
Abstract
The cause of megafaunal extinctions at the end of the last glaciation has been hotly debated during the last few decades, most recently at the global scale. In North America and elsewhere the debate centers on whether or not human hunters, who seemingly first entered the continent during the extinction period, caused the extinctions through over-hunting. An alternative explanation is that climate change during the terminal Pleistocene radically modified existing habitats and this caused the extinctions. Huston's (1979, 1994) dynamic equilibrium model (DEM) of community species richness provides a theoretical context for explanations of the extinctions in North America and highlights life history characteristics of extinct mammals. These life history traits and associated phenotypes are a seldom-explored line of evidence concerning the causes of the extinctions. In light of life history traits, environmental disturbance is implicated as the proximate cause of the extinctions, but the DEM does not preclude overkill as a contributing cause in North America.
Introduction
Explanations for the late Pleistocene extinctions of megafauna (e.g., mastodons, mammoths, camels, horses, and many other mammals) in North America vary between two extremes—those that advocate a central role for overkill by humans (Fiedel and Haynes 2004; Gillespie 2008; Lyons et al. 2004; Martin 1973, 1984) and those that advocate environmental change (Graham and Lundelius 1984; Graham and Mead 1987; Grayson and Meltzer 2003) although there is an expanding middle ground (Barnosky et al. 2004; Koch and Barnosky 2006; Owen-Smith 1987). Evidence to support or reject any particular explanation is neither purely empirical nor purely theoretical (Alroy 2001; Grayson and Meltzer 2003; Guthrie 1984a; Lyons et al. 2004; McDonald 1984; references in Martin and Klein 1984; Surovell et al. 2005). To date, theories that provide test implications for hypothesized explanations are scarce. Often an aspect of ecological theory is borrowed post hoc to support or reject a particular hypothesis (e.g., Brook and Bowman 2005; Graham and Lundelius 1984; McDonald 1984; Owen-Smith 1987). Alternatively, the empirical record is interpreted in diverse ways to allow assignment of a cause to the extinctions (Fiedel and Haynes 2004; Grayson 2001; Grayson and Meltzer 2003; Martin and Steadman 1999; McDonald 1984).
Proponents of overkill commonly argue that the beginning of the current interglacial represented no more severe of a climate change than the preceding ones; thus climate change is not a sufficient cause of the extinctions (Koch and Barnosky 2006; Lyons et al. 2004). On the other hand, those disputing the overkill hypothesis commonly point out that small-bodied taxa unlikely to have been prey also became extinct at the end of the Pleistocene; thus hunting by humans is not a sufficient cause (Grayson and Meltzer 2003; Grayson 2007). Both claims are empirical matters that are more clearly understood when life history evolution and the ecological parameters of extinction provide a theoretical context for the debate. We develop such a context in this paper.
In their attempts to explain the extinctions, archaeologists and paleoecologists have long borrowed frameworks that range from purely descriptive to environmental modeling to purely theoretical. Theory should drive scientific research design and its implementation (Dunnell 1982; Lewontin 1974), but there has been no unified theoretical framework that bridges alternative hypotheses. Framed as a question, one hypothesis is: Did humans hunt North American megafauna to the point of extinction during the late Pleistocene? A second hypothesis is: Did environmental disturbance to habitats associated with deglaciation during the terminal Pleistocene cause the extinctions? A third hypothesis implicates a middle ground between the first two: Were both agents responsible for producing sufficient cumulative stress to cause the extinctions? The third hypothesis is the null hypothesis that must be rejected to accept either of the others.
The life history evolution of taxa that became extinct during the Pleistocene is a critical factor that has received little attention (see recent summary by Koch and Barnosky [2006]; cf. Brook and Bowman 2005; Geist 1987, 1998, 1999; Guthrie 1984a,b; Kiltie 1984). There are several concepts relevant to life history evolution that are rarely considered from a synthetic perspective regarding the late Pleistocene extinctions. Many of these are framed by Geist (1978, 1987, 1998) in a series of books and papers, at least one of which deals directly with the terminal Pleistocene extinctions (Geist 1999). For example, Geist (1998) distinguishes between two types of evolution in large ungulates; evolution resulting from efficiency selection, which produces an efficiency phenotype, in contrast to evolution of novelty, which produces a dispersal phenotype. These are opposites on a continuum of life history evolution; efficiency selection is an increase in fitness through enhancing individual and/or population maintenance. Maintenance refers to energetic constraints of survival, and to be more efficient means to “do more with less” (Geist 1998:7). In the arena of life history evolution, this translates into enhancement of generalist diet, relatively small body size compared to competitors, and investment in high numbers of offspring (r-selection) instead of slower development of few offspring (K-selection). Efficiency phenotypes are fit in food-limited ecosystems.
On the other hand, evolution of novelty leads to specialized adaptations and highly enhanced competitive phenotypes. Geist terms the product of this kind of evolution a dispersal phenotype because though an efficiency phenotype is advantageous in food-limited ecosystems because it can “do more with less,” evolution of novelty occurs in food rich environments. It is perhaps unfortunate that Geist uses the term “dispersal,” but his point is that open habitats into which animals disperse tend to be food rich, which releases individuals from efficiency selection. Dispersal phenotypes can also evolve, however, in extremely energy-rich habitats or in ecosystems in which ungulate populations are constantly held below environmental carrying capacity by predators. An important term that Geist uses to describe dispersal phenotypes is hypermorphy. Hypermorphy, can be defined as extreme growth and specialization resulting from evolution of novelty. In contrast, along the continuum from dispersal to efficiency, Geist highlights paedomorphism as the opposite extreme from hypermorphy. Paedomorphic species represent the ultimate efficiency phenotypes with rapid ontogeny, high reproductive rates, and generalist diets. They waste little energy on specialization, social behaviors, and growth and are fit through efficient use of energy in food-limited ecosystems. We use paedomorphy and efficiency selection/phenotype and in contrast hypermorphy/dispersal phenotype interchangeably throughout the rest of the paper.
Indeed, it is in terms of hypermorphy that Geist's discussion of evolution of novelty is most cogent concerning the North American terminal Pleistocene extinctions. Novel growth (e.g., large body size and antlers/horns), novel adaptations (enhanced running ability for predator avoidance), and novel behaviors (e.g., social herding behaviors to protect young) are enhanced in hypermorphic ungulates. The evolution of specialization (novelty, hypermorphy) requires energy; Geist (1999) accurately describes the glacial environments of the North American Pleistocene to be extremely food rich because glaciers continually reworked existing soils (Huston 1993; Huston and Wolverton 2009) and because a highly evolved predator guild existed that limited populations of large herbivores below carrying capacity. With long term evolution in food rich environments, and with amplification of hypermorphic phenotypes, which indeed most of the extinct megaherbivores were (Guthrie 1984a,b), comes evolutionary risk. The energetic demands of hypermorphy represent an evolutionary tradeoff exchanging phenotypic plasticity, generalist diet, and reproductive maintenance (low energy adaptations) for enhanced competitive abilities that rely on large size, display organs (e.g., large horns), and/or complex behaviors (high energy adaptations). Most species cannot have it both ways, and in the long-term this tradeoff makes hypermorphic species relatively vulnerable to extinction compared to paedomorphs. Close examination of those taxa that became extinct at the end of the Pleistocene in North America and those that survived (see Pleistocene Losers and Holocene Winners below) reveals that hypermorphic evolution, the related energetic tradeoff, and resulting vulnerability to extinction adequately explains why the mass extinctions at the terminal Pleistocene occurred. It is puzzling that this broader context of life history evolution is under-emphasized or ignored in many discussions of the late Pleistocene extinctions.
Multiple studies assume that because taxa present at the end of the Pleistocene were present in North America during the Pleistocene, a period with multiple severe climate oscillations, those same megafauna should have been adapted to severe environmental fluctuations throughout the Pleistocene. This position assumes there was no natural-selection driven evolution during the Pleistocene. We suggest that it is the evolutionary enhancement of highly K-selected phenotypes in megafauna that made them vulnerable to extinction regardless of proximate cause, climate change or overkill (Brook and Bowman 2005; Johnson 2002; Lyons et al. 2004; Van Valkenburgh et al. 2004). Geist (1998, 1999) demonstrates that enhanced hypermorphy, the development of phenotypes including large body size in highly productive environments, evolved during the Pleistocene in response to glacial and interglacial oscillations (see below).
The dynamic equilibrium model (DEM) developed by Huston (1979, 1994) provides a theoretical structure that can encompass multiple causes of late Pleistocene extinctions because it provides a framework for understanding patterns of species richness at multiple spatial scales. The DEM offers a unique perspective on late Pleistocene extinctions and for evaluating the evidence for particular explanations because it accounts for shifts in environmental heterogeneity, changes in habitat productivity, and the role of hypermorphic life history traits –factors important during the late Pleistocene and important for understanding megaherbivores' biological responses to environmental changes. The DEM does not preclude either overkill or climate change as contributing causes. In this paper we first discuss spatial scales at which the extinctions have been debated. We then introduce hypermorphy as a life history characteristic relevant to the extinctions, followed by a discussion of overkill as an archaeological hypothesis. The DEM is then described, and we conclude by identifying alternative factors related to environmental disturbance that potentially caused the extinctions.
