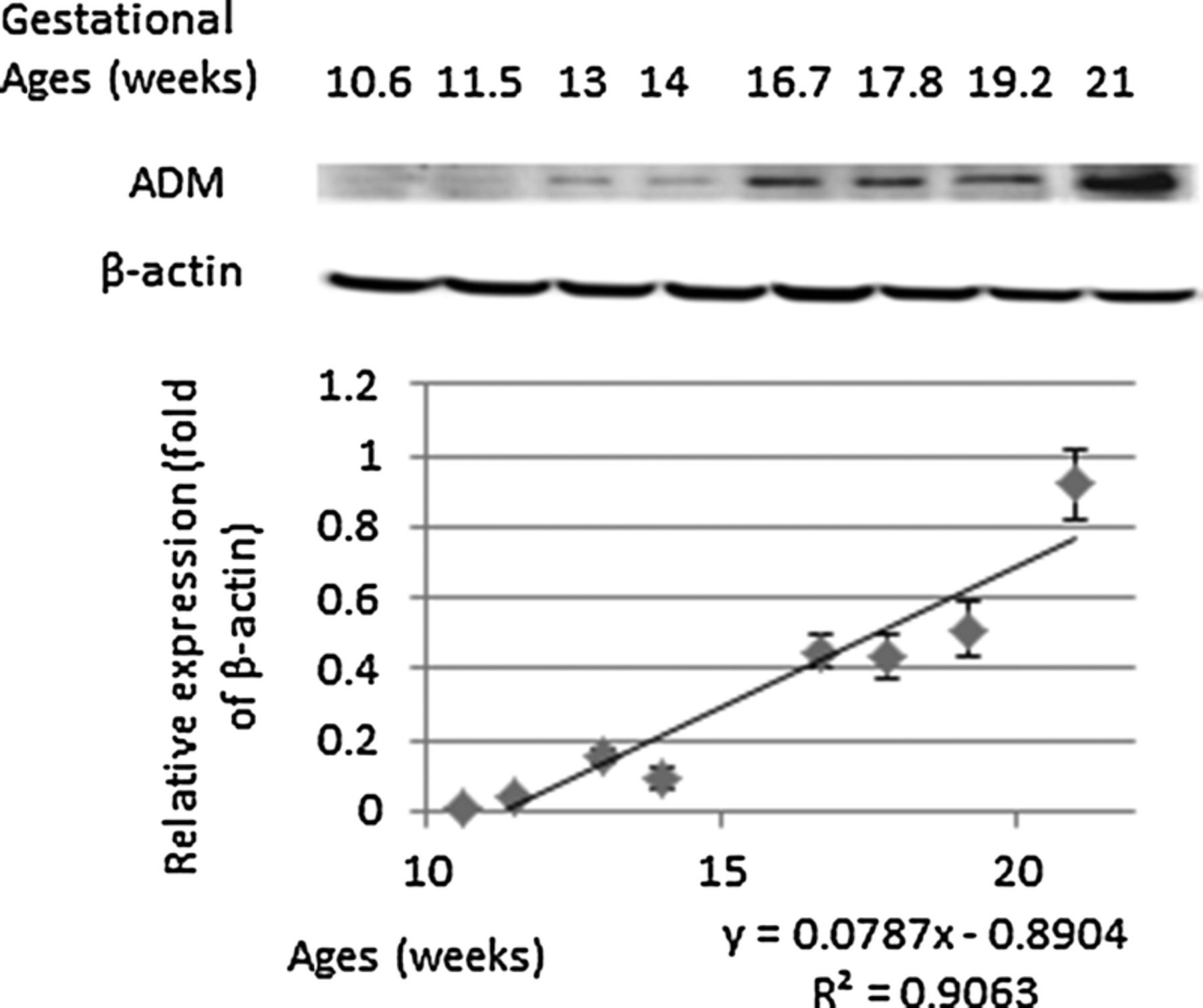
Protein expression of AM in human fetal lung measured by Western blot analysis. The relative expression of AM as fold of β-actin increased with advancing gestational age (R2, 0.9063; P < 0.01).
