
Abstract
The present paper examines the different types of plant and animal names used in Kakataibo (Pano, Peru) in terms of the typology of ethnobiological nomenclature proposed by Berlin et al. (1973; slightly modified in Berlin 1992). While doing so, this paper highlights and discusses the major issues posited by the Kakataibo ethnobiological lexicon, which mainly arise due to the pervasive presence of specific-folk generic polysemy in the language. More precisely, this paper shows that the pervasive presence of this type of polysemy creates a type of contrast set, whose members combine the taxonomic properties of secondary names and complex primary names of the productive type. This is a widespread property of the Kakataibo ethnobiological taxonomic system, which invites us to rethink the distinction between these two types of ethnobiological nomenclature. A comparison of the Kakataibo data with data from other Pano languages is also offered in this paper.
Introduction
Linguistic ethnobiology deals primarily with the naming of animals and plants by different ethnic groups. The scientific relevance of ethnobiological systems for naming animals and plants used by traditional societies was first noted by Lévi-Strauss (1966:153–154), who remarked that people from these societies are able to remember an impressively large number of lexemes that designate the flora and fauna in their surroundings. While Western scientific taxonomy has standardized rules for labeling scientific species and higher categories, indigenous peoples utilize multiple different strategies for naming their local flora and fauna, which are of linguistic and cognitive interest. This paper contributes to the understanding of folk biological nomenclature by describing the Kakataibo ethnobiological lexicon, showing how it fits in with the existing typologies of ethnobiological nomenclature (specifically, Berlin et al. 1973; slightly modified in Berlin 1992), and presenting its most salient properties and particularities. All the claims offered in this paper are based upon an ethnobiological lexical database with 1,233 lexical entries, that includes preliminary biological identifications and detailed descriptions given by the Kakataibo people for approximately 70% of the listed species (see Zariquiey et al. 2014).
According to Berlin (1992:20), “In any new summary of the patterned ways in which people think and talk about plants and animals, it is important that a clear distinction be made between the psychological conceptualization of plants and animals and the linguistic reflections of this underlying conceptual structure.” The present paper deals with the linguistic reflections (nomenclature) of the psychological categories with which the Kakataibo people conceptualize plants and animals; covert categories that are psychologically relevant, are not discussed.
The Kakataibo People
The Kakataibo people (also known as “Kashibo-Kakataibo,” “Kashibo,” and “Uni,” among other denominations) belong to the Pano language family and live in the Peruvian regions of Huánuco and Ucayali, along the Aguaytía, Shamboyacu, San Alejandro, Sungaroyacu, and (more recently) Pisqui Rivers (Figure 1). According to the most recent Census of Indigenous Communities of the Peruvian Amazon (INEI 2007), the Kakataibo people currently number about 1,879. However, the Kakataibo's political organization (FENACOCA) considered their number to be closer to 3,000 or 3,500 in 2007.
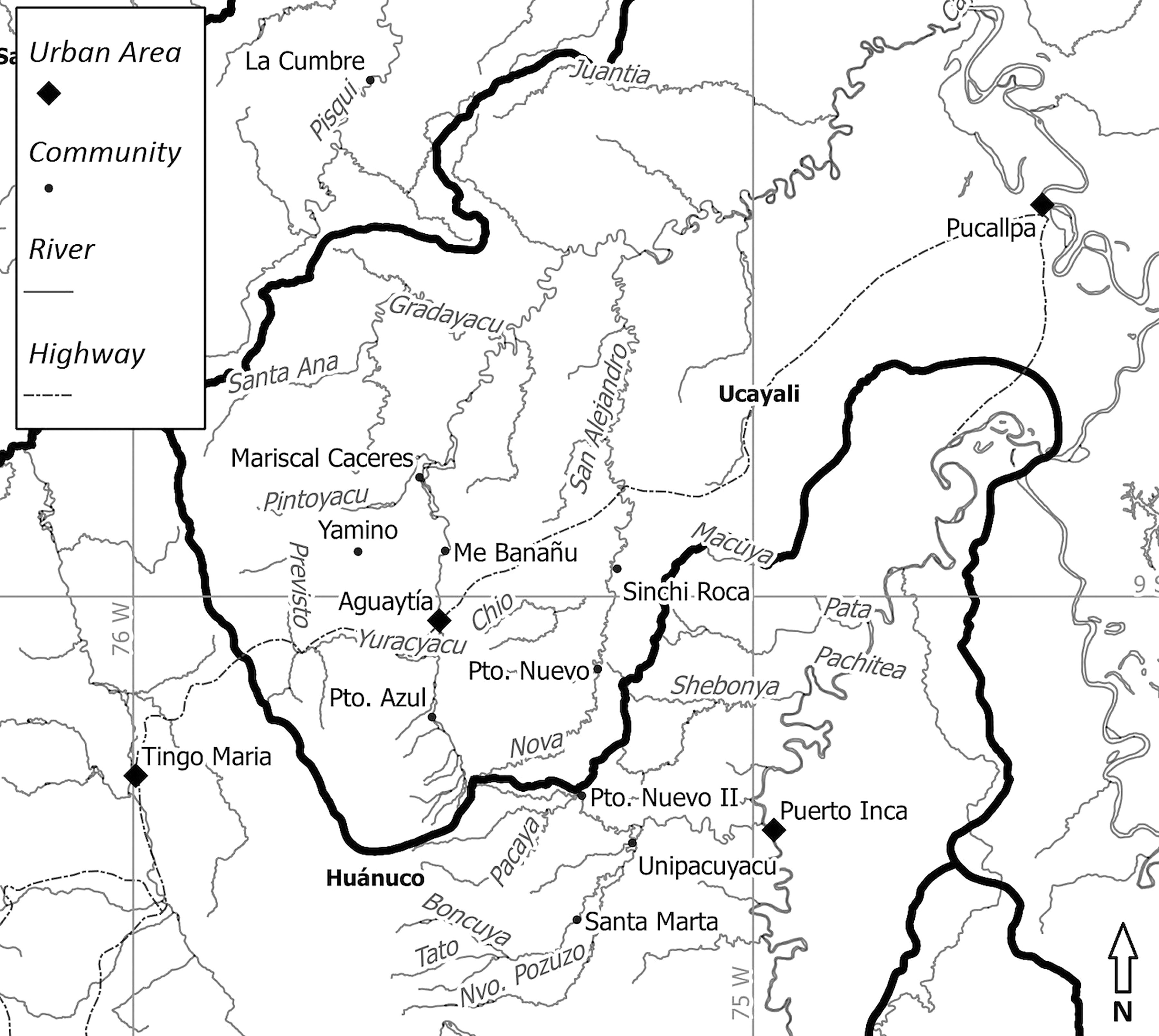
Location of Kakataibo communities.
