
Abstract
Vascular endothelial cadherin and β-catenin play a key role in establishment and maintenance of the endothelial monolayer integrity, regulation of vascular barrier function, and initiation of angiogenesis. The cadherin-catenin complex has been shown to be reduced in type 1 diabetic placenta, but the exact relationship between histopathologic findings and clinical data is not known. Immunohistochemistry of placental tissue from type 1, type 2, and gestational diabetes showed that diabetes per se might be compatible with normal levels of vascular endothelial (VE)-cadherin and β-catenin in fetoplacental vessels as long as the patient has not been treated with insulin. Immunoreactivity of VE-cadherin did correlate poorly with maternal glycemic control, as was investigated in this study, by birth weight, body mass index, and hemoglobin A1c (HbA1c). There was no correlation found between the immunoreactivity of β-catenin and birth weight, body mass index, or HbA1c. However our data did show a strong correlation between immunoreactivity and whether or not the patient had been treated with insulin. Patients diagnosed with gestational diabetes who had not been treated with insulin had similar levels of VE-cadherin and β-catenin to the control group, thus indicating that diabetes per se must not necessarily lead to a reduction. Our study suggests that therapeutic intervention using insulin in pregnancies complicated by diabetes might have potentially harmful effects on placental morphology. Future studies should further investigate these findings.
INTRODUCTION
Cadherins are a family of calcium-dependent, cell-cell adhesion molecules that mediate adhesive intercellular interactions and provide a dynamic cell-cell contact, wound healing, and positional clues to cells during development [1,2]. Vascular endothelial (VE) cells express a unique member of the cadherin family, termed VE-cadherin [3], which has been shown to play a central role in the establishment and maintenance of endothelial monolayer integrity [4], tubulogenesis [5], initiation of angiogenesis [6,7], and regulation of vascular barrier function [8]. The extracellular domain of VE-cadherin is coupled to the interior of cells through interactions with β-catenin, which is a member of the armadillo gene family [9]. β-Catenin associates with α-catenin, which plays a key role in linking the cadherin complex to the actin cytoskeleton [10]. Disruption of the cadherin-catenin complex from the cytoskeleton leads to the downregulation of cadherin-mediated cell adhesion [11] and, in some cases, the endothelial barrier function [12,13]. Breakdown of the endothelial barrier can lead to the unrestricted passage of molecules [14] and inflammatory cells [15,16], triggering uncontrolled inflammation and privileging further tissue damage [17,18].
In the human placenta fetoplacental vessels are found in chorionic villi, which are bathed in maternal blood [19]. This juxtaposition leaves the fetoplacental vessels vulnerable to any pathological alternations in maternal blood properties such as hypoxia, cytokines, hyperglycemia, hyperinsulinemia, and inflammatory mediators [20]. In the diabetic placenta a variety of pathological alterations have been reported, including hypovascularity, villous edema, and proliferation of the capillary endothelium [21]. Increased surface area and volume and enhanced angiogenesis have been reported even in the well-controlled diabetic placenta [22]. Previous studies showed a decrease in VE-cadherin and β-catenin in type I diabetic placenta [23], but the exact relationship between histopathologic findings and clinical data is not known. The aim of this study was to further investigate the expression of these 2 proteins in different diabetic subtypes, specifically analyzing the endothelium and trophoblast of stem villi, intermediate villi, and terminal villi, and correlate these findings with clinical data including body mass index, hemoglobin A1c (HbA1c), birth weight, and treatment modalities.
METHODS
The study adhered to the tenets of the Declaration of Helsinki. It was approved by the local ethics committee (Register Nr. 4517-12).
Eighty-six placentas were included in the study; they were collected immediately after delivery and processed in a single center (Department of Pathology, University of Bonn). Patients were type 1 diabetic patients (n = 10), type 2 diabetic patients (n = 5), and gestational diabetic patients, of which one subgroup had been treated with insulin previously (n = 20) and the other had never been treated with insulin (n = 21). All type 1 and type 2 diabetic patients had also been treated with insulin. Patients in the control group (n = 30) had uncomplicated term pregnancies with no known history of diabetes. The clinical details of patients are shown in Table 1.
