
Abstract
Background
Cilia are complex and powerful cellular structures of the respiratory mucosa that play a critical role in airway defense. Respiratory epithelium is lined with cilia that perform an integrated and coordinated mechanism called mucociliary clearance. Mucociliary clearance is the process by which cilia transport the mucus blanket overlying respiratory mucosa to the gastrointestinal tract for ingestion. It is the primary means by which the airway clears pathogens, allergens, debris, and toxins. The complex structure and regulatory mechanisms that dictate the form and function of normal cilia are not entirely understood, but it is clear that ciliary dysfunction results in impaired respiratory defense.
Methods
A literature review of the current knowledge of cilia dysfunction in chronic rhinosinsusitis was conducted.
Results
Ciliary dysfunction may be primary, the result of genetic mutations resulting in abnormal cilia structure, or, more commonly, secondary, the result of environmental, infectious, or inflammatory stimuli that disrupt normal motility or coordination. Patients with chronic rhinosinusitis (CRS) have been found to have impaired mucociliary clearance. Many biochemical, environmental, and mechanical stimuli have been shown to influence ciliary beat frequency, and common microbial pathogens of respiratory mucosa such as Pseudomonas aeruginosa and Haemophilus influenzae have developed toxins that appear to interrupt normal mucociliary function. Furthermore, inflammatory mediators known to be present in patients with CRS appear to impair secondarily mucociliary clearance.
Conclusion
The goal of this article is to summarize the recent developments in the understanding of cilia dysfunction and mucociliary clearance in CRS.

Scanning electron microscopy at nearly 4000× magnification of normal sinonasal epithelium. Cilia are densely packed on the mucosal surface.
The axonemes of motile cilia contain two central singlet microtubules surrounded by nine doublet microtubules (Fig. 2). Each doublet consists of 1 α-tubule, a complete circle of 13 protofilaments, and 1 β-tubule, an incomplete circle of 10 protofilaments. This structure is consistent among the motile cilia of the respiratory epithelium, oviduct, and cerebral ventricular ependymal cells. The two central microtubules are attached by paired bridges and the peripheral doublets attach to the central pair via radial spoke heads. Each outer doublet interacts with the adjacent outer doublets via inner dynein arms, outer dynein arms (ODA), and nexin, each having a distinct role in the dynamic motion of cilia bending. 3 Activation of the dynein arms generates a sliding motion of one microtubule doublet against the adjacent doublet. It is thought that phosphorylation of the ODAs regulates cilia beat frequency (CBF) while phosphorylation of the inner dynein arms regulates the wave form pattern of beating.4,5 Although the function of the radial spoke heads is not entirely understood, it seems they are involved in regionally limiting the sliding between the microtubules during the ciliary stroke, thus converting the sliding motion generated by the dynein arms into a bending motion of the axoneme. 6 In a manner that is incompletely understood, normal cilia remarkably coordinate this bending motion synchronously with surrounding cilia on nearby cells and metachronously with progressively more distant groups of cilia, to generate an elegant and unified fluent motion that transports the mucus blanket out of each paranasal sinus in a reproducible pattern.
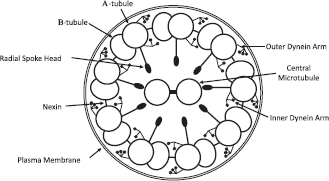
Schematic diagram of the ultrastructure of the axoneme of motile cilia containing two central singlet microtubules surrounded by nine doublet microtubules.
The overlying mucus blanket, produced by goblet cells interspersed among epithelial cells throughout the sinonasal mucosa and submucosal glands, possesses a dynamic gel-like composition in which its rheological properties have a tremendous influence on mucociliary clearance. Mucus is ∼1% NaCl, 0.5–1% free protein, 0.5–1% mucins (carbohydrate-rich glycoproteins), and 95% water. 1 Additionally, mucus contains innate immune proteins such as lactoferrin, lysozyme, and immunoglobulins, which aid in the local immune defenses. 7 The two extremes of the rheological behavior are viscosity (Newtonian fluid mechanics) and elasticity (non-Newtonian). Interestingly, mucus is viscoelastic: it is marked by both viscosity (a liquid property), because of its resistance to flow and its capacity to absorb energy when in motion, and elasticity (a solid property), because of its capacity to store energy that is used to move or deform mass. The physical properties of mucus include spinability, which describes its thread-forming capacity and its internal cohesive force; adhesivity, its ability to bind a solid surface; and wetability, its ability to spread on a surface. All of these rheological and physical properties are influenced by the degree of hydration and the glycoprotein composition, factors that are host regulated.1,8
Over the last 10 years, with the introduction of perfluorcarbon/osmium fixation, 9 detailed transmission electron microscopic examination of airway surface liquid has revealed at least two layers of mucus. The “gel phase” is the outer viscous layer, comprised of high-molecular weight, glycosylated macromolecules, that form a network of tangled polymers ideal for trapping inhaled debris. 10 The “sol phase” is the deeper periciliary layer, lower in viscosity and composed mostly of water and electrolytes. Within the sol phase are mucins that form an apical glycocalyx extending 500–1500 nm from the epithelial cell surface.11,12 The sol phase, both in composition and in size, appears to be critical for proper mucociliary transport in separating the mucus from the epithelial cell wall and membrane.13,14 If the sol phase is too short, the glycocalyx of the cell wall will interact with the gel phase and impair clearance of the mucus blanket.
