
Abstract
Congenital diaphragmatic hernia (CDH) is associated with lung hypoplasia and pulmonary hypertension and has high morbidity and mortality rates. The cause and pathophysiology of CDH are not fully understood. However, impaired angiogenesis appears to play an important role in the pathophysiology of CDH. Therefore, we examined different components of an important pathway in angiogenesis: hypoxia-inducible factors (HIFs); HIF regulators von Hippel–Lindau (VHL) and prolyl 3-hydroxylase (PHD3); and HIF target genes vascular endothelial growth factor A (VEGF-A) and vascular endothelial growth factor receptor 2 (VEGFR-2). Quantitative polymerase chain reaction of lung tissue showed a significantly decreased expression of VEGF-A mRNA in the alveolar stage of lung development in CDH patients compared with matched control patients. In the canalicular stage, no differences for VEGF-A were seen between the lungs of CDH patients and those of control patients. Other components of angiogenesis (VHL, HIF-1α,HIF-2α, HIF-3α, VEGFR-2 mRNA, PHD3 protein) that were analyzed showed no differences in expression between CDH and control patients, independent of the developmental stage. A lower expression of VEGF mRNA in CDH patients in the alveolar stage, possibly as a result of downregulation of HIF-2α might indicate a role for these factors in the pathophysiology of CDH.
Keywords
INTRODUCTION
The probability of survival in congenital diaphragmatic hernia (CDH) is mainly determined by the severity of lung hypoplasia and its associated pulmonary hypertension [1]. The lungs in CDH patients are characterized by maldevelopment of airways, impaired vascular development, and a disordered process of postnatal vascular remodeling [2]. Pulmonary vessel morphometry in CDH shows a decreased volume of the pulmonary vascular bed, an increased adventitial and medial thickness, and muscularized distal pulmonary arterioles [3,4]. Newborns with CDH have a delayed transition to postnatal circulation due to respiratory failure. In a number of cases, they develop pulmonary hypertension resulting in major treatment challenges and eventually death.
The most potent inducer of angiogenesis is vascular endothelial growth factor A (VEGF-A) [5]. Furthermore, VEGF-A is abundantly expressed in parenchyma of the developing lung [6]. Also during postnatal alveolarization of the lung, angiogenesis in general, and VEGF-A in particular, has an important role [7]. The VEGF-A receptor (VEGFR-2) mediates the major part of VEGF-A–driven vascular growth [5]. The transcription of both VEGF-A and VEGFR-2 is mainly regulated by hypoxia-inducible factor (HIF) [8].
The HIF transcriptional complex is a heterodimer composed of 1 of the 3 alpha subunits (HIF-1α, HIF-2α, HIF-3α) and a constitutive nuclear protein, the beta subunit (HIF-1β). Aside from common target genes, HIF-1α and HIF-2α also regulate unique sets of genes that play important roles during development [8]. Furthermore, HIF-3α appears to be reciprocally regulated in the response to hypoxia, resulting in negative feedback [9]. Hypoxia-inducible factor 2α has an important role in the remodeling of the primary vascular network [10].
Posttranscriptional regulation of the HIFα isoforms occurs through hydroxylation of specific proline residues by prolyl 4-hydroxylases (PHDs), which are active at hypoxic conditions. At low oxygen concentration, the hydroxylated HIFα proteins associate with the von Hippel–Lindau protein (pVHL) and are subsequently targeted for proteosomal degradation (Fig. 1). Prolyl 4-hydroxylase is expressed in early human lung development, and inhibition of PHDs prevents the degradation of HIFα at normoxia, which led to amelioration of the pathophysiology in an animal model with brochopulmonary dysplasia [11,12]. This indicates that PHDs are potential therapeutic targets.
In an earlier study, we showed that the expression of HIF-2α and VEGF in the human lung correlates with increasing gestational age, underscoring their importance in pulmonary development [13]. However, little is known about the expression of the various HIFs and their target genes in CDH patients. We hypothesize that these factors might be involved in the early (canalicular) and late transitional (alveolar) stages of the pathophysiology of CDH. Therefore, we analyzed with real-time polymerase chain reaction (PCR) the expression of HIF-1α, HIF-2α,HIF-3α, and VHL and 2 targets of HIF, VEFG-A and VEGFR-2, in human control and CDH lung samples in the canalicular and alveolar stage. Furthermore, the location and expression of PHD3, the predominant posttranscriptional regulator of HIF-2α, are evaluated by immunohistochemistry.