A Matter of Scale
Recent studies of Pleistocene extinctions examine the issue at the global scale and highlight correlations between human arrival in an area and the extinction of multiple megafaunal taxa (Lyons et al. 2004; Surovell et al. 2005). These studies demonstrate that overkill is a provocative hypothesis. However, the debate should be considered at multiple taxonomic and spatial scales (e.g., Barnosky et al. 2004; de Vivo and Carmignotto 2004; Grayson 2007; Grayson and Meltzer 2003, 2004; Hill et al. 2008; Koch and Barnosky 2006; Lyons et al. 2004; Stuart et al. 2004).
Lyons et al. (2004; Brook and Bowman 2005) emphasize that the late Pleistocene extinctions appear to be “size-selective”—relatively large-bodied species were most vulnerable to extinction. Johnson (2002), however, found that mammals with relatively low reproductive rates, which do not necessarily correlate with body size, were prone to extinction during the late Pleistocene (see also Brook and Bowman 2005). Johnson also found that a high proportion of surviving taxa are adapted to closed habitats (e.g., forested areas); some of these species are arboreal, others are nocturnal. According to Johnson, these adaptations would have made it possible for these species to avoid overkill by humans. Similarly, in a study of the distribution of body size in African mammals, Kelt and Meyer (2009) argue that African mammal communities are different from those on other continents because the large mammals there avoided anthropogenic extinction through coevolution with humans. In areas of the world where large and/or slow-reproducing megaherbivores survived, it is thought that coevolution with humans diminished prey naïveté. In contrast, North American extinct megaherbivore species are depicted as having been “naïve prey” – that is, poorly adapted to human predators. The “large size- and slow reproducing-selectivity” of the North American extinctions has, thus, been characterized as “compelling evidence that the extinction was precipitated by human activities” (Koch and Barnosky 2006:243). Johnson (2002) and Kelt and Meyer (2009) are advocating a “naïve prey dichotomy”; this dichotomy, however, overlooks the fact that such bimodal body size distributions and arboreal/nocturnal adaptations tend to occur in parts of the world with minimal variation in intra-annual growing seasons compared to the temperate regions where the extinctions occurred (e.g., Geist 1998; Huston and Wolverton 2009; Kiltie 1984; Wolverton et al. in press). We argue below that North American megaherbivores would have had little difficulty avoiding human predation, but they could not have avoided the continental-scale changes in intra-annual growing seasons that occurred at temperate latitudes during the terminal Pleistocene (sensu Kiltie 1984).
At the continental scale, the extinctions do not correlate temporally with climate change or human arrival in an area (Koch and Barnosky 2006; Lyons et al. 2004). Because the overkill hypothesis concerns human interactions with animals, it must be tested with archaeological data (Grayson and Meltzer 2002, 2003; Hill et al. 2008). The blitzkrieg model, which claims that overkill occurred so rapidly that it did not leave an archaeological or paleozoological signature (Martin 1984), is neither testable (Grayson 1984) nor probable from a taphonomic perspective, so we do not consider it further.
A continent-scale consideration of the late Pleistocene extinctions provides increased resolution relative to the global scale. Finer spatial resolution reveals several critical variables (e.g., regional paleoecological and archaeological records) that are less apparent at the global scale (Guthrie 2006; see de Vivo and Carmignotto 2004 [South America]; Grayson 2007 [Eurasia and North America]; Stuart et al. 2004 [Eurasia]; Miller et al. 2005 [Australia]). Grayson (2001, 2007) and others (Koch and Barnosky 2006) have argued for consideration of extinctions taxon by taxon (sensu Stuart et al. 2004; Surovell et al. 2005; Worman and Kimbrell 2008), and here we consider the North American terminal Pleistocene extinctions from an ecological perspective that can be applied on a taxon-by-taxon basis at the continental scale. These perspectives allow the unique cultural, environmental, and evolutionary histories of continents and their inhabitants to be more tightly linked (temporally and perhaps causally) to local extinctions.
Hypermorphy
To recapitulate, hypermorphy refers to phenotypes that, over evolutionary time, devote progressively less energy to reproduction and more energy to the enhancement of physical characteristics such as display and defense organs, social behaviors, often body size, and cursorial (running as opposed to leaping) locomotion (Table 1), all of which ultimately influence competition for mates, predator avoidance, and defense (see Van Valkenburgh et al. 2004 for discussion of carnivore extinction). In contrast, a maintenance/efficiency phenotype occurs where food availability and home-range space per animal are limited; thus maintenance of reproduction is paramount (Table 1). Highly productive habitats associated with the Pleistocene's waxing and waning glaciers resulted in highly developed morphologies related to food-rich conditions; in Geist's (1999:78–89, emphasis added) terms:
Geist's (1998:257) “axes of adaptation” for cervids.
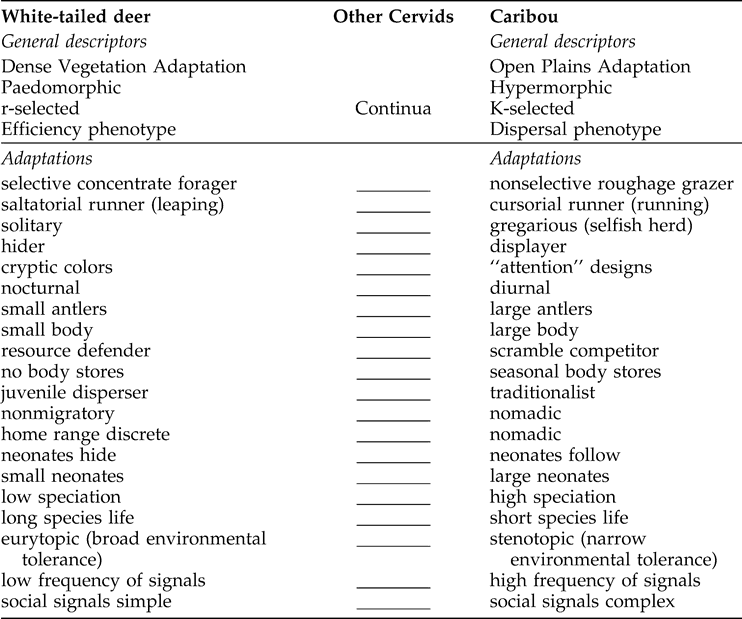
From the late Tertiary onward, a set of remarkable mammals evolved concurrently with the increased severity of seasonally cold habitats. They evolved into hypermorphs characterized by luxurious, often morphologically bizarre growth, associated with progressive colonization of [dispersal in] tropical, temperate, and periglacial, alpine, and arctic landscapes… When nutrients and energy are superabundant, which only occurs in colonization of an uninhabited habitat, a large, luxurious, mobile, strong phenotype develops that, in the process of maximizing reproduction, competes not for material resources, but for mates.
Indeed there is empirical support for Geist's statement beyond skeletal morphology of extinct Pleistocene mammals; analysis of strontium isotopes from teeth of extinct and extant Pleistocene mammals from Florida indicates that migration (dispersal) was comparatively high in extinct herbivores, such as mastodon, mammoth, and horse, but comparatively low in white-tailed deer (Hoppe and Koch 2007). This supports the argument, based in evolutionary biology and functional morphology, that mammals of the terminal Pleistocene were extremely K-selected to the point that they were hypermorphic, a condition that evolved during the Pleistocene (Geist 1999; Guthrie 1984b). The important implication here is that hypermorphic dispersal phenotypes are quite vulnerable to extinction because their reproduction rate is compromised, both because they are extremely K-selected and because of the energetic tradeoff that occurs through evolution of novelty in food-rich environments.
The extinctions might be more accurately characterized as “life history selective” with a tendency for taxa with large body size to meet their end. Indeed, Grayson (2007) points out that only a few small-bodied species became extinct at the end of the Pleistocene. Thus with few exceptions, relatively r-selected competitors (gregarious reproducers) with a range of body sizes survived the extinction (Lyons et al. 2004). This life history selectivity is scaled to taxonomy; for example, in the ungulate guild, r-selected taxa tended to survive. The same selectivity applies to carnivores. We surmise that competitive displacement may have been the causal factor in the extinction of smaller, highly r-selected taxa (e.g., the Aztlan rabbit, Aztlanolagus), a factor that is accounted for in the DEM. Here, however, we focus on body size as a phenotypic characteristic related to reproductive life history, and we acknowledge that data concerning the reproductive life histories of the smallest species that became extinct are limited.
Overkill: An Archaeological Hypothesis
One of the competing extinction hypotheses—overkill (sensu Martin 1973, 1984)—has recently received considerable attention in the ecological literature (Brook and Bowman 2005; Koch and Barnosky 2006; Lyons et al. 2004). The coincident timing of human entrance into the New World (and other continents) and extinctions is interpreted as causal rather than merely correlational. Overkill is primarily an archaeological hypothesis with archaeological test implications related less to the timing of human entry into the New World than to multiple instances of association between artifacts and remains of extinct animals (Grayson 2001, 2007; Haynes 2002; Meltzer 2004). We agree that the earliest Americans “entered the New World [as] skilled hunters with sophisticated weaponry” (Lyons et al. 2004:353). However, such weaponry is only clearly associated with two of the 35 genera that went extinct during the terminal Pleistocene (Grayson and Meltzer 2002, 2003; see Waguespack and Surovell 2003 for a contrary view). Further, only 16 of those genera are known to have been present on the landscape when the first American colonists arrived; the others seem to have been extinct, or were at least quite rare by the time people appeared on the scene (Grayson 2007).
