
Abstract
To investigate the correlation between maternal and fetal angiotensin-converting enzyme gene insertion or deletion polymorphism and birth size, angiotensin-converting enzyme gene insertion or deletion polymorphisms of 470 Japanese pairs of mothers and infants were genotyped. The relationships between maternal and fetal angiotensin-converting enzyme gene insertion or deletion polymorphisms and birth sizes (weight, length, head circumference, and the incidence of small-for–gestational age status) were analyzed. No relationship between fetal angiotensin-converting enzyme genotype and birth size was observed. In contrast, the incidence of small-for–gestational age status in babies whose mothers had 1 or 2 deletion alleles was greater than that in babies whose mothers had 2 insertion alleles (7% vs 3%, P = 0.045). Maternal deletion allele of angiotensin-converting enzyme is associated with fetal growth restriction in the Japanese normal population.
INTRODUCTION
Fetal growth is affected by interactions between fetal genes (such as insulin, insulin-like growth factor–I [IGF-I], IGF-II, and their receptors) and maternal uterine environment [1], but the detailed mechanism for this reaction remains unclear. The renin-angiotensin system (RAS) is one of the candidate factors that may affect intrauterine growth. Maternal RAS plays a significant role in regulation of uteroplacental blood flow by means of angiotensin-II receptors [2]. Disorders in the placental RAS may contribute to a decrease in uteroplacental blood flow, preeclampsia, and intrauterine growth restriction [3]. In addition, the polymorphism studies [4–6] of genes involved in RAS showed the positive correlation between RAS and insulin resistance. As insulin acts as a potent growth factor in placenta, fetal RAS may affect fetal growth through the interaction with insulin. These findings indicate that RAS, and particularly angiotensin-II, may also be fetal growth factors. However, it remains controversial which factor, maternal or fetal RAS, is more important for fetal growth.
In RAS, angiotensin-converting enzyme (ACE) plays an important role by catalyzing the conversion of angiotensin-I into angiotensin-II. In adults, plasma ACE activity level is associated with the 287–base pair fragment insertion (I)/deletion (D) polymorphism of the ACE gene intron 16, and the D allele is related to increased ACE activity [7,8]. It is generally accepted that ACE I/D polymorphism is a potent marker of personal RAS function. A large number of studies have confirmed that this polymorphism is associated with cardiovascular disorders [9,10]. On the other hand, the relationship between ACE I/D polymorphism and the incidents during pregnancy, such as pregnancy-induced hypertension (PIH), remains controversial [2,11–14].
A few studies [15–18] identified a positive correlation between the presence of the fetal ACE D allele and reduced fetal growth, but others [19,20] could not find positive correlation between these 2 factors. The correlation between maternal ACE I/D polymorphism and fetal growth has mainly been discussed in women with PIH [2,11,13]; only 1 study [2] in an Italian population has reported a tendency for intrauterine growth restriction with the maternal ACE D allele.
To reveal the correlation between ACE I/D polymorphism and fetal growth, we examined maternal and fetal ACE genotypes and compared them with birth sizes in the Japanese normal population.
MATERIALS AND METHODS
Study subjects
Study subjects were Japanese mothers and their term infants delivered between October 2004 and November 2007 at an obstetrics and gynecology hospital in Asahikawa. We included women for whom the 1st prenatal examination during the 1st trimester showed a structurally normal single fetus. The gestational age was calculated from the date of the last menstrual period and ultrasound examination. Mothers who were transferred to other hospitals because of pregnancy complications, such as threatened preterm delivery, were excluded. In addition, mothers who developed PIH (n = 14) were excluded from the group of study subjects, as PIH is closely related to fetal growth restriction by itself. Written informed consent was obtained from all of the participants. The Ethics Committee of Asahikawa Medical College approved this study protocol.
Measures
Midwives measured the birth weight, crown-to-heel length, and head circumference (HC) of the infants within 2 hours of delivery.