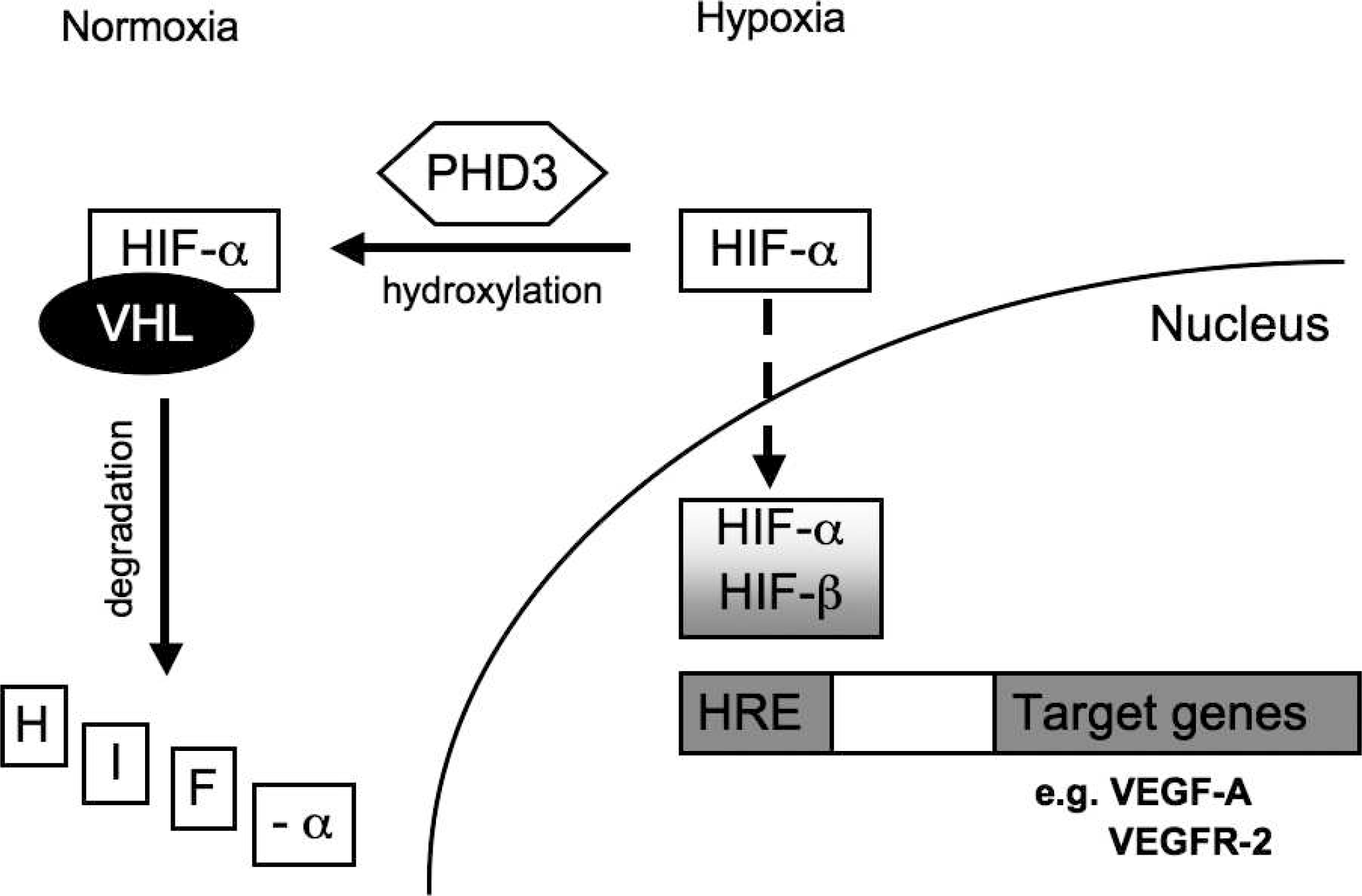
Overview of the regulation of hypoxia-inducible factor and its activation of target genes involved in angiogenesis.
