
Abstract
Background
Intelectin-1 is a new type of Ca2+-dependant soluble lectin in humans that has affinity for galactofuranose in carbohydrate chains of bacterial cell walls, indicating that intelectin-1 may play a role in immune defense against bacteria. The purpose of the current study was to determine the expression of intelectin-1 mRNA and protein and to localize intelectin-1 protein in nasal polyps and tissues from control subjects.
Methods
Normal sphenoid sinus mucosa was obtained from 10 patients undergoing surgery for pituitary tumor. Nasal polyp samples were obtained from 10 patients undergoing endoscopic sinus surgery for chronic polypoid rhinosinusitis. Real-time polymerase chain reaction (PCR) was performed for intelectin-1 mRNA. Immunofluorescent staining was done for localization of intelectin-1 and quantitatively analyzed using computer-based image analysis. Western blot analysis was performed.
Results
Real-time PCR and Western blot analysis showed that intelectin-1 expression in nasal polyps was increased compared with normal sinus mucosa. Using immunofluorescent staining, intelectin-1 was strongly stained in epithelium and submucosa of nasal polyps, and faint staining was found in normal sinus mucosa.
Conclusions
Intelectin-1 is expressed in human sinus mucosa and is increased in patients with nasal polyps. These results suggest a possible contribution for intelectin-1 in the pathophysiology of nasal polyps.
The sinonasal epithelium is open to the extraenvironment. The sinonasal epithelium is exposed to a variety of biological, chemical, and mechanical insults. Innate immune defenses fortify the epithelium by encompassing an array of mechanisms that shield the epithelial barrier. Innate immunity in the paranasal sinuses is composed of mucociliary clearance, pattern recognition receptors expressed on epithelial cells, secretion of inflammatory mediators, and antimicrobial peptides. 2 Additionally, it was suggested that host-derived lipids act as mediators of mucosal immunity. 3 Antimicrobial peptides expressed in the sinonasal epithelium include lactoferrin, lysozyme, cathelicidins, defensins, surfactant protein-A and -D, acidic mammalian chitinase, collectins, serum amyloid A, and secretory leukocyte proteinases inhibitor.4,5
Intelectin-1 is a recently described lectin thought to be involved in the innate immune response and mucosal defense. Using large-scale in situ hybridization screening methods, Komiya et al.
6
isolated a novel cDNA, the mRNA of which is expressed in small intestinal paneth cells in mice, and termed this gene intelectin because the deduced amino acid sequence is similar to the previously cloned oocyte lectin gene of Xenopus laevis.
6
Intelectin-1 has a Ca2+-dependant affinity for
Materials and Methods
Subjects
Nasal polyps were obtained from 10 patients (5 male and 5 female patients; age range, 21–53 years) undergoing endoscopic sinus surgery for chronic polypoid sinusitis. None of these patients had a history of nasal allergies, asthma, aspirin sensitivity, or ongoing drug treatment. The diagnosis of nasal polyposis was established based on medical history, endoscopic findings, and computed tomography scan results. All patients were asked to stop any anti-inflammatory treatment at least 1 month before the surgical procedure. No subjects had an infection of the respiratory tract 4 weeks before the study began. Smokers and subjects who had bronchial asthma that required the regular use of inhaled or systemic corticosteroids were excluded from the study.
Normal sphenoid sinus mucosae were obtained from 10 patients (7 male and 3 female patients; age range, 33–58 years) undergoing pituitary surgery. The subjects were free of nasal symptoms at the time of investigation, and rhinoscopy did not reveal any anatomic abnormalities or signs of mucosal injury. The subjects had no history of nasal infections, allergy, and smoking or ongoing drug treatment. Informed consent was obtained from all patients and control subjects, and the study protocols were approved by the Institutional Review Board of the Korea University College of Medicine.
Tissue Preparation
Tissue samples were cut into two portions; one portion was immediately flash-frozen in liquid nitrogen and stored at -70°C for subsequent RNA processing and Western blotting analysis, and the other portion was fixed with 4% paraformaldehyde overnight at 4°C, and then dehydrated in a graded serial of ethanol-to-xylene and embedded in paraffin wax for immunofluorescent staining.
