
Abstract
Background
Hepatocyte growth factor (HGF) is a growth factor thought to attenuate Th2-driven eosinophilic airway inflammatory responses. Increased expression of HGF and its receptor c-Met in nasal polyps suggests a role in disease pathogenesis. The effect of HGF on human sinonasal epithelial cell (SNEC) responses to Th2 inflammatory cytokines in chronic rhinosinusitis with nasal polyps (CRSwNP) has not been explored.
Methods
SNECs isolated from patients with CRSwNP and control subjects were grown in cell culture at the air–liquid interface. The Th2 cytokine IL-13 was applied for 24 hours in the presence or absence of HGF. Eotaxin-3 and c-Met expression was assessed using real-time PCR, immunohistochemistry, and flow cytometry.
Results
SNECs obtained from both CRSwNP and control subjects showed markedly increased expression of eotaxin-3 after exposure to IL-13. HGF significantly blocked IL-13–induced expression of eotaxin-3 in control SNECs, but not in SNECs derived from CRSwNP subjects.
Conclusion
SNECs are active participants in sinonasal mucosal immunity, expressing inflammatory mediators in response to potential pathogens and endogenous cytokines. Although Th2 cytokines can elicit expression of proeosinophilic mediators by SNECs, HGF appears to have a down-regulating effect on this response. In patients with CRSwNP, SNECs are resistant to this attenuation, showing continued IL-13–induced eotaxin-3 expression despite HGF treatment. Abnormalities in the regulation of epithelial cell responses to endogenous cytokines and growth factors may contribute to the persistent eosinophilic inflammatory state in CRSwNP.
Keywords
Histologically, CRSwNP is characterized by a predominance of eosinophils, goblet cell hyperplasia, and thickening of the basement membrane. The eosinophilic inflammation associated with CRSwNP occurs in a milieu distinguished by the prevalence of Th2 cytokines such as IL-4, IL-5, and IL-13. 4 Although the cause of the shift from Th1 to Th2 mediators in CRSwNP is not known, both host factors and external mechanisms have been proposed.
Hepatocyte growth factor (HGF) is a potent mitogen, which acts as a stimulator for various epithelial cell types such as those found in the liver, kidney, and in the upper airway. Its signal is potentiated through c-Met, a tyrosine kinase receptor encoded by a proto-oncogene. HGF and c-Met have been shown to be elevated in conditions involving acute tissue injury such as liver injury and acute pneumonia. 5 HGF and c-Met are also involved in the various phases of wound repair such as inflammation, epithelialization, and the formation of granulation tissue. Their role in upper airway inflammation and CRSwN has been established by several studies. Rho et al. confirmed the presence of c-Met and HGF in the submucosal glands and epithelium of nasal polyps. 6 A similar study found that the expression of these molecules is increased in inverted papillomas. 7 The objective of this study is to identify c-Met expression in human sinonasal epithelial cells (SNECs) and determine the functional consequences of HGF stimulation in nondiseased controls and patients with CRSwNP.
Materials and Methods
Human Subjects
Six patients with CRSwNP and six control subjects were enrolled in the study. The research protocol was approved through the Johns Hopkins Institutional Review process, and all subjects gave signed informed consent. The CRSwNP patients were defined by historical, endoscopic, and radiographic criteria and by meeting the definition of the American Academy of Otolaryngology–Head and Neck Surgery Chronic Rhinosinusitis Task Force. 8 Specifically, the patients had continuous symptoms of rhinosinusitis as defined by the Task Force report for >12 consecutive weeks, associated with computed tomography of the sinuses revealing isolated or diffuse sinus mucosal thickening or air–fluid levels. Nasal endoscopy revealed the presence of at least one sinonasal polyp before or at the time of ESS.