Each cilium has a forward power stroke followed by a recovery stroke. During the power stroke the cilium is fully extended and the distal tip reaches the viscous outer gel phase of the mucus layer, transmitting directional force to the overlying mucus layer. During the recovery stroke, the cilium bends ∼90° and sweeps back to its starting point within the thinner periciliary sol phase. This process thus creates a unidirectional transport of the outer mucus layer while simultaneously mixing vertically the entire mucus blanket, thereby increasing the efficiency of trapping inhaled debris and microbes. 10
The coordination of ciliary beating is thought to be secondary either to an intracellular calcium wave via gap junctions between epithelial cells that drives microtubule interactions 15 or to a hydrodynamic wave that forces a timed coordination of nearby cilia. 16 Although the mechanism of coordination that results in this remarkable ciliary wave is not entirely understood, it is clear that disease states alter the normal function of cilia, thereby disrupting the critical process of mucociliary clearance.
Furthermore, ongoing research has begun to explore how the structure of mucus itself is also altered by disease states, most likely via disruptions in mucosal ion transport. The significance of normal ion transport for mucociliary health is shown by the pathophysiology present in cystic fibrosis (CF), wherein dysfunctional chloride transport secondary to a mutation of the gene for the protein CF transmembrane conductance regulator results in abnormal mucus and severe mucociliary dysfunction. For example, several groups have indicated that cigarette smoke condensate, in addition to decreasing CBF, also substantially decreases transepithelial transport of chloride.17,18 More recently, investigators have found that this chloride transport inhibition occurs in both the primary chloride conduit, the CF transmembrane conductance regulator, and in secondary transport mechanisms such as calcium-activated chloride channels.19,20 These findings may explain why smokers show airway histopathology similar to that of CF patients, including goblet cell hyperplasia and mucus hypersecretion, and why cigarette smokers have increased rates of chronic rhinosinusitis (CRS).21,22 Furthermore, other investigators have demonstrated that sodium absorption may be increased in sinonasal epithelial cells derived from CRS patients, which would also contribute to increased mucus viscosity. 23 These ion transport mechanisms represent potential therapeutic targets, whereby investigational pharmacologic agents may be used to modify chloride transport and change the mucus structure to facilitate mucociliary clearance.24–26
Dynamic Regulation
Ciliary activity accelerates in response to a variety of mechanical,27–29 chemical,30,31 hormonal,32–34 pH, 35 and thermal stimuli (Table 1).36,37 Extracellular nucleotides (adenosine and uridine) are especially potent regulators of epithelial functions stimulating mucociliary clearance through mucus secretion, increasing CBF, and gating ion channels involved in the maintenance of epithelial surface liquid volume. 38 These nucleotides are released by the epithelium in response to mechanical and osmotic stimuli, and they work in a paracrine fashion through both metabotropic and ionotropic receptors to potentiate mucociliary clearance by recruiting adjacent cells to increase CBF. 38 Furthermore, adrenergic,34,39,40 cholinergic,41,42 and peptidergic43,44 stimulation have also been shown to stimulate ciliary motility. These environmental and host stimuli are transmitted via surface receptors and channels to trigger activation of second messenger cascades that regulate phosphorylation status of ciliary proteins thereby modulating the kinetics of microtubules sliding relative to each other. Recently, elegant experiments using fluorescence resonance energy transfer in primary ciliated cell culture showed direct evidence that activation of protein kinase A (PKA) coincides with an increase in CBF, and that the return to baseline frequency lags PKA inactivation, indicating that dephosphorylation by phosphatases is required to terminate CBF stimulation. 45
Factors influencing CBF

CBF = ciliary beat frequency.
