
Abstract
Background
Aspergillus fumigatus and Alternaría alternata are ubiquitous environmental fungal allergens that can exacerbate airway inflammation and contribute to the disease process in patients with chronic rhinosinusitis (CRS). These antigens have been shown to induce human sinonasal epithelial cells (HSNECs) to promote a proinflammatory response, but what is unclear is a means by which to reduce these effects. Inhaled pathogens can induce HSNECs to produce reactive oxygen species (ROS) that trigger cytokine production.
Objective
This study aimed to determine whether the free radical scavenger superoxide dismutase (SOD) could reduce HSNEC-derived inflammation, as measured by interleukin (IL)-6 and IL-8 production, in response to Aspergillus or Alternaria exposure.
Methods
Sinus tissue explants were collected at the time of surgery from control patients (n = 7) and patients with CRS with nasal polyps (CRSwNP) (n = 9). HSNECs were cultured from the explants and treated with Aspergillus, Aternaria, and SOD for 24 hours. Cell supernatants and lysates were collected, and IL-6 and IL-8 concentrations were measured using enzyme-linked immunosorbent assay.
Results
In control and CRSwNP HSNECs, Aspergillus and Aternaria both increased cytokine production (p < 0.05), as measured by IL-6 and IL-8 concentration. SOD treatment reduced the inflammatory response to fungal antigen exposure from CRSwNP HSNECs but not control HSNECs. In CRSwNP patients, SOD significantly decreased IL-6 and IL-8 production after Alternaria exposure and IL-8 after Aspergillus exposure (p < 0.05).
Conclusions
When HSNECs from CRSwNP patients are treated with SOD concurrently with Aspergillus or Alternaria, SOD treatment decreases the fungal antigen-induced inflammatory response. The ability to attenuate inflammation induced by common fungal allergens with SOD treatment could provide a novel therapeutic or preventative approach for patients with CRS or other allergic inflammatory airway diseases.
Fungal allergens are ubiquitous in the sinonasal environment 3 with a controversial role in the pathogenesis of CRSwNP. The presence of fungal allergens has been identified in nearly 100% of both CRSwNP and control patients.4,5 And among the few dozen species of fungus cultured, Aspergillus fumigatus and Alternaria alternata were two of the most common. 4 Conversely, other studies involving CRSwNP patients have identified fungal DNA, including Aspergillus and Alternaria, in polyps of only some patients and found significantly increased numbers of eosinophils in those patients compared with those without fungal DNA. 6 Additionally, in CRSwNP patients, fungal exposure has been shown to increase the secretion of a number of cytokines, including proinflammatory cytokines interleukin (IL)-6 and IL-8. 6 IL-6 has also been shown to be up-regulated in CRS, 7 and IL-8 has been correlated with disease severity. 8 The role of fungus in the pathogenesis of CRSwNP is not completely defined, but a number of studies report a contributory role for fungal allergens in producing inflammation.2,9,10
The human sinonasal epithelial cell (HSNEC) is an important regulator of airway inflammation, because it is the first barrier to environmental allergens and toxins and subsequently produces inflammatory mediators, including cytokines and chemokines that recruit additional inflammatory cells.11,12 Furthermore, HSNECs from CRSwNP patients have been shown to be hyperresponsive to fungal stimulation. 13 A role for reactive oxygen species (ROS) produced by HSNEC as drivers of inflammation has not been well explored but is supported by studies in human bronchial epithelial cells.14,15 ROS include free radicals such as superoxide and hydroxyl radicals, as well as derivatives of oxygen that do not contain unpaired electrons, including hydrogen peroxide and hypochlorous acid. 16 ROS are a product of normal cellular metabolism but in excess can damage important cellular components, including DNA, lipids, proteins, and carbohydrates, leading to cellular dysfunction. 16 Enzymes, like superoxide dismutase (SOD) and glutathione peroxidase, and nonenzymatic antioxidants, like ascorbic acid, are involved in converting ROS to less toxic products. Disruptions in the balance between ROS and antioxidants can lead to cellular damage and inflammation. 17 A number of studies have noted a relationship between free radicals and nasal polyps.17–19
In addition to elevated levels of ROS in patients with CRSwNP, there are also reductions in the levels and function of the antioxidant enzyme SOD,20,21 which is capable of reducing the damaging effects of ROS. SOD is an enzyme with the function of converting superoxide into less reactive peroxide and exists in three isoforms; extracellular SOD, intracellular SOD, and mitochondrial SOD. Studies have shown that intracellular SOD and mitochondrial SOD have decreased transcription and decreased enzyme activity in nasal polyp tissue compared with control tissue. 20 Additionally, certain isoforms of SOD have been reported to show an altered expression profile in polyp tissue, supporting the theory that oxidative damage is a contributor to polyp formation. 21 This study aimed to determine whether SOD could reduce HSNEC-derived inflammation in response to Aspergillus or Alternaria exposure.