Immunohistochemical Analyses of Fetal Lung Tissue
Immunohistochemical analyses were performed in fetal lung tissues of fetuses of 10.5, 10.8, 12, 15, 17, and 18 weeks of gestational age. We observed immunoreactivity against AM at all gestational ages, localized in lung mesenchyme and the mesenchymal smooth muscle cells (red arrows in Figs. 3, 4 and stained green in Fig. 5). The immunoreactivity against AM, measured as mean of fluorescence intensity, increased with advancing gestational age (n, 6; R2, 0.3081; P < 0.05; Fig. 3). One-way ANOVA showed increased fluorescence intensity with increasing gestational age (n, 6; R2, 0.8199; P < 0.001), with post hoc analysis showing significant increase between 10.5 weeks of gestation and 18 weeks and more (P < 0.05). Costaining with anti-SM22 alpha is shown in Figure 4, further delineating mesenchymal localization of AM. Staining with 4’-6-diamidino-2-phenylindole for nuclei labeling is shown in Figure 5, further confirming the absence of immunoreactivity against AM in the tubular epithelium.
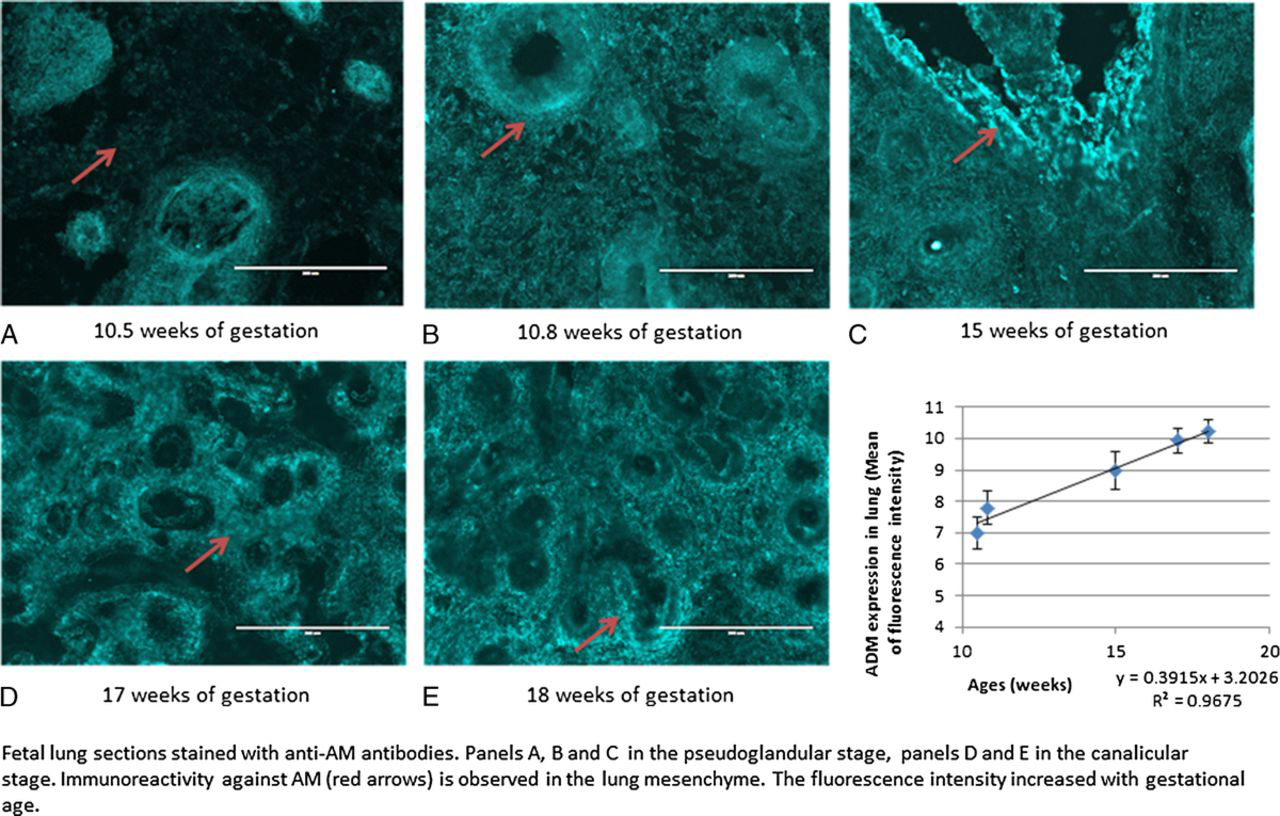
Immunohistochemical analyses of sections of human fetal lung tissue, shown at original magnification ×20. Immunoreactivity against AM was observed in fetal lung tissue of fetuses of 10.5, 10.8, 15, 17, and 18 weeks of gestational age and the immunoreactivity, measured as mean of fluorescence intensity using imageJ software, increased with advancing gestational age (R2, 0.3081; P < 0.05).