Small changes in both extracellular and intracellular pH can have a profound impact on CBF. An increase in intracellular pH produces an increase in CBF, whereas a decrease in pH produces a decrease in CBF. 35 However, it is not known whether this effect is caused by modulation of kinase activity, even though an acidic pH has been shown to inhibit PKA function, 46 or the direct activation of ODAs of the axoneme. 47 Temperature has also been shown in many investigations to influence CBF, most likely through protein kinase C modulation.,36,37,48 Lower temperature tends to slow CBF. 36
CBF is also regulated by mechanical factors. Direct mechanical stimulation of the cilia promotes an increase in CBF, which coincides with an increase in intracellular Ca2+. 27 It has been shown in models of mouse tracheal mucosa that shear stress applied to the apical surface of mucosa stimulates CBF through an intracellular pathway dependent on purinergic receptors activation as well as [Ca2+] and [ATP]. 28 Thus, these experiments suggest that CBF in the trachea coincides with the respiratory cycle, whereby CBF increases with inspiration and returns to baseline during exhalation to prevent microaspirations. Interestingly, in sinonasal cultures but not tracheal cultures, the application of a sudden apical pressure stimulus, much as would be encountered during a sneeze, stimulates increase in CBF (Fig. 3). 29
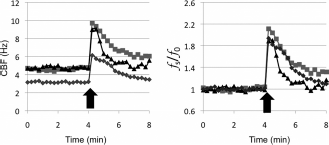
Dynamic regulation of ciliary beat frequency. Air-liquid interface cultures were established from three different mice (▪, ⋄, and δ). Continuous ciliary beat frequency was recorded before and after application of mechanical stimulation (arrow) delivered as an air puff. The panel on the left represents raw data (Hz), and panel on the right normalizes the data to the frequency just before the stimulation.
Acquired Cilia Dysfunction
Although intrinsic factors such as genetic mutations encountered in primary ciliary dyskinesia can alter cilia function with devastating sequelae, extrinsic factors such as pollutants and microbes can also directly and indirectly impact normal cilia function. This phenomenon is evident in patients with CRS who experience relentless cycles of infection and inflammation, resulting in cilia loss (Fig. 4) and a hyperviscous mucus blanket generating dysfunctional mucociliary coupling. In addition to direct ciliary loss, cilia surviving the microbial and/or inflammatory insults appear to not function normally. Although the literature is conflicting regarding CBF and its changes in patients with CRS,49–53 recent work has suggested that a subset of patients with CRS have a blunted ciliary response to environmental stimuli 54 that is reversible once the tissue is removed from the infected or inflamed sinonasal environment. 55 This finding suggests that local exogenous factors can negatively modulate the ciliary dynamic response to stimuli.

Scanning electron microscopy at 1500X magnification of chronic rhinosinusitis sinonasal mucosa showing substantial ciliary loss.
Microbial Factors
Because normal mucociliary clearance is an extremely effective way to combat the continual influx of inspired microbial pathogens, many infectious organisms have developed mechanisms to interfere with and combat this process. Common respiratory bacterial pathogens such as Pseudomonas aeruginosa, Haemophilus influenzae, Streptococcus pneumoniae, and Staphylococcus aureus produce specific toxins to impair ciliary motion and coordination. 56 Viruses responsible for common upper respiratory infections disrupt the microtubule function of ciliated columnar cells and change the viscosity of surrounding mucus. 57 Impairing the local defense system facilitates the infectious pathogens’ upper airway colonization.