Surgery was only performed if a patient's symptoms and radiographic findings failed to resolve despite at least 6 weeks of treatment with oral antibiotics, topical corticosteroids, decongestants, and mucolytic agents, as well as 3 weeks of systemic corticosteroid. The six control subjects had no evidence of sinus disease and were undergoing endoscopic approaches for transnasal endoscopic pituitary surgery, cerebrospinal fluid leak repair, or sphenoidotomy for biopsy of an isolated, noninflammatory sphenoid sinus process. Before surgery, the CRSwNP patients received 1 week of oral prednisone. All tissue specimens were taken from the middle turbinate, the resected uncinate process, or the anterior ethmoid sinus. The CRSwNP specimens were carefully selected to avoid polyps or tissue that was grossly polypoid in appearance.
Epithelial Cell Dissociation from Human Sinus Mucosal Tissue
Mucosal tissue removed during ESS was collected into sterile cold transferred to phosphate-buffered saline (PBS) supplemented by penicillin (100 U/mL; Gibco, Gaithersburg, MD), streptomycin (100 μg/mL, Gibco), amphotericin B (2.5 μg/mL, Gibco), and gentamicin (50 μg/mL, Gibco). The samples were placed through a cell strainer (BD Falcon, San Jose, CA) into Ham's F12 media containing 0.01% protease Sigma type XIV (Sigma, St. Louis, MO) and supplemented with antibiotics as mentioned previously. Digestion in protease was performed at 4°C overnight, with separation of cells by agitation the following day. The cells were separated by straining into a conical tube to which fetal bovine serum (FBS) was added to a final concentration of 10%, inactivating the protease. The cells were centrifuged twice at 1300 rpm for 10 minutes, after which the supernatant was aspirated and discarded. The washed passage 0 (P0) SNEC cells were then seeded, at a density ≥1.5 × 104 cells/cm2, onto Vitrogen 100–coated (1:75 in sterile water; Cohesion, Palo Alto, CA) P-100 dishes in bronchial epithelium growth medium (BEGM), as previously described by our group. 9 The cells were then grown at 37°C for 24 hours and then washed with Hank's balanced salt solution (HBSS) (Biofluids, Rockville, MD) to remove debris. The media was then changed every 48 hours until the cells reached confluence.
SNEC Culture at the Air–Liquid Interface (ALI)
Confluent cells were washed with HBSS before trypsinization and then treated at 37°C for 4 minutes with a solution containing 0.2% trypsin (Sigma), 1% polyvinylpyrrolidone (Sigma), and 0.02% EGTA (Sigma) in HBSS. The trypsin was then neutralized by the addition of an equal volume of cold soybean trypsin inhibitor at a concentration of 1 mg/mL in Ham's F12 media. Dissociated cells were washed and resuspended into BEGM media and plated into human type IV placental collagen (type VI; Sigma)–coated 12-well Falcon filter inserts (0.4-μm pore size; Becton Dickinson, Franklin Lakes, NJ). Although some P1 cells were directly used for flow cytometry, other P1 cells were grown to confluence with BEGM, above (1 mL) and below (2 mL) the cells. When confluent, medium is removed from above the cultures and the medium below the inserts was changed to ALI medium consisting of LHC Basal Medium/DMEM-H (50:50; Gibco) containing the same concentrations of additives as BEGM with the exception that the concentration of epidermal growth factor is reduced to 0.63 ng/mL and amphotericin B is omitted. Each set of cultures came from a separate patient source and was maintained at the ALI with the apical surfaces remaining free of medium for at least 3 weeks before study.
Stimulation of Cells
Ciliated epithelial cells at the ALI were stimulated basolaterally with 10 ng/mL of IL-13 and/or 50 ng/mL of HGF diluted in ALI medium for 24 hours.
RNA Extraction/Reverse Transcription
Total RNA was extracted using the RNeasy Kit by Qiagen (Valencia, CA) according to the manufacturer's directions. RNA was quantified spectrophotometrically and visualized by staining with ethidium bromide to determine RNA integrity. Absorbance ratios at 260/280 nm were >1.80 for all samples studied. Isolated mRNA was reverse transcribed with random hexamer primers using the Qiascript Reverse Transcription kit (Qiagen).