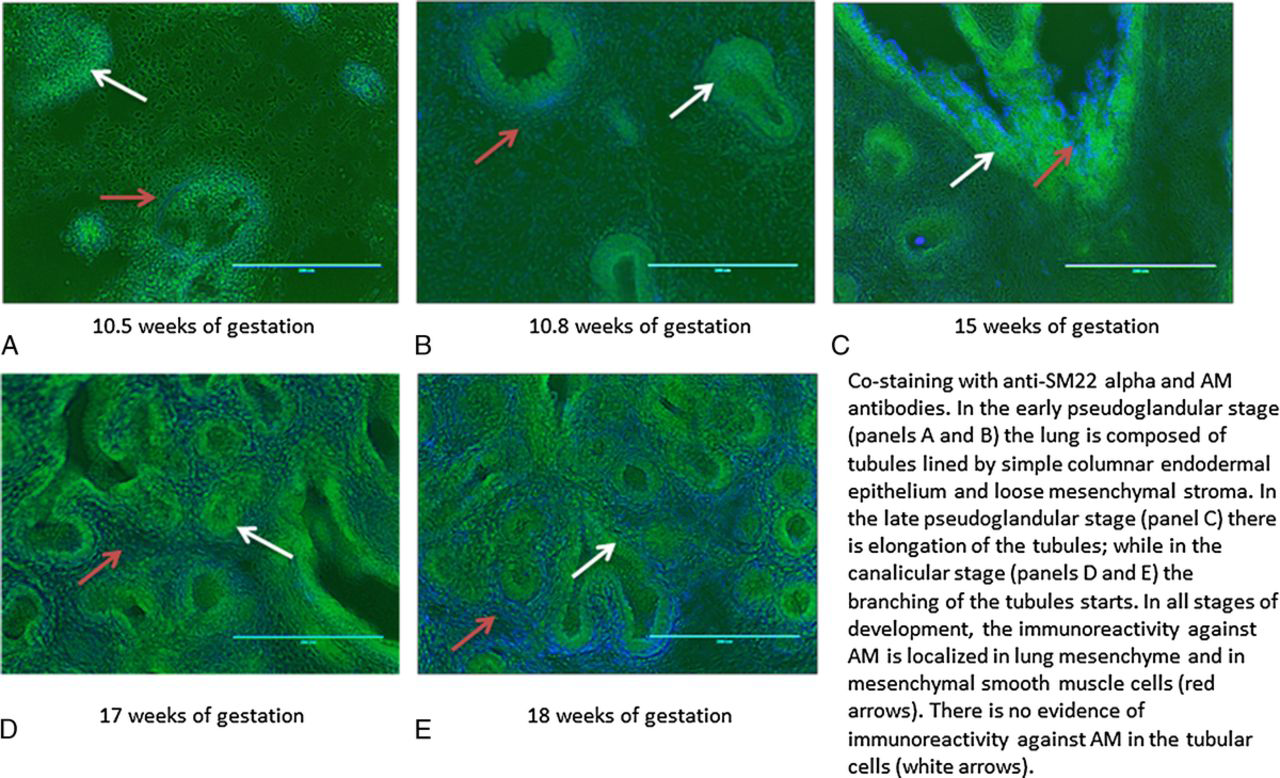
Costaining of sections of human fetal lung tissue at 10.5, 10.8, 15, 17 and 18 weeks of gestational age with anti-SM22 alpha antibody and AM antibody, shown at original magnification ×20. Immunoreactivity against AM was localized in the lung mesenchyme and in mesenchymal smooth muscle cells (red arrows). The lung tubules are lined with simple columnar endodermal epithelium (white arrows).