Molecular analysis of ACE I/D polymorphism
Maternal and cord blood samples were obtained, and ACE I/D polymorphisms were determined by polymerase chain reaction according to the standard method [21]. Genotyping was performed in a blind fashion. The subjects were then divided into 3 groups according to genotype (II, ID, and DD).
Statistical analysis
To compare the data at different gestational ages and between genders, weight, length, and HC were expressed as standard deviation scores (SDS) obtained using the reference of Japanese fetal growth [22]. A small-for–gestational age (SGA) infant was defined as the infant whose birth weight SDS was below −1.5 SDS. Subjects with the ID and DD genotypes were placed in the D+ group and compared with the II carriers (D– group). The birth sizes in these 2 groups were compared using an unpaired Student t-test. The incidence of SGA status in the 2 groups was compared by Fisher test. We considered P < 0.05 to be statistically significant. All statistical analyses were performed using the Statview program (version 5.0, SAS Institute Inc, Cary, NC, USA).
RESULTS
Our study subjects finally consisted of 470 pairs of mothers and infants. The mean gestational age of the study population was 278 days. Weight, length, and HC (SDS) of infants were 3083 g (–0.09), 48.7 cm (–0.44), and 33.8 cm (0.17), respectively. The male:female ratio of infants was 227:243.
The infant genotypes were II (38.5%), ID (47.7%), and DD (13.8%), and the maternal genotypes were II (40.4%), ID (48.3%), and DD (11.3%); these values were consistent with the Hardy Weinberg equilibrium. The relationships between infant ACE I/D polymorphism and birth size are shown in Table 1. There were no significant differences in birth size between the D+ and D– groups. The relationships between maternal ACE I/D polymorphism and birth