Kakataibo is the westernmost Pano language and, therefore, the one closest to the Andes Mountains. Shell (1965), D'Ans (1973), Loos (1999), and Fleck (2013) correspond in their treatment of this language as the only member of its branch. As described in Zariquiey (2011b), the Kakataibo language has (at least) four extant dialects and the data offered in this paper come from the Lower Aguaytía River dialect (see Zariquiey 2011a).
Folk Taxonomy
According to Berlin, the number of ranks in folk taxonomies does not exceed five or six. These include: unique beginner, life form, intermediate, generic, specific, and varietal. Berlin et al. (1973) graphically modeled the relationship of five of these proposed universal ethnobiological ranks (intermediates are not included), and in his subsequent monograph, Berlin (1992:21) refined and further elaborated on the nine principles of ethnobiological systems (as proposed in Berlin et al. 1973), culminating in his 12 general principles “that characterize a wide number of ethnobiological systems of classification from diverse parts of the world.” Five of these principles have to do with nomenclature and will be commented on throughout this paper in view of the Kakataibo data. Figure 2 (reproduced from Berlin et al. 1973) is a schematic representation of five of the six ethnobiological ranks in an idealized model.
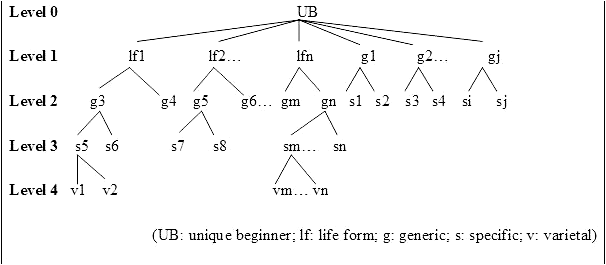
Schematic representation of five of the six ethnobiological ranks in an idealized model (reproduced from Berlin et al. 1973).
Typologies of Ethnobiological Nomenclature
Conklin (1962) proposed the first typology of ethnobiological nomenclature, which is summarized in Table 1. Conklin's typology distinguishes two main types of biological “lexemes” (or more properly,
Conklin's (1962) typology of ethnobiological nomenclature.

Berlin et al. (1973) elaborated on Conklin's (1962) proposal, introducing an additional distinction to produce the partially divergent typology represented in Table 2, where I have pointed out matches and mismatches with Conklin's proposal.
Berlin et al.'s (1973) typology of ethnobiological nomenclature.

This typology was retained by Berlin in his 1992 monograph, but the author changed his terminology as follows: primary lexemes =
According to Berlin's second principle of nomenclature, “
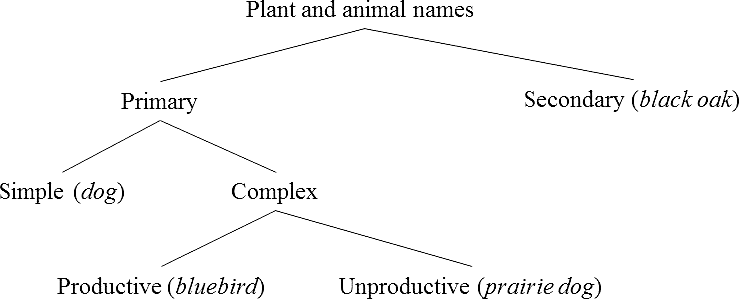
Typology of plant and animal names (Berlin et al. 1973; Hunn and Brown 2011).
In sum, the main difference between Conklin's (1962) typology and Berlin et al.'s (1973) proposal has to do with the introduction of one additional distinction within the category of primary complex labels, which are divided into productive and unproductive by Berlin and his associates. It is important to note that the distinction between productive complex names and secondary names is only based on taxonomic criteria (i.e., their contrast sets) and not on linguistic properties—in the sense that in both cases we have complex forms with one constituent that refers to a superordinate (see Berlin et al. 1973:217, note 4). I will return to this issue below.
Methods
The lexical database used in this paper is the result of a one-year collaborative documentation project, in which Kakataibo speakers were incorporated as members of our project team and not just as experimental subjects. Thus, the project has promoted the development of a local team, composed of eight members of the Yamino community (five men and three women), who have participated in different ways in the activities included in our project—group walks into the forest, preparation of a multi-authored ethnobiological dictionary (see Zariquiey et al. 2014), biological identification of species, audio and video recordings of cultural knowledge, mythology about plants and animals, and so on. The participation of the Kakataibo people has carefully satisfied all the requirements established by the Ethics Committee of the Pontificia Universidad Católica del Perú, including permission to work in the Yamino village and formal consents from team members. The project has also complied with all the obligations agreed to with the population of Yamino, which included fair salaries for the participants, support for the development and promotion of their local handicraft, and sharing of the results with the community.
Species Assignments
Our documentation project followed methods suggested by Fleck (2007) for obtaining scientific designations for plant and animal names in a research language when it is not possible to collect voucher specimens. Initially, lists of animal and plant names were compiled from research sessions with the Kakataibo members of our local team and augmented with plant and animal names that appeared in recorded texts and/or Wistrand de Robinson (1984), or overheard during residence in Yamino. Subsequently, these names were correlated with biological species (or higher-ranked biological taxa) and new names were elicited using drawings or photographs in field identification guides (Emmons [1997] for mammals; Schulenberg et al. [2007] and Clements and Shany [2001] for birds; Bartlett and Bartlett [2003] for reptiles and amphibians; Henderson et al. [1995] for palms; and Goulding et al. [2003] for fish, among others), while simultaneously discussing the natural history of the species and playing recorded vocalizations, if available (Emmons et al. [1997] for mammals; Schulenberg et al. [2000] for birds; and Cocroft et al. [2001] for frogs). David W. Fleck, who has conducted zoological and botanical inventory work and ethnobiological research with the Matses (also Pano) in Peruvian Amazonia, worked together with me and part of the Kakataibo team in Lima to associate the names with scientific designations. Only species expected to be in the area (according to the range map or range description in these field identification guides) were used in this type of group research.