Characteristics of the study subjects
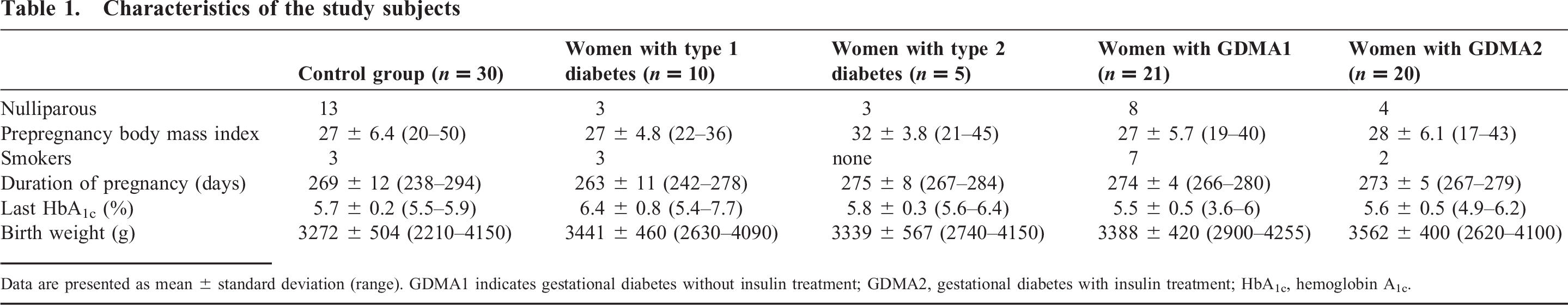
Data are presented as mean ± standard deviation (range). GDMA1 indicates gestational diabetes without insulin treatment; GDMA2, gestational diabetes with insulin treatment; HbA1c, hemoglobin A1c.
Tissue processing
Placental tissue was fixed in formaldehyde and embedded in paraffin according to well-established protocols. Serial sections were obtained (Mikrotom, Rotationsmikrotom HM 355; MICROM Laborgeräte GmbH, Berlin, Germany), collected on coated slides (SuperFrost Plus; Menzel, Braunschweig, Germany), dewaxed, dehydrated in ascending concentrations of ethanol, and placed in citrate buffer. To unmask antigens, an antigen retrieval procedure was performed by treating the samples in a microwave oven at 600 W. After cooling at room temperature, the sections were washed in phosphate-buffered saline and blocked with normal human serum. After 30 minutes incubation with blocking solution, sections were incubated in a moist chamber with the appropriate antibody against VE-cadherin (BD Transduction Laboratories, Franklin Lakes, NJ, USA; clone 75/cadherin-5; dilution 1∶100) and β-catenin (BD Transduction Laboratories; clone 14/beta-catenin; dilution 1∶100). Before washing, avidin, which binds selectively to complexed biotin, was added, and following a further wash step, color was developed (Vectastatin Elite ABC Kit Standard; PK 6100 Firma; Vector, Burlingame, CA, USA). The resulting signal was developed with diaminobenzidine. Immunoreactivities were assessed using a Carl Zeiss Axioskop (Leica DC 300; Jena, Germany), and densitometric readings were obtained using Borland Delphi 6 (Austin, TX, USA) as is described in detail previously [24]. In short, the principle behind this computer-based morphometry is based on the different staining patterns of cells, nuclei, and immunoreactivities. After storing the photographs as 1280 × 1024-pixel RGB bitmaps at a color resolution of 24 bits per pixel, a shading correction and a grey transformation of the green color channel are calculated. After application of a blurring filter based on an arithmetic mean filter, thresholding of the positively stained immunoreactivities is performed by a histogram analysis of the grey value distribution, resulting in a binary image in which binary object detection is performed and all objects with an area lower than a specific threshold, such as cell nuclei or debris, are eliminated. After this conversion, a precise and rapid mathematical quantification of individual colors is possible and the total area of the immunoreactivities can then be expressed as a percentage of the total parenchyma area. These steps are performed automatically. The sections were counterstained with hematoxylin solution.