size are shown in Table 2. The weight SDS and length SDS of infants in the maternal D+ group were less compared to those of the infants in the D– group, but the differences were not statistically significant. The incidence of SGA in the maternal D+ group was higher than that in the D– group (7% vs 3%, P = 0.045).
Fetal angiotensin-converting enzyme (ACE) insertion/deletion (I/D) polymorphism and birth size
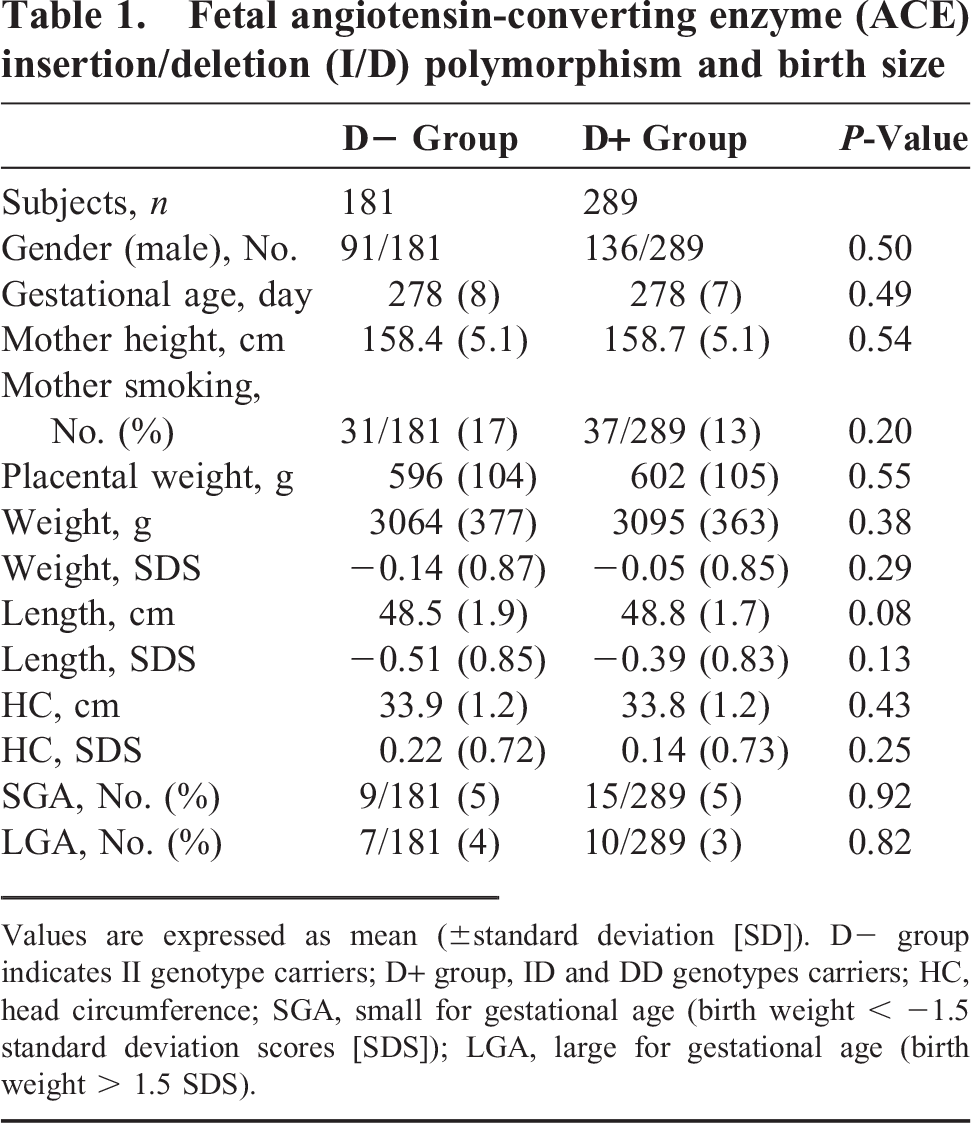
Values are expressed as mean (±standard deviation [SD]). D– group indicates II genotype carriers; D+ group, ID and DD genotypes carriers; HC, head circumference; SGA, small for gestational age (birth weight < −1.5 standard deviation scores [SDS]); LGA, large for gestational age (birth weight > 1.5 SDS).
Maternal angiotensin-converting enzyme (ACE) insertion/deletion (I/D) polymorphism and birth size
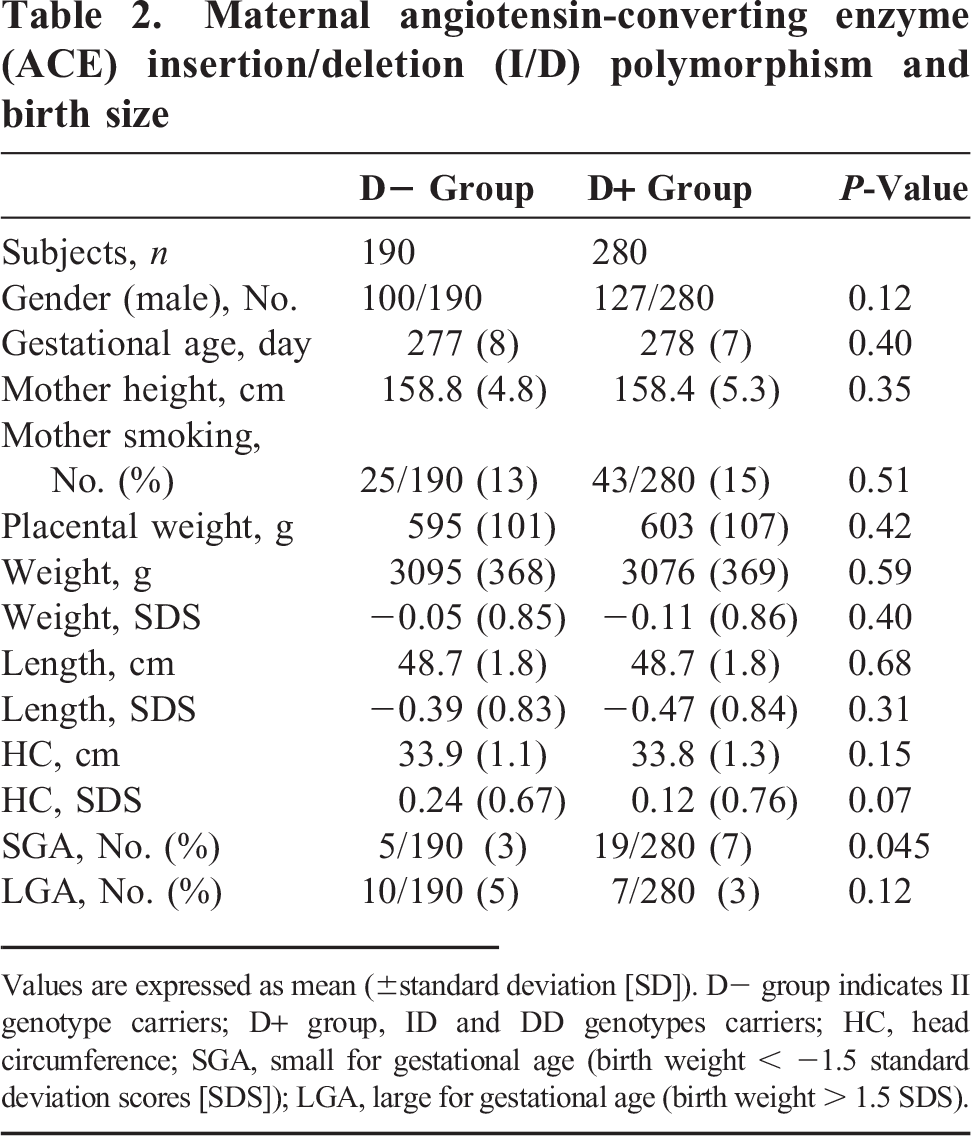
Values are expressed as mean (±standard deviation [SD]). D– group indicates II genotype carriers; D+ group, ID and DD genotypes carriers; HC, head circumference; SGA, small for gestational age (birth weight < −1.5 standard deviation scores [SDS]); LGA, large for gestational age (birth weight > 1.5 SDS).
DISCUSSION
This study provides novel evidence that the maternal ACE I/D polymorphism affects the birth size of Japanese newborns. However, we could not find any correlations between fetal ACE I/D polymorphism and fetal growth.
Our study has 2 significant points: (1) Many of the previous studies have investigated fetal or maternal genotype alone. We think it is important and necessary to investigate both fetal and maternal genotypes in the same study subjects; and (2) This is the 1st investigation regarding the relationship between ACE polymorphism and fetal growth in a Japanese population. The distribution of ACE polymorphism varies among