These methods were useful for collecting a near complete inventory of Kakataibo plant and animal names, both in the field and in research sessions conducted in Lima, with the participation of four speakers of the language. As no voucher specimens were collected to confirm the identifications, the scientific designations are necessarily tentative; however, for the larger and distinctive species, as well as for species in monotypic taxa, there is little room for doubt. In any case, since the present paper focuses on the form of the names rather than on a comparison with scientific systematics, it is usually not relevant whether some of the names for difficult-to-identify plants and animals actually refer to a sister species, or to more than one species that the Kakataibo people consider a single terminal taxon.
Higher-Order Groupings
Higher-order groupings are based on a total of 24 hours (divided into eight sessions of three hours each) of group discussion about the relationship among different animals and plants and their organization. These sessions of group discussion were led by the author and always included the participation of at least four people (mostly members of our local Kakataibo team, but also itinerant participants who occasionally volunteered to participate in a session). Men and women usually worked separately due to cultural preferences.
The methodology of these group sessions consisted of giving the participants a number of topics to discuss as a group (e.g., fish, parrots, monkeys, palms, and so on). They were asked to construct an agreed-upon organization of the category under discussion based on the names proposed by them and the ones that our project had previously documented. Notes on A2-sized pieces of papers, using markers of different colors, were taken mostly using tree figures to represent the conclusions reached by the group. On average, each topic took one hour to discuss so we were able to discuss three topics per session. From the eight sessions, four were recorded on audio and two were also partially recorded on video. This methodology allowed us to offer a first approximation of the internal structure of a considerable number of higher-order taxa, from unique beginners to generics.
Conventions Used in this Paper
All the examples offered in this paper observe the following orthographic conventions (phonetic symbols are given when they do not coincide with their corresponding orthographic representation): a, e, ë [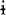 ], i, o, u, p, t, k [k], kw [kw], b [β, w], r [
], i, o, u, p, t, k [k], kw [kw], b [β, w], r [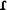 ], m, n, ñ [
], m, n, ñ [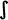 ], s [s, z], sh [
], s [s, z], sh [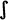 ], x [
], x [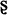 ], ts [ts], ch [t
], ts [ts], ch [t ], and [
], and [ ]. In addition, for names that have been identified only to the genus level, the following abbreviations are used: sp. = names that refer to only one biological species (of a genus), but it has not yet been identified; spp. = names that refer to more than one species (of a genus); and sp(p). = names known to refer to at least one species (of a genus), but it is not certain whether they refer to more than one species of the genus.
]. In addition, for names that have been identified only to the genus level, the following abbreviations are used: sp. = names that refer to only one biological species (of a genus), but it has not yet been identified; spp. = names that refer to more than one species (of a genus); and sp(p). = names known to refer to at least one species (of a genus), but it is not certain whether they refer to more than one species of the genus.
Ethnobiological Nomenclature in Kakataibo
In this section, I illustrate and describe the most salient types of Kakataibo's plant and animal names following Berlin and colleagues’ typology (but using the terminology employed in Berlin 1992). Some particularities of the Kakataibo data in relation to this typology are also discussed here.
Primary Labels
Simple Primary Labels
Simple primary labels are found at every level of the taxonomic classification of plants and animals used by the Kakataibo with no distributional restrictions, but they tend to appear in generic or higher taxonomic categories. The simple primary label nëpaxta is a generic name used to refer to all the sungrebes identified by the Kakataibo people (for instance, both Nomonyx dominicus [L.] and Heliornis fulica [Boddaert]), while the simple primary label ñutan refers to a specific type of tinamou (Crypturellus cinereus [Gmelin]). The term ñutan contrasts with other terms, including the simple name kuma, which refers to the black tinamou (Tinamus osgoodi Conover). However, the name kuma is also used as a generic, meaning simply ‘tinamou’. In this use, the semantic range of kuma includes both kuma in its specific sense and ñutan, among other terms used to name the different species of tinamous identified by the Kakataibo people. This situation, in which a polysemous term can refer to both a subgeneric taxon (which can be considered the prototype) and to one of its superordinate taxa, has been called
According to my current interpretation of the Kakataibo taxonomic system, ñutan (Crypturellus cinereus) belongs to the
Therefore, simple primary names appear in association with both generic and specific taxa. However, this is also true regarding other levels of the taxonomic hierarchy illustrated in Figure 2. For instance, the collective research conducted with the Kakataibo speakers led to an interpretation according to which the simple term ‘isá denoting ‘(inedible) small birds and passerines’ corresponds to the rank that Berlin calls
The total number of simple primary names in our database is 599, which corresponds to 48.6% of the total number of entries (1,233). Additional exemplification of the simple primary names found in our database is offered in Table 3.
Examples of simple primary labels in Kakataibo.

Unproductive Complex Primary Names
These forms are also found in Kakataibo. They appear in association with both generic and specific taxa, but they mainly relate to the latter. This is again an unusual property of the Kakataibo ethnobiological taxonomic system since unproductive primary terms would be expected to be generic binomials (see Hunn and Brown 2011). Table 4 presents some examples of this kind of name.
Examples of unproductive labels in Kakataibo.

All the examples in Table 4 refer to Kakataibo specific taxa and are considered “unproductive” because, for example, ‘isku xëta literally means ‘oropendola beak’ and does not include as its head a word meaning ‘
The total number of unproductive complex primary names in our database is 154, which corresponds to 12.5% of the total number of names. These 154 names include nominalizations like xëta ‘amiananti ‘rufous-tailed flatbill’ (literally, one that can harm with its beak) or taë tëbiskati ‘non-identified fish species’ (literally, one that can cut one's feet). Notice, however, that similar nominalizations may also carry an adjacent noun referring to a superordinate, as in the case of an nami pikë
Secondary Labels or Productive Complex Primary Labels?