Real-Time PCR of the Intelectin-1 Gene
Total tissue RNA was extracted with NucleoSpin RNA II (Macherey-Nagel, Düren, Germany) according to the manufacturer's instructions. For real-time reverse transcription (RT)-PCR, 200 ng to 1 μg of total RNAs, was reverse transcribed to cDNA using ReverTra Ace qPCR RT Kit (Toyobo, Osaka, Japan) following the manufacturer's protocols. Quantitative PCR was then performed in a 7300 Real-Time PCR System (Applied Biosystems, Foster City, CA) using 3 μL of cDNA template, 1 nmol of primers, and 12.5 μL of Power SYBR Green PCR Master Mix (Applied Biosystems), in a total volume of 25-μL reaction. The forward and reverse primers were as follows: GAPDH, 5′-GCA AAT TCC ATG GCA CCG T-3′ and 5′-TCG CCC CAC TTG ATT TTG G-3′; and intelectin-1, 5′-CCT GCT GTT TCT CAT AGC G-3′ and 5′-TGC ACT AGG ACA TTC GTC T-3′. The cDNA was amplified with initial denaturation at 95°C for 10 minutes, followed PCR by 40–50 cycles of 95°C for 15 seconds, 58°C for 60 seconds, and, finally, 1 cycle of melting curve after cooling at 60°C for 60 seconds. To confirm amplification specificity the PCR products from each primer pair were subjected to a melting curve analysis. Analysis of relative gene expression was done by evaluating q-RT-PCR data by 2(-ΔΔCt) method. Each experiment was repeated at least three times and GAPDH was used as housekeeping gene for internal control. The levels of gene expression were determined by normalizing relative to GAPDH expression.
Immunofluorescent Staining of Intelectin-1 Protein
Five-μm-thick sectioned tissues were fixed with 4% paraformaldehyde. The tissues were permeabilized with 0.2% TritonX-100 in 1% bovine serum albumin for 10 minutes, blocked with 5% bovine serum albumin for 1 hour at room temperature, and incubated overnight at 4°C with monoclonal mouse anti–intelectin-1 antibody (ENZO Life Science, Plymouth, PA). The tissues were then incubated with Dy-Light 549 horse anti-mouse IgG antibody (Vector Labs, Burlingame, CA) secondary antibody. Finally, coverslips were counterstained with 4′,6-diamidino-2-phenylindole. Each stained tissue was captured and visualized using a confocal z-stack laser scanning microscopy (LSM700; Zeiss, Oberkochen, Germany).
Ten images of the epithelium and submucosa of normal sphenoid sinus mucosa and nasal polyp were captured randomly from the biopsy sections that were immunostained with anti–intelectin-1 antibody at a magnification of X400. We measured the positively immunostained area and the total epithelium and submucosal, and then we expressed the data as the percentage of the stained area. The analysis was performed with Image J (National Institutes of Health, Bethesda, MD).
Western Blotting Analysis of Intelectin-1 Protein
Tissues were homogenized, and the cells were lysed in protein extraction solution (30 mmol/L This-hydrochloride [pH 8.0], 10 mmol/L of editic acid, 1% Triton X-100, 100 mM of sodium chloride, and 2 mM of phenylmethylsulfonyl fluoride) and centrifuged for 1 minute at 13,000 rpm. Proteins were separated by 12% sodium dodecyl sulfate–polyacrylamide gel electrophoresis and transferred to a polyvinylidene fluoride membrane (Millipore, Billerica, MA). The membranes were then blocked for 1 hour in Tris-buffered saline and Tween 20 buffer (20 mM of Tris-hydrochloride [pH 7.6], 135 mM of sodium chloride, and 1% Tween 20) with 4% nonfat dry milk, and incubated at 21°C for 2 hours with monoclonal anti–intelectin-1 antibody. After incubation, the membrane was washed twice for 10 minutes in Tris-buffered saline and Tween 20 buffer, and then treated with peroxidase-conjugated anti-mouse IgG antibody (Vector Laboratories) for 1 hour at room temperature. The membrane was washed and the substrate was added from an enhanced chemiluminescence kit (Intron, Seongnam Si, South Korea) and exposed to x-ray film for 1 minute. β-Actin antibody (Sigma, St. Louis, MO) was used as an internal control.