P. aeruginosa is a common respiratory pathogen that causes particularly severe infections in patients with a baseline mucociliary dysfunction, such as CF. Patients with CF produce abnormally viscous mucus, and as such, they are even more susceptible to the consequences of ciliary dysfunction. Pyocyanin, the pigment produced by P. aeruginosa that gives it its characteristic blue–green color, is, in fact, a potent factor that aids in the colonization and infection of respiratory epithelium. Although several pathogenic mechanisms of pyocyanin, such as free-radical generation, have been described, pyocyanin has also been shown to cause a progressive and concentration-dependent slowing of human nasal CBF in vitro. 58 It has also been shown that pyocyanin can reduce the velocity of mucus migration on rodent tracheal mucosa, 59 suggesting possible mechanisms for sinonasal mucociliary dysfunction in patients with chronic P. aeruginosa infections. Recently, conditioned media from P. aeruginosa has been shown in vitro not only to depress basal CBF, but also to inhibit rapidly the ability of sinonasal cilia to respond to mechanical stimulation thereby neutralizing the ability of the respiratory epithelium to accelerate mucus clearance in response to infection. 29
H. influenzae, another common respiratory pathogen, has been shown to produce ciliotoxic substances that facilitate the bacteria's colonization of respiratory epithelium. H. influenzae produces lipooligosaccharide and protein D, which have been shown to cause stasis and destruction of cilia and ciliated cells, although the specific mechanisms of these toxins remains unclear. 60 S. pneumoniae produces several toxins including the cytolytic agent pneumolysin and the radical oxidant hydrogen peroxide. These toxins have been shown, both alone and in combination, to cause a dose-dependent slowing of CBF, in addition to damage the of epithelium, on human ciliated epithelium in vitro harvested from inferior nasal turbinate brushings of healthy subjects. 61 In addition, the enterotoxin A produced by S. aureus in high concentrations may also decrease CBF of ciliated sinus epithelium by unclear mechanisms. 62
Inflammatory Mediators
The inflammatory responses found in CRS can be broadly divided into T-helper type 1 (Th1) and T-helper type 2 (Th2) cascades according to the different cytokines they produce, with a predominance of Th1 mediators found in CRS without nasal polyposis and Th2 mediators found in CRS with nasal polyposis. 63 Although no general consensus regarding inflammatory cytokines has been compiled, several molecules are consistently up-regulated in diseased mucosa, established by various techniques including ELISA, reverse transcriptase coupled to quantitative polymerase chain reaction, immunohistochemistry, and multiplex technology. The Th1 cytokines consistently elevated in CRS without nasal polyposis include TNF-α, interferon 7, and IL-8,64,65 while IL-5, eotaxin, and RANTES represent consistently elevated Th2 cytokines in CRS with nasal polyposis.66–69 Furthermore, several of these factors are reduced after treatment with glucocorticoids,64,69 a critical component of CRS medical management.
Although chemokines and cytokines are primarily responsible for inducing migration, differentiation, activation, and degranulation of subpopulations of leukocytes, several studies have reported that cytokines are also powerful modulators of respiratory cilia function. IL-8 has been shown to inhibit isoproterenol-stimulated CBF in bovine bronchial epithelial cells, 70 and, recently, TNF-α has been shown to inhibit viscosity-induced ciliary activity in primary human airway cultures. 71 Furthermore, IL-13 has been shown to decrease basal CBF in human respiratory epithelial cells in a dose- and time-dependent manner, 72 and IL-6 has shown similar effects at high concentrations on human fallopian tube ciliary activity. 73 Conversely, TNF-α and IL-β increase basal CBF in bovine bronchial epithelial cells. 32 Therefore, modulation of cilia physiology by CRS-specific inflammatory cytokines is a likely mechanism for decreased mucociliary clearance in the disease state.
Tobacco Smoke
The effects of tobacco smoke on the mucociliary function of epithelia have been of interest for more than 40 years. 74 Although studies reporting the effects of cigarette smoke on CBF have yielded conflicting results,75–78 histological studies of cilia from the airways of smokers consistently showed decreased cilia number.79,80 Additionally, cigarette smoke increases mucus production by airway epithelial cells,17,81,82 thus necessitating that the epithelium, with fewer and, most likely, dysfunctional cilia, increases the propulsive force to maintain homeostasis. Furthermore, to compound the insult to mucociliary clearance, tobacco-mediated blunting of stimulated CBF has been shown in lower airway epithelium 78 as well as in sinonasal epithelial cultures. 18 Although no clear mechanism has been established to explain the decreased number of cilia in smokers, 79 recent reports have illustrated an inhibition of ciliogenesis by tobacco smoke
Clinical Ramifications
As discussed previously, inflammatory mediators as well as toxins secreted by multiple respiratory pathogens associated with CRS disrupt ciliary function. Although pharmacologic intervention directed at restoring normal cilia function still has to be developed for commercial use, sinonasal saline lavage is an effective surrogate in removing the deleterious compounds in patients with hindered mucociliary clearance and thus aiding in restoring cilia function.
Conclusions
CRS, affecting >35 million Americans of all ages, 84 results in high morbidity, with patients showing worse quality-of-life scores (for physical pain and social functioning) than those suffering from chronic obstructive pulmonary disease, congestive heart failure, or angina. 85 Although multiple etiologies contribute to the development of CRS, ineffective sinonasal mucociliary clearance is the common fundamental pathophysiology. Multiple environmental and exogenous factors alter the normal physiological state, and the resultant inflammatory cytokines secondarily exacerbate the impaired mucociliary clearance.