One difficulty that surfaced while analyzing the Kakataibo terminology for naming plants and animals in terms of Berlin and colleagues’ typology was that, in many cases, determining whether a particular binomial or polynomial constitutes a secondary label or a productive complex primary label was not straightforward. This was due to the particularities of the contrast set within which it was included. Before discussing these difficult cases, let us start with others for which this distinction is straightforward. Figures 4 and 5 list the types of giant hunting ants and Spanish cedar identified and named by the Kakataibo people, respectively.
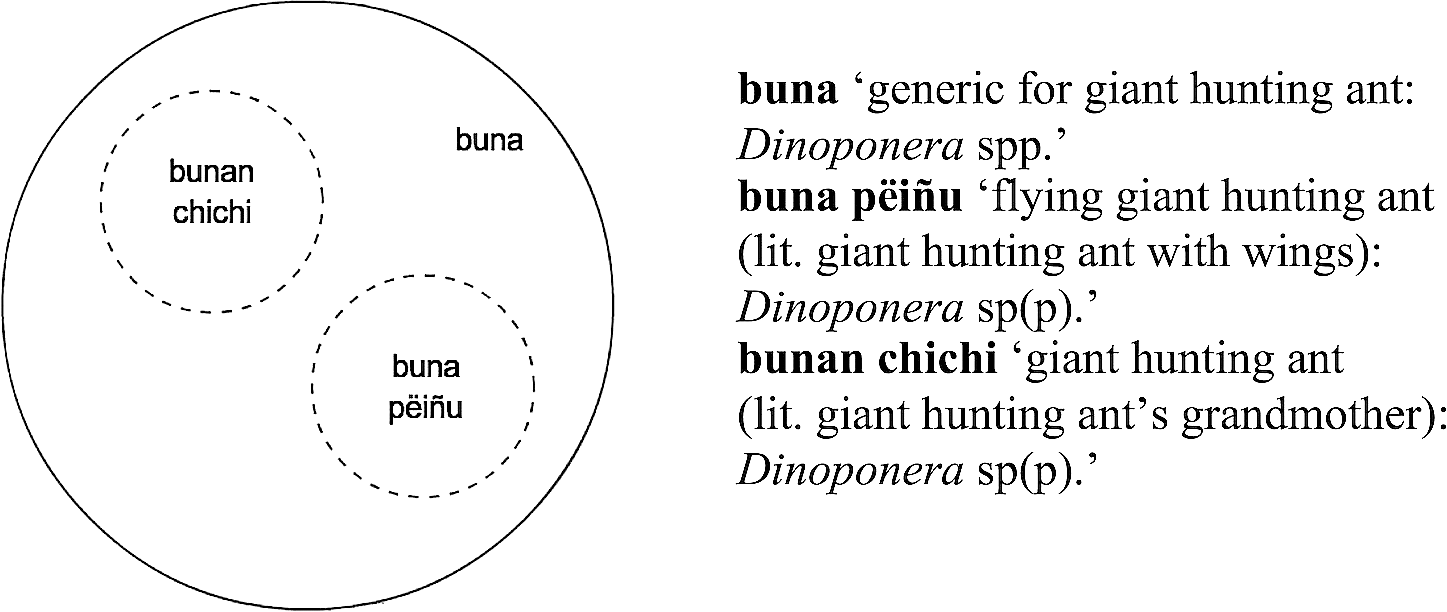
Types of giant hunting ants identified by Kakataibo people.
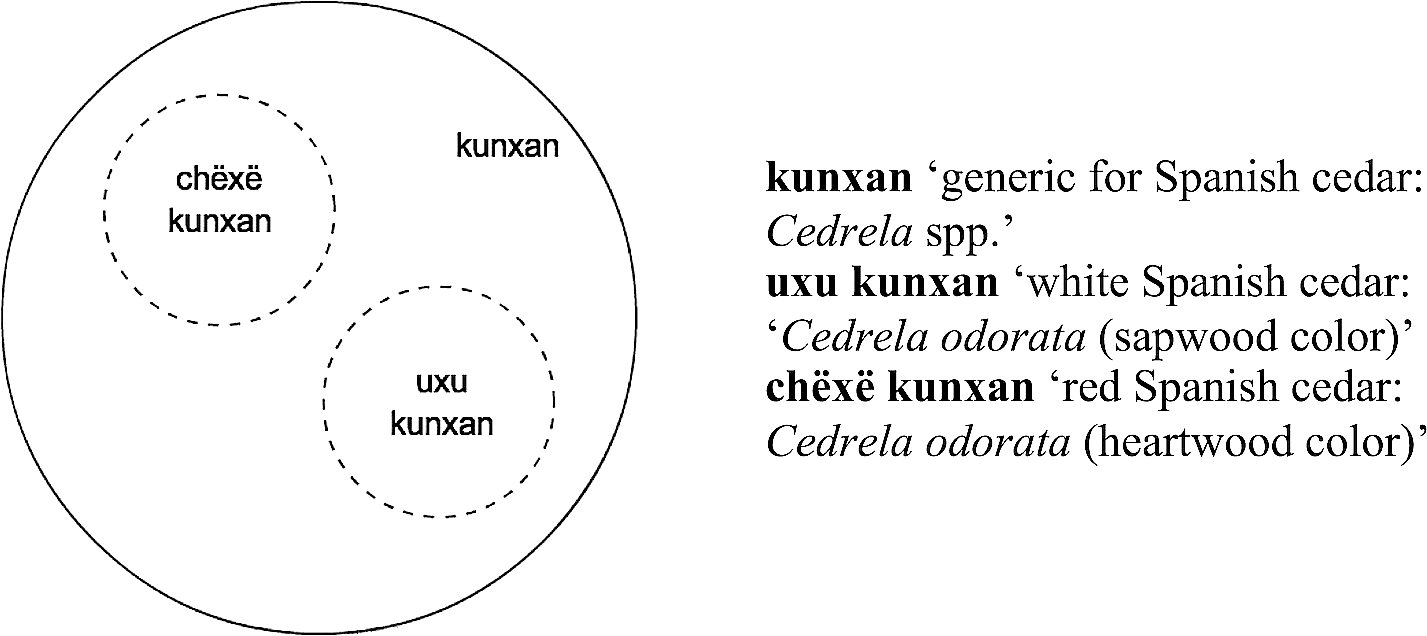
Types of Spanish cedar identified by Kakataibo people.