MATERIALS AND METHODS
Human lung tissue was retrieved from the archives of the Department of Pathology of the Erasmus MC-University Medical Center in Rotterdam, The Netherlands, after approval by the Erasmus MC Medical Ethical Committee. From formalin-fixed and paraffin-embedded lung tissue of a series of CDH patients, we selected 16 CDH patients who died within 24 hours after birth or from termination of pregnancy. Patients were grouped (n = 10 alveolar stage and n = 6 canalicular stage) based on their gestational age and histological appearance of their lung tissue. Neonates deceased from nonpulmonary causes served as controls and were matched for gestational age (n = 5 canalicular stage and n = 6 alveolar stage), duration of postnatal survival, and spontaneous breathing and/or ventilation (Table 1). Snap-frozen lung tissue of 7 CDH patients (n = 3 canalicular stage and n = 4 alveolar stage) was used for RNA isolation.
Real-time PCR
Total RNA was extracted from frozen lung tissue (3060 mg) using Trizol reagent (Life Technologies, Rockville, MD, USA) following the manufacturer's instructions. Complementary DNA was synthesized using 1 μg of total RNA by reverse transcription and used in real-time PCR as previously described [13]. A quantitative PCR core kit for SYBR Green I-assay (Eurogentech, Seraing, Belgium) and gene-specific primers for target genes VHL, HIF-1α, HIF-2α, HIF-3α, VEGF-A,and VEGFR-2 were used for real-time PCR. All reactions were performed in triplicate and controlled using standard nontemplate as a negative control and an identical positive control. The PCR results are shown as 2−ΔΔt using POLR2A as the endogenous reference control and a sample of 13.5 weeks of gestation as a “calibrator,” which was arbitrarily set at 100% (arbitrary value = 1). Because we used the same calibrator sample and performed the experiments under the same conditions, comparisons can be made between CDH patients and control patients who have previously been discussed in the literature. [13].
Characteristics of congenital diaphragmatic hernia (CDH) and control patients used for immunohistochemistry

Immunohistochemistry
Immunohistochemistry was performed on 4-μm sections of paraffin-embedded lung tissue, mounted on coated slides (Starfrost, Berlin, Germany) according to standard protocol, using the EnVision detection system (DakoCytomation, Glostrup, Denmark). [13] The primary antibody was a polyclonal rabbit anti-PHD3 (1:2000, Novus Biologicals, Littleton, CO, USA). Antigen retrieval consisted of boiling for 15 minutes in Tris-EDTA buffer (pH 9). Negative controls were prepared by omission of the primary antibody. Two independent observers (I.WJ.M.vdH. and P.vdV.) examined the slides for PHD staining and scored each sample as strong (3), clear (2), weak but visible (1), or negative (0).
Statistical analysis
For relative mRNA expression of the HIF pathway, differences between CDH and control patients were analyzed with an unpaired t-test. The chi-square test was used for proportions of PHD3 immunostaining scores of CDH versus control patients. A P value of 0.05 was considered statistically significant.
RESULTS
Real-time PCR
Previously, we have shown that VHL, HIF-1α, HIF-2α, and HIF-3α and their target genes VEGF-A and VEGFR-2 are expressed during the canalicular and alveolar stages of lung development [13]. In lung samples from CDH patients, all of the abovementioned factors were expressed at both stages of development (Fig. 2). Of all the expressed factors, HIF-2α and VEGF-A showed the highest relative expression in the canalicular and alveolar stages.
VEGF-A expression was significantly (P = 0.000) lower in CDH patients than in control patients in the alveolar stage. VEGF-A expression in the canalicular stage was similar in CDH and control patients. Expression of VHL, HIF1-α, HIF-2α, HIF-3α, and VEGFR2 showed no difference between CDH and control patients or in the alveolar or canalicular stage of lung development.
Immunohistochemistry
For localization and expression of PHD3, we used immunohistochemistry. Prolyl 3-hydroxylase showed cytoplasmatic expression, which was present in the bronchial epithelium and endothelium of arterioles. Moreover, PHD3 was expressed in most cell types of the lung, except for the arterial adventitia (Fig. 3). All cases showed strong staining in vessels of all sizes, as well as bronchial structures of the lung. We did not observe differences between samples from CDH and control patients in the canalicular or alveolar stages.
DISCUSSION
In this study, we evaluated the mRNA expression of the various HIFs, their target genes VEGF-A and VEGFR2, and the regulatory protein PHD in human CDH lungs and found that the abovementioned factors were expressed in the canalicular stage as well as in the alveolar stage. For the first time, a significantly lower mRNA expression for VEGF-A was seen in CDH patients in the alveolar stage. Due to the limitation of human patient material available, we have only been able to perform measurements at specific time points in development. Although our material is unique, our conclusions relate only to these time points and not to other stages of lung development.