Although Grayson and Meltzer (2002, 2003; Meltzer 2004) provide a conservative estimate for the number of terminal Pleistocene/early Holocene sites where faunal remains of extinct taxa are associated with artifacts (Barnosky et al. 2004; Fiedel and Haynes 2004), we regard their analysis as an important empirical test of the overkill hypothesis in North America (but see Surovell and Waguespack 2008). Their research does not assume a temporal correlation between human arrival and extinction (sensu Martin 1973), it does not use computer-generated simulations (sensu Alroy 2001), and it does not uncritically accept many North American associations of artifacts and faunal remains as kill sites.
Hill et al. (2008) consider the extinctions through an examination of a surviving large ungulate, Bison bison. With a large dataset, they show that bison on the Great Plains decreased in size from the late Pleistocene through the Holocene. Of critical importance in the late Pleistocene extinction debate is that bison exhibited no corresponding shift through time in population age structure. Under harvest pressure, which would have been severe if overkill by humans had occurred, prey populations become progressively juvenile dominated. This response to harvest pressure, particularly among ungulates, is well documented in the archaeological, ecological, and wildlife management literature at multiple temporal scales (Caughley 1977; Festa-Bianchet et al. 2003; Koike and Ohtaishi 1987; Lyman 1987; Munro 2004; Stiner 1990, 1994, 1998; Taber et al. 1982; Wolverton 2008; Wolverton et al. 2008). Hill et al. (2008) demonstrate that diminution of bison occurs independently of increased harvest pressure and represents an ecological response to a decline in habitat productivity (see Wolverton 2008; Crête 1999; Huston and Wolverton 2009). Further, this diminution coincides temporally with changes in terminal Pleistocene/early Holocene archaeological site-distribution and isotopic data from American bison tooth enamel, suggesting that human occupation of the high plains declined during the period of climate change that influenced the distribution and abundance of bison (Lovvorn et al. 2001).
Debate about the cause(s) of the late Pleistocene extinctions in North America should not be reduced to chronological problems rooted in correlations between entrance into the continent and climate change (but see Guthrie 2006). It is the taxa that survive the extinction event, what happens to them after the extinction, as well as the ecological characteristics of the species that became extinct that are critical to explaining the extinctions. Unique aspects of the late Pleistocene that are important include the evolutionary life history characteristics of the megaherbivores. Survivors tend to be species with relatively high reproductive capabilities, and those taxa that became extinct tend to be hypermorphic relative to the survivors. The DEM clarifies why this is the case.
The Dynamic Equilibrium Model: A Theoretical Framework
In the classic sense, competitive equilibrium means the evolution of communities through competitive interactions, including competitive displacement and exclusion (Lotka 1925; Vandermeer 1969; Volterra 1926). With respect to island biogeography, equilibrium refers to a stochastic balance between immigration of new species to an island and extinction of species already on the island (MacArthur and Wilson 1963, 1967). Once equilibrium is reached, community species composition can fluctuate, but richness remains relatively stable (e.g., Simberloff and Wilson 1969). Turnover occurs when a habitat becomes vacant through extinction and is subsequently filled through immigration or speciation of resident taxa.
According to Huston (1979, 1994), competitive equilibrium of species richness within a community rarely occurs because environmental change reduces populations of competing species (see also Connell 1978; Grime 1973; Harper 1967; Hutchinson 1953; Platt and Connell 2003; Pollock et al. 1998; and references in Huston 1994). Huston (1979) proposes that a dynamic equilibrium between two opposing forces explains species richness. One process involves the frequency of population reduction by mortality-causing and/or fertility-reducing disturbances coupled with the rate of population recovery related to a species' biotic potential. For example, changes in the seasonal duration of plant growth (e.g., phenology) disturb the timing of reproductive cues as well as food available for gestation, lactation, and growth in animal offspring and thus resource/food availability potentially becomes decoupled from animal reproductive biology (see below). In contrast to disturbance, the opposing process concerns the rate at which competing species displace or eliminate one another (see an early example in plants by Cowles 1899). Because the late Pleistocene extinctions are an example of changing species richness, the DEM and these processes are particularly salient.
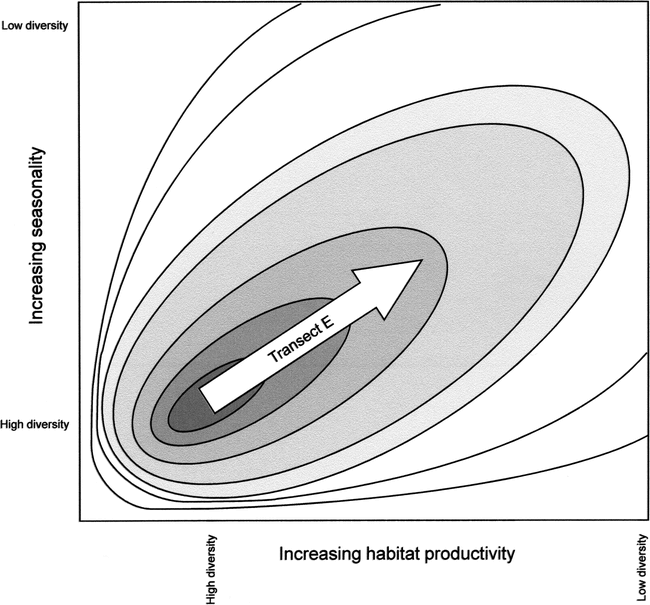
The DEM predicts that, across a landscape, high species richness should exist neither with low or high frequencies of population-reduction events, nor in habitats with high or exceptionally low productivity. High species richness should exist at moderate levels of disturbance in habitats of low to moderate productivity (Figure 1; see also Connell 1978; Pollock et al. 1998). At moderate disturbance levels, highly competitive species experience sufficiently frequent population reductions that they cannot fully displace weaker competitors (r-strategists). A reduction in disturbance frequencies allows highly competitive species with low biotic potential (K-strategists) to recover and to potentially exclude r-strategists (see also Butler and Delacorte 2004).
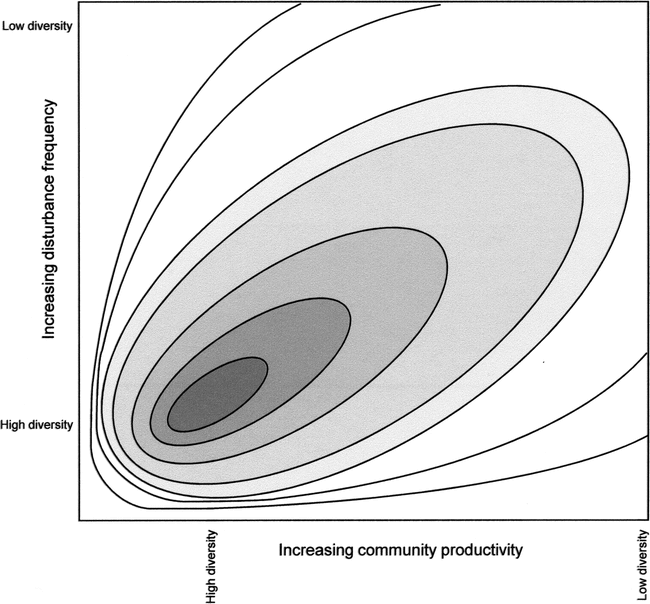
Dynamic equilibrium model of regional species diversity incorporating local and landscape effects (after Huston 1994:149, Figure 5.12). The third axis, diversity, is read as a topographic feature.
An important factor in a species' vulnerability to extinction relative to a competitor's vulnerability is the former's intrinsic population growth rate, specifically, biotic potential (r). If population reductions are too frequent, species with low biotic potential cannot recover fast enough to maintain viable population sizes (Figure 2, Transect A). A community's species richness erodes bit by bit with progressive, frequent population reductions (see also Harper 1967:254). On the other hand, if mortality-causing disturbances are infrequent, an increase in primary productivity can lead to a reduction in species richness (Transect B), because “much of the landscape will be occupied by communities close to competitive equilibrium, with low diversity as a result of competitive exclusion” (Huston 1994:148). Moreover, “species with life histories characterized by slow growth, late reproduction, large size, efficient resource use, and other late successional characteristics will dominate the landscape” (Huston 1994:148).
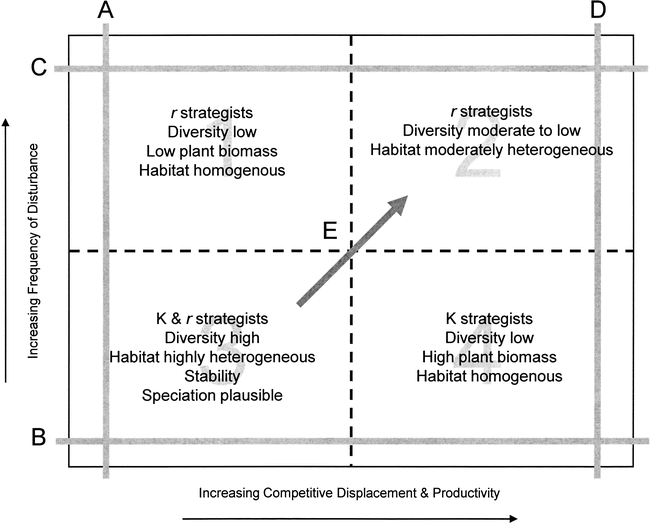
The dynamic equilibrium model and correlated biological responses to disturbance and competitive displacement. The quadrants (1–4) and transects (A–E) are discussed in the text.