Real-Time Polymerase Chain Reaction (PCR)
Real-time PCR was performed in a Light-Cycler 1.2 (Roche, Indianapolis, IN) using the SYBR Green PCR Kit (Qiagen). The 18S was used as a housekeeping gene with which to normalize expression and was determined in previous experiments to be stable across control and CRS sinonasal tissue samples. The reaction mix consisted of 0.8 μg of total RNA (target genes) or 0.08 μg of total RNA (18S RNA), 10 μL of QuantiTect SYBR Green PCR, 1.5–5 mol/L of target primers, or 1.0 mol/L of 18S rRNA primers, in total volume of 20 μL. All primers (18S, sense 5′ GTAACCCGTTGAACCCCATT-3′, antisense 5′-CCATCCAATCGGTAGTAGCG-3′); c-Met, sense 5′GGCATGTCAA-CATCGCTCTA-3′, antisense 5′ GTTGCAGATTCAGCTGTTGC-3′; eotaxin-3, sense 5′ AACTCCGAAACAATTGTACTCA-3′, antisense 3′ GTAACTCTGGGAGGAAACACC-5′) were commercially synthesized by Invitrogen (Carlsbad, CA). The cycle parameters used were 95°C for 15 minutes to activate Taq polymerase, followed by 45 cycles at 94°C for 15 seconds, 60°C for 30 seconds, and 72°C for 30 seconds. Amplicon expression in each sample was normalized to its 18S RNA content. The level of expression of target mRNA was determined as the delta CT (ΔCT). The ΔCT method uses the difference in CT value obtained between a normalizing housekeeping gene (18S) and the target gene to calculate relative quantification (ΔCT = the difference in threshold cycles for target and housekeeping gene). This normalization reduces sample-to-sample variations in signal strength. A decrease in the ΔCT by 1 U equals a doubling of the level of the target gene. Consistent use of the quantities of total RNA described previously (800 and 80 ng for target molecules and 18S RNA, respectively) resulted in highly reproducible real-time PCR cycle thresholds for each of the amplicons across all cell samples. Negative controls, consisting of reaction mixtures containing all components except target RNA, were included with each PCR run.
Flow Cytometry
Adherent cells SNECs were detached from 6-well plates as described previously and transferred into 1.5-mL microfuge tubes at a concentration of 200,000 cells/tube. Cells were incubated with 0.5 μL of biotinylated c-Met antibody (ebiosciences, San Diego, CA) for 30 minutes at 4°C. Cells were then centrifuged and washed twice and resuspended in cold PBS with sodium azide and 2% FCS. Cells were then stained with polycoerythrin (PE)-conjugated streptavidin for 30 minutes at 4°C followed by two washes and fixation with 2% paraformaldehyde. Isotype controls consisted of PE-conjugated rat anti-human IgG (eBiosciences). Analysis was performed on a FACS calibur flow cytometer (BD Biosciences, San Jose, CA) and data were analyzed using CellQuest software (BD Biosciences). Cell surface protein is expressed as the increase in mean fluorescence intensity over background (species-specific isotype-matched control) or percentage of positive staining cells.
Immunofluorescence and Confocal Microscopy
SNECs at the ALI were fixed in 4% paraformaldehyde for 15 minutes and washed twice in cold PBS. Cells were then incubated with 1% bovine serum albumin in staining buffer for 30 minutes followed by incubation with antibodies to b-tubulin and c-Met (Abcam, Cambridge, MA) diluted in 1% bovine serum albumin overnight at 4°C. After washing SNECs three times with PBS, secondary antibody was added for 1 hour at room temperature. Cells on inserts were then mounted on slides and imaged using a confocal microscope (LSM 510 meta, Carl Zeiss Microimaging, Thornwood, NY). Images are flattened z stacks taken at 1-μm intervals.