Interestingly, VEGF mRNA expression was significantly decreased in CDH patients compared with control patients, who were treated with similar ventilation strategies. In this study, we used lung tissue of patients who died within 24 hours after birth to look specifically at the transitional stage. In the transition to extrauterine life, pulmonary arteries must adapt to the gas exchanging function and increased blood flow [14]. Therefore, the capillary network must be recruited, resistance arteries need to dilate, and the pulmonary vasculature must remodel. Vascular endothelial growth factor is necessary for directing capillary sprouting and governing remodeling of the existing capillary network [15]. Inhibition of VEGF in this early postnatal period leads to impaired vascular maturation and subsequent arrest of alveolarization [16]. These features are seen in lung morphometry of deceased CDH patients [17]. Moreover, several animal studies show that decreased VEGF mRNA expression is associated with pulmonary hypertension, which is very common in CDH patients [18]. Conversely, similar and increased VEGF protein expression has been reported by our group in deceased CDH patients [19,20]. There are several reasons for the differences between these studies. The VEGF protein expression in these studies was measured with semiquantitative immunohistochemistry, and the antibody used detects more VEGF-A isoforms than the primer set used in this study. Furthermore, the VEGF mRNA was calculated in whole lung homogenate, whereas VEGF protein expression was counted specifically in the smaller capillaries.
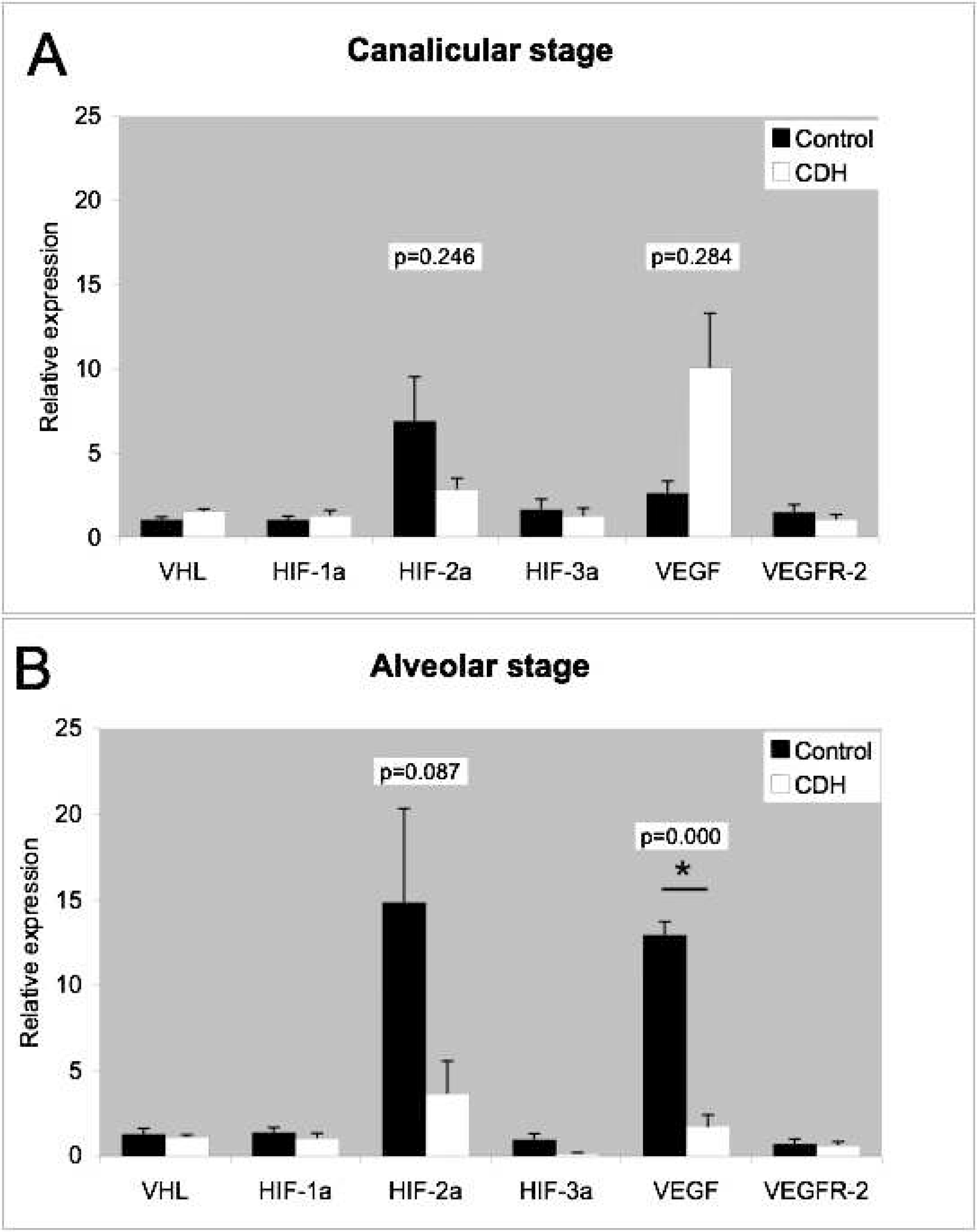
Relative mRNA expression of von Hippel-Lindau (VHL), hypoxia-inducible factors (HIF-1α, HIF-2α, and HIF-3α), and vascular endothelial growth factor (VEGF) and its receptor (VEGFR-2) measured by quantitative real-time polymerase chain reaction in the lung tissue of congenital diaphragmatic hernia (CDH) patients and matched controls in (