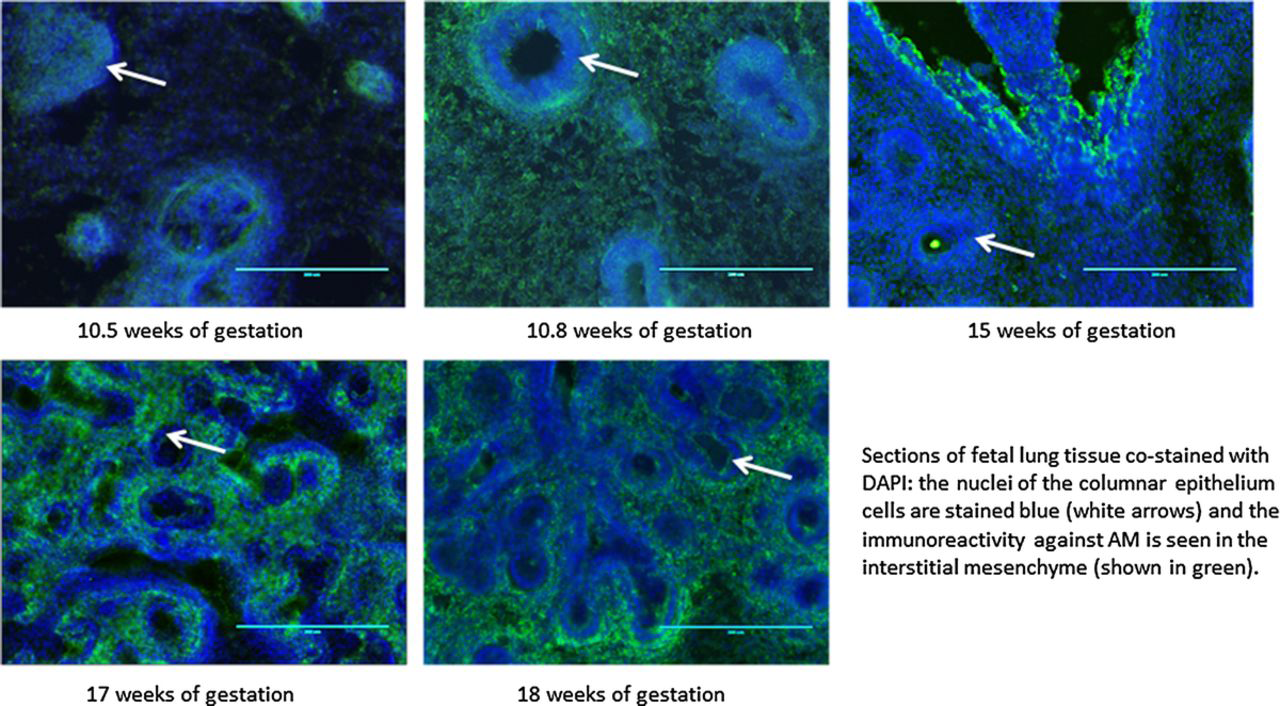
Immunohistochemical analyses of sections of human fetal lung tissue at 10.5, 10.8, 15, 17, and 18 weeks of gestational age labeled with 4’-6-diamidino-2-phenylindole and stained with AM antibody, shown at original magnification ×20. The nuclei of the columnar epithelial cells are stained blue (white arrows), and immunoreactivity against AM is seen in interstitial mesenchyme and mesenchymal smooth muscle cells (shown in green).