Environmental heterogeneity (patchiness of habitats) caused by the interaction of disturbance and productivity varies in a predictable fashion along individual transects (Figure 2). In Area 1 disturbance is frequent and productivity is too low to maintain anything but low species richness thus environmental heterogeneity is low. In Area 2 higher productivity allows plants and animals to recover from frequent population reduction, and thus heterogeneity increases left to right along Transect C. In Area 4 plant biomass is high because of high productivity, but diversity is low because population reductions in plants and/or animals are too infrequent to prevent competitive exclusion; environmental heterogeneity is low because a few highly competitive species dominate. Environmental heterogeneity decreases moving down Transect D. In Area 3 productivity is low-to-moderate and competitive displacement is low, thus most species are not competitively excluded. Further, disturbance is infrequent, and species with low biotic potential can recover from population reductions; richness and heterogeneity are highest in this quadrant.
Figure 2 can be conceptually superimposed on Figure 1 in that the highest species richness and habitat heterogeneity occur in the lower left corner at low frequency of disturbance and low to moderate productivity; richness and heterogeneity decrease rapidly along the x and y axes and decrease slowly toward the upper-right corners of the figures (Transect E). It is important to note that areas in Figure 2 are not discrete; rather continua exist among the biological variables portrayed in each area.
Causes of Extinction and the DEM
An important shift occurs when frequency of disturbance increases (Figure 2, Transects A and D); K-strategists can become extinct and r-selected species can become dominant—a situation we believe is analogous to the transition from glaciation through the terminal Pleistocene into the Holocene (Transect A or E depending on whether or not productivity increased simultaneously [see below]). This illustrates an important distinction between r- and K-strategists in terms of their vulnerability to extinction. Hypermorphic K-strategists are vulnerable to extinction by disturbance-mediated population reductions because their biotic potential is low (Figure 3a), but r-strategists recover rapidly from population reductions. However, r-selected species are poor competitors relative to specialized K-strategists, and through competitive exclusion, they are vulnerable to extinction during periods of high productivity (Figure 3b). This type of extinction process may explain the disappearance of the Aztlan rabbit prior to the last glacial maximum. The fact that the Aztlan rabbit was very small-bodied, was probably r-selected, and became extinct relatively early compared to many hypermorphic mammals suggests that such is the case. It is likely, however, that megaherbivores did not become extinct through competitive displacement and exclusion. Diversity in timing and cause of late Pleistocene mammalian extinctions underscores Grayson's (2007:201) assessment that “[p]iecemeal extinction, and thus complex causation, remains a real possibility for subglacial North America.” Nonetheless, the majority of mammalian extinctions during the period occurred among hypermorphs.
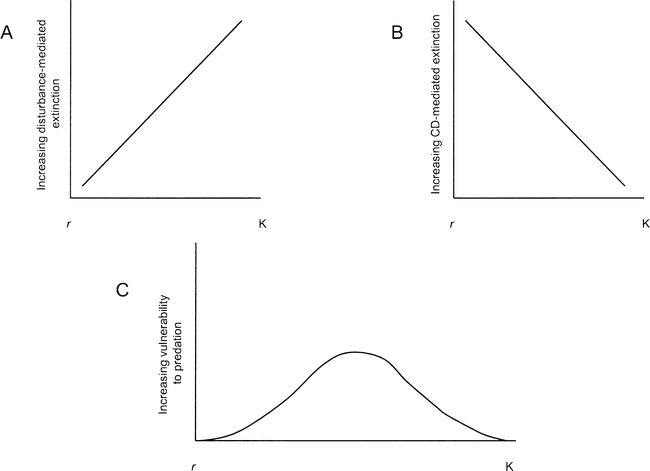
The relationship between reproductive strategy and a) vulnerability to disturbance-mediated extinctions, b) vulnerability to competitive displacement-mediated extinctions, and c) vulnerability to mortality by predation.
Two important relationships of life history evolution are implicit in the DEM. First, the longer the period of high productivity with low disturbance, the longer that intense competition and natural selection for competitive ability occurs. Low r, high K strategists thrive in this evolutionary scenario by becoming increasingly specialized and, thus, more competitive—this equates to evolution of novelty and hypermorphy. It is important to also recognize that the longer the period of high productivity/low disturbance, the more specialized K-strategists are likely to become, and the more vulnerable they will be to extinction from frequent population disturbances (moving up Transect A or up and right on Transect E; see Geist 1998, 1999; Pianka 1970; Southwood 1977, 1988; Van Valkenburgh et al. 2004). Second, anti-predation phenotypes (e.g., enhanced cursorial locomotion, social behavior, and/or large body size) evolve under the same circumstances (Geist 1987, 1999; Pianka 1970); as a result, hypermorphic K-strategists become more vulnerable to extinction due to disturbance but less vulnerable to predation during long periods of high productivity (Figure 3c). An obvious exception to this occurs on islands where K-strategists evolve in predator-free environments (Steadman 2006).
The DEM predicts that low r species (K-strategists) should become extinct through a series of frequent population reductions. It must be determined, then, whether or not the frequency of population reductions from human predation was substantial enough to spiral megaherbivores toward extinction at the end of the Pleistocene in North America. If population disturbance was the cause of extinction, then predation must have reduced all megaherbivore populations (especially of those species that became extinct) frequently enough that they could not recover relative to higher r herbivores, such as Bison bison and Odocoileus sp., that survived the extinction period. In particular, archaeological evidence for human predation must be ubiquitous across megaherbivore taxa, especially those that became extinct (e.g., Hill et al. 2008; Munro 2004; Wolverton 2008; Wolverton et al. 2008). Evidence that population-reducing disturbances were caused by predation must also demonstrate that a diverse range of K-selected megaherbivore species were frequently hunted. To state that overkill occurred without such archaeological evidence is to assume that humans frequently preyed upon all extinct taxa, and preyed upon them all at the end of the Pleistocene. Although it might eventually be shown that these assumptions and presumptions are true, none of them are as yet empirically warranted (Grayson 2007; Grayson and Meltzer 2003; Hill et al. 2008; Meltzer 2004; but see Kooyman et al. 2001).
Were the environmental disturbances related to late Pleistocene climate change frequent and intense enough to reduce populations of megaherbivores beyond the point of recovery? To answer this, we must evaluate the evidence for profound changes in frequency and magnitude of environmentally-mediated disturbances at the end of the last glaciation when the megaherbivores went extinct (see Discussion section). Signatures of these events should include evidence for the selection of hypermorphic K-selected phenotypes among extinct fauna, increases in environmental heterogeneity and in population-reducing disturbances through time, as well as the survival of relatively r-selected herbivores after the extinction period (following Transects A and E). In sum, the ultimate cause of the extinctions lies in the evolutionary ecology of the extinct taxa. Addressing the ultimate cause may lead to improved understanding of proximate cause(s) such as human predation and climatic change. The evolution of hypermorphy among species that became extinct at the end of the Pleistocene is germane to understanding the ultimate and proximate causes of the extinction event. We set up the role of hypermorphy as a hypothesis that potentially explains the North American terminal Pleistocene extinctions below.
The Hypermorphy Hypothesis
The DEM predicts two proximate causes for extinction: 1) population decline caused by environmental disturbance followed by poor recovery and/or 2) competitive displacement and exclusion of poor competitors (relatively r-selected) by good competitors (relatively K-selected). Traditional hypotheses regarding the terminal Pleistocene extinctions suggest that population decline was produced by either human predation or environmental disturbance, which reflects change on the y axis of the DEM (Figures 1 and 2). Consideration of life history evolution as a causal factor in the extinctions integrates the x and y axes. This is particularly the case for the evolution of hypermorphy because it responds to changes in habitat productivity (Geist 1998; Worman and Kimbrell 2008).
The hypothesis of hypermorphy evolution has several implications for the cause of the extinctions. First, if enhanced hypermorphy increased susceptibility to extinction, then the surviving taxa should be those with life history characteristics that provide an ability to recover from population reductions. Second, if hypermorphy evolved during the Pleistocene, the preponderance of large body size, luxury organs, and/or cursorial morphology should be relatively high during the late Pleistocene compared to earlier in the Pleistocene (sensu Geist 1987). Third, disturbances that cause extinction of hypermorphs should negatively impact dispersal phenotypes and favor gregarious r-selected taxa. Fourth, if dispersal (food availability) relates to large body and antler/horn size and contributes to the evolution of hypermorphy, then modern phenotypes should be relatively hypermorphic today in areas that are food-rich, whereas efficiency phenotypes should occur in areas that are comparatively food-limited. Of these test implications, we examine the first in reference to ungulates in the next section of the paper (see also Kiltie 1984); the second has been examined for cervids and other ungulates (Geist 1987, 1998, 1999; Guthrie 1984a, b); the third implication has been addressed in studies of body size diminution among surviving and extinct taxa (Graham 1991; Purdue 1989; Schultz et al. 1972; see below); and we address the fourth using data on white-tailed deer (Odocoileus virginianus) from central Texas in the Dispersal and Efficiency Selection in White-tailed Deer section below.