Statistical Analysis
The Mann-Whitney U test was used to compare the levels of intelectin-1 expression between the nasal polyps and the normal control tissues. Statistical analysis was performed using SPSS 12.0 software (SPSS, Inc., Chicago, IL). The results are expressed as the median and ranges. The values of p < 0.05 were accepted as statistically significant.
Results
Real-Time PCR of Intelectin-1 mRNA
Real-time PCR assay was performed to compare the presence of intelectin-1 mRNA in the control sphenoid sinus mucosa and nasal polyps. A significantly higher level of intelelctin-1 was found in nasal polyps compared with normal sinus mucosa (p < 0.05; Fig. 1).
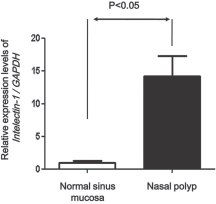
Real-time polymerase chain reaction (PCR) for mRNA for intelectin-1 in the normal sinus mucosa and nasal polyps. The intelectin-1 mRNA was expressed in normal sinus mucosa and nasal polyps. The level of expression of intelectin-1 mRNA was significantly increased in nasal polyps compared with normal sinus mucosa. GAPDH represents the gene for the glycolytic enzyme, glyceraldehyde-3-phosphate dehydrogenase.
Immunofluorescent Localization of Intelectin-1 Proteins
The tissue distribution of intelectin-1 protein was analyzed by immunofluorescent staining. Positively immunostained cells were red in color. The intelectin-1 protein was expressed in epithelial cells and inflammatory cells in control sinus mucosa and nasal polyps (Fig. 2). The expression of intelectin-1 protein in nasal polyps was consistently stronger than normal sinus mucosa. The percentage of the immunostained area in epithelium was significantly increased in nasal polyps compared with normal sinus mucosa (Fig. 3). The percentage of the immunostained area in submucosa was significantly increased in nasal polyps compared with normal sinus mucosa (Fig. 4).
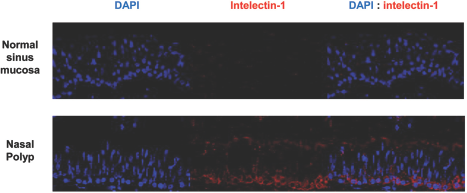
Immunofluorescent staining of intelectin-1. Intelectin-1 protein is expressed in the epithelium and submucosa. The expression of intelectin-1 protein in nasal polyps was consistently stronger than in the normal sinus mucosa (original magnification, X400). DAPI, 4′,6-diamidino-2-phenylindole.
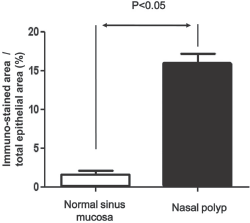
The percentage of the intelectin-1–immunostained area in the epithelium was significantly increased in nasal polyps compared with normal sinus mucosa. The results are expressed as the median and ranges.
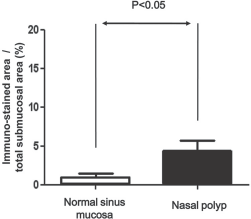
The percentage of the intelectin-1–immunostained area in the submucosa was significantly increased in nasal polyps compared with normal sinus mucosa. The results are expressed as the median and ranges.
Western Blotting of Intelectin-1 Protein
Western blotting showed a prominent immunoreactive band with an apparent molecular mass of 32 kDa reacting with anti–intelectin-1 antibody in nasal polyps (Fig. 5). The expression of intelectin-1 protein in nasal polyps was consistently stronger than normal sinus mucosa, and a less prominent band of the same molecular mass was detected in normal sinus mucosa.

Representative Western blots of intelectin-1 protein expression in normal sinus mucosa and nasal polyps. The number indicates the size of the immunodetected protein.