Statistical analysis
Pearson correlation was used to access the dependency between continuous variables. Analysis of variance followed by post hoc comparisons (Tukey) were carried out for between-group comparisons. Statistical calculations were performed using the Statistical Package for Social Sciences (SPSS, Chicago, IL, USA).
RESULTS
VE-cadherin
The median staining intensities found in the endothelium and trophoblast layers of terminal, intermediate, and stem villi are shown in Table 2. In the endothelial layer a significant reduction of VE-cadherin in terminal, intermediate, and stem villi compared to the control group was found in type 1 diabetic patients, type 2 diabetic patients, and gestational diabetic patients who had been receiving insulin treatment (Figs. 1,2). The VE-cadherin staining was similar in gestational diabetic patients compared to the control group. No differences were found in the trophoblast layer.
VE-cadherin; endothelial and trophoblastic staining intensity in the different types of chorionic villi
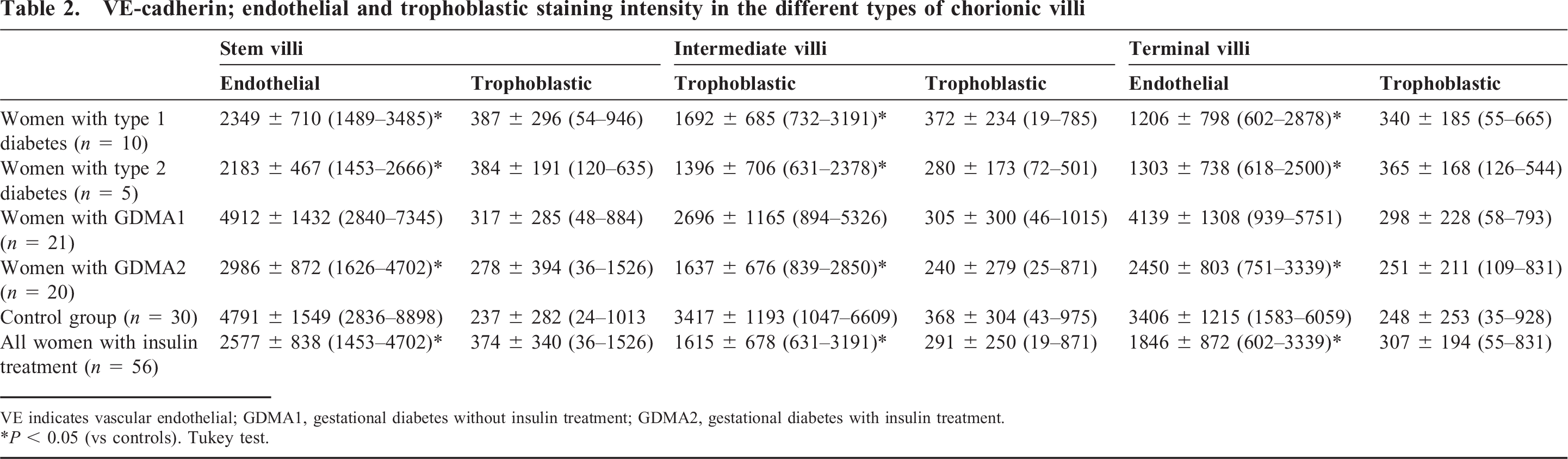
VE indicates vascular endothelial; GDMA1, gestational diabetes without insulin treatment; GDMA2, gestational diabetes with insulin treatment.
P < 0.05 (vs controls). Tukey test.