Statistical Analysis
Raw data from real-time PCR were entered into a spreadsheet (Excel; Microsoft Corp., Redmond, WA). Statistical analysis was performed using a software program (Excel; Microsoft Corp.). Data are expressed as mean ± SEM. Statistical significance of differences was determined using the t-test. Differences were considered statistically significant at p < 0.05.
Results
SNECs in Culture Express c-Met by Immunofluorescence and Flow Cytometry
SNECs from both CRSwNP and control patients express c-Met mRNA detected by reverse transcription-PCR and protein assessed by immunofluorescence and flow cytometry (Fig. 1; 95–100%). Flow cytometry was performed on submerged SNECs and therefore resemble the basal layer of cells in ciliated respiratory epithelium. Immunofluorescence of SNECs at the ALI shows c-Met staining primarily in the basolateral nonciliated epithelial cells. Additionally, stimulation of SNECs from both groups with the potent Th2 cytokine, IL-13, does not further augment c-Met expression.
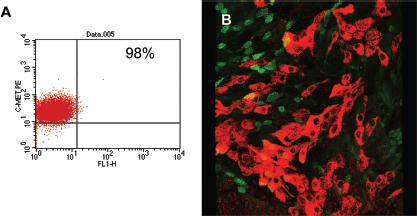
Sinonasal epithelial cells (SNECs) express membrane bound c-Met in both chronic rhinosinusitis (CRS) and control patients. (A) Flow cytometric analysis of control SNECs (grown in submerged culture) stained with c-Met phycoerythrin. Ninety-eight percent of the cells express c-Met protein on the surface of each cell. Each dot on this diagram represents one epithelial cell that stained positive for c-Met; the fluorescence intensity on the y-axis is indicative of the number of membrane bound c-Met receptors on each cell. (B) Immunofluorescence and confocal microscopy of SNECs at the air–liquid interface (ALI) from CRS with nasal polyps (wNP) stained with c-Met and tubulin (a marker for ciliated epithelial cells). There are few areas of double staining, indicating that c-Met is primarily expressed in basolateral cells that are nonciliated.
HGF Down-Regulates c-Met Protein in CRSwNP
To assess the effects of HGF on c-Met expression, SNECs at the ALI were stimulated with HGF. Subsequently, cDNA derived from these cells and real-time PCR was used to quantify c-Met levels. There is no statistically significant difference in c-Met mRNA levels between control and CRSwNP patients after HGF stimulation. To examine protein expression, flow cytometry was used to measure c-Met in CRSwNP and control SNECs stimulated with HGF before growth at the ALI. We observed a substantial decrease in membrane bound c-Met in CRSwNP (50%; range, 40–60%) but not in control patients (95%; Figs. 2 and 3).
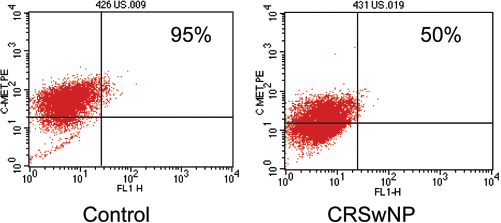
Flow cytometric sinonasal epithelial cell (SNEC) c-Met expression after hepatocyte growth factor (HGF) stimulation. SNECs from control and chronic rhinosinusitis with nasal polyp (CRSwNP) patients were stimulated with 50 ng/mL of HGF and stained with c-Met PE antibody and analyzed by flow cytometry. At baseline both controls and CRSwNP patients express 95–100% of c-Met. The panel on the left shows that controls stimulated with HGF do not modify c-Met expression. CRSwNP SNECs stimulated with HGF is depicted on the right, which shows nearly a 50% reduction of membrane bound c-Met protein.
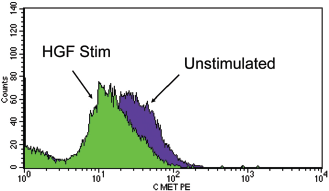
Reduced expression of c-Met after hepatocyte growth factor (HGF) stimulation in chronic rhinosinusitis with nasal polyps (CRSwNP). Flow histogram depicting sinonasal epithelial cells (SNECs)from CRSwNP before and after HGF stimulation as labeled. Fluorescence intensity decreases after HGF stimulation, indicating a reduction in membrane bound c-Met expression.