In Figure 4, we find one generic primary name for ‘giant hunting ant’ and two binomials that refer to two types of these ants and contrast with each other. These binomials equally include the name of the generic (buna) and constitute a contrast pair exclusively conformed by morpho-syntactically complex terms. Exactly the same situation is found in Figure 5. Therefore, the binomials in Figures 4 and 5 clearly constitute
In turn, Figure 6 presents a selection of the types of armored catfish (Fam. Loricariidae) that the Kakataibo people distinguish and name (the complete list includes at least 19 different terms, whose referents are still to be more precisely identified).
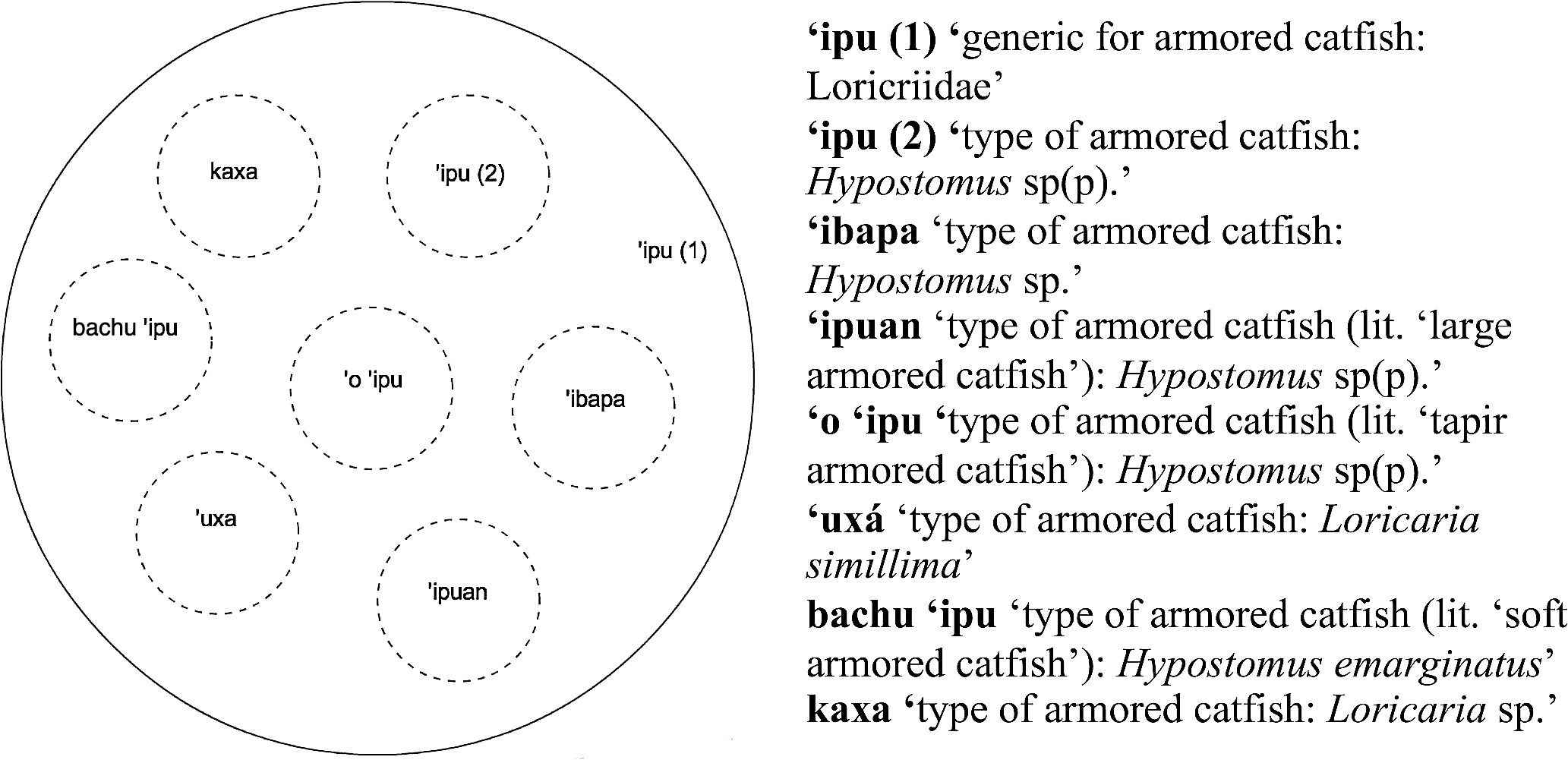
Types of armored catfish identified by Kakataibo people.
Although they contrast with each other and are formally identical to the secondary terms identified in Figures 4 and 5, the names ‘o ‘ipu and bachu ‘ipu fit Berlin et al.'s definition of
Notice that, as is usually the case in Kakataibo's taxonomic system, in Figure 6, the name ‘ipu is a generic name that refers to the whole class, but also the name of one specific type of armored catfish identified by the Kakataibo people. This type of polysemy may be problematic for the distinction between secondary names and productive complex primary names in cases like the one presented in Figure 7, where the different subtypes of the single local biological species of tapir that the Kakataibo people identify are listed.