Methods
Patients
The Institutional Review Board at the Medical University of South Carolina approved the study protocol before initiation, and written informed consent was obtained for all patients. Patients were included in two groups, CRSwNP (n = 9) and control (n = 7). Patients were included if they were undergoing endoscopic sinus surgery for CRSwNP, diagnosed as defined by the European Position Paper on Rhinosinusitis 2012. 2 Control patients were enrolled if they were undergoing endoscopic surgery for cerebrospinal fluid leak, removal of a nonhormone secreting pituitary tumor, or other skull base surgery and had no radiographic or clinical evidence of inflammatory sinus disease. Patients were excluded if they had received oral steroids or immunotherapy in the four weeks before surgery or had other medical conditions with immune effects, including cystic fibrosis, rheumatoid arthritis, and other immune disorders.
Tissue Procurement and Cell Culture
At the time of surgery, sinus mucosal samples were taken from the osteomeatal complex. HSNECs were cultured and purity confirmed as previously described.13,22,23 Tissue samples were digested mechanically for 30 seconds, then washed and resuspended in basal epithelial growth medium (BEGM). Cells were plated on T-75 cm2 cell culture flasks (Corning Inc., Corning, NY) and incubated at 37°C for two hours to remove contaminating fibroblasts, before transfer to a type IV human collagen (Sigma-Aldrich, St. Louis, MO)-coated T-75 cm2 cell culture flask (Corning Inc.). Cells were expanded for 5–10 days to 90% confluency, then trypsinized and reseeded evenly on six-well human collagen type IV-coated (Sigma-Aldrich) tissue culture plates (Corning Inc.) in BEGM.
Treatment Conditions
Once epithelial cells reached 90% confluency on the six-well tissue culture plates, cells were treated in triplicate with vehicle control (BEGM), A. fumigatus antigen, A. alternata antigen, and SOD from bovine erythrocytes (Sigma-Aldrich). Fungal antigens contained a mixture of mycelial extract and culture filtrate. A. fumigatus is a ubiquitous fungal antigen and the most common fungi in the airway.24,25 Alternaria is similarly prevalent.26,27 We have previously established an optimal dosing of 2 μg/mL for these fungal antigens (range, 0.1–20 μg/mL), which is consistent with other reported in vitro doses of A.fumigatus.28,29 The dose of A. alternata was matched to the dose of A. fumigatus. SOD dosing was based on previously described doses that have been shown to be efficacious and nontoxic to HSNEC. 12
Treatments included each fungal antigen (Greer Laboratories, Lenoir, NC) at concentrations of 2 μg/mL alone, SOD in concentrations of 100 and 1000 μ/mL alone, and combinations of each fungal antigen with each SOD concentration. Cells were then incubated at 37°C for 24 hours. Cells were also pretreated for 30 minutes before the 24–hour treatment, with either BEGM, 100 units/mL SOD, or 1000 units/mL SOD, to match the SOD concentration of the 24-hour treatment. After 24 hours, HSNEC supernatants and lysates were collected and stored at -80°C until use. All doses of A.fumigatus used were confirmed not to induce cell death as determined by LDH Cell Viability Assay (Bioassay Systems, Hayward, CA).
Cytokine Measurement
Cytokine levels in HSNEC supernatants and lysates were quantified using enzyme-linked immunosorbent assay. Enzyme-linked immunosorbent assay kits from BD Biosciences (San Diego, CA) were used to measure levels of IL-6 and IL-8 and were executed according to manufacturer's instructions. Total protein content in HSNEC lysate samples was measured to correct for variable epithelial cell mass within individual tissue culture wells using Pierce BCA Protein Assay kit (Thermo Scientific, Waltham, MA), performed according to manufacturer's instructions.