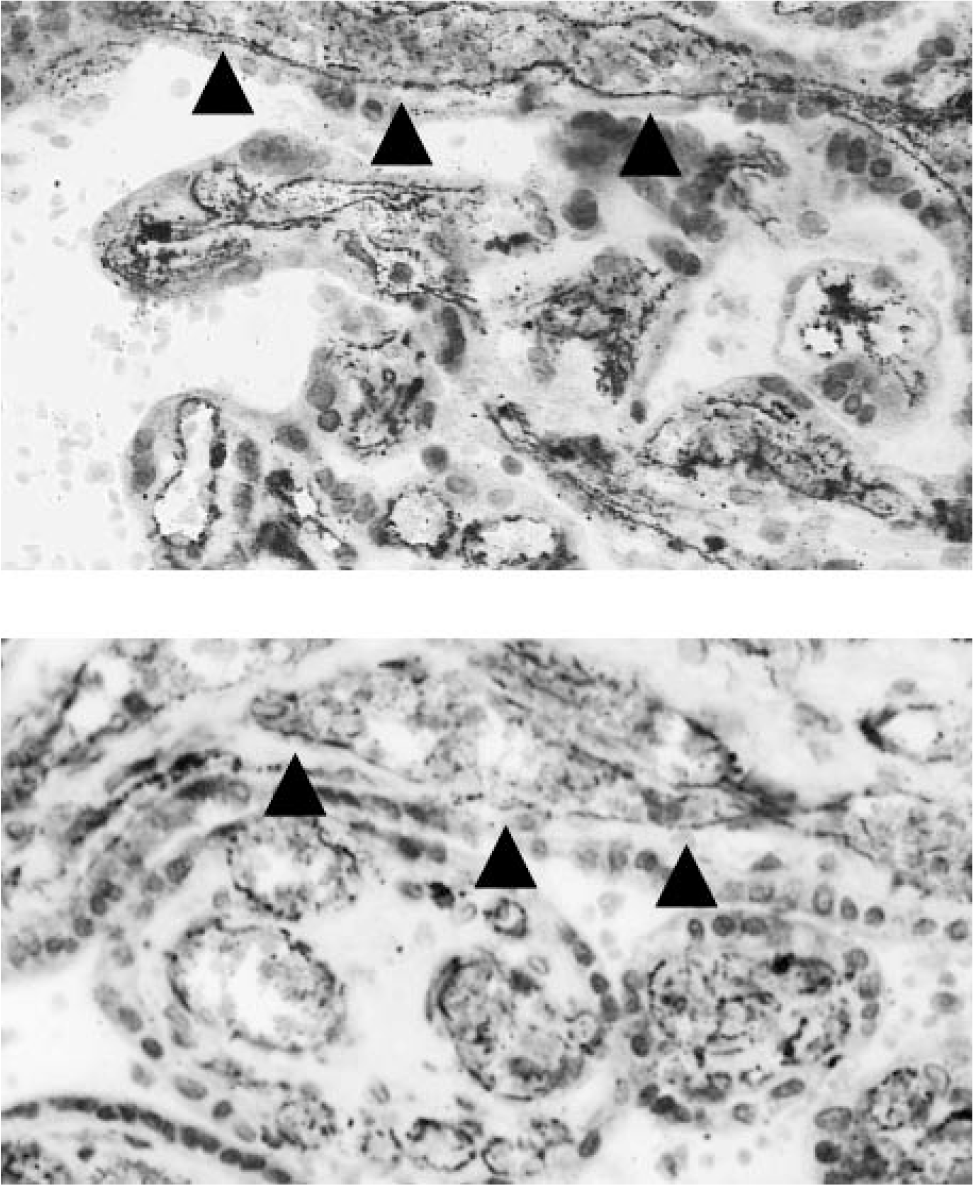
Micrographs showing localization of vascular endothelial cadherin (arrows) in a human placenta from a normal pregnancy (above) and one complicated by insulin-dependent diabetes mellitus (below) with reduced expression in the latter.