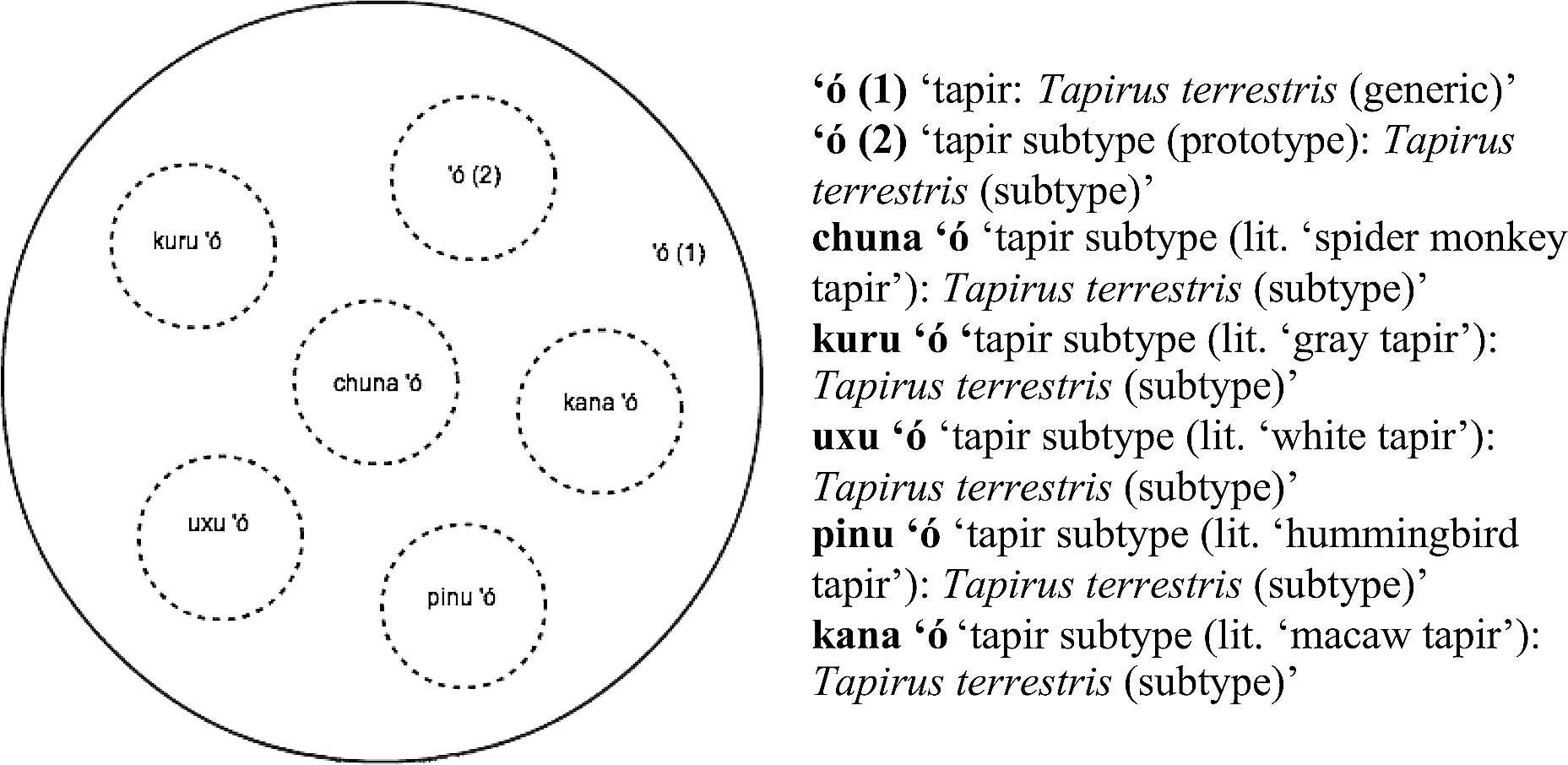
Types of tapir (Fam. Tapiridae) identified by Kakataibo people.
As can be seen in Figure 7, the Kakataibo people recognize and name (at least) six subtypes of the single local species of tapir, with five of the subtypes being labeled with descriptive binomials, and one with a one-word term. The number of different types of tapir identified and named by the Kakataibo people is remarkable, since Berlin (1992) suggests that domesticated species are usually over-differentiated, while wild species are not. Similar cases have been widely described in the literature. For instance, Hunn (1977) reported over-differentiation of peccaries by the Tzeltal Indians of Chiapas. In strict terms, binomials like the ones in Figure 7 would constitute productive primary complex names, since they do contrast with one primary name (‘ó in its specific sense). However, contrast sets like the one in Figure 7 are in some way different from the ones in Figure 6. In the latter, we find
Distribution of different types of names (following Berlin's [1992] terminology) in our Kakataibo database.

Summary
Table 5 summarizes the general characteristics of the Kakataibo ethnobiological database this paper is based on. It can be appreciated that the total number of complex names (binomials and polynomials) is 634 and, therefore, they are more numerous than simple terms (N = 599). In addition, among complex terms, productive primary ones are the more frequent (with 211 tokens), while we only find 47 names that clearly correspond to the category of secondary names. Furthermore, 202 complex names appeared in contrast sets that exhibit features associated with both secondary names and productive primary ones (similar to the one illustrated in Figure 7) and, therefore, have been called as “mixed” in the context of this paper.
Discussion
As may be concluded from the above analysis and from the results presented in Table 5, the pervasive presence of specific-folk generic polysemy has produced a type of contrast set that was not predicted by Berlin et al. (1973) or Berlin (1992) in their distinctions between productive complex primary names and secondary names. This kind of polysemy is, of course, not new in the literature and has been described for many other languages. In fact, in Berlin et al. (1973), it is possible to find several examples of this kind of polysemy in languages such as Hanunóo, Karam, and Guarani, among others. In turn, Taylor (1990:25–28) offers a detailed discussion of this issue in the ethnobiological taxonomic system of the Tobelo people (Moluccas, Indonesia), in which, due to polysemy, a single name may appear in different contrast sets. Taylor illustrates this by examining the most inclusive named Tobelo groupings of floral forms: o gota ‘tree’, o gumini ‘vine’, and o rurtibu ‘herbaceous weed’. The issue regarding these words is that they involve polysemous meanings that produce variations in membership of lower-level terms into these three major categories. For instance, among other meanings, o gota could be used to refer to ‘firewood’ and in this sense it includes any type of firewood, even if it comes from a tree-like palm or cycad, which do not belong to the o gota class and, therefore, cannot be referred to by this name if not used as firewood. Although polysemy is pervasive in Kakataibo, as far as I can tell, it does not produce a similar multi-membership effect.