Statistical Analysis
Statistical analysis was conducted using GraphPad Prism 6.01 software (GraphPad Software, La Jolla, CA). Two-way ANOVA was used to compare baseline differences between control and CRSwNP groups. One-way ANOVA with post hoc Fisher's least significant difference test was used to determine significance in cytokine measurements, concentrations, or percentage change, between the treatment groups. The mean of the vehicle control treatment was compared with the means of the other treatment groups. The means of the fungal antigen treatments were compared with the corresponding fungal antigen with SOD treatment groups. p < 0.05 was considered statistically significant.
Results
There Are Baseline Differences in Cytokine Production between Control and CRSwNP HSNECs
There are baseline differences in IL-6 production between control and CRSwNP HSNECs (Fig. 1). IL-6 production was significantly increased in CRSwNP, as compared with control HSNCEs, when both were treated with vehicle. Differences in IL-8 production were statistically significant only for IL-8 production in lysates (data not shown) but not for secreted IL-8. Cytokine production was not affected in either diagnostic group when treated both with 100- and 1000 units/mL SOD (Fig. 1). Mean concentrations of secreted IL-6 and IL-8 for all treatments are shown for control HSNECs in Table 1 and for CRSwNP HSNECs in Table 2.
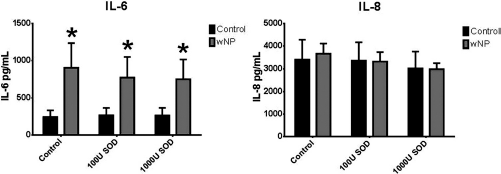
Cytokine secretion after control and SOD treatment of HSNECs derived from control and CRSwNP patients. CRSwNP HSNECs produce more IL-6 at baseline than control HSNEC, whereas IL-8 production is similar between groups. Baseline IL-6 or IL-8 production was not blocked by SOD. *p < 0.05 as compared with control HSNEC. SOD = superoxide dismutase, HSNEC = human sinonasal epithelial cell, CRSwNP = chronic rhinosinusitis with nasal polyps, U = units/mL.
Mean concentrations ∓ standard deviation of secreted IL-6 and IL-8 after various treatment conditions of HSNECs derived from control patients
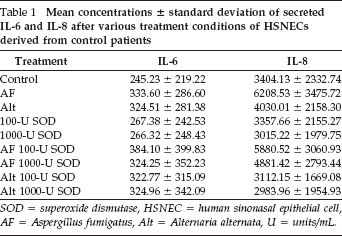
SOD = superoxide dismutase, HSNEC = human sinonasal epithelial cell, AF = Aspergillus fumigatus, Alt = Alternaria alternata, U = units/mL.
Mean concentrations ∓ standard deviation of secreted IL-6 and IL-8 after various treatment conditions of HSNECs derived from CRSwNP patients
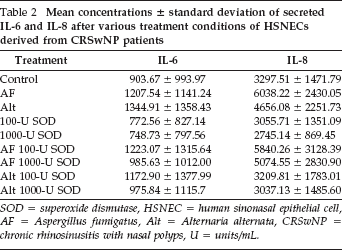
SOD = superoxide dismutase, HSNEC = human sinonasal epithelial cell, AF = Aspergillus fumigatus, Alt = Alternaria alternata, CRSwNP = chronic rhinosinusitis with nasal polyps, U = units/mL.
Fungal Antigens Stimulate Cytokine Secretion in Both Control and CRSwNP HSNECs
Aspergillus and Alternaria stimulate cytokine production in both control and CRSwNP HSNECs (Fig. 2). In control HSNECs, both fungal antigens stimulate IL-8 but not IL-6 secretion. In CRSwNP HSNECs, both fungal antigens stimulate both IL-8 and IL-6 secretion. Results were similar in HSNEC lysates.
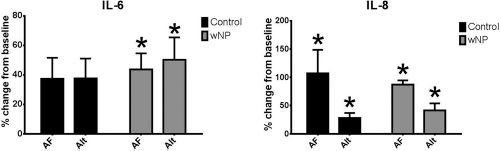
Percent change in cytokine secretion compared with vehicle treatment after fungal treatment of HSNECs derived from control and CRSwNP patients. AF and Alt triggered cytokine production in both groups with the exception of IL-6 in control HSNEC. *p < 0.05 versus vehicle treatment. AF = Aspergillus fumigatus, Alt Alien lana alternata, HSNEC = human sinonasal epithelial cells, CRSwNP = chronic rhinosinusits with nasal polyps.