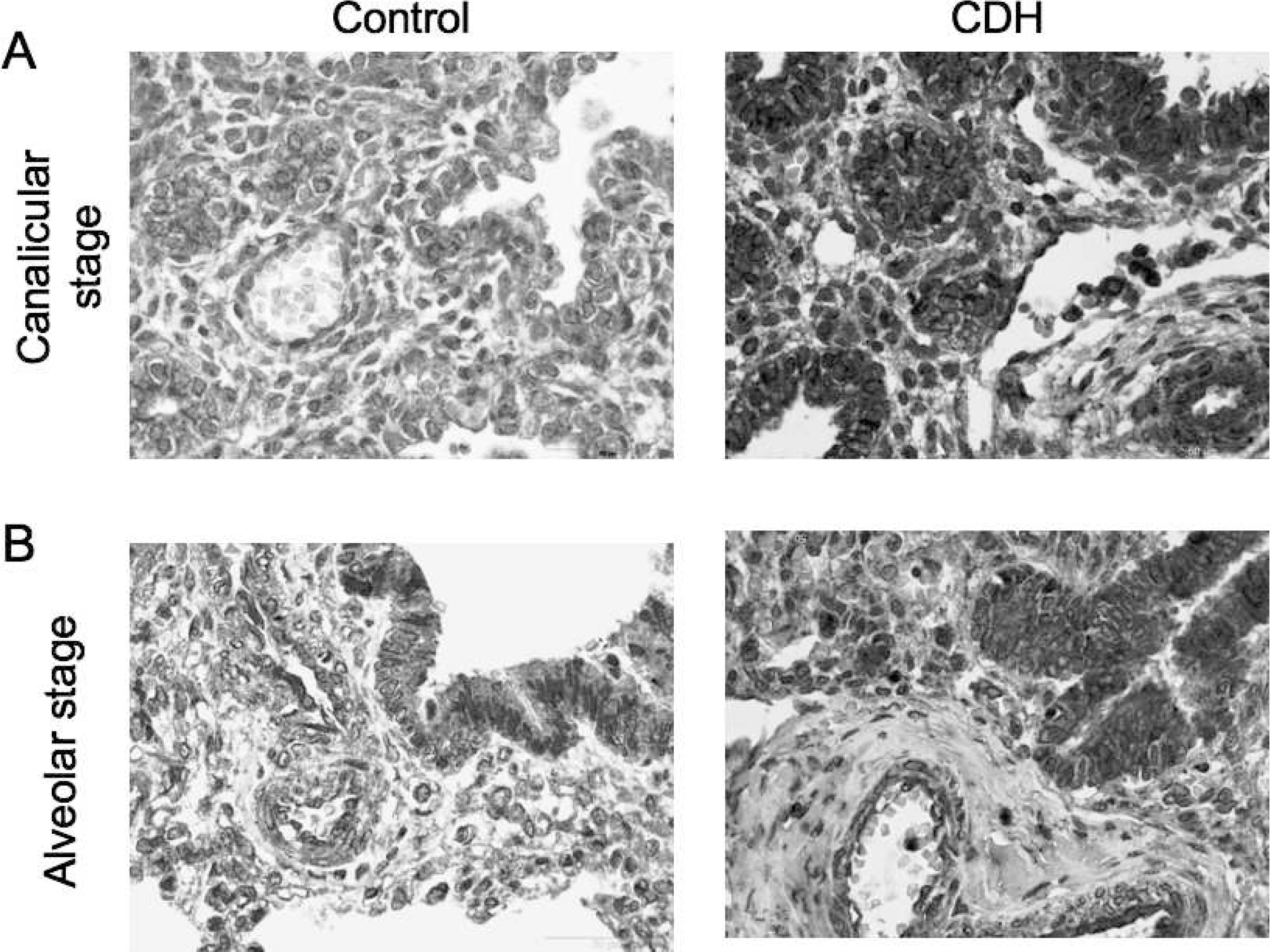
Immunostaining with anti–prolyl 3-hydroxylase (PHD3) of congenital diaphragmatic hernia (CDH) and control lung tissue in the (
Our results demonstrated similar mRNA expression of VEGF in CDH patients compared with control patients in the canalicular stage. From several animal studies, there are contradicting results: some report decreased expression of VEGF in canalicular stage lungs of nitrofen-induced CDH in mice and rats [21,22], whereas others report increased or no difference in VEGF expression in CDH rats during gestation [23,24].
Expression of VEGF receptors has been suggested to play a role in the variation in VEGF function. However, no change in the amount of VEGFR-2 mRNA was seen during lung development or between control and CDH lung tissue. Moreover, in the nitrofen-induced CDH rat model, no change in VEGFR-2 was observed [25]. Although VEGF and VEGFR-2 are both transcription targets of HIF, there is no evidence for a reciprocal regulation of both factors, which could explain an increased expression of VEGF without an increase in VEGFR-2 expression.
Although there appeared to be a trend for HIF-2a mRNA expression to be lower in CDH cases compared with controls in the alveolar stage of lung development, no significance could be reached. Therefore, we cannot conclude that HIF-2α has a role in the pathophysiology of CDH. In mice, HIF-2α in the lungs increases at the end of gestation and stays stable after birth, whereas HIF-1 a is postnatally downregulated [26]. This suggests a potential role for HIF-2α during the transitional period of the fetus to adapt to life in an oxygen-rich environment. Furthermore, HIF-2α is required for the remodeling of the primary vascular network into a mature pattern [10] and is closely related to VEGF expression [26].