Pleistocene Losers and Holocene Winners
Southwood (1977) explicates several options that organisms have in terms of life history when they encounter new environments either due to spatial variability or temporal change. In general, organisms can use energy in a variety of ways, for example to reproduce, eat, or rest here-now, here-later, there-soon, or there-much-later, but each option requires a difference in energy expenditure. Probability of survival relates to fitness, which relates to evolutionary biology; that is, what particular suite of adaptations can a particular individual of a species rely upon to address the spatio-temporal challenges of survival? Birds migrate, for example, to take advantage of highly productive growing seasons at high latitudes. This represents a there-much-later reproductive strategy that requires particular morphology resulting from the shaping of evolutionary history. There is diversity in fitness among different migrating bird species, each carving out its own niche related to its evolved life history strategy. In this complex evolutionary matrix, life history theory predicts that species with a low biotic potential are susceptible to extinction if their populations are frequently reduced because they cannot replace themselves rapidly enough (Cole 1954). Were extinct Pleistocene megaherbivores low r species that were vulnerable to extinction by population reduction? There is a strong relationship between the biotic potential of a species and its age at first reproduction and gestation length (Birch 1948; Cole 1954; Peters 1983). Although it is impossible to determine the ages at first reproduction or lengths of gestation for extinct megaherbivores, it is possible to use analog species to infer such variables (Table 2; Guthrie 1984a; Kiltie 1984; McDonald 1984), and these analogs can be compared to herbivore species that survived the late Pleistocene extinctions. North American ungulates that survived the extinction period have relatively short gestation periods (none longer than 10 months) and early ages at first reproduction (none older than 3 years of age) (Figure 4). Extant large-bodied mammals in other parts of the world tend to be those with lower reproductive rates and older ages at first reproduction (Figure 4; Calder 1984; Peters 1983). McDonald (1984) and others (Brook and Bowman 2005; Guthrie 1984a; Johnson 2002; Kiltie 1984; Koch et al. 1998) demonstrate that it is likely that Pleistocene megaherbivores were K-strategists with long gestation periods and relatively low biotic potential. Several extant Old World ungulates are similar in terms of body size to species that became extinct in North America at the end of the Pleistocene and can serve as analogs. Figure 5 is a bivariate scatterplot of gestation length (x axis) against body mass (y axis) comparing several extant North American ungulates to the extinction analogs (data are in Table 2). In terms of body size, there is separation between extinction analogs and North American survivors on the y axis, and there is complete separation in terms of gestation length on the x axis. These data indicate that relative to the extinct megaherbivores, surviving taxa have higher biotic potential and tend to be r-selected (Figures 4 and 5).
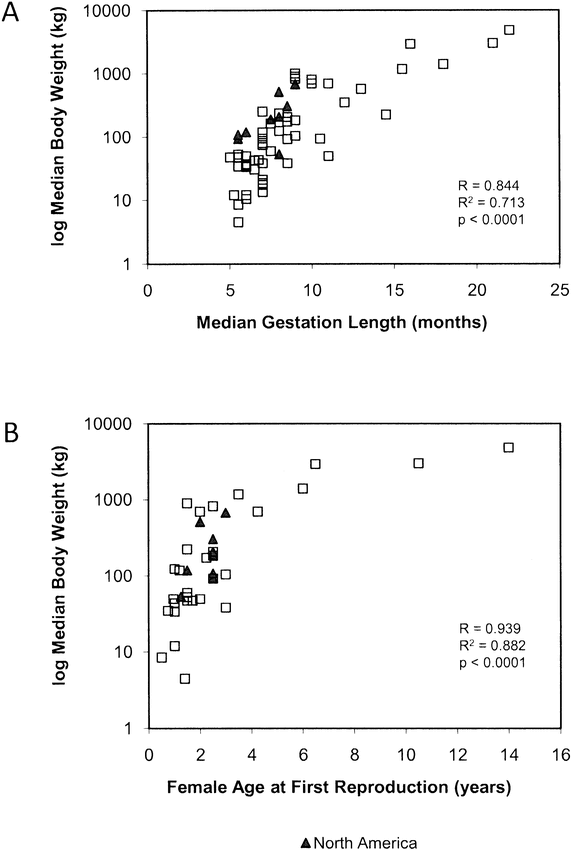
A) median (range midpoint) gestation lengths of extant North American ungulates relative to those of species from other parts of the world ([open squares] primarily Africa). B) the same relationship as in A in terms of age at first reproduction. The graphs highlight the relatively high biotic potential and small body size of North American ungulates that survived the extinction period (data from Nowak and Paradiso 1983).
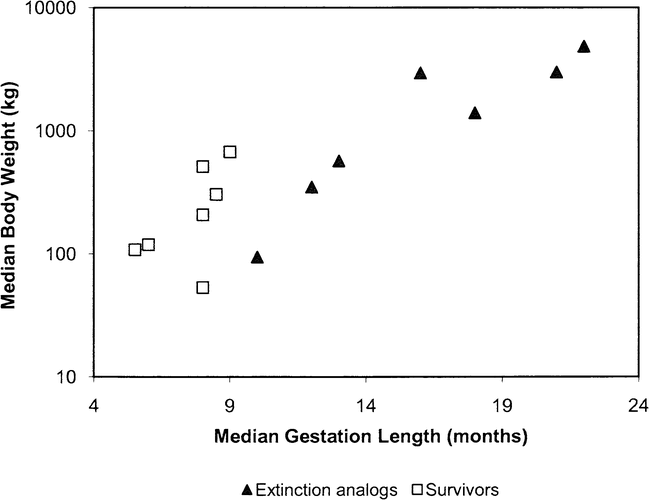
Bivariate scatterplot of median (range midpoint) gestation-lengths versus median (range midpoint) body size for several North American ungulates that survived the extinction period and extinction analogs from other parts of the world that survived in seasonally equable environments (data from Nowak and Paradiso 1983).
Gestation length and body mass for large mammalian herbivores (data from Nowak and Paradiso 1983).
The evolutionary biology of the extinct megaherbivores and of surviving ungulates supports the hypermorphy hypothesis. The extinction event “left behind the specialists in noncompetition [in terms of exclusion]. Examples of such include mobile r-strategists, that is, species with high reproductive rates, short, individual life expectancies [gestation periods], and excellent mechanisms of juvenile dispersal, such as white-tailed and black-tailed deer, pronghorn, peccaries, and coyotes” (Geist 1999:84 emphasis added). This follows closely the predictions of the DEM (Figure 2, Transect A or E). Further, Holocene lineages of Pleistocene fauna (e.g., wolves, bears, bison) exhibit a reduction in body size suggesting that diminution among r-selected species was advantageous (e.g., Gordon 1986; Graham 1991; Guthrie 2003; Hill et al. 2008; Lyman 2004; Purdue 1989; Schultz et al. 1972). Indeed, members of at least one taxon, Equus, (Guthrie 2003) exhibit diminution prior to extinction. The overkill hypothesis is not supported by these data because if increased predation had been a factor, there would have been a decrease in population density and intraspecific competition, which would have increased ontogenetic growth rate and adult body size (Kie et al. 1983; Wolverton et al. 2007; Wolverton 2008; Wolverton et al. in press).
Why did bison and white-tailed deer/mule deer survive in North America while horses became extinct at the terminal Pleistocene? The answer lies not just in analysis of body size, but in consideration of hypermorphy and its implications for reproduction, such as the relationship between hypermorphy and lengthy gestation. For example, gestation length in the modern Burchell's zebra (Equus burchelli) is 12 months, for mule deer (Odocoileus hemionus) it is 6 months, and for the American bison it is 9 months. In terms of reproductive ecology, contingencies for survival into the Holocene were less flexible for the horse, despite its diminution, than for mule deer and bison. We discuss the implications of seasonality of growth and reproductive seasons below (see Seasonality and the DEM).
Analysis of stable carbon isotope data from animal tooth enamel indicates that diets of extinct mammoths (Mammuthus) and horses and surviving bison overlapped (Connin et al. 1998; Feranec and MacFadden 2000; Koch 1998; Koch et al. 2004), and that these taxa are “intermediate feeders with a strong preference for grasses and sedges” (Feranec 2004:364). These species represent distinct body sizes. If their diets overlapped, what led to the demise of mammoths and horses but not bison and white-tailed deer/mule deer? In addition to reproductive ecology discussed above, part of the answer lies in the mammalian herbivores' dietary adaptations that are not registered in stable isotopes. Resource partitioning may occur in competing herbivores with similar diets. For example, white-tailed deer are selective concentrate feeders that preferentially browse high quality portions of plants, such as highly nutritious new growth. They are “by no means ‘super ruminants’—they cannot use some woody browse species as well as cattle [or other Old World ungulates] can” (Verme and Ullrey 1984:111). This is so pronounced that in areas of North America, such as south-central Texas, where free-ranging Old World fallow deer (Dama dama) and sika deer (Cervus nippon) share habitat with the native white-tailed deer, the exotic species more efficiently digest native forage than does the white-tailed deer (Demarais et al. 2003). When white-tailed deer switch to less nutritious forage, their body size declines considerably (Simard et al. 2008; Tremblay et al. 2005). In terms of digestion efficiency, white-tailed deer would have been at a disadvantage during the late Pleistocene leading to their competitive displacement by other herbivores despite overlap in diet. With the extinction event, however, competitive release occurred, and this generalist, left alone on the landscape, has thrived. The white-tailed deer's generalist dietary adaptation allowed it to survive competitive displacement on the margins when the North American late Pleistocene ungulate guild was crowded with specialists and to thrive in the absence of competition once those specialists became extinct.