Fungal Stimulation of Cytokine Production Responds to SOD in CRSwNP, but Not Control, HSNECs
Cytokine secretion in control HSNECs did not significantly respond to SOD treatment (Fig. 3). Only intracellular, but not secreted, IL-8 stimulated by Alternaria in control HSNECs was reduced by SOD. Conversely, cytokine stimulation from fungal antigens in CRSwNP was ameliorated by SOD (Fig. 4). In conjunction with Alternaria, both concentrations of SOD, 100 and 1000 units/mL, reduced both IL-6 and IL-8 production as compared with Alternaria alone. In contrast to Alternaria, only Aspergillus with 1000 units/mL SOD reduced IL-8 production, as compared with Aspergillus alone. Reductions in IL-6 production did not reach statistical significance. Results were similar in HSNEC lysates.
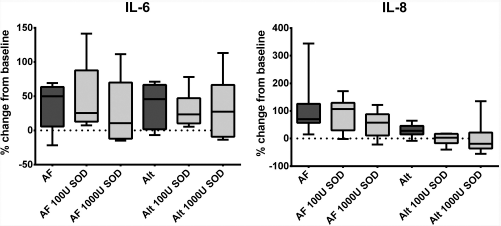
Percent change in cytokine secretion compared with vehicle treatment after fungal treatment and SOD of HSNECs derived from control patients. None of the fungal-induced cytokine production was blocked by SOD. AF Aspergillus fumigatus, Alt = Alternaria alternata, SOD = superoxide dismutase, HSNEC = human sinonasal epithelial cells, U = units/mL.
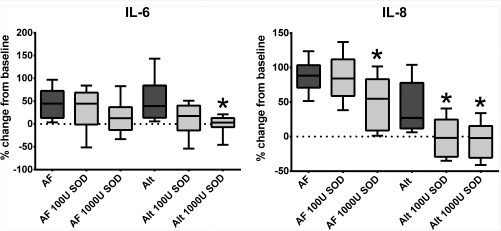
Percent change in cytokine secretion compared with vehicle treatment after fungal treatment and fungal treatment with SOD of HSNECs derived from CRSwNP patients. SOD blocked Alt induced IL-6 and IL-8 production, as well as AF induced IL-8 production. *p < 0.05 versus corresponding fungal treatment without SOD. AF Aspergillus fumigatus, Alt = Alternaria alternate, SOD = superoxide dismutase, HSNEC = human sinonasal epithelial cell, CRSwNP = chronic rhinosinusitis with nasal polyps, U = units/mL.
Discussion
IL-8 and IL-6 secretion was increased in HSNECs from CRSwNP patients when exposed to both A. fumigatus and A. alternata. IL-8 and IL-6 are proinflammatory cytokines that can be produced by the airway epithelium to instigate an inflammatory signaling cascade and have a potential role in the pathogenesis of CRSwNP. 8 IL-8 is chemotactic for neutrophils, eosinophils, and other inflammatory cells, with subsequent infiltrating neutrophils releasing additional IL-8 and attracting more neutrophils.11,14 Neutrophils and eosinophils can release ROS and further potentiate inflammation. Additionally, IL-6 induces T-cell proliferation and activation and stimulates fibroblast proliferation and collagen synthesis, further contributing to the inflammatory process. 8
Baseline differences in IL-6 between control and CRSwNP in this study are consistent with previous reports.7,30 Differences in IL-8 between control and CRSwNP were seen in lysates but not supernatants, although a positive correlation has been found between IL-8 concentration in nasal secretions of CRSwNP patients and the severity of sinus disease, as assessed by Lund-Mackay score. 8 The in vitro model used may not reflect the same conditions that lead to increased IL-8 secretion in CRSwNP at baseline that occurs in vivo. IL-6 is also elevated in nasal mucus in CRS patients, both with and without polyps, and correlates with tissue levels. 7 Additionally, elevations in levels of IL-6 in nasal polyp tissue versus control tissue and alterations in components of the IL-6 signaling pathway in CRSwNP have been reported. 30 Variable response to fungal antigens in control and CRSwNP HSNECs, with increases in IL-6 production in CRSwNP but not controls, is also consistent with reports of increased cytokine production with stimulation of peripheral blood mononuclear cells from CRSwNP patients. 31 IL-8 production was increased in both controls and CRSwNP patients and may be reflective of different stimulation pathways for IL-6 and IL-8.