HGF Down-Regulates Eotaxin-3 Levels in Control Patients but not CRSwNP
Although SNECs at baseline express low levels of eotaxin-3, IL-13 stimulation potentiates eotaxin-3 secretion. Therefore, to assess the functional consequences of HGF stimulation, SNECs at the ALI were exposed to a combination of IL-13 and HGF and eotaxin-3 levels were determined by real-time PCR. SNECs from CRSwNP and control patients produced similar amounts of eotaxin after IL-13 stimulation. However, HGF blocked IL-13–induced expression of eotaxin-3 in control SNECs by fourfold (two-cycle real-time PCR threshold, p < 0.02), but not in SNECs derived from CRSwNP subjects (Fig. 4).
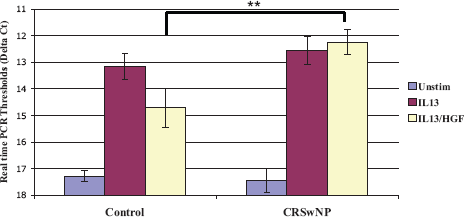
Eotaxin-3 production from sinonasal epithelial cells (SNECs). SNECs at the air–liquid interface (ALI) from control and chronic rhinosinusitis (CRS) patients were stimulated with either 10 ng/mL of IL-13 alone or 50 ng/mL of hepatocyte growth factor (HGF) and 10 ng/mL of IL-13. Realtime polymerase chain reaction (PCR) was then used to measure eotaxin-3 levels from cDNA made from cells. All values are normalized to 18S and depicted as delta cycle thresholds. Error bars represent ± SE of the mean. HGF attenuates IL-13–induced production of eotaxin in controls but not CRS with nasal poltps (wNPs). **This reached statistical significance at p < 0.02.
Discussion
Although CRSwNP inflammation is produced in part by an exaggerated production of Th2 cytokines and mediators, the cause has not been well defined. Multiple groups have proposed that an abnormal immune response may occur as a result of exposure to microorganisms and/or their products such as fungi, staphylococci, or pseudomonal biofilms.10–15 Exotoxins from Staphylococcus aureus, acting as superantigens, are able to induce a polyclonal T-cell response that activates larger subpopulations of T cells than a typical antigen. S. aureus toxins have been shown to increase production of Th2 cytokines such as IL-4 and IL-5, leading to increased lymphocytic and eosinophilic infiltration. 11 Bachert et al. showed the presence of IgE antibodies to the S. aureus superantigens, SEA and SEB, in nasal polyps.12–14 The presence of S. aureus superantigens in nasal polyps suggests a potential role for exogenous factors as a contributor to the Th2 immune response in CRSwNP. However, these innocuous and ubiquitous agents, such as S. aureus and fungi, do not typically generate an exaggerated Th2 response in a healthy host, which alludes to the presence of an additional, endogenous abnormality in affected CRSwNP patients. These findings have been the primary impetus to better understand host gene expression in CRSwNP.