Discussion
In the present study, we have shown the expression of intelectin-1 mRNA in normal sphenoid sinus mucosa and increased expression of intelectin-1 mRNA in nasal polyps. We also showed that intelectin-1 protein localizes to the sinus mucosa and that this expression is augmented in patients with nasal polyps compared with normal sinus mucosa, particularly in the epithelium. These findings indicate that expression of the intelectin-1 gene in sphenoid sinus mucosa is not only constitutively expressed, but is also increased during nasal polyp formation. The increased expression of intelectin-1 mRNA and protein in nasal polyps suggests a role in abnormal local defense mechanisms of the nasal mucosa in nasal polyps.
A nasal polyp is a manifestation of a chronic inflammatory disease of the upper airway with an unknown etiology. Polyp tissue is infiltrated predominantly by eosinophils, lymphocytes, plasma cells, and mast cells. There is no question that these inflammatory cells have an important role in nasal polyp formation; however, it is not known what recruits these inflammatory cells into nasal polyps. Although several hypotheses have been suggested as the underlying mechanisms of nasal polyps, chronic persistent inflammation and Th2 polarization are considered major factors in nasal polyp formation regardless of the etiology. 9 Indeed, an abnormal host response to the pathogen could be the basis for persistent inflammation. Overexpression or production of antimicrobial peptides could lead to persistent and potentially damaging inflammation.
The immune system is divided into innate and adaptive immunity. T and B cells, the major components of the adaptive immune system, have individually unique receptors for a specific antigen. Binding of an antigen to receptors causes activation and proliferation of the cells. Even though the adaptive immune system is specific and efficient, it takes several days for clonal expansion of these cells. In contrast, the innate immune system protects the host in the early phases of an infectious insult and has completely different mechanisms and receptors used for immune recognition. The innate immune response is mainly mediated by recognition of only a few highly conserved structures present in large groups of microorganisms (pathogen-associated molecular patterns, including bacterial lipopolysaccharide, peptidoglycan, lipoteichoic acids, mannans, bacterial DNA, double-stranded RNA, and glucans). The receptors of the innate immune system for these pathogen-associated molecular patterns are called pattern recognition receptors. There are three classes of pattern recognition receptors (a secreted class that is best characterized by man-nan-binding lectin, an endocytic class that occurs on the surface of phagocytes, and signaling receptors that recognize pathogen-associated molecular patterns and activate signal transduction pathways inducing the expression of a variety of immune response genes, including inflammatory cytokines). 10
Intelectin-1 is a new type of Ca2+-dependant soluble lectin in humans that has an affinity to galactofuranose in carbohydrate chains of bacterial cell walls, indicating that intelectin-1 may play a role in immune defense against bacteria as another secreted class pattern recognition receptor. 7 Intelectin-1 is also known as omentin and intestinal lactoferrin receptor.11,12 Although intelectin-1 was first known to be expressed in the small intestine, several recent studies have shown expression of intelectin-1 in lower airway epithelium. The expression of lower airway epithelial intelectin-1 has been observed in secretory cells with quantitatively decreased expression in smokers. 8 Intelectin-1 expression has been reported to be increased in bronchial epithelial cells from mice allergic to ovalbumin. 13 A recent study showed that intelectin-1 plays a role in interleukin-13–mediated increased expression of the chemokines, monocyte chemotactic protein-1 and -3, and allergic airway inflammation. 14
At present, there are no published studies that have determined the level of intelectin-1 in the upper airway. The current study showed that intelectin-1 mRNA and protein in nasal polyps are increased. Although, the present study can not explain causative relationship between increased expression of intelectin-1 and nasal polyps, we speculate that overexpression or production of this antimicrobial peptide could lead to persistent damaging inflammation and partially contribute to nasal polyp formation by Th2 polarization, as occurs in asthma. Further work is needed to determine whether or not regulation of the level of intelectin-1 can be a target for development of novel therapeutic modalities for nasal polyps.
Conclusion
Intelectin-1 is expressed in nasal polyps and in normal sphenoid sinus mucosa. Intelectin-1 mRNA and protein are more intensively expressed in nasal polyps than in normal sinus mucosa. These results suggest that overexpression or production of this antimicrobial peptide could lead to persistent damaging inflammation.
Footnotes
The authors have no conflicts of interest to declare pertaining to this article