Another important issue raised by Taylor has to do with the need of distinguishing between polysemy and homonymy. Although the distinction between polysemy and homonymy is not always clear, for Taylor (1990:28) the criterion of distinction should be relatedness of meaning (as is traditionally proposed by dictionary makers): “two lexemes may be said to be homonymous if all their forms [i.e., sounds] are the same, but they have unrelated meanings … Polysemy, on the other hand, refers to related meanings of the same lexeme” (Taylor 1990:28). This distinction is certainly useful for Kakataibo. Although all the examples presented in this paper are clearly cases of polysemy in Taylor's terms, homonymy does exist in the inventory of animal and plant names of the language. One example of this is found in the pair ‘aku ‘type of catfish’ (Cetopsis sp.) and ‘aku ‘unidenfied tree’. These two words have exactly the same sounds but unrelated meanings. One interesting type of meaning relatedness among the senses of a single lexeme is metonymy. Words are metonymously related when “we use [one] word not in its established sense but to name a category in contextual association with the category usually named by the word” (Waldron 1967, quoted in Taylor 1990:28). Examples of metonymy are also found in Kakataibo, as can be seen in the pair xëox ‘tangarana tree’ (Triplaris sp.) and xëox ‘type of ant that lives in the tangarana tree’ (Pseudomyrmex gracilis Fabricius). Thus, although there is evidence of honomymy and metonymy in the Kakataibo language, this paper is exclusively based on cases in which a lexeme presents two related meanings that exhibit a taxonomic relation (i.e., a species and its superordinate).
This type of polysemy corresponds to what Forth (1995) calls
As we can see, polysemy is not a clear-cut and transparent phenomenon. On the contrary, it must be distinguished from homonymy and, at least partially, from metonymy, as it has been done by Taylor (1990). In addition, as proposed by Forth (1995), it is also relevant to distinguish between taxonomic polysemy (inclusion) and non-taxonomic polysemy (encompassment). All these distinctions can be established in Kakataibo, but since only inclusion has direct taxonomic consequences (Forth 1995), this paper is exclusively based on this type of polysemy (called here
As shown in the previous section, due to the pervasive presence of specific-folk generic polysemy, a considerable number of binomials (which may largely contrast with other binomials) also contrast with the non-generic use of the simple name that refers to their superordinate. This produces a situation in which the distinction between secondary names and primary names of the productive type (in Berlin's [1992] terms) is not always easy to apply. This issue is explored in greater detail below.
According to Berlin (1992:28, nomenclature principle 2), “secondary forms differ from primary productive expressions in that the former occur, with predictable exceptions, only in contrast sets whose members share a constituent that labels the taxon that immediately includes them.” In cases like the one illustrated in Figure 7, the non-generic use of the polysemous term shares a constituent with the binomials it contrasts with and, crucially, this constituent refers to their superordinate (e.g., compare
However, in Kakataibo it is possible to find binomials or polynomials that neither contrast exclusively with other complex names with which they share a constituent referring to their superordinate, nor contrast with lexically different primary terms. Therefore, they are neither prototypical secondary terms nor prototypical complex primary names of the productive type. It is true that the typology proposed by Berlin and his associates predicts exceptions to these prototypes (see Berlin 1992:28, nomenclature principle 2), but the issue in Kakataibo is that contrast sets like the ones in Figure 7 are not exceptional but very common, due to the pervasive presence of specific-folk generic polysemy. Although this type of polysemy is not new in the literature, what seems to be unusual regarding Kakataibo taxonomy is that this kind of polysemy is quite common for polytypic categories and constitutes a well-established systemic pattern. Thus, any analysis that treats cases like the one in Figure 7 as exceptional would be problematic for this language.
Conklin (1962) developed his typology of ethnobiological nomenclature based on two linguistic features: 1) the number of roots that make up a term; and 2) whether a multi-root term carries the semantic property of descriptive force. That is, whether a term involves a head noun referring to a superordinate category, which is modified by a second root. Berlin et al. (1973) and, more recently, Berlin (1992) based their typology on these same linguistic characteristics, but added a taxonomic criterion, specifically related to the properties of the contrast set in which we find a multi-root term with a head that refers to its superordinate in order to introduce the distinction between productive complex primary names and secondary names. The type of contrast set exhibited by a multi-root term is more a taxonomic property than a properly linguistic one. However, it was argued by Berlin et al. (1973) that this decision was based on the need of distinguishing between multi-root terms that would have equally been
A property-driven approach to the classification of plant and animal names (similar to the one that is being argued for in contemporary linguistic typology; e.g., Plank 2001, Hyman 2009) is useful to offer a satisfying analysis of the Kakataibo data presented in this paper, without introducing a new distinction into the typology. Basically, there are two properties which are relevant for the distinction between prototypical secondary names and prototypical productive complex primary names in Berlin's terms: 1) to
A property-based approach to the distinction between secondary names and productive complex primary names.

As we can see, the property-driven approach proposed here produces a three-way distinction which allows us to offer a satisfying description of the Kakataibo data, without abandoning the typology proposed by Berlin, which has proven to be useful for other languages. Basically, in this approach, we have three possible combinations of the two features that define the two prototypes described by the categories
The two Pano languages whose ethnobiological taxonomic systems have been studied in more detail are Shipibo-Konibo (e.g., Tournon 1991, 1994; Valenzuela 1998, 2000) and Matses (e.g., Fleck and Voss 2006; Fleck et al. 2002). Valenzuela (1998) has shown that in Shipibo-Konibo it is very usual to find that names of plants and animals are compounds. Indeed, Valenzuela (1998) mentions that more than 86% of the compound names found in the whole language are associated with names of animals and plants. For instance, as indicated in Valenzuela (1998), the Shipibo-Konibo people distinguish around 30 types of plantain and all the names used to refer to them are complex. In many of the complex terms analyzed by Valenzuela, we find a nominal head referring to a superordinate accompanied by a pre-head modifier, which expresses a property (shape, color, behavior, etc.). However, complex names might also include heads that are not related to a superordinate, but to some other referent, which may or may not be semantically related with the named species by means of a metaphoric or metonymic relation. In general, the scenario in Shipibo-Konibo is similar to what we find in Kakataibo regarding the extended presence and use of binomials/polinomials in their ethnobiological taxonomic systems (recall that in our lexical database we have more complex names than simple one). However, we do find differences between the two languages regarding polysemy. While polysemy is a pervasive feature of the Kakataibo taxonomic system, Valenzuela (2000) only reports a few cases of polysemy, like the word banabu, which can mean ‘cultivated plants’, but also ‘plant’. Tournon (1994) gives a similar example with the word yuina, usually translated as ‘edible or game animal’ (Valenzuela 2000), which can also refer to its superordinate ‘vertebrates’. What we find in Kakataibo is clearly different from Shipibo-Konibo both because polysemy is mostly found between generic and specific taxa and not at higher ranks, and because polysemy constitutes one of the most significant and systematic properties of the whole taxonomic system.