Not all extinctions are created equal, and ungulates are a highly diverse group. For example, compared to probicideans and horses, bison and white-tailed deer have shorter gestation periods. In addition, the white-tailed deer is not only an r-selected species, but it is highly generalized in terms of diet, which enhances its non-migratory, low-dispersal, highly philopatric (overlapping with maternal range through adulthood), maintenance phenotype (Comer et al. 2005; Geist 1998; Purdue et al. 2000). Despite its poor digestion efficiency, the white-tailed deer thrives by switching foods in poor-quality habitat (Tremblay et al. 2005) and through potentially dramatic diminution in body size (Kie et al. 1983; Wolverton et al. in press). The plasticity of this phenotype is highly advantageous should environmental disturbance increase—hence the ability of Odocoileus to thrive in nearly all parts of the New World today. Species without such adaptive flexibility and phenotypic plasticity are at greater risk of extinction during periods of environmental stress. It is likely that in some species, body size, reproductive ecology, and dietary adaptations shifted considerably during the Pleistocene. Together, however, these three realms of ecology and evolution converge as ecological energetics. It is also likely that an evolutionary threshold was crossed late in the Pleistocene affecting a variety of evolutionary processes and adaptations that resulted in the extinction of some species and the survival of others in just the piecemeal fashion described by Grayson (2007).
Dispersal and Efficiency in White-tailed Deer
The evolution of dispersal phenotypes can occur over short time spans such that factors relating to the evolution of hypermorphy can be easily observed. Data collected from deer from 2004 to 2008 at Fort Hood, Texas illustrate the relationship between resource availability and phenotypes. Population density, habitat quality, and body and antler size data indicate that significantly larger body size and antler size occur in areas where dispersal is possible (Table 3)—that is, in food-rich areas with relatively low deer population densities. West Fort Hood (WFH) is somewhat isolated from the adjacent West Region (WR) by the town of Killeen and by US Highway 190, but the two areas comprise oak-juniper habitat with continuous soils and similar vegetation (Teague and Reemts 2007). Although the taxonomic composition of vegetation is similar between WFH and WR, the proportion of poor quality (food limited) white-tailed deer habitat is higher on WFH than WR (Wolverton unpublished data; Teague and Reemts 2007). In addition, population density of white-tailed deer has been historically high in WFH relative to other areas of the fort (Figure 6), harvest rates by sport hunters have been relatively low, and deer in this area have small bodies and antlers (Figure 7; Table 3). In contrast, WR has a relatively low population density related to higher harvest rates and contains a higher proportion of food rich habitat. In Geist's (1987, 1998) terms, white-tailed deer in WFH exhibit an efficiency phenotype in food-limited conditions, and the deer in WR represent, relative to deer in WFH, a dispersal phenotype in food rich habitat. In a relatively short period, a hypermorphic (dispersal) phenotype of deer has begun to evolve in WR an area with relatively high food availability. This supports the notion that hypermorphy evolves under conditions with high nutrient availability, a setting Geist (1987, 1999) ascribes to the Pleistocene evolution of megaherbivores. This pattern has been observed at several spatial and temporal scales in cervids including other studies on white-tailed deer, moose (Alces alces), caribou (Rangifer tarandus), and elk (Cervus elaphus) (Ferguson et al. 2000; Kie et al. 1983; McCullough 1979; Reimers et al. 1983; Sand et al. 1995; Simard et al. 2008; Teer et al. 1965; Teer 1984; Tveraa et al. 2007; Wolverton et al. in press).
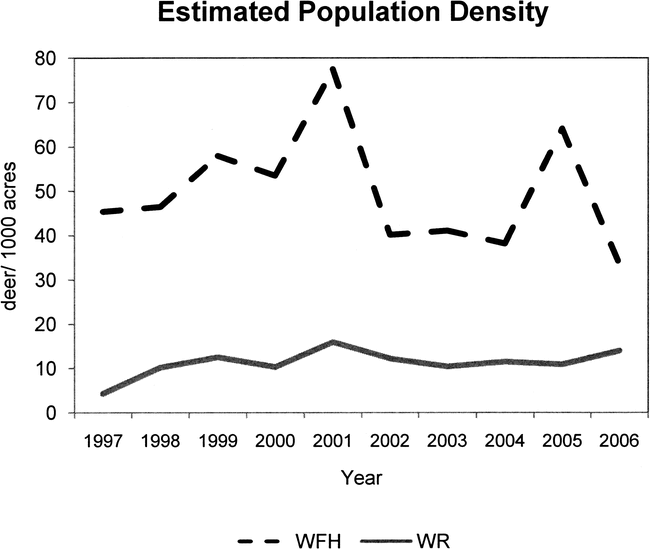
Estimated white-tailed deer population density in two management areas of Fort Hood, central Texas. Population density is estimated using spotlight surveys each year to determine harvest quotas for management areas, and WFH is consistently more densely populated with deer.
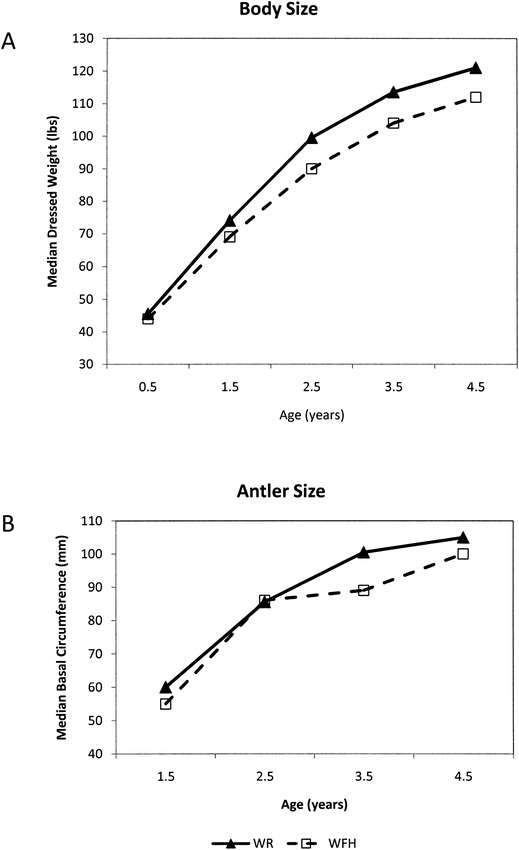
a) Median body mass (field dressed weight) of white-tailed deer bucks in WFH and WR. b) Median basal antler circumference for bucks in WFH and WR. Deer are consistently larger in mass and antler size in adjacent, less densely populated regions than in WFH.
Body mass and antler size data for white-tailed deer from Fort Hood, Texas.
Summary
We have argued that hypermorphic competitors (e.g., K-strategist megaherbivores) evolved their unique dispersal phenotypes during extended pulses of high growing season productivity in periglacial environments during the Pleistocene. These high productivity periods would have produced “conditions favorable to maximum body growth” (Geist 1999:89; see also Guthrie 1984a, b) that culminated in the late Pleistocene. During these periods megaherbivores experienced intense competition, which selected for dietary specialization, specific defense mechanisms (e.g., cursorial locomotion), large body size, and luxury organs (e.g., large antlers and horns). The Pleistocene would have been an extended period of fluctuating competitive displacement among ungulate species (moving back and forth along Transect B of the DEM). An extended period of competitive displacement eventually reached a time when large-bodied K-strategists were vulnerable to population reductions. The remains of white-tailed deer, a highly r-selected cervid, are low in abundance and comparatively rare in late Pleistocene faunal assemblages (Ferring 2001; Geist 1998) and become increasingly common and abundant in Holocene ones (e.g., Baker 1998; McMillan and Klippel 1981; Wolverton 2005; Wolverton et al. 2008). This suggests that this generalist was competitively displaced prior to megaherbivore extinction and was released from competition thereafter—a conclusion that makes sense in light of modern wildlife biology.
Prior to the terminal Pleistocene, megaherbivores may not have been far enough along the hypermorphic evolutionary trajectory to have been vulnerable to extinction because of disturbance, or the disturbances may not have been sufficiently frequent or severe enough to cause extinction. This unique aspect of the megafauna's evolutionary histories culminated in the terminal Pleistocene and relates directly to the extinction event. The late Pleistocene appears to have been a period of high productivity combined with an increase in population-reducing disturbances (Transect E); the high productivity would have promoted the competitive displacement begun earlier in the Pleistocene and the disturbances would have caused the extinctions of hypermorphic mammals. Different species had different ecological energetics (Table 2), thus all extinctions would not have occurred at precisely the same time. This implication conforms to the empirical record of terminal dates for multiple species that became extinct near the end of the Pleistocene (Grayson 2007).
Discussion
How do the implications of the DEM and hypermorphy relate to the hypothesized overkill and climate change causes of extinction? Overkill implies that humans were super-predators and that extinct megaherbivores were naïve prey. It has also been argued that habitat productivity increased during deglaciation, which is thought to weaken the case for climate change. We discuss these arguments in detail below and summarize other explanations that are relevant to the extinction debate in light of the DEM and hypermorphy.