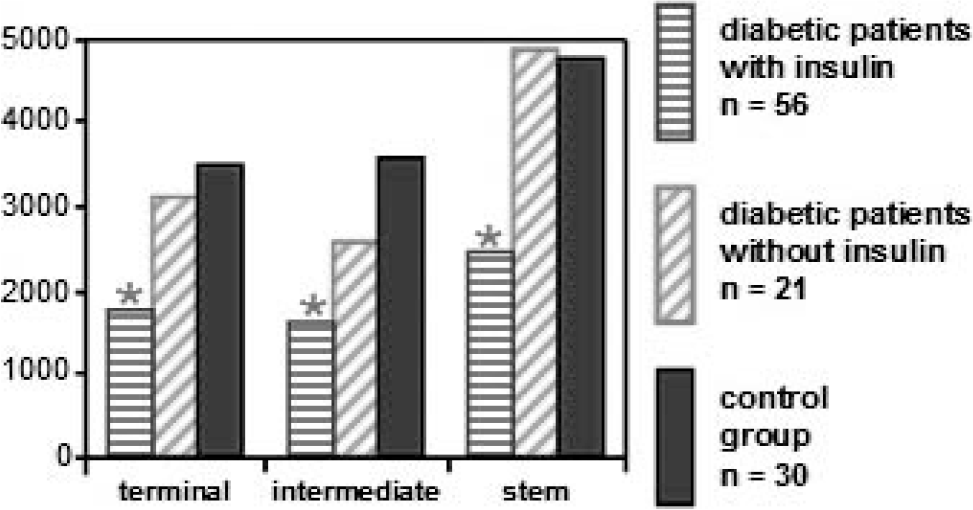
Histograph of the mean of endothelial immunofluorescence of vascular endothelial cadherin in terminal, intermediate, and stem villi. * P < 0.05 (vs control group). Tukey test.
β-catenin
The median staining intensities found in the endothelium and trophoblast layer of terminal, intermediate, and stem villi are shown in Table 3. In the endothelial layer a reduction of β-catenin in terminal, intermediate, and stem villi was found between patients receiving insulin treatment and the control group (Figs. 3,4). In the trophoblast layer a reduction of β-catenin was found only in intermediate villi between patients receiving insulin treatment and the control group.
β-catenin; endothelial and trophoblastic staining intensity in the different types of chorionic villi

GDMA1 indicates gestational diabetes without insulin treatment; GDMA2, gestational diabetes with insulin treatment.
P < 0.05 (vs controls). Tukey test.
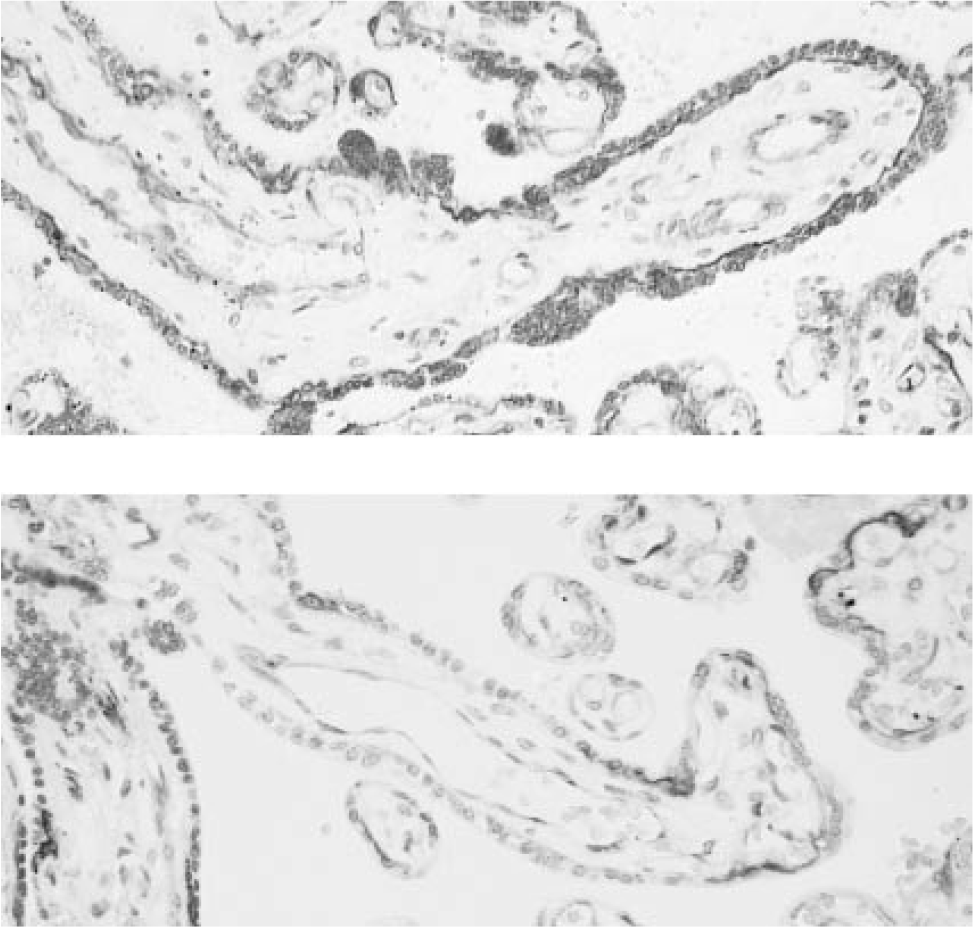