different populations [23], and the frequency of D allele is lower in the Japanese population compared with other populations [23,24]. In our study, the incidence of D allele was similar to that reported in previous studies [24,25] involving the Japanese population. We think that the efficacy of the D allele for fetal growth could be more emphasized in the Japanese population.
There are currently 2 hypotheses that aim to explain how ACE I/D polymorphism affects fetal growth: (1) Fetal hypothesis: Insulin secretion has been associated with ACE polymorphism, and the presence of the ACE I allele has been associated with a greater insulin response in adults [4,5]. As insulin acts as a placental growth factor, the presence of fetal ACE I allele may have a potential for increasing fetal growth. (2) Maternal hypothesis: There is a reported connection between the maternal RAS and placental circulation; ACE activation may result in redistribution of the placental circulation [26], and the presence of the maternal ACE D allele has been associated with increased uteroplacental vascular resistance [2]. Thus, fetal growth could be restricted by the reduced blood flow in the fetus whose mother has D allele. Neither of these hypotheses has been confirmed until now; however, our investigation supports the maternal hypothesis.
Our study has 2 limitations. (1) Our study subjects consisted of healthy mothers; therefore, in this study we did not analyze other factors known to affect the size at birth, such as PIH and maternal diabetes. (2) We did not measure the ACE activity of study subjects. However, it is generally accepted that ACE D allele is associated with high plasma ACE activity in pregnant women [27,28] as well as in healthy adults. Thus, our investigation indicates that maternal RAS is more important than fetal RAS for fetal growth.