Megaherbivores as Prey
Vulnerability to extinction is not the same as vulnerability to predation (Figure 3). Those species most susceptible to population regulation through predation are species exhibiting reproductive adaptations that are intermediate between r- and K-strategists, which Southwood (1977) terms reproductive polymorphs (Figure 3c). K-strategists, with the exception of those that evolved on predator free islands (Nagaoka 2002a, 2002b; Steadman 2006), are less vulnerable to predation at the individual-phenotype scale because individuals tend to have defense adaptations that co-evolved with their reproductive strategies (Owen-Smith 1987), one of which could be large body size (see above). According to Southwood (1977:352–353, emphasis added),
The combination of characters at each extreme [K versus r strategists] will lead to different forms of population dynamics. The role of predators at both extremes will be small, at the r-end because of the organisms' high mobility, at the K-end because of defense mechanisms… However, for species that are intermediate in the continuum [e.g., among cervids, white-tailed deer], predators are important and, taken with other forms of interspecific competition, frequently maintain a population equilibrium below the carrying capacity as determined by food and space resources. Successful biological control [of predators] and the outbreak of ‘upset pests’ following the destruction of predators provide many field examples to support these theoretical conclusions.
The most abundant survivors of the late Pleistocene extinctions in North America, white-tailed deer and mule deer are relatively r-selected ungulates. In fact, today in the absence of substantial predation in eastern North America, white-tailed deer have become “upset pests” (Côté et al. 2004; references in McShea et al. 1997). Although ungulates tend to be large-bodied K-strategists (Wemmer 1997), both species of Odocoileus fall at the r-strategist end of the cervid continuum. As reproductive polymorphs, their populations are likely to be regulated through predation. At the end of the Pleistocene, the ungulate species theoretically least vulnerable to the effects of predation became extinct, yet those predicted to be most impacted by predation survived. Undoubtedly, reproductively polymorphic ungulates survived the extinction period because they were able to recover from any form of population reduction given their higher biotic potential.
Proponents of overkill stress that K-selected megaherbivores would have been relatively vulnerable to extinction compared to other herbivores with greater biotic potential (e.g., Alroy 2001; Brook and Bowman 2005; McDonald 1984), but such K-selected megaherbivore species likely possessed effective defense adaptations related to their co-evolution with sizeable predator guilds (see discussion in Grayson 2001; Owen-Smith 1987). Further, the overkill argument does not take into account the fact that extinct terminal-Pleistocene mammals were not simply K-selected, they were hypermorphic—their phenotypes were pushed to an extreme in terms of display, defense, and body size through evolution leading up to the terminal Pleistocene (Geist 1987, 1999; Guthrie 1984b). The skeletal morphologies of several ungulate and carnivore taxa indicate an increase in hypermorphy culminating at the end of the Pleistocene (see above). Because there is little archaeological evidence for human predation on megaherbivores (other than the proboscideans) and no evidence of prey depression associated with predation, such as shifts in prey population age structure and body size, the implications of hypermorphy evolution in extinct species are important (Grayson and Meltzer 2003; Hill et al. 2008).
Our discussion of predation vulnerability is enmeshed in two competing assumptions, one ecological and one archaeological. First, we assume that because hypermorphic phenotypes constitute defense morphologies and behaviors, megaherbivores in particular would not have been especially vulnerable to human predation. Thus we believe that late Pleistocene fauna were effectively not naïve to predation by humans. There are no empirical data to support this belief, but general life history characteristics of hypermorphic phenotypes suggest it is plausible. Second, as discussed earlier, it is often assumed the late Pleistocene North American humans were super-predators with highly effective hunting technology and behaviors (Lyons et al. 2004; Martin 1973). This assumption may be tested through analysis of artifact function, but more importantly it can be assessed by examining the distribution and number of extinct-megaherbivore kill sites (Grayson and Meltzer 2003) and by examining evidence for harvest pressure on prey (Hill et al. 2008). That is, overkill must be treated as a hypothesis that relies on archaeological evidence of the association between hunting weaponry and remains of extinct animals. To date, such evidence is quite rare and, at best, mounting.
Habitat Productivity during the Terminal Pleistocene
McDonald (1984) and others (Ferring 2001; Lyons et al. 2004) argue that environmental change was unlikely to have been an important factor in the extinction of megaherbivores because habitat productivity appears to have increased during the late Pleistocene (Figure 8). McDonald's paper introduces a quandary in the context of our discussion; he (1984:404–409) suggests, “the combination of extensive deglaciation, warmer mean air temperatures, and ample available moisture over much of the continent created a physical environment which substantially increased North America's primary productivity” and that as a result “modern ecological and evolutionary theory… predicts that most changes in the North American environment during the period of extinction should have resulted in an increase in megafaunal biomass and perhaps even in diversity” (Figure 8). This prediction is in line with the DEM in that high productivity at low disturbance frequency favors K-strategists (Figure 2, Transect B). McDonald's map (Figure 8) indicates that central and northern North America is where productivity is thought to have increased. This distribution fairly precisely overlaps with the ranges of two of the comparatively K-selected ungulates that survived into the Holocene, bison and caribou. However, McDonald's prediction also implies that productivity did not increase so much that r-selected species sensitive to extinction through competitive exclusion were wiped out (Figure 3b). We note that habitat productivity and environmental disturbance may increase simultaneously and produce a reduction in diversity (Figures 1 and 2). It is important to distinguish the life history effects of changes in productivity from population reducing effects of environmental disturbance. Moreover, although McDonald suggests there was an increase in habitat productivity, an increase in biomass is not equivalent to an increase in highly nutritious, edible forage (Graham 1986). In fact, Graham determines that highly nutritious forage was in decline by the late Pleistocene. Hypermorphs would have been vulnerable to population disturbances (whatever the proximate cause) even if productivity increased after the last glacial maximum. Below, we postulate one such disturbance that may have contributed to the extinction.
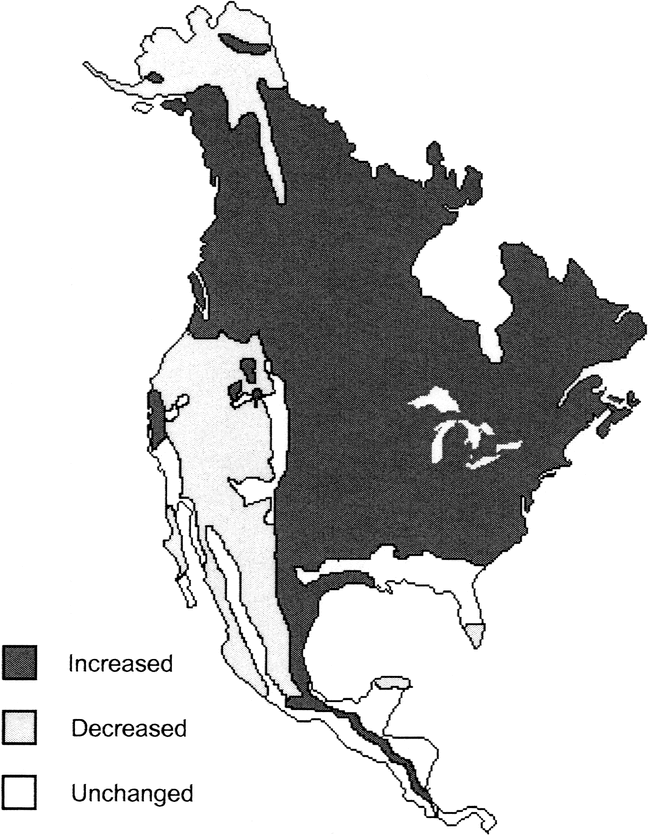
Reproduction of McDonald's map (1984) of North America depicting regions where productivity is thought to have increased, decreased, and remained unchanged during the terminal Pleistocene.
Seasonality and the DEM
Climate change at the end of the Pleistocene reorganized biotic communities such that there are no modern analogs because species responded individually to environmental change (FAUNMAP 1996; Graham and Lundelius 1984; Grayson 2007; Grimm and Jacobsen 2004; Lyman 2008; Martin and Martin 1987; S. D. Webb et al. 2004; Williams et al. 2001, 2004; Williams and Jackson 2007). At the end of the last glaciation and into the Holocene, seasonality in insolation—solar radiation received at the earth's surface—in the Northern Hemisphere increased, peaked, and decreased, acting as an external force on climate (Jackson and Williams 2004; Kutzbach and Ruddiman 1993; Kutzbach et al. 1998; Williams et al. 2001; T. Webb et al. 2004). The peak in seasonal insolation occurred roughly 12,000 14C years BP (approximately 14,000 calendar years BP). Williams et al. (2001:3357–3358) describe the peak of high seasonality during the latest Pleistocene as a “‘hypercontinental’ combination of hot summers, cold winters, and low precipitation.” Similarly, Prentice et al. (1991) report increasing summer temperatures after the glacial maximum in eastern North America. Webb et al. (1993:460) describe the terminal Pleistocene as a “‘transition’ period in which increased seasonality in insolation combined with the still significant Laurentide ice sheet to create combinations of climate variables that do not exist today… seasonal contrast was pronounced at this time.”