Micrographs showing localization of β-catenin in a human placenta from a normal pregnancy (above) and one complicated by insulin-dependent diabetes mellitus (below) with reduced expression in the latter.
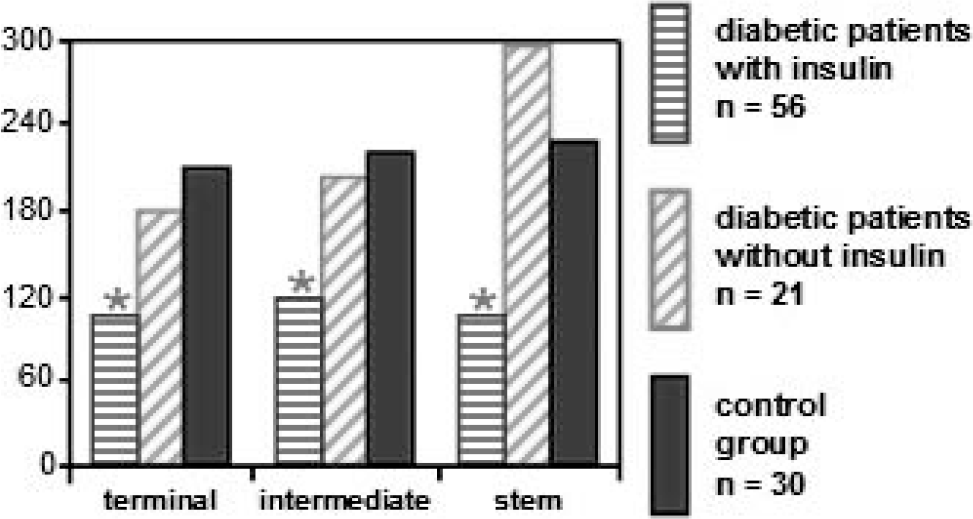
Histograph of the mean of endothelial immunofluorescence of beta catenin in terminal, intermediate, and stem villi. * P < 0.05 (vs control group). Tukey test.
Clinical data
The median birth weight was 3377.9 ± 473 g (n = 85, minimum [min] = 2210 g, maximum [max] = 4230 g). A significant correlation between birth weight and VE-cadherin was found in the endothelial layer of intermediate villi (P = 0.041) (Table 4). The median HbA1c was 5.7% ± 0.6% (n = 59, min = 3.6%, max = 7.7%). The HbA1c did significantly correlate with the VE-cadherin expression in the endothelial layer of the terminal (P = 0.023) and stem villi (P = 0.005) (Table 4). The median body mass index was 27.6 ± 6.4 (n = 85, min = 17, max = 50). No significant correlation was found regarding body mass index (Table 4), smoking, nulliparity, or duration of pregnancy.