Discussion
In the fetus, the placenta functions as the organ for gas exchange, and the fetal lungs receive only 5% to 10% of the combined cardiac output. As a result, pulmonary hypertension is normal during fetal life, and the lungs develop in a relative hypoxic environment. Despite the fact that pulmonary vascular surface area increases with fetal lung growth, pulmonary vascular resistance increases with gestational age, suggesting an increase in vascular tone. 22
Although the role of AM in fetal lung vascular development is not known, we speculate that the elevated pulmonary vascular pressures in the fetus may lead to an increase in AM expression. In support of this speculation, there is evidence of a marked increase in AM expression in rats with hypoxia-induced pulmonary hypertension, 18 and we are showing immunoreactivity against AM in the human fetal lung as early as 10 weeks of gestational age with increased immunoreactivity as gestation progresses. This rise in immunoreactivity correlates with increased protein and mRNA expression. In addition, data from previous investigators have shown the presence of immunoreactivity against AM in human fetal lung tissue starting at 18 weeks of gestation with progressive increase during fetal life. 23
Adrenomedullin is a vasoactive peptide with multiple biological actions in vitro and in vivo: it induces angiogenesis, 24 and it is a protective factor for blood vessels. 25 It has been characterized as a potent long-lasting systemic vasodilator after intravenous administration, 12 and it has been shown to produce a sustained decrease in pulmonary artery pressures after continuous inhalation, both in animal studies 17 and in patients with idiopathic pulmonary arterial hypertension. 26 Adrenomedullin might play a role in fetal lung vascular development, and there is evidence that AM and its receptor components are highly expressed in human reproductive tissues, including fetal membranes and placenta. 27
The expression of the AM1 receptor increased with advancing gestational age; this rising expression of both CGRP and the AM1 receptors in the canalicular period of lung development, which coincides with the completion of the preacinar vascular patterning and multiplication of capillaries, supports a role for AM in human vascular development. This is consistent with findings in murine models, where AM, acting on the AM1 receptor, facilitates the development of the murine fetal cardiovascular system.16,28 Normal fetal lung development occurs in relatively hypoxic conditions, and there is evidence of increased AM production and up-regulation of the AM1 receptor during hypoxia. 29 This finding would also explain the increased expression of AM and the AM1 receptor observed by our group.
The expression of the AM2 receptor decreased with advancing gestational age. This finding could be explained based on the available evidence in rat models of the separate biological functions of the AM1 and AM2 receptors in disease states; furthermore, such differences seem to be dependent on the receptors’ particular location and pathophysiological conditions. 16 In the normal mouse, there is increased expression of CRLR and RAMP2 in the lungs with very little expression of RAMP3, similar to our findings in human fetal lungs. 30 Sepsis induces higher circulating levels of AM in humans 31 and in mice 30 ; and in the murine sepsis model, there is marked decrease in the CRLR and RAMP2 mRNA expressions in the lung, with a significant increase in the RAMP3 gene expression.30,32
Our study has several limitations. We could only obtain fetal lung tissue from 10 to 24 weeks of gestational age, which corresponds to the pseudoglandular and canalicular periods of lung development. At this early stage of development, the conducting airways are completed but alveolar epithelial cell differentiation is just beginning. Also in this period, there is exponential growth and multiplication of pulmonary capillaries with the formation of the primitive blood-air barrier. 33 Adrenomedullin expression could be different, as gestational age further advances and alveolar development begins. The nature of the samples posed particular challenges: we could not process the tissue before 1 to 2 hours after termination and we did not have access to clinical and demographic data about the women undergoing the procedures. However, our data are consistent with the results obtained in animal models16,29,30 and in humans 24 by different investigators. The RAMP family interacts with other receptors of the calcitonin peptide family besides CRLR 34 and as such, the expression levels of RAMP2 and RAMP3 in fetal lung observed by our group may be related to their activity with other G protein–coupled receptors.
In conclusion, we found that pulmonary mRNA AM expression increased in the human fetal lung from 10 to 24 weeks of gestation, with a concomitant increase in AM immunoreactivity. In addition, we found significant differences in the trajectories of expression of the AM receptor components RAMP2 and RAMP3. Further studies are required to elucidate the mechanisms by which AM contributes to pulmonary vascular and alveolar development.
Footnotes
Acknowledgments
The authors thank Dr. Curtis Boyd and the staff at Southwestern Women's Options, as well as Suzanne McConaghy for technical assistance.