ROS have been shown to contribute to the inflammatory process in CRSwNP in a number of other studies. Using chemiluminescence, the level of free radicals was noted to be higher in polyp tissue from CRSwNP than mucosal tissue from control patients. Additionally, chemiluminescence level correlated with endoscopy score and the number of eosinophils in polyp tissue. 19 Dagli et al. reported lower levels of antioxidants in both the blood and tissue of CRSwNP as well as increased levels of malondialdehyde-thiobarbituric acid, a product of free radical induced lipid peroxidation. 17 This is consistent with other reports of increased levels of malondialdehyde-thiobarbituric acid in nasal polyp tissue. 18 Additionally, ROS have been shown to cause ciliary dysfunction in human sinonasal epithelium, 32 and oxidative stress has been correlated with the severity of nasal obstruction and congestion in CRSwNP patients. 33
ROS may be produced exogenously, in the form of toxins in cigarette smoke or diesel exhaust particles, or may be produced endogenously. 12 ROS are produced by all cells in the body during regular metabolic processes, by the respiratory burst of activated granulocytes, and potentially by nonimmune cells of the airway for signaling purposes. 16 A role for epithelial cell-driven ROS in the immunopathology of airway diseases is supported by studies in human bronchial epithelial cells and human tracheal epithelial cells that have reported hydrogen peroxide release in response to inflammatory mediators such as phorbol myristate acetate and platelet activating factor.14,15,34 Epithelial cells are targets for airborne microbes, allergens, and toxins but also serve as effectors in the generation of inflammatory mediators that can function in an autocrine or paracrine fashion. 14 ROS have an identified role in modulating transcription factors, including nuclear factor-B and activator protein-1, which induce gene expression of a number of proinflammatory cytokines.16,35
Although ROS were not measured directly in this study, evidence for epithelial cell-driven ROS in the inflammatory process initiated by fungal antigen exposure is provided by the ability of SOD to attenuate the inflammatory cytokine production. SOD appeared to be more effective in reducing inflammation from Alternaria then Aspergillus. Alternaria and Aspergillus may use different mechanisms for causing inflammation, with Alternaria potentially inciting more ROS than Aspergillus. Alternaria has proteases on its surface, which nonspecifically activate protease-activated receptors on the surface of HSNECs and lead to the degranulation of eosinophils.2,36 Possibly, ROS are mediators in this pathway, or other yet to be delineated mechanisms of Alternaria and Aspergillus HSNEC-derived inflammation.
There is therapeutic potential for the treatment of CRSwNP with SOD. SOD has been tested for its therapeutic potential in a rabbit model of chronic allergic asthma, which showed a trend of improvement in airway responsiveness when treated with SOD. 37 SOD has also been shown to inhibit airway inflammation and bronchial hyperreactivity produced by ovalbumin in a murine model of allergy.38,39 Additionally, in CRSwNP HSNECs, SOD reduces inflammation induced by cigarette smoke. 12 Limitations to the therapeutic use of SOD in the airway include the ability to maintain mucosal contact long enough for effective treatment, which is hindered by normal mucociliary clearance resulting in removal of topical agents every 15–20 minutes. 40 The loss of SOD activity at room temperature is also a potential therapeutic barrier. 41
Conclusion
HSNECs are the first barrier to toxins, allergens, and microbes in the sinonasal environment and drive the subsequent inflammatory process with ROS production contributing to the inflammatory signaling cascade. When HSNECs from CRSwNP patients are treated with SOD concurrently with Aspergillus or Alternaria, SOD treatment decreases the fungal antigen-induced inflammatory response, as measured by IL-6 and IL-8 concentration. The ability to attenuate inflammation induced by common fungal allergens with SOD treatment could provide a novel therapeutic or preventative approach for patients with CRS or other allergic inflammatory airway diseases.