The Matses case is also very interesting. Fleck and Voss (2006) offer a detailed description of the Matses classification and terminology for mammals. One of the most fascinating properties of the terminology presented by the authors is the presence of unusually large synonym sets to refer to a single mammal species or variety identified by the Matses (and other small Pano groups from the area). Interestingly, based on the information given, binomials seem to be clearly less common than simple terms in the terminology used by the Matses to name mammals. In fact, only a few complex names are found in the paper. These names usually include the name of the superordinate followed by an adjective like çhëxë ‘dark’ or uxu ‘white’. Simple names are clearly the norm in the Matses’ mammal terminology and this makes the situation in this language different from what we have found in Kakataibo and Shipibo-Konibo. It is important to notice that Fleck et al. (2002) find large sets of descriptive phrases formally similar to the binomials and polinomials in Matses’ bat terminology described by Fleck and Voss (2006) and by Valenzuela (1998). Yet, these descriptive phrases are not lexicalized terms and might be cognitively different from the complex terms discussed in this paper. Regarding polysemy, the situation is similar to what we find in Shipibo-Konibo. There is also no pervasive specific-folk generic polysemy indicated in Matses and, in fact, the single case of this type of polysemy given by Fleck and Voss (2006) is only partial—the term bëchun could mean ‘capuchin monkey’ but it could also mean ‘brown capuchin monkey’ (Cebus apella, syn. Sapajus apella L.), in which case it is usually followed by the modifier çhëxë ‘dark’. Therefore, based on the available references, it is possible to conclude that the type of polysemy described here is unusual from the perspective of the Pano language family and represents a specific trait of Kakataibo.
Conclusions
The present paper has listed and illustrated the different types of plant and animal names that are used in Kakataibo. The discussion has mainly followed the nomenclature typology proposed by Berlin et al. (1973; terminologically modified in Berlin 1992) and attention has been given to how the Kakataibo data interact with the taxonomic generalizations (particularly with respect to nomenclature) summarized in Berlin (1992). The primary focus of this study has been on issues concerned with the morpho-syntactically complex names and their classification in the Berlin et al. (1973) typology. While many of them can be easily classifiable (see Figures 4, 5, and 6), the pervasive presence of polysemous terms in the Kakataibo ethnobiological taxonomic system has produced a very particular situation, in which classifying morpho-syntactically complex names in terms of Berlin et al.'s (1973) typology is not always straightforward. Complex forms in contrast sets like the one in Figure 7 might simply be considered productive complex simple names, but they would not be prototypical examples of this nomenclature type since they do not contrast with lexically different primary names. These morpho-syntactically complex terms are also reminiscent of secondary names in the sense that they only contrast with other terms with which they share a constituent (that refers to their superordinate). However, prototypical secondary names only contrast with other secondary names and examples like the ones in Figure 7 contrast as well with one primary name—a polysemous form that also names the superordinate. In fact, contrast sets like the one found in Figure 7, which are very common in Kakataibo, exhibit characteristics that were not predicted by Berlin et al.'s (1973) typology. This problematic situation has been solved by means of a property-driven approach, which produces a three- (and not two-) way distinction, more appropriate for the organization of Kakataibo's plant and animal names (see Table 6).
This proposal constitutes a new approach to the nomenclature typology proposed by Berlin and his associates. This new approach facilitates a description of the complex situation found in the Kakataibo data without abandoning the distinctions proposed by Berlin et al. (1973). Interestingly, this type of pervasive polysemy is not found in the other Pano languages whose ethnobiological taxonomic systems have been described; it is a distinctive characteristic of the Kakataibo language.
The distribution of primary terms throughout the Kakataibo data is also relevant since it does not necessarily observe Berlin's generalizations about ethnobiological nomenclature. While secondary names are systematically used for subgeneric taxa (with almost no exceptions), simple primary names and unproductive complex primary names distribute widely throughout the taxonomic system of Kakataibo and are relatively common for subgeneric taxa. The presence of simple primary names and unproductive complex primary names in association with subgeneric taxa is not predicted by Berlin, who states that, “A specifiable relationship can be observed between the names of taxa and their rank. Life-form and generic taxa are labeled by primary names; subgeneric taxa are labeled, in general, with secondary names” (Berlin 1992:34, nomenclature principle 3). It is true that Berlin's (1992:34, nomenclature principle 4) generalizations predict that under some conditions (polysemy and great cultural importance) subgeneric taxa might be named by primary names; however, not all the contradictory cases in Kakataibo satisfy these two conditions (cf. ñutan ‘type of tinamou’ [Crypturellus cinereus] and kaisa ‘fish type’ [Astyanax sp.], which are non-polysemous terms that equally denote specific taxa and are not neccesarily of great cultural importance). In fact, from the Kakataibo perspective, there is perhaps no more culturally important animal than the tapir but there are no non-polysemous primary names to refer to the different subtypes of tapir identified by the Kakataibo. Therefore, while the association between secondary names and subgeneric taxa finds solid support in the Kakataibo data, these data suggest at the same time that the alleged association between primary names and higher ranks might require further evaluation.
Footnotes
Acknowledgments
The present paper is a product of a one-year documentation project on the ethno-biological knowledge of Kakataibo people (Proyecto n° 70242.2024, “Etno-biología de los Kakataibo: una Aproximación al Saber Sobre la Naturaleza de un Pueblo Amazónico Peruano”), conducted in 2012 and funded by the Pontificia Universidad Católica del Perú, through its Dirección de Gestión de la Investigación. At this institution, I would like to thank Pepi Patron and Carlos Chávez and his team for all their support and help. I would also like to thank the Kakataibo speakers—Emilio Estrella, Ricardo Odicio, Salomón Estrella, Magaly Estrella, Karen Estrella, Irma Odicio, Ricardo Pereira and Alfredo Estrella—who helped with the elaboration of the lexical database used for this paper. A special thanks to David Fleck, who, as my co-researcher, has shared with me all his expertise in biology, without which this paper would have not been possible. I also thank William Aranda, who worked as a research assistant in the first stage of our project, Gabriel Martínez for proof-reading a previous version of this paper, and Hugo Ponce de León for creating the excellent map figure. Finally, I thank the editors of the Journal of Ethnobiology and the three anonymous reviewers for all their help and useful comments, which have enormously improved this paper.