Consistent with our interpretations of the DEM's heterogeneity predictions, Prentice et al. (1991:2054, emphasis added) recognize that “climatic changes affect the natural disturbance regime and thereby alter the proportions of the landscape in different successional stages.” Increased intra-annual seasonality would have had a relatively severe impact on hypermorphic species because of their relatively long gestation periods, leading ultimately to their inability to recover from population reductions. Kiltie (1984:300–307) discusses three adaptive challenges that megaherbivores would have faced as seasonality changed during the late Pleistocene. First, if shifts in intensity of seasonal insolation affected season duration, then cues such as temperature, precipitation, and photoperiod, which initiate breeding seasons, would have been disrupted. This is especially problematic in situations where favorable reproductive periods became shorter than gestation periods. Megaherbivores had long gestation periods and were thus more vulnerable than relatively r-selected species to this kind of disruption. Second, gestation length is often depicted in terms of mean duration, but intraspecific variation in length of the reproductive period would exclude some individuals from reproducing—those with longer gestation periods. Kiltie (1984:301 emphasis added) states, “such [intraspecific] variation [in gestation length] is relevant to the question of adaptation to seasonal environments because even if a species uses mating cues that perfectly correlate with future conditions affecting maternal/offspring survival, there can still be an element of uncertainty in the timing of birth with respect to season.” As with the first adaptive challenge, a change in seasonality decouples periods of reproduction from periods of favorable intra-annual resource availability—that is, growing and breeding seasons would no longer have been aligned with reproduction and development in slow-growing, slow-reproducing animals. This relates to Kiltie's final challenge, that a change in seasonality disrupts the coordination between reproductive and environmental cycles. In effect, species lose reproductive time. “If individuals of a species time their mating activities so that young are reproduced only in certain seasons, some time may be lost that could have otherwise been spent in production of young. This lost time, which is not necessarily equal for all species in a given environment, can reduce the reproductive rate of the population” (Kiltie 1984:304).
There is increasingly abundant evidence that changes in seasonality were significant during the terminal Pleistocene (e.g., Broughton et al. 2008; Kutzbach and Ruddiman 1993; Kutzbach et al. 1998; Prentice et al. 1991; Webb et al. 1993; T. Webb et al. 2004; Williams et al. 2001). Variability in seasonal weather shifts would have altered the timing of optimal birthing and rearing; periods of the year during which optimal forage would have been available would have changed as winters became colder and summers became warmer (Anderson et al. 2007; Pielou 1991). Under the DEM, disturbance of birthing and rearing seasons would have caused frequent population reductions among slow-reproducing megaherbivores. Although not concerning a North American species, a recent study on Irish elk (Megaloceros giganteus) relates extreme hypermorphy in antler growth and highly cursorial anatomy directly to contingencies of reproduction in females (Worman and Kimbrell 2008). Those authors conclude that Irish elk could only have evolved where the seasonal resource pulse is highest (Geist 1998), and this species' demise most likely relates to a decrease in growing season length at the end of the Pleistocene.
In western North America, population reduction related to increasing seasonality appears to have occurred in multiple species of artiodactyls (Broughton et al. 2008). It is also significant that low r, K-strategists that survived the extinction period (e.g., extant elephants and rhinoceroses) inhabit areas of the world that maintain relatively equable seasonality during the Holocene (Kiltie 1984:309, figure 14.5). Johnson (2002) and others (Kelt and Meyer 2009) suggest that arboreal and nocturnal species survived extinction because they avoided human predation, but these species also occur in seasonally equable environments. Perhaps they avoided extinction because they were inhabitants of the subtropics and tropics (and thus were not subjected to increased seasonality) rather than because they avoided human super-predators (which has been assumed to be the reason for their survival in some cases).
The cause of many of the extinctions might not have been an increase in mortality, but a decrease in fertility, perhaps due to changes in seasonality. This is an important possibility because previous studies have searched mainly for cause in mortality. If a shift in seasonality occurred, Kiltie's (1984) potential mechanism for extinction offers a rarely acknowledged but striking alternative explanation to the overkill hypothesis.
Environmental Heterogeneity during the Terminal Pleistocene
Another hallmark of gradual increases in productivity and disturbance (e.g., seasonality) is a decrease in environmental heterogeneity. Movement upward and right on Transect E (Figure 2) is described by Huston (1994:149): “frequent disturbances will produce many patches that are denuded or have low density populations. However, recovery from disturbance will be rapid [because of high productivity]… Dominant life histories will be characterized by rapid growth, early reproductive maturity, small size, [and] low efficiency of resource use.” Heterogeneity in such habitats will be “exceeded only by low-growth, low-disturbance conditions” (to the left and down on Transect E). Indeed, patchiness and heterogeneity of habitats decreased from the late Pleistocene into the Holocene (FAUNMAP 1996; Graham 1986), echoing that both productivity (McDonald 1984) and disturbance (Prentice et al. 1991; Williams et al. 2001) increased during the same period (up and right on Transect E).
It has long been a truism in ecology that smaller populations have a higher probability of dying out than larger populations (MacArthur and Wilson 1967; Shaffer 1981). Further, the greater the degree to which a population is isolated from other populations of conspecifics (together comprising a metapopulation), the greater the chance that such a population will go extinct for lack of immigrant recruitment (Levins 1969). This would especially be the case for hypermorphic, K-strategists with dispersal phenotypes, which require large tracts of high-productivity habitat within low environmental disturbance regimes. The stochastic nature of terminal Pleistocene climates would have fragmented (meta)populations and habitats into discontinuous, isolated, and variously-sized patches. If a few small populations located between large populations were extirpated for any reason, then the remaining populations would have been relatively more isolated and less likely to recruit through immigration even if the surviving populations increased. It is easy to envision several climatic fluctuations over a few thousand years—for example, between about 15,000 and 10,000 B.P.—eventually extirpating isolated, stochastically depleted populations of K-strategists, and ultimately resulting in their extinction. That is indeed what seems to have occurred with alpine and subalpine small mammals (r-strategists) on Great Basin mountain-tops during the Holocene (Beever et al. 2003; Grayson 2006) and with some populations of the genus Oreamnos (Lyman 1998; Nagorsen and Keddie 2000).
Conclusion
Arguments that focus on the large size of extinct megafauna and consider them to be slow-breeders (e.g., Johnson 2002; Lyons et al. 2004) turn too quickly to overkill as the cause of extinction. For example, Brook and Bowman (2005) evaluate many of the same life history parameters that we do but conclude that humans are the only mortality agent that could have exerted sufficiently regular population reductions to cause the extinctions. As discussed above, an increase in seasonality might have caused a decline in fertility (sensu Kiltie 1984), and evidence for this kind of climate change exists. Further, studies that implicate size-selectivity are quick to cite K-strategist life history traits as indicative of vulnerability to extinction through population reductions, but tend to neglect other life history traits in hypermorphic mammals that are relevant to the debate (e.g., development of anti-predator phenotypes).
Grayson (2001, 2007) suggests that the evidence for human hunting and timing of extinction should be considered independently for each taxon; this strategy has produced interesting results at several spatial scales (Stuart et al. 2004; Surovell et al. 2005; Worman and Kimbrell 2008; see also Guthrie 2003). However, there is a recent tendency to choose the (safer) “middle ground,” citing correlations between human arrival and extinctions (e.g., Steadman 2006; see discussion in Koch and Barnosky 2006; Grayson and Meltzer 2003). These “middle ground” arguments also focus on the weakness in climate models – that the terminal Pleistocene does not appear to have been unique in terms of environmental disturbance. The middle ground is unsatisfactory because temporal correlations are only weakly documented with empirical evidence. Thus, each new fossil specimen that is well dated, has a strong association with human artifacts, or both, is significant.
Pleistocene climate change in North America meets several predictions of the DEM regarding extinction. There is empirical evidence that environmental heterogeneity decreased, habitat productivity may have increased, and disturbance increased. In general, the ecological product of the late Pleistocene extinctions was an overall reduction in species diversity, which represents movement upward and right along Transect E in the DEM (Figure 9).
The dynamic equilibrium model as applied in this study to the late Pleistocene. Transect E corresponds to Figure 2, and the y-axis highlights increasing seasonality as an important environmental disturbance in the extinctions.
The DEM offers a previously unconsidered theoretical perspective with which to evaluate evidence surrounding the hotly debated causes of late Pleistocene extinctions because it highlights the evolutionary life history relationships underlying community species richness. An important contribution of the model is that it helps sort out the role that an increase in productivity might have played in structuring biological communities prior to and during the extinction period. The model also highlights the importance of frequent population reductions resulting in extinction. Although frequent human predation could have been a mechanism of population reduction, archaeological evidence is necessary to support such a conclusion. Increased seasonality after the last glaciation, for which there is evidence, would have been a persistent, frequent disturbance that was ubiquitous at temperate latitudes. There is minimal evidence for frequent predation of multiple species of megaherbivores by humans. Elevated productivity and increased seasonality appear to have worked in tandem to contribute significantly to, if not simply cause, the late Pleistocene extinctions.
Footnotes
Acknowledgments
We thank Valerius Geist, Michael Huston, Ernie Lundelius, David Meltzer, and Lisa Nagaoka for comments on early drafts and/or discussion of ideas expressed in the paper. Kevin Cagle provided the Fort Hood deer data. David Steadman provided comments that challenged us to better communicate our thoughts on models, data, and opinions. Two anonymous reviewers provided comments that improved the paper as did the copy editing by Kelly Ferguson, Heather Trigg, and Virginia Popper. Eva Ruiz translated the abstract to Spanish, and Brannon Barr translated it to French. This research was supported by in-kind support from the Institute of Applied Sciences and the Department of Geography at the University of North Texas.