In the canalicular stage, the expression of HIF-2α mRNA in CDH patients’ lungs was comparable to that in control patients’ lungs. Moreover, loss of HIF-2α in HIF-2α−/- mice did not affect lung development during the canalicular stage [27]. The function of HIF-2α during early lung development and the consequences of altered expression remain elusive.
The expression of 2 HIF-2α-related genes, HIF-1/α and HIF-3α, did not differ between control and CDH patients, nor in the canalicular or alveolar stage. In particular, HIF-1α is mainly posttranslationally regulated; its protein expression is increased by hypoxia [8]. Previously, we showed that protein expression of pVHL and HIF-1α was decreased in CDH patients [20]. Because all of our investigated patients died under hypoxic conditions, this could account for the increased presence of the protein regardless of increased mRNA expression.
Upstream in the regulation of HIF we showed similar protein expression of PHD3 in control versus CDH lung tissue as well as in the canalicular and alveolar stages. Of the 3 known human PHDs (PHD1, PHD2, and PHD3) we were specifically interested in PHD3, because this isoform has the highest affinity for HIF-2α and is functionally important during prolonged periods of hypoxia [28]. Immunohistochemistry is not quantitative and therefore may mask any possible differences in PHD expression. From the literature, we know that HIFs and PHDs are reciprocally regulated and play key roles in the pathogenesis of hypoxia-induced pulmonary hypertension [29]. The possible therapeutic inhibition of PHDs and the consequent stabilization of HIF could enhance postnatal lung growth, which might be beneficial to CDH patients [30].
The results of our study suggest that lower VEGF-A expression, possibly as a result of selective downregulation of HIF-2α, may contribute to the pathophysiology of the transitional/alveolar lung development of CDH patients.