SNECs are active participants in mucosal immunity, expressing inflammatory mediators in response to potential pathogens and endogenous cytokines. Although many studies have implicated a dysregulation of epithelial innate immune genes in potentiating Th2 inflammation, few studies have examined the role of growth factors and genes involved in repair. 15 In 2006, Rho et al. described the overexpression of c-Met, a proto-oncogene expressed on epithelial cells, and its growth factor ligand HGF in nasal polyps compared with inferior turbinate mucosa. 6 Subsequently, Stankovic et al. performed a gene chip microarray analysis of nasal polyps compared with normal mucosa and also found overexpression of c-Met. 16
Although the focus of research on c-Met/HGF has been on neoplastic disease, very little is known about its role in sustaining chronic inflammation. HGF, the ligand for c-Met (or HGF receptor), is a potent mitogen and morphogen that promotes motility, proliferation, and differentiation in a number of different cell types, including upper airway epithelial cells. 17
In this study, we show the expression of c-Met mRNA and protein in SNECs cultured from control and CRSwNP patients. At baseline, close to 100% of SNECs from controls and CRSwNP express membrane bound c-Met. Interestingly, both Rho et al. and Stankovic et al. found an increased expression of c-Met in the submucosal glandular epithelium and the basal layers of respiratory epithelium in nasal polyps compared with inferior turbinate mucosa from controls. This difference may perhaps be explained by the fact that our study examined c-Met expression in SNECs isolated from mucosal tissue with polypoid inflammation and not whole nasal polyps that contain large amounts of submucosal glands. Additionally, when CRSwNP SNECs are stimulated with HGF, there is a profound decrease in membrane bound c-Met protein but not in mRNA expression. These findings are consistent with a study by Shen et al., who quantified c-Met mRNA and protein levels via Northern and Western blots in tracheal epithelial cells after stimulation with HGF. This study found that while mRNA levels do not change with HGF stimulation, total protein expression was decreased. 18 Additionally, our study found that the potent Th2 cytokine IL-13 does not modulate or augment mRNA or protein expression of c-Met in SNECs from either control or CRSwNP patients.
Eotaxins are among the primary chemokines produced by epithelial cells that act as potent chemotactic factors for eosinophilic inflammation. 19 A few studies in the literature have reported the effects of HGF on eosinophilic inflammation in bronchial asthma, another Th2-mediated disease that shares many similarities with CRSwNP. Okunishi et al. in 2009 examined the effect of HGF on asthmatic airway inflammation in the ovalbumin-allergic mouse model, before ovalbumin inhalation. They found that intratracheal administration of HGF profoundly decreased eosinophilic airway inflammation and down-regulated antigen presentation to T cells in the lung. 17 Similarly, Ito et al. show that HGF treatment of eosinophils suppressed CD69, a marker of eosinophil activation, and also suppressed the release of effector inflammatory molecules such as reactive oxygen species and eosinophil-derived neurotoxin. 20 To study the functional consequences of HGF stimulation on the nasal epithelium, we stimulated SNECs from controls and CRSwNP with IL-13 and HGF. Our study indicates that HGF attenuates IL-13–induced eotaxin production from SNECs in control patients, but not CRSwNP. This suggests that the ordinary ability of HGF to down-regulate Th2 inflammation is dysfunctional in the nasal epithelium in CRSwNP. To our knowledge, this is a novel finding not previously reported. The decreased c-Met receptor found after HGF stimulation in CRSwNP may perhaps explain the inability of HGF to attenuate IL-13–induced eotaxin production in this population. Further research is needed to explore this mechanism.
One inherent difference in our methodology is that preoperative systemic corticosteroids were given to the CRSwNP patients but not to the control patients. Although it is possible that treatment with steroids potentially impacts the expression of c-Met, it is important to note that flow cytometric analysis of SNECs grown in submerged culture showed similar c-Met expression between the two groups. With respect to SNECs grown at the ALI, because this culture model system involves multiple weeks of growth and differentiation in media free of corticosteroids, it seems unlikely that there would be significant residual effects from prior exposure in vivo. However, it can not be ruled out that remote exposure to steroids might have a small effect on the subsequent physiology of c-Met in vitro.
Although our study population was relatively limited and understanding that cell culture systems imperfectly model the in vivo environment, these findings highlight that abnormalities in the regulation of epithelial cell responses to endogenous cytokines and growth factors may contribute to the persistent eosinophilic inflammatory state in CRSwNP. Because mucosal innate immunity is closely tied to growth and repair responses, dysfunction of the c-Met/HGF pathway may represent a mechanism through which persistent Th2 inflammation can be propagated by the sinonasal epithelium. A better understanding of how growth factors and their signaling pathways interact with epithelial cell innate immune function may contribute to new therapeutic strategies for CRSwNP.