Pearson product-moment correlation coefficient of clinical data
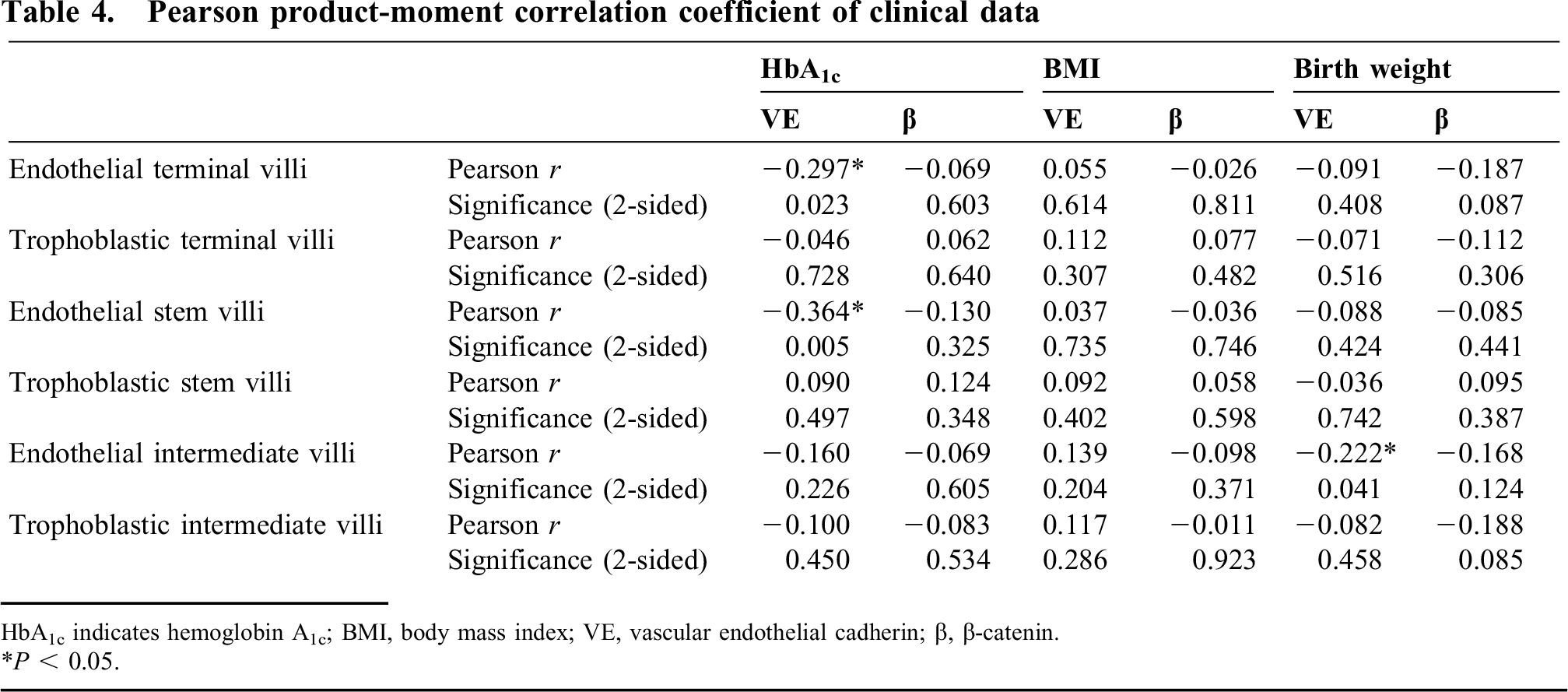
HbA1c indicates hemoglobin A1c; BMI, body mass index; VE, vascular endothelial cadherin; β, β-catenin.
P < 0.05.
DISCUSSION
This study showed that diabetes per se might be compatible with normal levels of VE-cadherin and β-catenin in fetoplacental vessels as long as the patient has not been treated with insulin. This study showed an inverse correlation between VE-cadherin expression and HbA1c only in the endothelium of terminal villi and stem villi and an inverse correlation between birth weight and VE-cadherin expression in the endothelium of intermediate villi. Statistical analysis using the Pearson correlation coefficient did not show a relation between the clinical data obtained and immunoreactivity of β-catenin in the endothelium and trophoblast layer in stem, intermediate, or terminal villi (Table 4). In contrast, the data did show a strong correlation between immunoreactivity and whether the patient had been treated with insulin. Those patients who had been diagnosed with gestational diabetes and had not been treated with insulin had similar levels of VE-cadherin and β-catenin to the control group, thus indicating that diabetes per se does not necessarily lead to a reduction. The inverse correlation between birth weight and HbA1c on VE-cadherin expression may confound the effect of the insulin treatment. The effect of the insulin treatment appeared to be stronger, because it was present in all types of villi, but further studies seem to be necessary to investigate this effect in greater depth.
The well-known phenotype in the type 1 diabetic placenta constitutes vessels that exhibit altered junctional maturity and molecular occupancy with increased leakiness. During critical periods of placental development these altered conditions may compromise efficient functioning throughout gestation and may have ramifications for fetal growth and well-being [23]. Endothelial cells in type 1 diabetic placental vessels have been shown to exhibit altered VE-cadherin and β-catenin in more than 50% of vessels [25]. But the exact mechanism remains unclear. Hyperglycemia has been shown to promote angiogenesis [26], constriction [27], inflammation [28], and permeability [29]. Hence, hyperglycemia might exhibit basic pathophysiologic properties that would support its role in causing vascular dysfunction, but increasing evidence contradicts this paradigm. Increased angiogenesis was found even in well-controlled type 1 diabetic placenta [30], and macrosomic neonates were still present even in diabetic patients with optimal glucose control [31]. According to our data, hyperglycemia seems to play only a minor role in causing vascular dysfunction, as neither birth weight nor HbA1c had a significant impact on the expression of VE-cadherin and β-catenin.
Circulating insulin might be an alternative agent that could cause a decrease in VE-cadherin and β-catenin. The terminal villous capillaries and the adjacent trophoblast are rich in vascular endothelial growth factor (VEGF) receptors [32,33]. Vascular endothelial growth factor is found to be increased in diabetic placentas and is present in both the endothelium and trophoblast layers of terminal, intermediate, and stem villi in type 1 diabetic placentas, as opposed to terminal villi only in normal pregnancies [25]. Perfusion studies of the placenta have shown that high insulin perfusion of the fetal circulation results in increased endothelial VEGF and junctional disruption, of which the exact pathomechanism remains unclear but might be related to trophoblast signaling [34]. In the retina, acute intensive insulin treatment has been shown to activate hypoxia-inducible factor 1-α, which in turn raises VEGF levels [35]. In our study maternal insulin treatment had a strong impact on the expression of VE-cadherin and β-catenin and might explain the effects of diabetes on human fetoplacental vessels even in well-controlled diabetic pregnancies.
Limitations of the study include the variety of effects of quantitative immunohistochemistry such as tissue fixation, antigen retrieval, antibody lot, color reaction development, and a lack of type 1 and 2 diabetics who were not treated with insulin. Future studies using different measurements should enhance our understanding of the dynamic pathogenesis of vascular dysfunction in pregnancies complicated by diabetes. The current analysis of immunohistochemistry of placental tissue from type 1, type 2, and gestational diabetes shows that diabetes per se might be compatible with normal levels of VE-cadherin and β-catenin in fetoplacental vessels as long as the patient has not been treated with insulin. The inverse correlation between birth weight and HbA1c on VE-cadherin expression may confound the effect of insulin treatment on VE-cadherin expression. Therefore, future studies are needed to investigate this effect further.